Курсовая работа
Генотипическое и фенотипическое разнообразие населения Украины по группам крови
ВВЕДЕНИЕ
Группы крови формируются в результате действия различных эволюционных факторов. Актуальность темы данной работы состоит в том, что группа крови может определять устойчивость к определённым заболеваниям, распределение групп крови территориально может указывать на такие генетические процессы как дрейф генов, миграции.
Целью данной работы является исследование генетической структуры (полиморфизма) украинских популяций по локусам групп крови.
Задачи:
- установить фенотипическую, генотипическую структуру популяций разных областей Украины по группам крови по системе АВ0 и по системе Rh;
- определить частоты аллелей групп крови в популяциях разных областей Украины по группам крови по системе АВ0 и по системе Rh;
- сравнить и проанализировать полиморфизм на территории Украины.
1. ОБЗОР ЛИТЕРАТУРЫ
1.1 История происхождения территориально-административного деления Украины
Долгие века нынешняя территория Украины была разделены и находилась под властью различных государств. Рассмотрим историю некоторых нынешних городов Украины.
В начале нашего века территория Житомирской области
заселялась древними восточнославянскими племенами, север – древлянами, центральная часть и юг – полянами. В 945 году древлянские земли окончательно вошли в состав владений киевских князей – Киевской Руси. В X- начале XII веках район проживания древлян и полян входил в состав мощного древнерусского восточнославянского государства – Киевской Руси, затем в состав Владимиро-Волынского, а с 1199года – Галицко-Волынского княжества. Волынь и Полесье принадлежали к славянской прародине, были колыбелью украинского народа. В первой половине XIII века вместе с другими славянскими землями Житомирщина попала под гнет татаро-монгольских завоевателей. В 1362 году этой территорией завладели литовские феодалы, а после объединения Литвы и Польши в 1569 году в единое государство Речь Посполитую, волынские земли попали под власть польских магнатов. После воссоединения правобережной Украины с Российской империей в 1793 году, территория Житомирщины вошла в состав Волынской и частично Киевской губерний.
Край, где находится современная Черновицкая область
, заселялся в эпоху палеолита, о чем свидетельствуют обнаруженные здесь стоянки первобытных людей, возраст которых от 40 до 15 тыс. лет. Северная часть современной области в X-XII веках входила в состав Киевской Руси, позже была частью Галицко-Волынского княжества, которое со временем попало под власть Золотой Орды. В 1345 году венгерские войска выгнали татар с правобережья Днестра и Северная Буковина отошла Венгерскому королевству. В 1359 году Молдавское воеводство, входившее в состав Венгерского королевства, объявило о своей независимости. Образовавшееся Молдавское княжество распространило свою власть на всю территории Буковины. В 1514 году Буковина перешла под власть Турции. После русско-турецкой войны (1768-74гг.) Буковина перешла к Австрии, в составе которой и находилась до 1918 года. Этот период стал временем интенсивного развития региона, в 1849 году было образовано герцогство Буковина в составе Австрии. После распада Австро-Венгерской империи, была попытка присоединения ее к Украине, но армия Румынии оккупировала территорию Буковины. В состав современной области также входит часть исторической области Бессарабии (Хотинский район). Бессарабия – древнерусская земля, входящая в состав Галицкого, а позже Галицко-Волынского княжества, в XIV – XV вв. находилась в составе Молдавского княжества, а в XVI веке южные районы Бессарабии стали турецкими. В 1812 году по Бухарестскому мирному договору, заключенному между Российской империей и Турцией, территория Бессарабии отошла к России, на ее территории была образована Бессарабская губерния, одним из уездов который был Хотинский. В 1918 году Бессарабия была оккупирована Румынией. Территории северо-восточной части Королевства Румынии - Бессарабия и Северная Буковина были аннексированы Советским Союзом и 7 августа 1940 года, после присоединения Северной Буковины и Хотинского уезда Бессарабии к Украинской ССР, была образована Черновицкая область.
Закарпатье
, отделенное от остальной Украины естественной преградой – Карпатскими горами с несколькими перевалами - столетиями формировало свою особую историю и культуру, которые выделяют самую западную область страны от других украинских регионов. Первые упоминания о жизни людей на территории области относятся к периоду раннего палеолита. Прошлое Закарпатья неразрывно связано с восточными славянами. Отдельные славянские группы здесь появились в первое тысячелетие нашей эры и были выходцами из Прикарпатья и Волыни. В 896 году начался переход венгерских кочевых племен через Карпаты в Среднее Придунайье, освоение которого длилось на протяжении всего Х века. В Закарпатье, которое далеко находилось от основного центра славянства, еще некоторое время сохранялась власть славянской знати. Однако, начиная с XI века и завершая ХIII веком, Закарпатье постепенно вошло в новообразованное европейское государство – королевство Великая Венгрия и здесь окончательно утвердились венгерские короли. До средины ХХ века Закарпатье входило в состав разных государств, но всегда сохраняло свою славянскую сущность. Некоторое время часть Закарпатья входила в состав Галицко-Волынского княжества. Долгое время вся территория Закарпатья принадлежала венгерским правителям, но в 1541 году центральной Венгрией овладели турки, после чего она была разделена, Закарпатье оказалось поделенным на две части. Впоследствии центральные и восточные районы края вошли в Турецкую империю, а западная часть попала под власть Габсбургов. После 160-летнего турецкого ига, в конце XVII века вся территории Венгрии, в том числе и Закарпатье, перешла под власть Габсбургов. После поражения Австрии в австро-прусской войне 1866 года, было создано двуединое государство - Австро-Венгрия, после распада которого, в 1918 году образовался ряд государств, в том числе и Чехословакия. В 1918-19-х годах западная часть территории края была оккупирована чехословацкой армией, а румынские войска заняли юго-восточную часть. В то же время в Венгрии провозглашается Советская республика, включившая в себя большую часть Закарпатья.
Возле Запорожья
исследовано несколько поселений периода позднего палеолита. В VII веке до н.э. в Северном Причерноморье проживали скифы. На протяжении многих столетий территория края была своеобразным коридором, которым проходили разные народы, где скрещивались разные культуры. После татаро-монгольского нашествия в 1237-1240 годах, территория Запорожья на два столетия вошла в состав Золотой Орды. После порабощения Украины литовцами, а затем поляками, на запорожских землях в XV веке появились казаки («вольный человек» в переводе с тюркского). Южно-украинские земли стали одним из центров формирования запорожского казачества, которые осваивали земли у порогов Днепра.
Бердянск
– город областного значения в Запорожской области Украины, административный центр Бердянского района. На территории современного Бердянска жили люди еще в глубокой древности, о чем свидетельствуют обнаруженные в урочищах Сладкий Лиман и Ближний Шпиль остатки двух поселений эпохи неолита и ранней бронзы (III тыс. до н.э.). После ликвидации Запорожской Сечи и раздачи казацких земель помещикам многие казаки (около 5-7 тыс.), бежали во владения турецкого султана, который был заинтересован в том, чтобы запорожцы перешли к нему на службу, и разрешил им селиться на пограничной территории под Очаковым. Когда Екатерина II потребовала выдачи беглецов, турки на это не пошли и переселили казаков подальше от границы, на правый берег Дуная и в 1778 году султан официально признал запорожских казаков своими подданными. На турецких землях казаки основали Задунайскую Сечь, которая несколько раз меняла свое местонахождение. По словам украинского историка Михаила Грушевского, жилось казакам в Турции неплохо, только «мучила совесть запорожцев, что приходится помогать бусурманам воевать против христиан», поэтому с Задунайской Сечи убегали отдельные отряды запорожцев к своим единоверцам в Россию. В ответ на массовый уход казаков с Украины в Турцию, русское правительство арестовало бывших войсковых старшин и кошевого атамана Петра Кальнишевского, который более 25 лет провел в заключении в Соловецком монастыре. Однако, когда в 1787 году началась очередная война России с Турцией, российское правительство обратилось за помощью к бывшим казакам, которые оставались на территории Украины, из которых было создано «войско верных казаков», переименованное в Черноморское казачье войско в 1788 году. В русско-турецкой войне 1787-1791 годов черноморским казакам пришлось сражаться с казаками из Задунайской Сечи, т.е. своими бывшими соотечественниками.
Совсем другая ситуация сложилась в начале русско-турецкой войны 1828-1829 года, когда часть «турецких» казаков под Измаилом перешла на сторону русской армии. Из-за этого турки уничтожили Задунайскую Сечь. Из тех казаков, кто решил остаться на территории России, было сформовано Азовское казачье войско, которое поселили между Мариуполем и Бердянском.
История Сумской области
неразрывно связана с развитием Украинского государства - территория современной области заселялась в конце позднего палеолита. В VIII веке до н.э.- вошла в состав большого государственного формирования племен скифов – Скифии. В начале нашего века на территории Сумщины появились племена древних славян. В Х веке территория Сумщины входила в состав Киевской Руси, в Липовицкое княжество. В XI – XIII веках Сумщина входила в состав Черниговского и Северского княжеств. В 1360-е годы юго-западная часть нынешней области вошла в состав Великого Княжества Литовского, а затем - стала частью Речи Посполитой. После того, как императрица Екатерина II в 1764 году отменила гетманскую власть на Левобережье, а в 1765 году отменила полковой уклад и самоуправление в Слободской Украине, на Сумщину было распространено административно-территориальное деление Российской империи.
Территория нынешней Черкасской области
была заселена в древнейшем периоде каменного века – палеолите. Территория современной области входила в состав тех земель Приднепровья, где формировалось ядро восточнославянских племен, из которых возникла могучая феодальная держава – Киевская Русь. В 1239-40-х годах эти земли были опустошены татарами. Воспользовавшись татаро-монгольским нашествие и феодальной раздробленностью Руси, в XIV – XVI веках территорию современной области захватили литовские и польские феодалы.
2. ПОЛИМОРФИЗМ
Согласно определению, полиморфизм — это существование в популяции двух или более резко различающихся (прерывистых) форм, при котором частота более редкой формы определяется не одним лишь мутированием. Иными словами, полиморфизм — это такая изменчивость в локальной воспроизводящейся популяции, при которой проявляется чётко выраженное или резкое менделевское расщепление.
Такое определение полиморфизма не позволяет относить к нему некоторые типы изменчивости. Оно исключает чисто фенотипическую изменчивость (поскольку это негенетическая изменчивость); оно исключает географическую изменчивость (которой не существует в одной популяции); оно исключает полигенную изменчивость (при которой не происходит расщепления на резко различающиеся классы); и, наконец, оно исключает генетическую изменчивость, обусловленную новыми или повторными мутациями.
На основе разных критериев можно выделять различные типы полиморфизма. Важно различать генетический полиморфизм и хромосомную изменчивость. Генетический полиморфизм — это прерывистая изменчивость по гомологичным аллелям одного и того же генного локуса. Хромосомным называют полиморфизм по типам хромосом, например по половым хромосомам, или по таким перестройкам, как инверсии.
Различают также переходный и сбалансированный полиморфизм. В случае переходного полиморфизма разнообразие носит временный характер: оно наблюдается до тех пор, пока происходит процесс замещения одной формы другой при контролирующем действии естественного отбора. При сбалансированном полиморфизме разные типы представляют собой более или менее постоянные компоненты данной популяции благодаря естественному отбору, который благоприятствует сохранению разнообразия.
