Николай Клягин
Современная научная картина мира

«Клягин Н.В. Современная научная картина мира»: Логос; Москва; 2007
Аннотация
Рассматривается современная научная картина мира, охватывается широчайший спектр данных космогонии, биогенеза, антропогенеза, социогенеза и др. Эти данные настолько разнородны, что подпадают под общий знаменатель только на уровне универсального причинно-следственного закона. Для понимания его происхождения необходимо объяснить, как возникли законы сохранения и неубывания энтропии. Чтобы протянуть нить причинности от физического и астрономического мира к человеческому обществу, нужно осмыслить пути и результаты биоэволюции. Все это дает нам возможность понять, где находится центр Вселенной, как и почему возникли и угасли динозавры, отчего человек единственный на планете обладает наукой и искусством.
Для студентов высших учебных заведений, изучающих курс «Концепция современного естествознания». Может использоваться аспирантами при подготовке к экзамену по кандидатскому минимуму в области истории и философии науки.
Оглавление
Введение
Глава 1 . Вселенная
1.1. Микромир
1.2. Макромир
1.3. Сингулярность
1.4. Энтропия
1.5. Закон сохранения
1.6. Причинно-следственный закон
1.7. Мироздание за пределами Вселенной
1.8. Симметрия
Глава 2 . Земная жизнь
2.1. Биогенез
2.2. Геология
2.3. Эволюция
Археозой (4,55-2,5 млрд лет назад)
Протерозой (2,5-0,544 млрд лет назад)
Палеозой (544-251 млн лет назад)
Мезозой (251-65 млн лет назад)
Кайнозой (65-0 млн лет назад)
2.4. Гоминизация
Неотения и акселерация
Демография
Красота
Глава 3 . Человек
3.1. Материальная культура
Технология
Социальность
3.2. Духовная культура
Интеллект
Религия
Нравственность
Искусство
3.3. Цивилизация
Заключение
Приложение 1 . Палеолитическое искусство
Приложение 2 . Статистика сюжетов палеолитического искусства
Приложение 3 . Местонахождения мобильного искусства нижнего и среднего палеолита (стиль 0)
Приложение 4 . Местонахождения франко-кантабрийского наскального искусства
Приложение 5 . Хронология кайнозойских оледенений
Приложение 6 . Классические археологические культуры палеолита
Литература:
Приложение 7 . Расчетная убыль постоянной Хаббла к окраинам Вселенной
Библиография
Введение
Людям свойственно делить окружающий мир на явления, данные в ощущениях, и сущности вещей, постигаемые мысленно. Например, всем знакомы простодушные воробьи, изящные трясогузки и мудрые вороны, но никто и никогда не видел «птицу вообще», хотя суждения об этом есть в любом пособии по зоологии. Конкретные явления воплощают как типичные, так и неповторимые особенности объектов, и двух абсолютно одинаковых явлений мы не найдем. Сущности вещей, напротив, выражают постоянно встречающиеся черты явлений определенного класса и дают нам общее представление о нем, в силу чего мы знаем, что такое «человек вообще», «планета вообще», «дождь вообще» и т. д. Может быть, сущности явлений – это просто слова, обозначающие сходные объекты? Да, это так. Сущности – это понятия, фиксируемые словами. Однако их анализ с точки зрения количественной теории информации К.Э. Шеннона показывает, что реальное положение дел куда сложнее (см. разд. 3.2).
Вопрос о природе сущностей захватил воображение уже первых древнегреческих философов. Они решали проблему чрезвычайно смело. Для них было мало понять, что такое, например, «камень вообще» – они пытались разгадать, что такое бытие, материальный мир вообще, и какими общими принципами он управляется. Собственно, благодаря такой интеллектуальной дерзости эти мыслители и сохранились в памяти многих поколений.
Деталей творческой лаборатории первых философов мы, к сожалению, не знаем, но догадываемся, что, ориентируясь на доисторические верования о водной колыбели бытия, возможно, дошедшие до них, Фалес Милетский (ок. 625 – ок. 547 до н. э.) усмотрел первооснову всех вещей в водной стихии [104, с. 100–115]. Первый классический философ полагал, что, испаряясь, вода способна порождать воздух, а сгущаясь, – земную твердь, что не лишено оснований, особенно если принять во внимание скудость научных знаний. Его ученик Анаксимандр (610 – ок. 540 до н. э.) возвысился до воззрений на природное первоначало как на некий апейрон, «бесконечное» [104, с. 116–129]. Выражаясь языком нашего времени, мы допускаем, что древний философ приблизился к понятию материи как философской категории для обозначения объективной реальности, данной человеку в ощущениях [64, с. 131]. Подобное определение материи беспроигрышно, поскольку под него подходит любой материальный объект. По той же причине оно неконкретно и дает нам мало знаний о первооснове бытия. Что же оно означает вообще?