Все формы полиморфизма — генетический, хромосомный, переходный и сбалансированный — весьма обычны и широко распространены в живой природе. В популяциях организмов с половым размножением полиморфизм в сущности наблюдается всегда.
Явление полиморфизма подводит нас к концепции генофонда, которая в свою очередь даёт возможность по-иному взглянуть на локальную воспроизводящуюся популяцию. Рассмотрим популяцию, полиморфную по гену A и содержащую аллели A1, A2 и A3. В такой популяции будут возникать диплоидные генотипы A1A1, A1A2, A2A2 и т. п., и обусловленные этими генотипами формы будут представлены в любой выборке особей, однако ясно, что одна из главных черт данной популяции — это генетический полиморфизм, лежащий в основе наблюдаемого разнообразия. Можно сказать, что генофонд такой популяции содержит аллели A1, A2 и A3. Кроме того, эти аллели встречаются в генофонде с определённой частотой; допустим, что их частоты равны соответственно 60, 30 и 10%. Следовательно, популяцию можно описать количественно, используя типы генов, содержащихся в её генофонде, и их частоты.
Следует отметить, что концепция генофонда шире, чем концепция полиморфизма. Генофонд популяции слагается из всех имеющихся в ней генов. Так, генофонд нашей гипотетической популяции может быть полиморфным по гену А, содержать редкий мутантный аллель гена В и быть мономорфным по генам С и D.
Особи, из которых состоит данная популяция, в каждом данном поколении представляют собой различные генотипические продукты гамет, взятых из генофонда этой популяции в предшествующем поколении.
2.1 Полиморфизм по группам крови у человека по системе АВ0
Различия в групповой принадлежности крови зависят от различий в строении определенных сложных углеводных компонентов мембраны эритроцитов; это относится, по крайней мере к группам ABO и Lewis, которые наиболее изучены с химической точки зрения.
Групповые вещества крови представляют собой антигены,
т. е. при введении индивидууму, лишенному именно этого антигена, вызывают образование специфических антител. Многие из этих антигенов представляют собой белки, однако большинство близки к углеводным антигенам, описанным у бактерий. Антитела — это белки (иммуноглобулины), которые выделяются в плазму крови особыми антителообразующими клетками. Группоспецифические антитела, используемые для выявления антигенов групп крови, обычно обнаруживаются в сыворотке беременных женщин (иммунизированных антигенами собственного плода) или в сыворотке индивидуумов, которым многократно переливали кровь. Иногда такие антитела возникают «спонтанно» у индивидуумов с особыми группами крови; некоторые же (например, анти-М и анти-N) получают путем иммунизации кроликов эритроцитами человека.
Главный метод определения различий по группам крови — агглютинация.
Если отмытые эритроциты смешать с сывороткой, содержащей специфические антитела, то они покрываются белком антител и агглютинируют. Однако некоторые специфические антитела, связываясь с эритроцитами, не вызывают их агглютинации; такие антитела называют неполными.
Наследование различных вариантов групп крови удивительно точно следует законам Менделя. Этим можно воспользоваться при решении вопроса об отцовстве, для различения монозиготных близнецов от дизиготных, в работах по изучению сцепления генов. Многие различия по группам крови встречаются в разных популяциях с высокой и неодинаковой частотой; поэтому они представляют ценность и для антропологии. Более того, многие из них могут быть определены сразу после рождения и впоследствии изменяются лишь при нескольких редких заболеваниях. Вначале казалось, что правило «один ген — один антиген» приложимо и в этих случаях; однако с развитием молекулярной генетики и после описания ряда необъяснимых явлений стала очевидной ограниченность этого правила. Известно, что структурные гены представляют собой участки ДНК, в которых закодированы аминокислотные непоследовательности полипептидных цепей белковых молекул. Поэтому углеводные антигены, выявляемые при тестировании групп крови, не могут быть прямыми продуктами генов. Соответствующие гены, как полагают, контролируют ферменты (гликозилтрансферазы), участвующие в синтезе углеводных цепей. Далее, гены состоят из множества азотистых оснований, способных к мутациям. Возможно, что мутация, затрагивающая тот или иной фермент, приводит к изменению не одной антигенной специфичности групповых веществ крови, когда определенная группа антигенных специфичностей наследуется как целое. Ситуацию, когда определенная группа антигенных специфичностей наследуется как целое, можно теоретически объяснить, постулируя существование ряда тесно сцепленных генов; однако если сцепление столь тесно, что кроссинговер практически никогда не наблюдается, этот постулат остается лишь удобным предположением, не проясняя истинной структуры этих генов. Серологические методы чувствительны и высокоэффективны, но для понимания природы генов они дают нам меньше сведений, чем химия. К сожалению, почти ничего не известно о физиологическом значении групповых веществ крови, а ведь знание их функций позволило бы успешно вести поиски агентов, избирательно действующих на эти вещества.
В 1968 г. Рзйс и Сенджер в своей ставшей классической монографии перечислили 14 различных систем групп крови, т. е. группоспецифических веществ, которые контролируются независимыми генными локусами. Кроме того, существует немалое число антигенов групп крови, которые не варьируют (общие антигены), а также антигенов, найденных лишь в одной или нескольких семьях (личные антигены).
2.2 Система А1А2В0
Эта система была обнаружена первой; в 1900 г. Ландштейнер показал, что сыворотка некоторых индивидуумов вызывает агглютинацию отмытых эритроцитов ряда других индивидуумов. Это открытие позволило разработать метод безопасного переливания крови, а также предоставило в распоряжение генетиков и антропологов набор просто наследуемых вариантов, довольно часто встречающихся в большинстве популяций, легко обнаруживаемых и практически не зависящих от пола, возраста и окружающей среды. Затем лишь в 1924 г. Бернштейн установил тип наследования групп крови системы ABO, существо которого представлено в табл. 1. Эритроциты могут быть носителями антигенов А, или В, или А и В, или же ни одного из них; таким образом, мы получаем четыре фенотипа: группы А, В, АВ и О; наследование определяется тремя аллелями— А, В
и О,
которые при расщеплении дают 6 генотипов — АА, АО, ВВ, ВО, АВ, 00.
Таблица 1
Генетическая и серологическая характеристики групп крови системы ABO

Присутствие антигена А можно определить, смешивая клетки с сывороткой, содержащей анти-А, после чего они агглютинируют; антиген В определяется сходным образом с помощью анти-В. Истинный анти-0 не найден; поэтому мы не можем серологически отличить гомозиготы А или В от гетерозигот А или В (АО и ВО). Отсюда также следует, что мы не можем определить частоты этих генов в популяции путем простого подсчета; для этого приходится применять косвенные методы, исходя из допущения, что в данной выборке для этих аллелей соблюдается равновесие Харди — Вейнберга.
Анти-А, как правило, присутствует в сыворотке индивидуумов, лишенных этого антигена, а анти-В — у тех, кто лишен антигена В. По-видимому, эти естественно встречающиеся антитела образуются в ответ на A-подобные и В-подобные антигены бактерий и других организмов, проникающие в пищеварительный тракт человека вскоре после рождения.
Группу А можно подразделить на группы А1 и А2. Эритроциты А2 слабо реагируют с обычным анти-А, однако у нас нет специфических анти-А2, чтобы идентифицировать этот вариант. Правда, в организме некоторых А2- и А2В-индивидуумов образуются специфические анти-А1,которые можно применить для субтипирования A-клеток. Тем не менее генотипы А1А2серологически неотличимы от А1А1 или А10 , aA2А2— от A20.Описаны различные другие количественные варианты группы А (например, A-intermediate), довольно часто встречающиеся у африканцев.
Правила переливания крови.
Во многих случаях возникает необходимой переливания крови одного человека (донора) – другому (реципиенту). При переливании крови необходимо учитывать агглютиногены донора и агглютинины реципиента. При переливании небольших количеств крови агглютинины донора значительно разводятся и теряют способность агглютинировать эритроциты реципиента. У лиц с I (0) группой крови эритроциты не содержат агглютиногенов (ни α, ни β), такую кровь можно переливать лицам с любой группой крови. Кровь II (А) группы можно переливать только лицам со II (А) или IV (АВ) группой крови (у которых нет агглютининов a). Кровь III (В) группы можно переливать только лицам с III (В) или IV (АВ) группой крови (у которых нет агглютининов b); кровь IV (АВ) группы можно переливать только лицам с IV (АВ) группой, у которых вообще нет агглютининов. Таким образом, лица с I (0) группой крови являются универсальными донорами, а лица с IV (АВ) группой крови – универсальными реципиентами.
Однако изложенные правила пригодны лишь при переливании небольших количеств крови. При необходимости переливания больших количеств крови используется только одногруппная кровь.
2.3 Система
Rhesus
(Rh)
Резус-фактор — это антиген (белок), который находится на поверхности красных кровяных телец (эритроцитов). Он обнаружен в 1919 г в крови обезьян, а позже — и у людей. Около 85 % европейцев (99 % индейцев и азиатов) имеют резус-фактор и соответственно являются резус-положительными. Остальные же 15 % (7 % у африканцев), у которых его нет, — резус-отрицательный.
Резус-система определяется тремя сцепленными генами (CDE); все эти гены локализованы в 1-й хромосоме. Наиболее сильным антигеном резус-системы является антиген RhD, который контролируется соответствующим геном D. При этом резус-положительная группа крови доминирует над резус-отрицательной.
Наследование резус-фактора происходит сложным образом, но, учитывая ведущую роль гена D, его можно представить как моногенное наследование с полным доминированием: при генотипе DD или Dd резус положительный (Rh+), а при генотипе dd – отрицательный (Rh–). Переливание резус–положительной крови человеку с резус-отрицательной группой крови приводит к развитию иммунной реакции на резус-антиген: синтезируются резус-антитела, то есть происходит иммунизация организма против резус-антигена. Этот процесс продолжается в течение 2…4 месяцев. Резус-антитела сохраняются в сыворотке длительное время. Если иммунизированному человеку перелить резус-положительную кровь, то резус-антитела разрушают резус-положительные эритроциты, что приводит к гемолитическому шоку, а нередко – и к смерти человека.
Резус-конфликт
. Большое значение резус-совместимость имеет при вынашивании плода. Если резус-отрицательная женщина (dd) вынашивает резус-положительный плод с генотипом Dd (аллель в приходит от резус-положительного отца DD или Dd), то происходит иммунизация организма матери резус-антигеном.
Существуют различные точки зрения на механизм иммунизации. Согласно одной из них, часть эритроцитов плода попадает в кровоток матери при родах. Однако имеются данные о возможности проникновения резус-антигенов через плаценту еще до родов; в то же время, в некоторых случаях иммунизации не происходит.
Если организм женщины оказался иммунизированным резус-антигеном, то при повторной беременности резус-антитела через плаценту попадают в кровоток плода. Если генотип второго ребенка dd, то резус-антигены отсутствуют, и резус-конфликта не происходит. Но если генотип второго ребенка Dd, то резус-антитела матери разрушают резус-положительные эритроциты плода. Это приводит или к гибели плода, или к рождению неполноценного ребенка.
Таким образом, браки ♀ Rh– × ♂ Rh+ (например, ♀ dd × ♂ Dd или ♀ dd × ♂ DD) оказываются неблагоприятными, т.к. в этом случае велика вероятность рождения неполноценного ребенка. Однако в каждом конкретном случае все может быть не так страшно.
Во-первых, в браках ♀ dd × ♂ Dd (то есть при гетерозиготности отца) вероятность образования резус-отрицательного плода с генотипом dd составляет 50%. Тогда никакой иммунизации не происходит. Вполне возможно, что и последующие дети окажутся резус-отрицательными, и никакого резус-конфликта не возникнет.
Во-вторых, даже если в браке ♀ dd × ♂ Dd первый плод окажется резус-положительный (Dd), и произойдет иммунизация материнского организма, то последующие дети могут оказаться резус-отрицательными, и резус-антигены не окажут на их эритроциты вредного воздействия.
В-третьих, при обнаружении в крови резус-отрицательной женщины резус-антигенов (например, сразу после рождения резус-положительного ребенка) ей переливают сыворотку крови от мужчин-добровольцев, искусственно иммунизированных резус-антигенами. Такая сыворотка содержит резус-антитела, которые разрушают эритроциты с резус-антигенами, появившиеся в женском организме. В результате иммунизации женского организма не происходит.
И, наконец, при рождении резус-положительного ребенка, в крови которого обнаруживаются резус-антитела, ему производят полное (заместительное) переливание крови на кровь, не содержащую резус-антитела.
2.4 Географическое распределение групп крови А1А2
B0
Прежде всего, мы должны понять, что гены O, A, и B являются очень древними. Они не являются результатом недавних мутаций. Например, анализ египетских мумий показывает, что три гена, отвечающих за образование групп крови, присутствуют там в таких же пропорциях, как и у современных египтян.
На сегодня наиболее обоснованной является теория, согласно которой современный человек сформировался в Центральной Азии в то время, когда климатические условия там были более благоприятными, чем сейчас. Эта самая ранняя группа современных людей могла иметь гены с повторяемостью 25 A, 15 B, и 60 O на каждую сотню A-B-O генов групп крови. Примерно такова повторяемость генов в Центральной Азии сегодня.
По мере приближения к историческим временам и роста человеческой популяции на Земле, отток людей из Центральной Азии увеличивался. Эмиграционные группы были достаточно велики, чтобы перенести с собой ген B. Наиболее приближенные к Центральной Азии районы, например, Маньчжурия и северная Индия, получили наибольшую порцию. Восточная Европа получила большее количество на этом континенте, и по мере продвижения на запад частота гена B исчезает. Северо-восточная Африка получила больше всего на своем континенте, и частота гена B уменьшается по мере продвижения на запад и юг.
Народы Азии и Африки в общем имеют высшую частоту гена B, чем другие люди. Они отличаются друг от друга в частоте Rh. Азиаты имеют выше частоту гена, называемого Rhz, чтобы отличить его от других генов ряда Rh. С другой стороны, африканцы имеют высокую повторяемость другого гена Rh, называемого Rho.
Американские, австралийские и азиатские группы имеют небольшое количество или вообще не имеют гена rh. Африканская группа имеет небольшое количество гена rh. Жители Европы (включая американцев и австралийцев, ведущих свое происхождение от европейцев), тем не менее, имеют значительное количество гена rh; примерно каждый седьмой из них является Rh-отрицательным.
Если мутации, в результате которых появились антигены A и B являются древними, то группа крови O гораздо старше.
Другой параметр, свидетельствующий о древности группы O, следует из анализа физической антропологии и предполагает, что большую часть своего существования человечество принадлежало исключительно к группе O.
Новые исследования по митохондриальной DNA (mtDNA) подтверждают теорию о появлении Homo sapiens в Африке и последующем распространении в других регионах.
Высокий коэффициент группы крови O позволяет предполагать, что американские индейцы и эскимосы напрямую произошли от кроманьонских предков, вероятно, монгольских, которые мигрировали примерно в 15,000 до н.э. в Америку.
Малая часть коренных индейцев-американцев имеют группу крови В, таким образом они должно быть мигрировали в Америку достаточно поздно для получения положительного гена Rh, но слишком рано, чтобы получить ген для B.
Группа крови A в наибольших концентрациях обнаруживается у западноевропейцев. В отличие от групп крови B и O, группа A имеет множество вариаций. Основная группировка, A1, встречается примерно у 95 процентов группы A. Самая большая подгруппа, A2, встречается в основном на Северном Кавказе. A2 в очень больших концентрациях обнаруживается в Исландии и Скандинавии, особенно у лаппов, древнейших обитателей этого района. Они почти уникальны в высокой частоте группы A, и имеют самую высокую частоту A2, 42 процента в одной группе населения. A2 ген почти полностью сконцентрирован у населения Кавказа.
Частота группы А в Европе снижается по мере продвижения на восток. На большей территории Европы повторяемость гена А превышает 25 процентов. Он так же в значительных количествах обнаружен на территориях вокруг всего Средиземного моря, особенно на Корсике, Сардинии, Испании, Турции и Балканах. Ясно, что человечество чаще основывало постоянные поселения в тех местах, где условия обещали им самые высокие шансы на выживание.
Видимо ген В мог попасть на Индийский субконтинент через завоевания. При исследовании распределения ABO вдоль Шелкового пути Северо-западного Китая наблюдалось явное увеличение частоты группы крови В, особенно когда сравнивались субъекты монголоидний выборки с субъектами европеидной выборки.
Почти непрерывная цепь гористой территории протянулась от Уральских гор до Кавказа в Азии, и далее до Пиренеев в южной Франции. Этот барьер разбил миграцию групп крови на два основных маршрута; северный поток и южный. Переселенцы, выбравшие южный путь, стали предками жителей Средиземноморья и западноевропейцев, они несли с собой ген группы А. Уральские горы остановили большую миграцию на запад из Азии, хотя небольшое количество европеидов попало в восточную Европу, принеся с собой ген группы крови В, который они получили, смешавшись с азиатскими монголоидами. Это барьер послужил для разделения групп крови на западную A; и восточную B группы.
Монголоиды, носители группы крови B; продолжали двигаться на север, в направлении современной Сибири.
Для современных антропологов группа крови В продолжает и на сегодняшний день оставаться ‘Восточной’ группой крови. В больших количествах она обнаруживается у азиатского населения, такого как китайцы, индийцы, сибиряки. В Европе группа крови В чаще всего встречается у венгров, русских, поляков и других восточно-европейских народов. Среди до-неолитических народов, таких как баски и американские индейцы, группа крови В практически не встречается.
Из всех групп крови ABO, B демонстрирует четко выраженное географическое распространение. Протянувшись, как огромный пояс, вдоль равнин Евразии до Индийского субконтинента, группа крови В в повышенных количествах насчитывается на территориях от Японии, Монолии, Китая и Индии до Уральских гор. Отсюда в западном направлении процентное соотношение уменьшается, пока не достигает самого низкого значения в самой западной точке Европы.
Группа крови B является явно не индоевропейской группой крови. В Европе, только две области выделяются высоким соотношением группы крови В: одна среди не индоевропейских народов, известных как финно-угры (венгры и финны), другая – среди центрально-славянских народов (чехи, южные поляки, и северные сербы). Вторгающиеся Викинги также могли иметь относительно высокий процент гена В, поскольку многие города Британии и Западной Европы, связанные с побережьем только внутренними коммуникациями такими, как большие реки, имеют, в сравнении с окружающими территориями, непропорциональное количество группы В.
Небольшие количества групп крови у западноевропейцев отражают миграцию азиатских кочевников на запад. Это наиболее явно прослеживается у восточных западноевропейцев, немцев и австрийцев, которые имеют неожиданно высокий процент группы В по сравнению со своими западными соседями. Высокая повторяемость группы В у немцев имеет место на территории около верхней и средней Эльбы, важной естественной границы между ‘цивилизацией’ и ‘варварством’ в древние и средние века.

Рис. 1. Частота гена 0 групп крови системы АВ0

Рис. 2. Частота гена А групп крови системы АВ0

Рис. 3. Частота гена В групп крови системы АВ0

Рис. 4. Распределение аллеля В у населения Европы

Рис. 5. Распределение групп крови по системе Rh у населения Европы
кровь группа полиморфизм аллель
3. ИМУНОХИМИЧЕСКИЕ ХАРАКТЕРИСТИКИ ГРУПП КРОВИ
Принадлежность индивидуума к той или иной группе крови определяется наличием или отсутствием в эритроцитах группоспецифических веществ – антигенов А и В. Вещества с такими свойствами и такой же специфичностью были найдены в большинстве тканей организма. Исключения составляют яички, хрусталик, хорион, плацента, хрящ и эпителий кожи. Группоспецифические вещества тканей относятся к так называемым спирторастворимым и представляют собой гликолипиды. Водорастворимые группоспецифические соединения в больших количествах встречаются в различных слизистых секретах (слюне, слизи желудочно-кишечного тракта и др.) и являются гликопротеинами. Важно подчеркнуть, что антигены А и В обнаруживаются в семенной жидкости, плазме, слюне и женском молоке, причем именно в жидкостях выявляется в растворенном состоянии наибольшее количество этих веществ. Таким образом, спирто- и водорастворимые субстанции относятся к различным классам макромолекул, обладающих одинаковыми антигенными свойствами, детерминированными одним и тем же локусом.
Химическая структура определена для водорастворимых группоспецифических веществ, которые состоят из большого числа относительно коротких олигосахаридных цепей, соединенных ковалентными связями с полипептидными основами. Молекулярная масса этих гликопротеидов – около 500 000 дальтон, их углеводные цепи состоят из 7…8 остатков сахара, и каждая молекула содержит около 300 углеводных цепей. Групповую специфичность антигенов А и В определяют терминальное положение сахаров в цепи молекулы, причем существенно, что молекула вещества А заканчивается остатком N-ацетилгалактозамина, а цепь молекулы антигена В остатком галактозы. В остальном обе цепи идентичны. При 0-группе крови в цепи отсутствуют терминальные остатки, т. е. N-ацетилгалактозамин или галактоза, и молекула приобретает так называемую Н-специфичность. Установлено, что у носителей аллеля IА имеется специфическая трансфераза, переносящая N-ацетилгалактозамин к концевой группе олигосахаридной цепи. Аналогичным образом у носителей аллеля IВ, обладающих D-галактозилтрансферазой, к концевой группе олигосахаридной цепи присоединяется галактоза. У индивидуумов с группой крови 0, т. с. гомозигот по аллелю I0 соответствующие трансферазы отсутствуют, и к углеводной цепи никакой дополнительный остаток не присоединяют. Без этого дополнительного остатка иммунологически определяется Н-антигенная специфичность. Н-антигены как продукты промежуточной реакции присутствуют у индивидуумов с А- и В-группами крови, но естественно, что концентрация Н-антигенов выше у индивидуумов с группой крови 0.