На этот вопрос, похоже, отвечает современная физика – точнее, так называемая физическая Теория Всего Сущего (ТВС). За первоэлемент физического мира она принимает суперструны – складки мировой поверхности, непрерывно перемещающиеся и колеблющиеся с релятивистскими скоростями, т. е. со стремительностью, близкой к скорости света (см. разд. I.1). Этот заманчивый подход заведомо рискован. Он правомерен применительно к положению дел в нашей Вселенной. Однако мы не вправе исключать, что за ее пределами существуют иные миры (см. разд. 1.7), в которых материя организована иными способами, нежели в форме суперструн.
У суперструн есть своя философская биография. В известном смысле подобный первоэлемент материального бытия интуитивно был предвосхищен Гераклитом Эфесским (ок. 520 – ок. 460 до н. э.) в образе стихии неугомонного огня [104, с. 176–257]. Действительно, мировая поверхность, вспыхивающая гребешками суперструн, больше всего напоминает пожар на нефтепромысле. Иносказательный философ двинулся еще дальше. Он полагал, что космос периодически разгорается и угасает. Подобный образ мироздания типологически напоминает космологическую модель пульсирующей Вселенной, которая то рождается в Большом Взрыве и бурно расширяется, то схлопывается в Большом Коллапсе (по-английски говорят в «Большом Схрусте», Big Crunch) [724]. Разумеется, чересчур модернизировать античные взгляды не следует. Однако вряд ли можно отрицать, что многие коцептуальные образы современной науки родились не сегодня. Так, Левкипп и его ученик Демокрит (ок. 460 – ок. 360 до н. э.) создали учение об атомах, усвоенное с надлежащими уточнениями современной физикой.
Венцом гераклитовской мысли стал логос – своего рода универсальный закон взаимопревращения всех вещей, послуживший основой философского учения о диалектике. Подметив, что в больших масштабах отрезок окружности представляется прямой линией (подобно тому, как обозримая сферическая земная поверхность выглядит плоской), Гераклит справедливо заключил, что противоположные состояния объектов не абсолютны, а представляют собой крайние пункты в цепочке своих переходных состояний. В рамках этой философской традиции Г. В. Ф. Гегель (1770–1831) создал грандиозную картину развертывания некой абсолютной идеи, порождающей все наблюдаемые и мыслимые вещи сообразно законам диалектики [26]. Можно ли полагать, что «общая теория всего на свете» [63, с. 269] состоялась? По-видимому, это было бы преждевременно, а существо затруднения состоит в следующем.
Известные нам законы природы, в том числе законы диалектики, поддерживают устойчиво повторяющиеся связи явлений и их сущностей, что обеспечивает стройность, упорядоченность и целостность нашей Вселенной. Для всех этих законов характерна повторяемость, которая, в свою очередь, обусловлена причинно-следственным законом
. Последний означает, что в сопоставимых условиях из одинаковых причин вытекают сходные следствия. Почему так происходит, науке не известно. Тем не менее ясно, что без причинно-следственного закона не смог бы действовать ни один закон природы или общества, а мироустройство распалось бы, чего, однако, не наблюдается. Таким образом, открывается неожиданное обстоятельство, что глубже всех мыслимых законов любой степени фундаментальности всегда лежит закон причинности (каузальности).
Таким образом, для понимания природы вещей нашего мира требуется объяснить происхождение закона каузальности. Для его функционирования необходимо совместное действие двух факторов. Во-первых, закон причинности нуждается в том, чтобы что-то вообще происходило и менялось, а во-вторых, причинно-следственный закон не обойдется без того, чтобы при этом что-то сохранялось неизменным. Следовательно, нам придется объяснить, почему во Вселенной наблюдается течение событий и при этом действуют законы сохранения (энергии и пр., о чем подробнее см. разд. 1.4–1.6).
Решение этих непростых задач потребует анализа проявлений каузальности в основных сферах реальности, что даст нам современную научную картину мира в наиболее существенных ее чертах. Мы предполагаем проследить действие закона причинности в наиболее важных областях и феноменах действительности. Во-первых, возникновение Вселенной (см. гл. 1). Во-вторых, зарождение и развитие жизни (см. гл. 2). В-третьих, становление человека, появление материальной и духовной культуры, а также происхождение цивилизации (см. гл. 3). Нашей задачей будет показать, что перечисленные события шли закономерно под влиянием причинности.
Действие закона каузальности способно объяснить поступательность в развитии нашего мира. В самом деле, Вселенная зреет, стареет и не проявляет тенденций к омоложению. В развитии земной жизни господствует закон Долло о необратимости эволюции, поэтому на смену примитивным формам жизни приходят более сложные организмы, в результате чего биологическая эволюция протекает прогрессивно. В социальной жизни мы тоже наблюдаем общий поступательный прогресс – отдельные случаи социального регресса лишь подтверждают правило. Иными словами, из причин вытекают следствия, но не наоборот, хотя следствия могут влиять на причины, меняя условия их действия. Например, если холодильник станет производить холод без остановки, то в конце концов замерзнет сам и перестанет производить холод. Это явление называется в кибернетике принципом обратной связи. Однако оно не отменяет закона причинности, а всего лишь искажает его проявления за счет нарушения условий выполнения каузального закона, что подразумевается его определением (он осуществляется без искажений лишь в сходных условиях).