Обнаружен специфический локус, контролирующий синтез Н-антигена. Так, при генотипах НН или Hh образуется Н-вещество как конечный продукт реакции у индивидуумов с группой крови 0 и как предшественник А- или В-веществ у индивидуумов с группой крови А, В или АВ. При генотипе hh Н-антиген не образуется, и у всех гомозигот в этом случае серологически будет определяться группой крови 0, хотя они и могут быть носителями генов А и В. Такое явление лежит в основе так называемого «бомбейского феномена», при котором не синтезируется ни гликолипиды, ни гликопротеины А и В. Другой локус – Se (secretor) – контролирует образование Н-специфической группировки только в углеводных цепях гликопротеидов: при генотипах Se/Se и Se/se синтезируются Н-специфические гликопротеиды, а при генотипе se/se Н-гликопротеины не синтезируются, и соответственно в железистых секретах не образуются А- и В-антигены. Вместе с тем при генотипе se/se синтез Н-гликопротеинов происходит, и при наличии соответствующих трансфераз образуются А- и В-антигены как в эритроцитах, так и других клетках организма. Ген Le (Lewis) детерминирует синтез вещества, находящегося в конкурентных отношениях с Н-антигеном. Описаны и некоторые другие пути биосинтеза группоспецифических веществ. Показано, что их образование контролируется многими локусами, и в процессе их биосинтеза возможны разнообразные типы взаимодействия генов. (Явление подавления синтеза А- и В-специфичности имеет важное значение для объяснения биологической роли отсева несовместимого потомства.)В настоящее время установлена локализация локуса АВ0 – в длинном плече 9-й хромосомы. Однако доказательств сцепления между генами АВ0 и Н до сих пор не получено. В то же время известно, что гены Se и Le локализованы в 19-й хромосоме.
В сыворотке крови неиммунизированных к АВ0-антигенам людей содержатся так называемые нормальные агглютинины – антитела к группоспецифическим веществам системы АВ0. Они вырабатываются к отсутствующим у данных лиц веществам. Так, у индивидуумов с группой крови 0 имеются анти-А (α)-и анти-В (β) -антитела, у лиц с группами крови А и В – соответственно анти-В- и анти-А-антитела, при группе крови АВ подобные антитела не вырабатываются. Эти изоантитела (α и β) представляют собой 19S иммуноглобулины (IgM) с молекулярной массой около 900 000 дальтон.
В ответ на введение чужеродного антигена в организме образуются иммунные агглютинины. В отличие от нормальных, иммунные агглютинины представляют собой 7S-иммуноглобулины (IgG) и имеют молекулярную массу около 160 000 дальтон. Важным отличительным свойством этих агглютининов является их способность проникать через плаценту и большая агглютинационная способность. Происхождение нормальных агглютининов до сих пор не выяснено. Некоторые исследователи полагают, что изоагглютинины образуются в результате иммунизации антигенами, близкими по своей структуре к веществам А и В. Такие антигены довольно часто встречаются во многих продуктах питания и медикаментах. По мнению других авторов, самостоятельное образование изоагглютининов у плода генетически обусловлено. При сравнительном изучении АВ0-системы были получены весьма интересные данные. Оказалось, что АВ0-антигенная дифференцировка тканей встречается у многих видов приматов, в том числе и человекообразных обезьян. Антигены, идентичные или подобные антигенам АВ0-системы, широко распространены также во всем остальном животном и растительном мире. Например, у свиней, коров, баранов и многих других животных в эритроцитах содержатся агглютиногены А и В, идентичные или близкие человеческим. Многие препараты (из крови человека, иммунные сыворотки животных, прививочный материал, экстракты из органов животных и гормональные препараты), применяемые с лечебной, профилактической и диагностической целями, содержат вещества, аналогичные антигенам АВ0-системы. Вещества, сходные с антигенами АВ0-системы, выявлены у вирусов, риккетсий, бактерий, грибов и высших растений.
Такая структурная конвергенция соединений, образующихся у организмов, которые находятся на самых различных уровнях организации, поразительна. Не обладая свойствами ферментов, эти соединения представляются еще загадкой в молекулярно-функциональном аспекте, если не учитывать, конечно, того обстоятельства, что их идентифицируют в качестве структурного компонента мембран в большинстве клеток человека и других организмов. Необходимо также отметить, что АВ0-система является одной из основных систем совместимости у человека, а ее гены обладают выраженным плейотропным эффектом в отношении предрасположенности к некоторым заболеваниям. Следует подчеркнуть, что АВ0-локус, по-видимому, репрессируется лишь на сравнительно короткий период эмбриогенеза или вообще не репрессируется.
3.1 Закономерности гибели зародышей при АВ0-несовместимости матери и плода
Интенсивность отсева при несовместимости можно рассчитать достаточно точно, принимая, что все достоверные отклонения в рождении детей от теоретически ожидаемых (или контрольных) показателей являются результатом внутриутробной гибели зигот. При некоторых типах браков элиминируется 80 % и более несовместимого потомства, однако также как и для характеристики элиминации зигот с хромосомными аномалиями, интенсивность отсева при АВ0-несовместимости должна рассчитываться на всю популяцию. Расчет интенсивности отсева по парам мать–ребенок дает несколько заниженные оценки, поскольку при этом учитывается только элиминация несовместимого потомства. Так, по данным Головачева (1983), в группе, состоявшей из 2827 пар мать–ребенок, интенсивность элиминации составляла 9,93 %, что в пересчете на всю выборку из 9813 пар индивидуумов равно 2,8 %.
При расчете интенсивности отсева по расщеплению потомства в семьях получают более правильные оценки, так как такой расчет позволяет учитывать отсев, направленный против и несовместимого и совместимого потомства. Так, интенсивность отсева при АВ0-несовместимых браках составляет 3,98 % для выборки, состоящей из 2250 семей, и 3,28 % для популяции г. Ленинграда с учетом резус-отрицательных браков. Согласно некоторым данным, репродуктивные потери при АВ0-несовместимости составляют не менее 3…5%. При этом необходимо иметь в виду, что суммарная оценка отсева зависит от демографических характеристик соотношения числа перво- и повторнобеременных женщин и возрастного состава популяции. Если эти факторы не учитывать, то оценка будет заниженной из-за разнонаправленности векторов отбора.
Репродуктивные потери при АВ0-несовместимости относительно высоки, а их биологический смысл неясен. Последнее особенно бросается в глаза при сравнении с отсевом хромосомных аномалий, при котором биологическое значение гибели носителей аберрации очевидно. Поэтому проблема отсева при АВ0-несовместимости требует самого подробного обсуждения, главным образом в аспекте общих закономерностей отбора и взаимодействия между организмами матери и плода. Высокую интенсивность отсева по АВ0-системе можно объяснить следующим:
1) существующее сейчас распределение групп крови в различных странах возникло недавно, вероятно, в результате смешения рас;
2) отсев при несовместимости является «расплатой» за адаптивность постнатального полиморфизма;
3) биосинтез группоспецифических веществ является одним из обязательных биохимических путей нормального развития, а эффекты действия АВ0-генов (гетерозис, плейотропия и др.) изменяют жизнеспособность эмбрионов;
4) изменения в окружающей среде, воздействующие на современного человека, неблагоприятно отразились на физиологических последствиях АВ0-несовместимости.
Гипотеза 1
Всесторонне проверить первое объяснение в приложении к Rh- и АВ0-системам в настоящее время не представляется возможным. Генетики рассчитали изменение частоты генов рА, qB и r0 в Европе, учитывая факторы миграций, внутриутробного отбора и интегрального гетерозиса. Оказалось, что равновесие частот генов может наступить примерно через 20 поколений, однако, при таком равновесии внутриутробный отсев будет происходить почти с такой же интенсивностью. Таким образом, эти данные не противоречат первой гипотезе, но и прямо не отвечают на поставленный вопрос, так как констатируют лишь факт нестабильности частоты генов среди населения Европы.
В большей степени первую гипотезу подтверждает хорошо известный факт повышения частоты гена r0 (также как иногда и гена cde системы Rh) в некоторых горных изоляторах Европы. Отсутствие гена qB, а иногда даже и рА, у коренного населения Америки также согласуется с этим предположением, хотя в данном случае не исключен дрейф генов.
Гипотеза 2
Обсуждение второй гипотезы целесообразно начать с вопроса о факторах, поддерживающих полиморфизм групп крови. Популяционно-генетические расчеты свидетельствуют о том, что для повышения до существующего уровня частоты мутантных генов системы АВ0 требовалось мощное давление отбора, однако отбор в пользу мутантного гена должен «преодолеть барьер несовместимости между матерью и плодом». Иными словами, интегральный вектор отбора в пользу гетерозигот должен быть больше вектора, направленного в пользу гомозигот, поскольку часть гетерозигот элиминируется при несовместимых по системе АВ0 беременностях. В связи с этим проблема внутриутробной гибели плода при АВ0-несовместимости приобретает принципиальное значение, особенно, если ее анализировать в связи с другими факторами, влияющими на изменение частот генов АВ0-системы в популяции. По данным литературы, представляется ясным только то, что, с одной стороны, имеет место отсев несовместимых с матерью гетерозиготных плодов, а с другой стороны, частоты генов в популяции в течение длительного периода практически не меняются. Следовательно, можно предположить, что существуют какие-то механизмы, обеспечивающие эту стабильность.
По мнению некоторых исследователей, гетерозиготное потомство в браках типа ♀А×♂0; ♀В×♂0 и т. д. (т. е. реципрокных несовместимым), более жизнеспособно, чем гомозиготное, и, следовательно, гетерозиготы могут поддерживать частоту генов в популяции на определенном уровне. Другие исследования показали, что в АВ0-локусе при отборе проявляется эффект гетерозиса. Анализ распределения частот генов групп крови в нескольких европейских и американских популяциях приводит к заключению об адаптивном преимуществе гетерозигот по локусу АВ0. Отечественные генетики показали наличие гетерозиса по АВ0-локусу для популяций коренного населения Сибири. Следует подчеркнуть, что во всех этих исследованиях определялись интегральные векторы отбора, и предполагалось, что гетерозиготы имеют адаптивное преимущество в постнатальный период.
На молекулярном уровне гетерозис по АВ0-локусу объясняется возможностью образования гибридного гетеродимера гликозил-трансферазы у A1B-гетерозигот. Из этого следует, что жизнеспособность особей с генотипом АВ всегда должна быть наивысшей. Такое преимущество является необходимым условием, поддерживающим равновесие генных частот. Однако, по данным некоторых авторов, относительная адаптивная ценность индивидуумов с генотипом АВ ниже, чем индивидуумов со всеми другими генотипами. Кроме того, у женщин с группой крови АВ спонтанные выкидыши наблюдаются достоверно чаще, чем у женщин с группой крови 0.
Однако известно, что у гетерозиготных родителей, например, АВ×АВ или MN×MN, или Нр2-1×Нр2-1, может наблюдаться избыток гетерозиготных детей. Стабильность генных частот в популяции может обеспечиваться также посредством различий в фертильности у индивидуумов с различными группами крови.
Значительное место в процессах, поддерживающих полиморфизм, могут занимать корреляции между группами крови и различными заболеваниями. Однако некоторые исследователи считают, что вопрос о взаимосвязи групп крови с болезнями неинфекционной природы и отбором остается нерешенным. Большинство изученных заболеваний (такие, как рак и язва желудка, инфаркт миокарда) поражают людей уже в том возрасте, когда репродукция прекращается или снижена, и поэтому они незначительно влияют на относительное число аллелей в последующих поколениях. Имеющиеся в настоящее время данные о связи между группами крови и неинфекционной патологией, по-видимому, подтверждают лишь то, что гены групп крови потенциально подвержены действию естественного отбора. Существование таких связей свидетельствует в пользу этиологической роли генетических факторов, однако отсутствие таких связей вовсе не исключает роли генетических факторов.
Ассоциации групп крови А и 0 с предрасположенностью к оспе или чуме, несомненно, могли быть важными факторами полиморфизма человеческих популяций в прошлом. Однако в настоящее время они, очевидно, не могут иметь такого значения. Весьма вероятно также, что в некоторых регионах действовали частные факторы, например географические и климатические. Возможно, меньшая предрасположенность индивидуумов с группой крови 0 к ревматическим заболеваниям могла способствовать распространению гена 0 среди древних обитателей атлантического побережья Европы – кельтов.
Нельзя также игнорировать более общие механизмы, поддерживающие полиморфизм в человеческих популяциях. В основе таких общих механизмов могут лежать процессы адаптации микроорганизма к обмену веществ хозяина. Очевидно, что в популяциях с достаточным числом полиморфных систем процессы адаптации к инфекции занимают больше времени, и в связи с этим же индивидуумы имеют больше, возможностей эффективно мобилизовать специфические иммунные реакции. Эти общие соображения ни в коем случае не умаляют значения поисков конкретных механизмов, поддерживающих полиморфизм генов системы АВ0.
Таким образом, имеющиеся в настоящее время данные до конца не вскрывают механизмы компенсации АВ0-отсева. Эти данные также не проливают света на биологическую роль репродуктивных потерь в АВ0-несовместимых браках. Некоторые исследователи отмечают, что «для АВ0- и Rh-несовместимости положение еще хуже, поскольку в обеих системах отбор направлен против гетерозигот, и, таким образом, они совершенно неустойчивы». Однако эта крайняя точка зрения не разделяется многими исследователями и противоречит нашим данным о существовании внутриутробного отбора в пользу гетерозигот.
Гипотеза 3
Напомним, что АВ0-локус, по-видимому, репрессируется лишь на сравнительно короткий период эмбриогенеза или вообще не репрессируется. Таким образом, необходимо учесть, что принципиально возможно устранение иммунных взаимодействий между матерью и плодом за счет репрессии АВ0-локуса в эмбриогенезе. Данные по биохимии и генетике АВ0-системы свидетельствуют о существовании нескольких путей подавления биосинтеза А- и В- группоспецифических веществ, причем описано несколько рецессивных локусов, контролирующих на различных этапах подавления биосинтеза антигенов А, В или Н(0). Однако ни один из таких рецессивных аллелей не получил достаточного распространения ни в одной из обследованных популяций. Кроме того, АВ0-локус практически функционирует в течение всего онтогенеза. Следовательно, потенциальные возможности регуляции локуса системы АВ0 в раннем онтогенезе не реализуются, и поэтому можно думать об определенном участии АВ0-антигенов в нормальном эмбриогенезе человека.
Однако, поскольку при 10…35 % беременностей выявлены несовместимость матери и плода по системе АВ0, то должен существовать эффективный барьер, препятствующий иммунному конфликту. Действительно, в настоящее время доказано, что между матерью и плодом имеется морфофункциональный барьер, в известной степени предотвращающий иммунные конфликты. Этот барьер создается за счет функционирования различных звеньев — от структурных до динамических, – возникающих при взаимодействии иммунных систем, например Rh и АВ0. Но такой барьер полностью не препятствует возникновению иммунологических реакций со стороны материнского организма ни при нормально протекающей беременности, ни при ее осложнениях. Итак, даже при неполностью эффективном барьере не происходит отбор генов, подавляющих биосинтез группоспецифических веществ в раннем онтогенезе. Это еще раз свидетельствует о самостоятельной роли группоспецифических веществ во внутриутробном развитии и, следовательно, противоречит предположению о пассивной связи этих соединений с постнатальным полиморфизмом.
Исследователи, изучавшие антигены у эмбрионов и новорожденных, установили, что эти антигены присутствуют на ранних эмбриональных стадиях, затем исчезают и могут быть вновь идентифицированы при наступлении эритропоэза. Таким образом, АВ0-антигены имеют определенное значение в развитии клеточного метаболизма в течение одного или более критических периодов развития эмбриона и новорожденного. Некоторые авторы полагают, что вредное воздействие АВ0-несовместимости проявляется в течение ограниченного числа критических периодов (гаметический, эмбриональный и ранний фетальный). Принципиально трудно возражать против гаметической селекции в АВ0-несовместимых браках, однако нельзя и переоценивать этого типа отбора. Так, преобладание АВ0-несовместимого потомства служит еще одним аргументом против мнения о заметном вкладе гаметической селекции в отсев.
Вероятно, гибель зародышей совпадает с критическими периодами, приходящимися на имплантацию бластоцисты (на 4…6-й неделе развития) и период, предшествующий особенно быстрому росту зачатка плаценты. Начиная с 4…6-й недели беременности и кончая, примерно, 15…17-й неделей, действие АВ0-несовместимости на плод проявляется в учащении самопроизвольных выкидышей. Когда плацента формируется и начинает функционировать как орган, т. е. примерно с 14…15-й недели развития, гибель эмбрионов и последующие выкидыши, ассоциирующиеся с АВ0-несовместимостью, не наблюдаются вплоть до перинатального периода.
Эффект несовместимости в перинатальный период связан, вероятно, с повышением проницаемости плаценты для антител, что наблюдается незадолго до родов. Физиологический характер подобного явления не вызывает сомнений, поскольку при этом обеспечивается пассивная иммунизация плода материнскими антителами. Но вместе с антителами, направленными против инфекций и неспецифических гуморальных факторов иммунитета, в организм плода также проникают анти-А- и анти-В-антитела, которые при несовместимости вызывают патологические процессы у новорожденных, в частности эритробластоз. Летальные формы АВ0-несовместимости крайне редки, хотя легкие формы несовместимости могут быть выявлены у 7,1% новорожденных. Как подчеркивает Ж. Доссе (1959), незначительные последствия несовместимости по системе АВ0 наблюдаются в 2—3 раза чаще, чем по другим системам. АВ0-несовместимость также является частой причиной изменения функционального состояния новорожденных. Редкость тяжелых форм осложнений при АВ0-несовместимости объясняется существованием эффективных защитных механизмов, направленных против иммунологического конфликта в перинатальном периоде. Одним из таких механизмов является низкая антигенная активность А- и В-субстанций в эритроцитах плода и новорожденного, т. е. частичная репрессия локусов А и В, которая но каким-то причинам не происходит на ранних этапах эмбриогенеза. Таким образом, отсев потомства при АВ0-несовместимых браках различается по механизмам в зависимости от периодов внутриутробного развития.
Данные о преобладании несовместимого потомства вносят новый аспект в проблему внутриутробного отсева. Благодаря преобладанию то несовместимого, то совместимого потомства направления отбора еще в период утробного развития могут быть сбалансированы. Естественно, что динамическая компенсация имеет преимуществ тем больше, чем на более ранних стадиях развития она происходит. Поэтому пренатальное поддержание полиморфизма генов групп крови должно иметь большое биологическое значение. По аналогии с постнатальным периодом не исключено, что в основе преобладания несовместимого потомства у молодых первобеременных женщин лежит гетерозис по АВ0-локусу. При последующих беременностях происходит иммунизация материнского организма, и логично предположить, что «повреждение» зародыша материнскими анти-А- и анти-В-антителами может превысить эффект гетерозиготности. Тогда проявляется классический феномен внутриутробного отсева несовместимого потомства. Таким образом, учитывая смену направлений отсева, поставленный выше вопрос об имманентности утробного отбора по отношению к полиморфизму генов групп крови в известной мере теряет свой смысл. Данные по биохимической генетике системы АВ0 также говорят не в пользу этой гипотезы. Следовательно, сравнительно высокая эмбриональная смертность при АВ0-несовместимых браках не может рассматриваться только как дань адаптивности постнатальному полиморфизму.
С позиций эволюционной генетики также представляется маловероятным, что полиморфизм групп крови пассивно связан с нормальным эмбриогенезом. Система АВ0 или близкие к ней системы найдены у многих приматов, в том числе и среди человекообразных обезьян, что свидетельствует о вероятном существовании подобной системы и среди прямых предков человека. В этом аспекте представляется неправдоподобным существование пренатального иммунологического груза «в чистом виде» в филогенезе рода Homo на протяжении нескольких миллионов лет. Следовательно, логично предположить, что иммунологические механизмы, связанные с функционированием АВ0-локуса вовлекаются в процессы нормального эмбриогенеза. В связи с этим интересна гипотеза, согласно которой несовместимость между матерью и плодом является необходимым условием развития нормальной беременности. По аналогии с некоторыми явлениями трансплантационного иммунитета автор предположил, что пролиферативные процессы при образовании плаценты обусловлены несовместимостью между матерью и зародышем, и антигенное сходство при прочих равных условиях ведет к снижению массы плаценты и плода. При клинических исследованиях, посвященных изучению влияния АВ0-несовместимости на массу плаценты не получены однозначные результаты. Однако рассматриваемая гипотеза нашла подтверждение в экспериментальных работах, в которых показано, что при антигенной несовместимости имеется достоверное физиологическое увеличение массы плаценты. Также оказалось, что при иммунизации самки антигенами самца массы плаценты и плода увеличиваются, при этом также повышается вероятность успешных имплантаций.
Отечественные исследователи, проводя межлинейную пересадку бластоцист у мышей, выявили, что увеличение массы и жизнеспособности гибридных эмбрионов обусловлено не гетерозиготностью как таковой, а их большой антигенной несхожестью с организмом самки. Однако снижение массы плаценты и плода при приобретении организмом самки толерантности к антигенам самца может быть связано не только с отсутствием иммунологического распознавания, но и с явлением трансплантационной депрессии. Последним можно объяснить противоречивость клинических данных о влиянии несовместимости на массу плаценты.
Гибель совместимого потомства при АВ0-несовместимых браках позволяет предположить, что гетерозис по АВ0-локусу основан на механизмах иммунного взаимодействия между матерью и плодом. Правда, механизмы гетерозиса могут быть множественными даже в отношении одного локуса, и поэтому вряд ли целесообразно настаивать на исключительности иммунного механизма. Действительно, локализация АВ0-субстанций в клеточных мембранах подавляющего большинства тканей свидетельствует об участии этих веществ в функциональной организации клетки и, следовательно, позволяет предполагать о других путях реализации гетерозиса по АВ0-локусу в эмбриогенезе. Вместе с тем иммунобиологический механизм гетерозиса лучше согласуется с представлениями об эмбриональном гомеостазе, поскольку другие эффекты гетерозиса обычно выявляются в экстремальных условиях.
По системе HLA также получены данные, свидетельствующие об иммунобиологической природе эмбрионального гетерозиса. Так, показано, что несовместимость по системе HLA благоприятно влияет на течение беременности и развитие плода. He получены убедительные данные о связи HLA-несовместимости со спонтанными выкидышами. Отбор в пользу несовместимого по HLA-системе потомства выявлен и при обследовании детей от кровнородственных браков, при которых гетерозиготность оставалась высокой, несмотря на тесный инбридинг.
Итак, по-видимому, гены АВ0-системы участвуют в процессе нормального эмбриогенеза, поэтому они не репрессируются. Благодаря неоднозначности своего проявления в различные периоды эмбриогенеза и таким эффектам, как плейотропия и гетерозис, гены АВ0-системы изменяют жизнеспособность эмбрионов, что иногда ведет к селективной гибели носителей определенных генотипов.
Гипотеза 4
На четвертый вопрос, действуют ли на внутриутробный отсев изменения в окружающей среде и некоторые другие факторы, помогают ответить полученные нами данные о влиянии медицинских абортов, токсикозов беременности и возрасте женщины.