Казалось бы, общеизвестно, что немногое на свете протекает беспричинно. Однако с философской точки зрения ситуация представляется куда более сложной. В самом деле, если причинность распространена тотально и способствует повторяемости событий, то во Вселенной повторяющегося, однотипного должно быть так много, что мир выглядел бы однообразно. Наш опыт свидетельствует о противном. Очень многое в природе и обществе представляется оригинальным и неповторимым. В принципе, это обстоятельство противоречит убеждению в глубокой фундаментальности причинно-следственного закона. Именно поэтому он никогда не привлекал специального внимания мыслителей. Отсюда наша задача состоит в том, чтобы продемонстрировать, что в природе и обществе гораздо больше типологически подобных явлений и сущностей, чем кажется на первый взгляд.
Глава 1
Вселенная
В данной главе мы предполагаем рассмотреть:
1) новейшие сведения об устройстве микромира, составляющие основу так называемой Теории Всего Сущего, опирающейся на физические теории суперструн, суперсимметрии и супергравитации;
2) новые факты и гипотезы относительно строения Вселенной, которая в свете ряда недавних парадоксальных открытий астрофизики представляется анизотропной, неоднородной, имеющей центр и периферию;
3) акаузальную («беспричинную») природу загадочной сингулярности, положившей начало нашей Вселенной и обладавшей необычными свойствами;
4) происхождение Второго начала термодинамики, или закона неубывания энтропии, которое в свете современных представлений поддается объяснению;
5) происхождение фундаментальных законов сохранения (энергии, массы, квантовых чисел), которое также поддается естественнонаучному объяснению;
6) происхождение причинно-следственного закона, основополагающего для всех прочих законов природы, также поддающегося естественнонаучному объяснению;
7) современные гипотезы происхождения нашей сингулярности и природы мироздания, окружающего нашу Вселенную, анизотропная природа которой предполагает некое внешнее бытие;
8) общие черты в организации законов природы на разных уровнях и в различных сферах бытия – черты, поддающиеся не метафизическому, а естественнонаучному истолкованию, хотя и ведущемуся в духе философского обобщения.
1.1. Микромир
Современная физическая картина микромира строится на основе теорий суперсимметрии [5; 21, с. 9–113; 24; 27; 101, с. 17–56; 304], супергравитации [21, с. 121–170; 101, с. 57–68] и суперструн [32; 33; 9; 48; 101, с. 277–304; 351; 369; 370; 551; 791]. Приставка «супер» означает, что при очень высоких энергиях ранней Вселенной существующие ныне первичные элементарные частицы имели так называемых суперпартнеров, отличающихся квантовым числом (свойством), спином
, или собственным угловым моментом количества движения, выраженным в постоянной Планка (в кванте действия ħ
= h
/(2π) = 6,6260688(37) × 10–34Js
/(2π) [618], или ħ
= h
/(2π) = 6,6260755(40) × 10–34Js
/2π [650]).
Современная наблюдаемая Вселенная сложена всего из двенадцати кирпичиков вещества – элементарных частиц со спином 1/2, называемых фермионами
. В их число входят шесть относительно массивных кварков
(с массой до 178,0 ± 4,3 ГэВ (гигаэлектронвольт) у истинного кварка
[125, с. 638]) и 6 относительно легких лептонов
, состоящих из тройки электроноподобных частиц (электрон
, мюон
и тау-частица
) и тройки нейтрино (электронное
, мюонное
и тау-нейтрино
с массами от 0,07–11 эВ (электрон-вольт) у электронного нейтрино (рис. 1) [597; 158; 276; 528; 530; 606; 740]).
В первые мгновения после Большого Взрыва, породившего наш мир, когда Вселенная была неимоверно горячей (1033К), элементарные частицы двигались настолько энергично, что сближались друг с другом на минимальные дистанции. От расстояний взаимодействия зависят массы участвующих в нем частиц: чем короче дистанции, тем предельные массы взаимодействующих частиц больше. Поэтому в те времена наши обычные фермионы имели массивных двойников-суперпартнеров со спином 0, т. е. бозонов (см. ниже). Нынешним кваркам отвечали суперкварки, или скварки; нынешним электронам, мюонам и тау-частицам соответствовали сэлектроны
, смюоны
и стау-частицы
, а теперешним нейтрино – три снейтрино
. Когда Вселенная расширилась достаточно, чтобы заметно остыть, характерные дистанции взаимодействий между частицами возросли, а массивные суперчастицы перестали проявляться и в известном смысле вымерли. В принципе их можно получить на сверхмощных ускорителях (суперколлайдерах), однако необходимые для этого энергии пока недостижимы.


Рис. 1.
Фундаментальные элементарные частицы (суперструны – первоосновы физического микромира). На схеме опущены античастицы, которые отличаются от обычных частиц ориентацией спина (у нейтрино) или знаком электрического заряда (так, античастицей отрицательно заряженного электрона является положительно заряженный позитрон, а W–-бозон является античастицей для W+-бозона). Под каждой частицей в скобках дана ее масса в млрд электрон-вольт (ГэВ).