Отсутствие влияния на отсев такого сильного вмешательства, как медицинские аборты, является достаточно веским аргументом против предположения о том, что изменения условий жизни повлекли за собой увеличение интенсивности отсева.
Отсутствие эффекта токсикозов беременности также подтверждает эту точку зрения. Однако увеличение материнского возраста не повысило интенсивность отсева, а лишь изменило его направленность. До сих пор мы не касались вопроса о причинах смены вектора отсева у повторнобеременных женщин. Возможно, что такое изменение у повторнобеременных женщин старшего возраста связано, во-первых, с некоторым возрастным изменением общих и местных иммунных реакций материнского организма па плод и, во-вторых, с увеличением вероятности внутриутробной гибели плода. При такой ситуации, естественно, лучше развиваются более несовместимые с матерью зародыши, что и ведет к изменению направления векторов отсева.
4. ГРУППЫ КРОВИ И ЭВОЛЮЦИЯ
Большинство популяций человека полиморфно по группам крови АВ0, т. е. содержат все три аллеля IA, IB, I0. На основе относительных частот этих аллелей жителей различных областей земного шара можно характеризовать как разные «серологические расы». Большинство рас содержат особей со всеми четырьмя группами крови 0, А, В, АВ и различаются только по распространенности отдельных групп. Однако у некоторых рас те или иные группы крови вовсе отсутствуют. Например, южноамериканские индейские племена были до прихода европейцев или остаются до сих пор изогенными по группе 0. Аллели IA, IB довольно редки также среди индейцев Северной Америки. Индейцы племен блад и черноногих, обитающие на юге Канады и в Монтане, представляют исключение; в отличие от южноамериканских племен они содержат аллель IA, причем частота этого аллеля очень высока, достигая 80%, чего не наблюдается ни в одной другой популяции человека. Американские индейцы произошли, по-видимому, от азиатской (монголоидной) расы, небольшая группа представителей которой в свое время пересекла Берингов пролив. Хотя у азиатской расы встречаются все четыре группы крови, частота группы 0 относительно мала; поэтому среди этих основателей, пришедших из Азии, или в небольших сформировавшихся позже изолятах могли сложиться условия, благоприятствовавшие дрейфу генов, и либо просто случай, либо случай в сочетании с естественным отбором мог привести к такому необычному распределению.
В большинстве популяций человека частота аллеля IA колеблется между 15 и 30%, а аллеля IB – между 5 и 20%. Например, частоты этих аллелей в больших популяциях эскимосов, распространенных по всему континенту от Аляски до Гренландии и Лабрадора, равны соответственно 30 и 6%. Но в небольшом племени северных эскимосов в Туле (Гренландия), которое насчитывает менее 300 человек и почти лишено притока генов из других популяций, частота IA составляет всего 9%. Кроме того, в некоторых изолятах Лабрадора и Баффиновой Земли ген IB совершенно отсутствует. По-видимому, все эти случаи можно отнести за счет дрейфа генов.
Интересные данные получены по общине дункеров в Пенсильвании (США). Дункеры (данкеры) — сектанты, эмигрировавшие в начале XVIII века из Германии и с тех пор живущие небольшими общинами, генетически почти изолированными от остального населения. Следовало ожидать, что частоты аллелей в этой группе будут промежуточными между частотами, наблюдаемыми на родине дункеров в Рейнской области, и частотами среди живущих вокруг них восточноамериканских популяций. Однако эти ожидания не подтвердились. Среди американцев, не принадлежащих к дункерам, так же как среди немцев (по данным 1939 года), люди с группой крови А составляют 40—45%, а среди дункеров — почти 60% (IA = 0,38); что касается частоты аллеля IB, то она снижена почти до нуля (IB = 0,025).
Есть много других примеров резких различий по частоте генов между небольшими родственными племенами или между религиозными изолятами и соседними популяциями с более высокой численностью. О том, что изоляция играет большую роль в такой дифференциации, свидетельствует совершенно иное распределение по частоте групп крови в больших популяциях человека. В больших смежных популяциях с практически неограниченной миграцией частоты генов обычно сходны.
Согласно концепции Форда, полиморфизм контролируется главными генами (или генами-переключателями), проявляющими заметное плейотропное действие. Появление и сохранение полиморфизма в популяциях обусловлено равновесием сил отбора, действующих на все развивающиеся при этом морфологические и физиологические признаки. Поэтому существование полиморфизма обычно свидетельствует о важности затрагиваемых им признаков. Можно предположить, что полиморфизм в отношении групп крови сбалансирован, и что селективное значение участвующих в этом генов обусловлено их влиянием на плодовитость, жизнеспособность или устойчивость к болезням.
В настоящее время изучена зависимость такой хорошо известной гемолитической болезни новорожденных как эритробластоз плода от Rh-несовместимости матери и плода. Вкратце, эритробластоз возникает у Rh-положительного плода, развивающегося во чреве Rh-отрицательной матери, сенсибилизированной чужеродными для нее Rh-антигенами плода во время данной или чаще предыдущих беременностей. Клетки крови плода содержат Rh-антиген. Если этот антиген попадет в кровоток матери, то он индуцирует образование в плазме матери Rh-антител (изоиммунизация). Это ведет к разрушению эритроцитов плода (гемолиз) и анемии.
Несовместимость матери и плода по антигенам системы АВ0 также ведет иногда к эритробластозу новорожденных или чаще к выкидышам на ранних сроках беременности. Одна из АВ0-несовместимых комбинаций такова: наличие у матери анти-А- или анти-В-антител против одноименных антигенов, имеющихся у плода. У женщин группы 0, в крови которых содержатся оба вида антител, вероятность гибели плода максимальна, а у женщин группы АВ – минимальна. Ознакомившись с основными генетическими и серологическими механизмами, характерными для систем групп крови, читатель может самостоятельно рассчитать потенциально неблагоприятные браки и показать, что внутриутробный отбор неизменно выступает во всех случаях против гетерозигот. Отсюда следует, что одни лишь силы отбора не могут обеспечить сохранение полиморфизма, поскольку отбор против гетерозигот теоретически приводит к состоянию неустойчивости, благоприятствующей закреплению наиболее распространенного аллеля.
Была установлена связь с АВ0-группами крови и ряда других заболеваний.
I (0) группа крови
. Частота среди больных язвой двенадцатиперстной кишки на 17% а среди больных язвой желудка на 10% выше среднепопуляционной. Среди лиц с группой крови 0 чаще встречается гипофизарная аденома.
II (A) группа крови.
Частота среди больных раком желудка на 10% выше среднепопуляционной. Сахарный диабет – на 8% выше. Среди лиц с группой крови А чаще встречаются переломы бедра, детская летальная пневмония, пернициозная анемия, рак женских половых органов.
Тем не менее, становится все более и более очевидной справедливость предположения Форда о том, что дивергенцию по частоте групп крови нельзя объяснить одними лишь случайными флуктуациями. Если бы в этом был замешан только дрейф генов, то вряд ли можно было бы ожидать, что частоты, обнаруженные во всех (а их было немало) изученных популяциях, займут лишь одну пятую всего возможного диапазона частот. Кроме того, установлено, что различия между популяциями по частоте аллелей системы АВ0 значительно больше, чем различия по аллелям систем MN и Rh; это показывает, что аллели систем АВ0 изменяются под влиянием внешних условий быстрее, чем другие аллели.
4.1 Отбор по системе групп крови АВ0
Рассмотрим некоторые проблемы, возникающие при анализе полиморфизма по системе групп крови АВ0. Один аспект отбора по этой системе – серологическая несовместимость матери и плода – общепризнан, хотя относительно интенсивности отбора общего мнения нет. Эта несовместимость приводит к нестабильному равновесию и к медленному изменению частот генов. В отсутствие других форм отбора полиморфизм по АВ0 должен постепенно исчезнуть. Однако, вопреки этому предсказанию, он присутствует почти во всех популяциях человека. Следовательно, по этой системе должны существовать другие селективные факторы. Рассмотрим возможность отбора по системе групп крови АВ0 на фоне инфекционных заболеваний.
4.2 Группы крови АВ0 и инфекционные заболевания
В настоящее время продемонстрирована ассоциация групп крови АВ0 с очень многими заболеваниями. Например, лица с группой крови А чаще заболевают раком, тогда как лица, имеющие группу крови 0, более подвержены к язве желудка и двенадцатиперстной кишки. Связь с группами крови показана и для ревматизма, иммунный механизм которого бесспорен: риск заболевания ревматизмом самый низкий среди лиц группы 0. Хотя эта связь, возможно, повышает вероятность достижения носителями группы 0 пожилого возраста, она вряд ли оказывает влияние на естественный отбор, поскольку ревматизм, как правило, поражает людей среднего и более старшего возраста, т.е. после завершения репродуктивного периода. Обнаруженные корреляции групп крови системы АВ0 с заболеваниями демонстрируют фундаментальное влияние антигенов этой системы на физиологию организма. Например, данные по ревматизму свидетельствуют о том, что это влияние может иметь какое-то отношение к иммунному ответу. Даже связь группы А с раковыми заболеваниями и группы 0 с язвой желудка может быть обусловлена различиями иммунного ответа.
Инфекционные заболевания оказывают специфическое воздействие на иммунный ответ организма. Если группы крови АВ0 влияют на иммунный ответ, то отбор, возникающий из-за дифференциальной восприимчивости к инфекционным заболеваниям, может привести к дифференциальной детской и юношеской смертности.
Распространение аллелей АВ0 в мировом населении
Распределение аллелей А, В и 0 свидетельствует о наличии естественного отбора по данной системе. Если бы отбора не было, а распределение отражало только случайные колебания генных частот, то в популяции присутствовали бы все комбинации генных частот, возможные в системе из трех аллелей. В действительности дело обстоит по-другому: наблюдается только небольшое число из возможных комбинаций.
Какая форма отбора могла привести к повышению аллельных частот в сравнительно изолированных популяциях или, наоборот, к их понижению в популяциях, расположенных «в русле» мировых миграций? Одна из вероятных возможностей – влияние инфекционных заболеваний, особенно крупных эпидемий прошлого.
Перечислим группы заболеваний, которые, возможно, имеют селективное значение:
а) эпидемические заболевания, например чума, холера, оспа;
б) хронические болезни, например туберкулез и сифилис;
в) кишечные инфекции (в основном детские);
г) тропические болезни (в детском и юношеском возрасте).
В отличие от полиморфных вариантов гемоглобина, которые встречаются только в тропических областях, полиморфизм по системе АВ0 распространен по всему миру. Следовательно, тропические болезни вряд ли играют роль селективного фактора в этой системе. Для формулировки рабочей гипотезы можно использовать три факта.
1. Заселение Америки человеком произошло примерно 10…12 (18) тыс. лет назад. До экспедиций Колумба население Центральной и Южной Америки было почти полностью изолировано от остальной части человечества. Возможно, что для американских популяций того времени была характерна особая группа инфекционных заболеваний, отсутствующих в остальной части мирового населения, например сифилис и связанные с ним болезни, вызываемые трепонемой (Treponema pallidum). Частота группы крови 0 в популяциях коренного населения Америки очень высока.
2. Эпидемии чумы многократно опустошали Европу, в основном поражая густонаселенные области. В краевых и частично изолированных популяциях, которые, вероятно, были затронуты чумой в меньшей степени, группа 0 обычно встречается с высокой частотой.