Физические взаимодействия между фермионами осуществляются бозонами
– частицами с целочисленными спинами (0, 1, 2). Массу покоя от частицы к частице передают бозоны Хиггса
(спин 0, масса 96–117 ГэВ, или ок. 115 ГэВ [125, с. 639; 640]). Четыре основные силы природы – сильное взаимодействие, электромагнетизм, слабое взаимодействие и гравитация – переносятся, соответственно, глюонами
(спин 1, масса 0), фотонами
(спин 1, масса 0), Z-
и W-бозонами
(спин 1, масса 93 и 81 ГэВ) и гравитонами
(спин 2, масса 0).
Глюоны склеивают пары кварков в мезоны
, а тройки кварков – в нуклоны
: протоны
(время жизни св. 1031 лет [34, с. 61]) и нейтроны
(полураспад 10 мин 16 с [374]), а также нуклоны – в ядра атомов. Фотоны обслуживают все электромагнитные взаимодействия, Z– и W-бозоны – превращения кварков и прочих частиц, а гравитоны – взаимопритяжение частиц и особенно крупных их скоплений, физических тел, так как поодиночке гравитоны предельно слабы и становятся заметны лишь при массовых испусканиях от больших серий фермионов, собранных в массивные физические тела, а именно – в астрономические объекты. При повышенных энергиях проявления слабого и электромагнитного взаимодействий сливаются в электрослабой силе. В эпоху ранней, горячей Вселенной перечисленные бозоны имели суперпартнеров-фермионов со спином 1/2: бозонам Хиггса отвечали хиггсино
, фотонам – фотино
, глюонам – глюино
, а Z– и W-бозонам – зино
и вино
.
Древним партнером гравитона являлся гравитино
со спином 3/2. Суперпартнер последнего, гравифотон
со спином 1, стягивает антивещество и расталкивает обычное вещество, т. е. может послужить окном в антигравитацию; ср. [327]. Его суперпартнер, голдстино
со спином 1/2, в свою очередь, располагает суперпартнером гравискаляром
со спином 0 (т. е. имеется пара фермионов на три бозона).
Агент миллислабого и сверхслабого взаимодействия (5-й силы) аксион
(с массой менее 0,001 эВ) обусловливает нарушение симметрии зарядового сопряжения и четности (это – неравноправие вещества и антивещества при некоторых слабых взаимодействиях: например, преобладание рождений позитронов при распадах нейтральных К-мезонов на веществе и антивеществе, хотя в одном из случаев ожидалось бы преобладание электронов, чего не наблюдается). Поэтому существование аксионов объясняет подавляющее преобладание во Вселенной вещества над антивеществом [1; 4; 432; 529; 586; 705; 760].
В реакциях микрочастиц (например, при распаде того же К-мезона) набор исходных и конечных свойств участников событий в сумме должен быть идентичным. Если мы наблюдаем своего рода «перекос» в пользу вещества или антивещества, значит некая легкая и малозаметная частица «унесла» с собой качества (энергии и заряды), недостающие реакции для того, чтобы отвечать ожиданиям, соблюдать законы сохранения и выдерживать равноправие вещества и антивещества. Аналогичные «недостачи» энергии в ядерных реакциях привели в 1930–1933 гг. к открытию легкой частицы нейтрино, ответственной за «унос» энергии. В нашем случае с аномалией вещества и антивещества подозрение в искажении ожидаемого хода событий падает на гипотетический аксион. Будущие эксперименты позволят уточнить предположения физиков на этот счет.
За вычетом бозонов Хиггса, которые могут состоять из пары так называемых техникварков, лишь упомянутые частицы по-настоящему элементарны (хотя высказывается предположение, что электрон состоит из нескольких электрино
[242]). Прочие микрообъекты вроде мезонов или нуклонов композитны. В строении материи этот принцип выдерживается универсально: нуклоны с электронами образуют атомы, они, в свою очередь, объединяются в молекулы, составляющие физические тела, иерархия которых завершается небесными светилами. Подобная стратификация природы соответствует классификации фундаментальных сил: субатомный мир подчинен сильному взаимодействию, атомный и молекулярный – электромагнитному, астрофизический – гравитационному. Иными словами, структура макромира не случайна – она задается особенностями микромира. Не составляет исключения и иерархия астрономических объектов, что мы продемонстрируем при разборе многомерности суперструн.
Пространство нашей Вселенной ни в одной своей точке не является подлинно пустым. В соответствии с принципом неопределенности В. Гейзенберга, согласно которому координаты центра инерции физической системы и ее импульс одновременно не могут принимать вполне определенные и точные значения [19; 24, с. 63–64], в любом самом свободном от наполнения веществом и полем уголке Вселенной могут рождаться и исчезать так называемые виртуальные частицы
. Это обстоятельство заставляет считать среду Вселенной по-своему сплошной и расценивать ее как «мировую поверхность
», родственную другим трехмерным средам (воздушной, водной и т. п.).
Гуще или реже мировая поверхность повсеместно покрыта мельчайшими складками, которые именуются суперструнами
. Их размеры отвечают так называемой планковской длине
в 1,62 × 10–33 см (по имени физика М. Планка, 1858–1947). С точки зрения квантовой механики планковская длина является минимальной размерностью, физически разрешенной в нашей Вселенной, и ничего меньше этой длины существовать не может. Суперструны отвечают элементарным частицам микромира.
Со времени своего рождения в Большом взрыве суперструны пребывают в безостановочном движении, являющемся отдачей или отголоском этого Взрыва, что обусловливает философский постулат об абсолютности движения и относительности покоя. Во-первых, суперструны безостановочно колеблются с предельными световыми скоростями. Во-вторых, суперструны непрерывно перемещаются в пространстве: безмассовые, вроде фотонов, – со скоростью света, а обладающие массой покоя – с околосветовыми, релятивистскими скоростями.
Колебательным движениям суперструн присущи характерные особенности, именуемые модами
. По природе все суперструны тождественны, но в зависимости от моды колебания воспринимаются как разные частицы. Их можно рассматривать как элементарное начало всех физических вещей, как первоэлемент. Представляя собой вечно колеблющиеся частицы, ведущие себя и как волны, и как точечные объекты, суперструны разрешают философскую дилемму волны-частицы, давно тревожащую мыслителей. Ее существо состояло в том, что при определенных обстоятельствах элементарные частицы подавали себя как волны: например, плавно огибая препятствия и проникая в область геометрической тени подобно волнам на воде, что называется дифракцией
. В иных случаях (например при столкновении с мишенью) элементарные частицы проявляли себя как крохотные, точечные тела. Эти особенности присущи математической модели суперструн.
Все суперструны открыты, т. е. условно напоминают отрезки трепещущих нитей накаливания, – лишь гравитоны замкнуты в кольцо. Если макромир существует в трех пространственных измерениях (длина – ширина – высота), то мир суперструн пространственно девятимерен. Альтернативная теория супермембран
[299; 313а] видит все суперструны замкнутыми и пространственно десятимерными. Принятие состоятельности данной версии микромира может навести на мысль, что суперструны бывают 10-, 26-, 506-мерными [663] и вообще n-
мерными [447, с. 36].
На наш взгляд, природа данного затруднения скорее мировоззренческая, нежели физическая. Математическая многомерность понимается пространственно (как если бы рядом с нами находились некие вместилища иных измерений), для чего нет экспериментальных оснований. Логичнее считать, что микрообъекты, помимо трех привычных (длина – ширина – высота), имеют шесть дополнительных степеней свободы движения: колеблясь «влево-вправо», суперструна одновременно колеблется не только «вверх-вниз» и «взад-вперед», но еще в шести направлениях, уподобляясь в разрезе профилю противотанкового ежа. Девять подобных направлений минимальны, однако порция энергии может добавить десятое, одиннадцатое и т. д., ничем не подрывая основ теории суперструн. Математическое описание их поведения при этом останется «пространственным», хотя природа будет несколько иной.
На заре существования Вселенной распадающаяся сингулярность оставила следы своей многомерности в устройстве нашего макромира. Сингулярность представляла собой объект наподобие суперструны и колебалась одновременно по меньшей мере в девяти плоскостях. То же справедливо и для нынешних макрообъектов. Человек способен двигаться в трех измерениях и в то же самое время вращается вокруг центра Земли (4-я степень свободы движения). Вместе с Землей он одновременно обращается вокруг Солнца (5-я степень свободы), а вместе с ним – вокруг центра Галактики (6-я степень). Вместе с Галактикой мы ежеминутно перемещаемся вокруг центра Местной Группы (наша Галактика Млечный Путь, Большое и Малое Магеллановы Облака; 7-я степень свободы). В составе Местной Группы всякий человек непрерывно движется относительно центра Галактической Империи (галактики Дракон, Малая Медведица, Лев II, Лев I, Секстант, Большое Магелланово Облако, Малое Магелланово Облако, Млечный Путь, Киль, Скульптор, Печь [268]; 8-я степень свободы). Наконец, в составе Местного Сверхскопления галактик мы вместе со Сверхскоплениями Девы, Павлина-Индейца, Гидры-Кентавра и другими неуклонно стремимся к Великому Аттрактору (9-я степень свободы). Сам того не замечая, каждый человек живет во Вселенной по меньшей мере с 9-ю степенями свободы движения, и такое положение вещей, несомненно, является наследием раннего этапа развития нашего мира. Оттуда же проистекает устройство астрофизического макромира вообще.
В самом деле, чем объяснить то обстоятельство, что Вселенная под влиянием сил гравитации не выглядит как общий бесформенный ком, разбухающий в силу вселенского расширения, а представляет собой виртуозное образование из девяти (или более) центров вращения и притяжения, движущихся относительно друг друга? Почему она выглядит как матрешка взаимовложенных вращающихся сфер, о чем интуитивно догадывался Пифагор (2-я половина VI в. – начало V в. до н.э.) [104, с. 149, № 5]? Астрономическая наука привыкла к подобному положению вещей, общественность – тоже, однако философия требует разъяснения и здесь. Если многоэтажно закрученная вокруг различных центров Вселенная отражает своим обликом древнюю многомерность (девять степеней свободы) первичной сингулярности, тогда ясность наступает.