3. До середины 70-х годов во многих странах регистрировались случаи оспы. Имеются статистические данные о частоте встречаемости и смертности от оспы, особенно для населения Африки и Индии. Для этих регионов известно распределение групп крови АВ0. Если носители какой-то группы крови обладают повышенной восприимчивостью к оспе, то в областях с высоким уровнем заболеваемости эта группа должна встречаться реже.
Сифилис и группа крови 0
Сифилис – широко распространенное венерическое заболевание, которое вызывается особым микроорганизмом – бледной спирохетой (бледной трепонемой). Передается половым путем, через плаценту, реже – при использовании зараженных предметов. Как и другие венерические заболевания, сифилис – это не столько медицинская, сколько социальная проблема.
Антигенная структура бледной спирохеты очень сложна и до конца не изучена.
Перед нами стоит задача оценить возможное влияние сифилиса на дифференциальную биологическую приспособленность человеческой популяции. Как и в случае серповидноклеточности и малярии, наиболее убедительным косвенным доказательством такого влияния было бы доказательство того, что носители группы 0 имеют повышенную устойчивость к этой болезни. Это предположение в настоящее время проверить нельзя, так как сифилис настолько успешно вылечивается пенициллином, что индивидуальных различий в исходе болезни, обусловленных различием иммунного ответа, просто не существует. Однако в 1920-е годы лечение пенициллином еще не применялось. В это время был собран обширный материал по группам крови и заболеваемости сифилисом, анализ которого позволил сделать следующие выводы:
а) связь между заражением сифилисом и группами крови системы АВ0 отсутствует;
б) после лечения распространенным тогда препаратом неосальварсаном носители группы 0 имели гораздо большую вероятность стать «серонегативными», чем лица с другими группами крови.
в) третичный сифилис, так же как и общий паралич, реже встречается у лиц с группой 0, чем у лиц с другими группами крови системы АВ0.
Итак, все данные вместе свидетельствуют о том, что группа крови 0 имеет преимущество в иммунном ответе на сифилис. Известно, что влияние сифилиса на репродукцию осуществляется в основном путем заражения плода больной матерью. Зачастую такое заражение ведет к гибели плода на поздних стадиях развития. Можно высказать предположение, что широкое распространение группы 0 у индейцев Центральной и Южной Америки обусловлено отбором за счет сифилиса и связанных с ним инфекционных заболеваний, вызываемых трепонемой.
Холера и группа крови 0
Холера – это инфекционное заболевание, известное с древнейших времен. Её эпидемический очаг находится в бассейне рек Ганга и Брахмапутры (Индия, Бангладеш). Возбудителями холеры являются бактерии – холерные вибрионы (классический азиатский вибрион, вибрион Эль-Тор и другие). Антигены, сходные с антигенами человека, у холерных вибрионов не обнаружены. Дифференциальный диагноз холеры, позволяющий отличить ее от других острых кишечных инфекций (сальмонеллеза, дизентерии, отравлений), в прошлом не всегда был возможен; до начала XIX в. не исключалось смешение холеры с другими кишечными инфекциями.
Летальность при холере в прошлом составляла 50…60% (при заражении вибрионами Эль-Тор в 80…90% случаев отмечаются легкие формы). Непосредственной причиной смерти является обезвоживание организма.
У лиц, перенесших холеру, вырабатывается стойкий иммунитет. Вакцинация при холере малоэффективна, поствакциональный иммунитет сохраняется 6 месяцев.
Недавние обширные исследования холеры в Бангладеш убедительно показали существование связи между системой АВ0 и летально эндемичной инфекционной болезнью. Больные диареей, вызванной ротавирусом, шигеллой. токсикогенной Е. coli или нетоксикогенными возбудителями холеры, имели частоты группы 0, сходные с контрольными (около 30%), между тем больные, зараженные токсикогенным холерным вибрионом с группой крови 0, составляли 57%. Эта разница статистически высоко достоверна. Среди членов семей, зараженных токсикогенным штаммом холеры, наблюдается статистически достоверная тенденция к повышению частоты группы крови 0 с увеличением тяжести протекания диареи. Эпидемии тяжелых кишечных заболеваний, описанные в прошлом в этом регионе, вероятнее всего, были эпидемиями холеры. Низкая частота группы крови 0 в этом регионе, возможно, обусловлена более высокой восприимчивостью к холере и смертностью от нее лиц с группой крови 0. Механизм этого взаимодействия остается невыясненным.
Чума и группа крови 0
Чума – это общее особо опасное инфекционное заболевание. Различают следующие формы (проявления) чумы: бубонную, легочную, кишечную, септическую, кожную.
Возбудитель чумы – чумная палочка – характеризуется высокой изменчивостью (вирулентные R-формы, авирулентные S–формы, промежуточные О-формы; изменение свойств происходит под влиянием бактериофагов). У чумной палочки обнаружены антигены, сходны с антигеном 0 системы АВ0 (подробнее см. ниже).
Постоянным резервуаром чумы в природе являются грызуны (свыше 300 видов) и другие животные. Чума передается воздушно-капельным и воздушно-пылевым путем; наибольшую роль в распространении чумы играют блохи, в кишечнике которых чумная палочка размножается.В прошлом эпидемии чумы неоднократно приводили к гибели множества людей: летальность достигала 40…100%. В Древнем Риме страшная эпидемия чумы свирепствовала в 293 г. до н.э. В середине XIV в. (1345–1352 гг.) в Европе от чумы умерло около 50 млн. человек (по другим оценкам – 25 млн.); в 1665 г. бубонная чума унесла жизни 10 млн. человек. В России одна из сильнейших эпидемий чумы совпала по времени с пугачевским восстанием.
У переболевших чумой людей вырабатывается стойкий и длительный иммунитет. Вакцинация при чуме малоэффективна, поствакциональный иммунитет сохраняется 6…12 месяцев.
С повышением качества жизни (улучшение питания и санитарно-гигиенических условий) и появлением антибиотиков (стрептомицина) чума перешла в разряд очень редких болезней; она встречается только в областях, недоступных для исследователей. Поэтому приходится анализировать косвенные данные.
При частотно-зависимом отборе одним из способов адаптации паразита к хозяину является выработка поверхностных антигенов, аналогичных антигенам хозяина: таким образом обеспечивается его защита от иммунного ответа. Данные, подтверждающие существование такого способа адаптации, получены для позвоночных и их паразитов. С конца 1950-х годов известно, что человек имеет общие АВН-подобные антигены со многими бактериями, особенно с бактериями группы Е. coli. Даже «нормальные» изоантитела анти-А и анти-В считаются иммунными антителами против широко распространенных кишечных инфекций. Поэтому интересно было выяснить, имеет ли чумная палочка ABH-подобные антигены. Такой антиген действительно обнаружен (антиген Н). Он распространен в основном среди лиц с группой крови 0. Этот факт, во-первых, свидетельствует о том, что обладатели первой группы крови имеют более слабую иммунную реакцию против чумной палочки и, во-вторых, согласуется с предположением о селективной невыгодности группы 0 вследствие высокой смертности её носителей от чумы. Конечно, эти результаты нельзя считать подтверждением гипотезы.
Группы крови АВ0 и оспа
Оспа – это эпидемическое особо опасное заболевание, поражающее весь организм человека. Оспа вызывается ДНК-содержащим вирусом. У этого вируса обнаружены антигены, сходные с антигеном А системы АВ0. Поэтому лица с группами крови А и АВ наименее устойчивы к этому заболеванию.
Источником инфекции является больной человек. Вирус оспы передается воздушно-капельным и воздушно-пылевым путем, а также при прямом контакте через нарушенные покровы тела. Различают несколько формы оспы: типичную, легкую, сливную, геморрагическую (черную), оспенную пурпуру. Смертность при типичной и легкой форме оспы составляла ~6% (иногда до 15…20%), при сливной ~45%, при черной и пурпурной оспе – до 100%.
Оспа известна с древнейших времен: мумии египетских фараонов возрастом более 6 тысяч лет несут следы перенесенной оспы. Однако массовое распространение оспы в Старом Свете относится к V…VI вв. н.э. Около 910 г. н. э. арабские медики дали первые четкие описания оспы. В Средние века в некоторых государствах Европы от оспы погибало до половины населения.
В начале XVI в. оспа была занесена в Новый Свет. В результате за несколько лет в Мексике от оспы погибло ~3,5 млн. человек, что стало одной из причин гибели цивилизации майя. В дальнейшем от оспы ежегоно погибали сотни тысяч человек; всего от этой болезни погибла треть коренного населения Южной Америки. В XVII в. оспа проникла в Северную Америку; в результате эпидемий этой болезни вымирали целые племена североамериканских индейцев. В России до 1917 г. оспой ежегодно переболевало 100 тысяч человек, из которых 20 тысяч умирали.
С незапамятных времен до конца XVIII в. в качестве профилактики против оспы использовалась вариоляция (оспопрививание) – прививка заразным материалом от больных оспой. В России вариоляция насаждалась Екатериной Великой. Однако вариоляция неоднократно сопровождалась осложнениями. После работ Э. Дженнера (1796 г.) надежной защитой от оспы стала вакцинация, которая дает стойкий (хотя и не пожизненный) иммунитет.
В 1958 г. по инициативе СССР была развернута программа ВОЗ по ликвидации оспы. В 1980 г. ВОЗ объявила о ликвидации этой болезни во всем мире.
После разработки гипотезы о том, что распределение групп крови АВ0 в популяциях человека может быть связано с крупными эпидемиями, а решающим фактором является общность антигенов возбудителя и хозяина, было решено исследовать активность АВН у возбудителя оспы. По чисто методическим причинам эти исследования провели не на вирусе натуральной оспы (вариола), а на близко родственном ему вирусе оспы коров. Была выявлена высокая активность антигена А, что с очевидностью предполагает наличие иммунологического механизма, определяющего ассоциацию с данной болезнью. При попадании в организм человека вирус, имеющий антиген А, будет частично инактивирован антителами анти-А, которые присутствуют только у людей с группами крови В и 0. У лиц с группами крови А и АВ (у них этих антител нет) инактивации вируса не произойдет. Следовательно, можно ожидать, что у больных с группами крови А и АВ болезнь будет протекать тяжелее. Поскольку оспа поражает детей и часто заканчивается смертельным исходом, различия по группам крови должны оказывать сильное влияние на отбор.
Было высказано предположение, что обнаруженный антиген А принадлежал не самому вирусу, а среде, в которой вирус выращивался: тогда вирус может включать материал хозяина в собственный капсид. Сейчас получены данные в пользу существования такого механизма. Ими объясняют различную клиническую реакцию на заражение вирусом гепатита В. Вероятно, этот вирус включает компоненты белков сыворотки (особенно гамма-глобулина) и переносит их к новому хозяину, иммунная реакция которого может зависеть (по крайней мере, частично) от сходства этих компонентов с его собственными белками.