В свете вышесказанного основной объект теорий суперструн и супермембран видится особым образом. В принципе это может быть точечный отпечаток сингулярности, суперточка
. Родившись как зеркальное отражение распадающейся сингулярности, суперточка пришла в колебательное движение со световой скоростью и тут же приняла облик сплошного протяженного объекта, т. е. суперструны. Далее она развивала релятивистские колебания в 9 или 10 плоскостях, представая как классическая суперструна или супермембрана с 9-ю или 10-ю измерениями, оставаясь в основе одним и тем же образованием, суперточкой, адекватно отвечающей гибкости суперструнных-супермембранных теорий по части многомерности.
Распад сингулярности можно представить в виде расходящейся во все стороны ряби от трепещущей первичной суперточки. Вероятно, истинное число ее степеней свободы составляло примерное количество суперструн нынешней Вселенной, помноженное на 9 или 10 степеней свободы. Сингулярность теряла свои исходные количественно астрономические степени свободы, сбрасывая их в виде 9– или 10-мерных суперструн. Это могло бы выражать действующий во Вселенной закон сохранения суммарного количества степеней свободы суперструн – закон, исходный для законов сохранения квантовых чисел (спина, электрического заряда и других свойств, заданных степенями свободы суперструн).
Передвигаясь по мировой поверхности как по среде с определенной степенью вязкости [552, с. 31–32; 803], суперструны испытывают ее сопротивление. Это происходит следующим образом. Вероятность встречи с короткоживущими виртуальными частицами мала. Однако для суперструн, перемещающихся с релятивистскими скоростями, она заметна. Причем чем выше частота их колебаний, тем она ощутимей. «Цепляясь» за встречные виртуальные частицы, суперструны претерпевают деформации, выражающиеся в уплотнении суперструны, т. е. в сокращении ее объема, падении амплитуды и соответствующем подъеме частоты колебаний.
Поэтому, как следует из теории относительности [120; 377], микрообъекты, ускоряясь на релятивистских скоростях, сокращаются в линейных размерах, прибавляют в массе и тормозятся во времени. Последнее обусловлено тем, что физическое время представляет собой дистанцию между событиями. Подскакивая при ускорении в частоте, суперструны сокращают эту дистанцию между своими колебаниями и, следовательно, замедляют свое время (при скорости света – вообще останавливают). Указанные релятивистские эффекты сродни эффекту Доплера в макромире (т. е. зависимости частоты колебания и длины волны от скорости движения источника звука или света). Масса покоя и масса движения у безмассовых частиц зависят от частоты колебания суперструны, что объясняет физическую природу формулы E = mc
2
: чем энергичнее колебания суперструн, тем значительнее их массы (что подробно разъяснялось выше). У частиц с массой покоя имеется нижний предел частот, ответственных за эту массу. У безмассовых частиц он не фиксирован – поэтому масса «плавает» и не расценивается как масса покоя.
Описанные физические эффекты объясняют конечность скорости света. Чем заметнее ускоряется микрообъект, тем труднее он сокращается в размерах и неохотнее растет в частоте колебаний (вследствие собственного внутреннего сопротивления). На его преодоление уходит все больше энергии разгона, и частица «зависает» на отметке скорости света.
Теория суперструн не может обойтись без понятия мировой поверхности, а оно органично объясняет происхождение виртуальных частиц. Представим себе, например, простыню или что-либо подобное. Допустим, на ней имеются складки. Если потянуть простыню за край, складки разгладятся, но возникнут в другом месте. Они поведут себя аналогично исчезающим и вспыхивающим на мировой поверхности виртуальным частицам (коротко живущим складкам-суперструнам). Разумеется, мировая поверхность – это не двумерная простыня, но принципы ее поведения аналогичны.
Природа основных взаимодействий микромира также вытекает из поведения суперструн. Для гравитации, электромагнетизма и сильного взаимодействия характерны притяжение и отталкивание. Эти эффекты обусловлены волновой природой суперструн-бозонов, которые подобно волне от судна на воде способны – в зависимости от фаз – как притягивать, так и отталкивать партнера по взаимодействию. Деформируя моду колебаний партнеров, частицы передают качества: цветовой заряд (особое свойство кварков, переносимое глюонами), электрический заряд (переносимый фотонами, Z– и W-бозонами), массу покоя (транслируемую хиггсами) и т. д. Обмениваясь частотами колебаний, частицы обмениваются энергией. Влияя на моды колебаний друг друга, частицы регулируют свои взаимопревращения и распады (например при слабых взаимодействиях).
Суперструны ответственны за происхождение таких фундаментальных свойств вещества, как твердость, непроницаемость, устойчивость, которые в античной философии (например у Нигидия Фигула) приписывались заботам «демиурга» – метафизического начала, поддерживающего формы всех вещей. Лопасти вентилятора или пропеллера в покое легко огибаются, однако в стремительном вращении превращаются в сплошной, непроницаемый диск. Колеблющиеся со световой скоростью суперструны так насыщенно заполняют объем с поперечником в планковскую длину, что делают его плотной, непроницаемой частицей, свойства твердости которой в скоплениях частиц передаются физическим телам макромира.
Можно констатировать, что необычайная философская продуктивность физической теории суперструн делает ее незаурядным мировоззренческим достижением.
1.2. Макромир
Вселенная родилась в Большом Взрыве и с тех пор непрерывно расширяется. Скорость этого расширения называется постоянной Хаббла, обозначаемой символом Н
0
. Две трети обратной ей величины (1/H
0) отвечают возрасту Вселенной при условии, что средняя плотность масс во Вселенной делает ее гравитационно замкнутой. При этом средняя плотность масс принимается за единицу (Ω = 1). Если средняя плотность масс меньше единицы (Ω < 1), Вселенная не замкнута и будет расширяться вечно. Если средняя плотность масс равна или больше единицы (Ω > 1), Вселенная замкнута, и ее расширение должно смениться сжатием (коллапсом). Эмпирические оценки средней плотности Вселенной разноречивы: Ω = 0,3–1,3 [255; 270; 609; 832]. Однако средняя плотность Вселенной, в принципе, не может быть «отрицательной», т. е. иметь Ω < 1 (так принято выражаться в астрофизике, хотя «омега» не может быть меньше «нуля» – она всего лишь меньше «единицы»).
Как отмечалось выше (см. разд. 1.1), Вселенная – это сплошная мировая поверхность, которая по квантово-механическим причинам не мыслится «тоньше» планковской длины. Допустим, Вселенная «попробует» расширяться вечно. Тогда, предельно растянувшись в расширении и достигнув «по толщине» планковской размерности, Вселенная непременно прекратит рост и под влиянием собственной внутренней гравитации обязательно начнет сжиматься. Поэтому ее вероятная средняя плотность больше или равна единице (Ω ≥ 1).
Между тем реальное положение вещей намного сложнее. Теоретическая оценка «омеги» – в том случае, когда она больше или равна единице, приложима лишь к изотропной модели Вселенной, т. е. ко Вселенной, повсеместно однородной. Однако современные астрофизические исследования недвусмысленно показывают, что Вселенная анизотропна, т. е. глубоко неоднородна. Она имеет центр и периферию (о чем речь пойдет ниже). В неоднородной же (анизотропной) Вселенной средняя плотность вещества, по-видимому, составляет приблизительно ΩUniv = ~ 0,245 (см. приложение 7). И все-таки данное обстоятельство не делает Вселенную открытой (вечно расширяющейся), поскольку в ее центральных областях местная плотность существенно превосходит критическую (т. е. Ωloc > 1). Это превращает Вселенную в образование, склонное к поэтапному схлопыванию на манер «гармошки», и пока периферические области Вселенной продолжают вяло расширяться, центральные области уже властно сжимаются и шаг за шагом втягивают в себя беспомощную периферию. Подобная современная картина мира куда сложнее традиционной, однако лучше отвечает новым фактам (см. далее).
Наблюдательные астрономические оценки постоянной Хаббла колеблются в пределах 39–105 ±11 км/с на мегапарсек (мегапарсек равен 3,26 млн световых лет) [151; 174; 186; 243; 269; 331; 336; 341; 355; 404; 420; 428; 435; 527; 531; 624; 748; 792]. Для определения возраста Вселенной такие данные непригодны, поскольку дают колоссальный разброс величин и не учитывают неоднородность Вселенной (споры по поводу оценки постоянной Хаббла даже получили хлесткое наименование «войны Хаббловой константы» [269] – участники ее были беспощадны друг к другу). Имеется, правда, и другая теоретическая возможность.
Представления о расширяющейся Вселенной, родившейся в Большом Взрыве, носят наименование моделей «горячей Вселенной». При расчетах моделей горячей Вселенной используется теоретическая величина постоянной Хаббла H
0 = 50 км/с на мегапарсек [611; 673; 705], которой отвечает возраст Вселенной в 13,(3) млрд лет [336]. Эта оценка оптимальна, поскольку независимые от нее определения возраста нашего мира по старым белым карликам в Галактике [550; 792], по тяжелым радиоактивным элементам на звездах [233; 409; 717] и по шаровым скоплениям звезд [235; 404; 475; 792] предусматривают вселенский возраст от 10,3 до13 млрд лет. Поясним, что белые карлики – это крохотные, сверхплотные звезды класса светимости VII, некоторые из которых вдвое старше нашего Солнца. Добавим, что тяжелые радиоактивные элементы имеют тенденцию распадаться. По их остаткам можно определить возраст древних звезд, которые также вдвое старше Солнца. Напомним также, что шаровые скопления – это гало, своего рода шаровидные облака звезд вокруг галактик, и эти звезды населения II также вдвое старше Солнца, т. е. звезды населения I, распространенного не в гало, а в дисках галактик.