Исследования ассоциации с группами крови, проведенные на больных оспой, дали противоположные результаты. Предположение, что лица, с группами крови А и АВ будут чаще и тяжелее болеть оспой, проверялось многократно, но результаты были получены неоднозначные. В одной из работ, в которой исследовались 986 случаев оспы (давних и возникших вновь), показано, что относительная частота заболеваемости значительно выше у лиц с группами А и АВ по сравнению с В и 0. Та же тенденция обнаружилась при анализе тяжести клинических симптомов и смертности. Более того, среди людей, переживших эпидемии оспы. наблюдался небольшой избыток обладателей групп крови В и 0, что указывает на более высокую смертность лип с группами крови А и АВ. Среди выживших больных тяжелые поражения кожи наблюдались чаще у лиц с группами А и АВ. Это исследование было проведено в индийской деревне во время эпидемии оспы; в качестве контроля использовались сибсы пораженных пробандов, которые остались здоровыми, несмотря на то, что также соприкасались с инфекцией. Почти все жители деревни, как заболевшие, так и здоровые, никогда не подвергались вакцинации. Такая постановка эксперимента подчеркивает разницу в распределении групп крови у больных и здоровых индивидов и в то же время до минимума снижает возможность ошибки, связанной со стратификацией популяции.
Обследование пациентов городских больниц Индии и изучение более легкой формы оспы в Бразилии не подтвердили ассоциацию оспы с группами крови. Это расхождение, возможно, объясняется тем, что рассмотренное выше исследование оспы в деревнях Индии выполнено в основном на детях. В работах, проведенных в городских больницах Индии, возрастное распределение пациентов не приводится, однако на основе некоторых косвенных сведений можно предположить, что большинство больных были взрослыми.
При серповидноклеточной анемии селективное преимущество гетерозигот HbA/HbS характерно только для детей раннего возраста; выжившие взрослые высокоиммунны независимо от того, каким типом HbS они обладают. Вполне возможно, что это справедливо и в случае оспы. До ликвидации оспы эта болезнь в некоторых областях Индии была почти эндемической. Действительно, высокий титр препятствующих гемагглютинации антител вариолы был найден у многих жителей этих областей, никогда не имевших клинических проявлений оспы и не подвергавшихся вакцинированию. Однако данные по этому вопросу остаются противоречивыми, а возможность его окончательного решения невелика из-за практически полной ликвидации оспы.
5. МАТЕРИАЛЫ И МЕТОДЫ
5.1 Объём исследования
В ходе работы были исследованы популяции нескольких областей Украины: Житомирской (41 человек), Черновицкой (81), Закарпатской (23), Запорожской (г. Запорожье – 53, г. Бердянск – 25), Сумской (78), Черкасской (137). Путём опроса была установлена принадлежность каждого представителя популяции к группе крови по системам АВ0 и Rh.
При анализе результатов использовались следующие подходы:
1. Закон Харди-Вайнберга;
2. Расчёт статистической ошибки выборочной доли, выраженной в процентах;
3. Дисперсионный анализ качественных признаков
5.2 Закон Харди-Вайнберга
Закон Харди-Вайнберга гласит, что процесс наследственной преемственности сам по себе не ведет к изменению частот аллелей и (при случайном скрещивании) частот генотипов по определенному локусу. Более того, при случайном скрещивании равновесные частоты генотипов по данному локусу достигаются за одно поколение, если исходные частоты аллелей одинаковы у обоих полов.Равновесные частоты генотипов задаются произведениями частот соответствующих аллелей. Если имеются только два аллеля, А и а, с частотами р и q, то частоты трех возможных генотипов выражаются уравнением:
(p+q)2
= p2
+ 2рq + q2
А а АА Аа аа,
где буквам во второй строке, обозначающим аллели и генотипы, соответствуют расположенные над ними частоты в первой строке.
Если имеются три аллеля, скажем А1, А2 и А3, с частотами р, q и r, то частоты генотипов определяются следующим образом:
(p+ q + r)2
= р2
+ q2
+ r2
+ 2pq+ 2рr + 2qr
А1 А2 А3 А1А1 А2А2 А3А3 А1А2 А1А3 А2А3
Аналогичный прием возведения в квадрат многочлена может быть использован для определения равновесных частот генотипов при любом числе аллелей. Заметим, что сумма всех частот аллелей, так же как и сумма всех частот генотипов, всегда должна быть равна 1. Если имеются только два аллеля с частотами р и q, то р + q = 1, и, следовательно, р2
+ 2рq + q2
= (р + q)2
= 1; если же имеется три аллеля с частотами р, q и r, то р + q + r = 1, и, следовательно, также (р + q + r)2
=1 и т.д.
5.3 Расчёт статистической ошибки выборочной доли, выраженной в процентах

где р% - выборочная доля в процентах, п - численность выборки.
5.4 Дисперсионный анализ качественных признаков
Построение дисперсионного комплекса для анализа качественных признаков имеет ряд особенностей: поскольку дисперсионному анализу подвергаются доли, в градации заносят число объектов с наличием признака и общее число объектов. Сумма квадратов доли вычисляется по формуле:

Где
 - -
доля объектов с признаком, m – число объектов с признаком, n – общее число объектов в градации.
Из этой формулы следует, что:

Формулы для вычисления дисперсий (сумм квадратов) для долей:

где N – общее число объектов в дисперсионном комплексе, n1, n2, … - число объектов в первой, второй и последующих градациях, m – общее число объектов с данным признаком, m1, m2, … - число объектов с данным признаком в первой, второй и последующих градациях.
С рассчитанными суммами квадратов производят те же действия, как и в дисперсионном анализе количественных признаков: находят факториальные, случайные и общие средние квадраты, степени свободы, вычисляют показатель силы влияния, статистический критерий.
5.5 Результаты и обсуждения
Таблица 2
Фенотипическая структура популяций по группам крови по системе AB0 разных областей Украины, %
| Регион |
Частоты фенотипов |
| I(0) |
II(А) |
III(В) |
IV(АВ) |
| Житомирская область |
31±7,21 |
42±7,68 |
17±5,83 |
10±4,69 |
| Черновицкая область |
30±5,09 |
52±5,57 |
13±3,74 |
5±2,45 |
| Закарпатская область |
30±9,56 |
43±10,34 |
17±7,83 |
10±6,26 |
| Запорожская область г. Бердянск |
44±9,93 |
28±8,98 |
16±7,33 |
12±6,49 |
| Запорожская область |
28±6,16 |
45±6,83 |
17±5,16 |
10±4,12 |
| Сумская область |
30±5,19 |
40±5,55 |
24±4,83 |
6±2,69 |
| Черкасская область |
33±4,02 |
43±4,23 |
17±3,21 |
7±2,18 |
Таблица 3
Фенотипическая структура популяций по группам крови по системе AB0 разных областей Украины, %
| Регион |
Частоты генотипов |
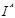  |
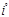 |
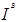 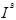 |
 |
|
  |
| Житомирская область |
9 |
33 |
2 |
17 |
30 |
9 |
| Черновицкая область |
12,1 |
37,1 |
1,2 |
12,1 |
30 |
7,5 |
| Закарпатская область |
10,2 |
35,3 |
1,7 |
14,4 |
30 |
8,4 |
| Запорожская область, г. Бердянск |
4,7 |
29 |
1,3 |
15,7 |
44 |
5,3 |
| Запорожская область |
10,9 |
35 |
2 |
14,9 |
28 |
9,2 |
| Сумская область |
7,4 |
29,8 |
3,3 |
19,8 |
30 |
9,7 |
| Черкасская область |
8,5 |
33,7 |
1,8 |
15,4 |
33 |
7,6 |
Таблица 4
Частоты аллелей групп крови по системе АВО у популяций разных областей Украины, %
| Регион |
Частоты аллелей |
 |
 |
 |
| Житомирская область |
30 |
15 |
55 |
| Черновицкая область |
34 |
11 |
55 |
| Закарпатская область |
32 |
13 |
55 |
| Запорожская область, г. Бердянск |
22 |
12 |
66 |
| Запорожская область |
33 |
14 |
53 |
| Сумская область |
27 |
18 |
55 |
| Черкасская область |
29 |
13 |
58 |
Таблица 5
Частоты фенотипов групп крови по системе Rh у популяций разных областей Украины, %
| Регион |
Частоты фенотипов |
| Rh + |
Rh - |
| Житомирская область |
88±5.09 |
12±5.09 |
| Черновицкая область |
83±4.12 |
17±4.12 |
| Закарпатская область |
83±7.83 |
17±7.83 |
| Запорожская область, г. Бердянск |
88±6.49 |
12±6.49 |
| Запорожская область |
81±5.39 |
19±5.39 |
| Сумская область |
85±4.04 |
15±4.04 |
| Черкасская область |
83±3.21 |
17±3.21 |
Таблица 6
Частоты генотипов групп крови по системе Rh у популяций разных областей Украины, %
| Регион |
Частоты генотипов |
| RR |
rr |
Rr |
| Житомирская область |
81 |
12 |
7 |
| Черновицкая область |
69 |
17 |
14 |
| Закарпатская область |
69 |
17 |
14 |
| Запорожская область, г. Бердянск |
42 |
12 |
46 |
| Запорожская область |
31 |
19 |
50 |
| Сумская область |
37 |
15 |
48 |
| Черкасская область |
69 |
17 |
14 |
Таблица 7
Частоты аллелей групп крови по системе Rh у популяций разных областей Украины, %
| Регион |
Частоты аллелей |
| R |
r |
| Житомирская область |
65.5 |
34.5 |
| Черновицкая область |
83.2 |
16.8 |
| Закарпатская область |
83.2 |
16.8 |
| Запорожская область, г. Бердянск |
65.5 |
34.5 |
| Запорожская область |
56 |
44 |
| Сумская область |
61 |
39 |
| Черкасская область |
83.2 |
16.8 |

Рис. 6. Круговые диаграммы распределения частот фенотипов по группам крови по системе АВ0
Список литературы
1. Айала Ф., Кайгер Дж. Современная генетика т.3. – М.: Мир, 1988.
2. Амеханян С.И., Акифьев А.П. Общая генетика. – М.: высшая школа, 1985.
3. Атраментова Л.А., Утевская О.М. Статистические методы в биологии. – Горловка, 2008.
4. Васецкий С.Г. Генетика и наследственность. – М.: Мир, 1987.
5. Грушевский М. Иллюстрированная история Украины. – Донецк: Бао, 2001.
6. Дубинин Н.П. Генетика. – Кишинёв: Штиинца, 1985.
7. Инге-Вечтомов С.Г. Генетика с основами селекции. – М.: Высшая школа, 1989.
8. Инге-Вечтомов С.Г. Введение в молекулярную генетику. – м.: Высшая школа, 1983.
9. Каминская Э.А. Общая генетика. – Минск: Высшая школа, 1982.
10. Корочкин Л.И. Геном, клонирование, происхождение человека. – Фрязино: Век 2, 2004.
11. Смиряес А.В., Кильчевский А.В. Генетика популяций и количественных признаков. – М.: КолоС, 2007.
12. Стрельчук С.І., Демидов С.І. Генетика з основами селекції. – К.: Фітосоціоцентр, 2000.
13. Харрисон Дж. И др.. Біологія человека. – М.: Мир, 1979.
14. Электронный источник // Le mappe genetiche di Cavalli Sforza -http://s155239215.onlinehome.us/turkic/63_Blood_Types/Blood_TypesRu.htm
15. Электронный источник // http://afonin-59-bio.narod.ru/2_heredity/2_heredity_individual/her_ind_07.htm
16. Электронный источник // William C. Boyd Ph.D. and Isaac Asimov Ph.D Генетика групп крови // http://www.balzan.it/english/pb1999/cavalli/paper.htm
|