Как можно видеть, возраст Вселенной по белым карликам, тяжелым элементам и шаровым скоплениям (13 млрд лет) на 300 млн лет уступает возрасту Вселенной по теоретической постоянной Хаббла (13,(3) млрд лет). Это лишний раз свидетельствует в пользу оптимальности последней величины, ибо 300 млн лет как раз потребовались юной Вселенной для того, чтобы сформировать древнейшие галактики и звезды с будущими белыми карликами и шаровыми скоплениями. Ниже возраст первых сверхскоплений галактик оценен в 200 млн лет после Большого Взрыва, однако они древнее первых звезд, а потому результаты вполне удовлетворительны. Дело в том, что ранние сверхскопления будущих галактик поначалу формировались как крупномасштабные возмущения вселенского вещества ок. 200 млн лет после Большого Взрыва, и лишь затем, ок. 300 млн лет после Большого Взрыва, в них сложились конкретные галактики с древнейшими звездами.
На наш взгляд, существует еще один путь расчета возраста Вселенной. Со времен Большого Взрыва ее внешние границы расширялись со скоростью света. Поделив скорость света на местную скорость вселенского расширения и взяв 2/3 от результата, мы получим нынешний радиус Вселенной в мегапарсеках (и световых годах), а значит и время, затраченное на достижение этого радиуса. При H
0 = 50 км/с на мегапарсек радиус Вселенной составляет 13,031 млрд светолет и отличается от предыдущей оценки на 302,4 млн световых лет. Это расхождение означает, что Земля расположена к краю Вселенной на 302,4 млн светолет ближе, нежели ее центр, который отстоит от нас на ту же дистанцию.
Поиск такого центра отталкивается от наблюдения, что Южное полушарие звездного неба ощутимо глубже Северного [546; 695]. Это означает, что на Севере звездной сферы к нам ближе край Вселенной, а на Юге – ее центр. В Южном полушарии звездного неба по направлению к сверхскоплению галактик Гидра-Кентавр на расстоянии ок. 300 млн светолет от нас лежит Великий Аттрактор. Этот объект соответствует 5,4 × 1016 солнечных масс, имеет поперечник в 0,6–6 млн светолет и вдвое превосходит среднюю плотность Вселенной [524; 134; 296; 372, с. 62; 473; 704; 785; 830]; cp. [254].
Сверхмассивный Великий Аттрактор притягивает окружающие скопления галактик, в том числе наше Местное Сверхскопление, которое вместе с соседними сверхскоплениями стремится к Великому Аттрактору («Великому Притяжателю») в составе сверхпотока Персей – Рыбы протяженностью в 423,8 млн светолет [808; 548; 514; 605]. Это означает, что гравитоны Великого Аттрактора успели охватить область Вселенной с диаметром ок. 1 млрд светолет, а стягивающая Вселенную деятельность Великого Аттрактора началась ок. 500 млн лет назад.
Великий Аттрактор отождествляется с рентгеновским галактическим скоплением Абелль 3627, обладающим массой в 5,1 × 1015 солнечных масс и поперечником в 1,956 млн светолет, укладывающимся в поперечник Великого Аттрактора. Абелль 3627 является источником самого яркого рентгеновского потока среди всех известных скоплений (0,1–2,4 кэВ). Его отделяют от нас 303,18 млн светолет [473; 830]. Сравнивая наше расстояние от центра Вселенной (302,375 млн светолет) с расстояниями от Великого Аттрактора (ок. 300 млн светолет) и Абелля 3627 (303,18 млн светолет), мы заключаем, что речь идет об одном и том же образовании, три ипостаси которого выявлены совершенно различными, независимыми методами.
Абелль 3627 в 10 раз уступает расчетной массе Великого Аттрактора, которая определялась гравиметрически (по силе тяжести, т. е. однозначно), в то время как масса Абелля 3627 оценивалась оптически (по внешнему виду, т. е. далеко не однозначно). Расхождение объясняется тем, что 0,9 реальной массы Абелля 3627 состоят из невидимого «темного вещества».
«Холодное темное вещество» не испускает ни тепла, ни света и взаимодействует с обычным «горячим светлым веществом» лишь гравитационно, сильно влияя на движение галактик и их скоплений, но оставаясь невидимым во всех областях спектра. Оно было открыто благодаря одному астрофизическому парадоксу. Наша и другие галактики вращаются столь интенсивно, что центробежная сила могла бы их разметать. Видимой «светлой» массы им совершенно недостаточно, чтобы сцементировать их силой тяжести. Значит, существует некая невидимая «темная» масса, которая восполняет недостачу вещества, необходимого для консолидации галактик силой тяжести. Гравитационные оценки свидетельствуют, что по массе «темное вещество» образует не менее 90% состава Вселенной [333; 431; 581; 617; 629; 664; 739; 760; 783; 785; 795]; cp. [176]. Состав «темного вещества» – проблематичен.
Барионное (кваркосодержащее) «темное вещество» заключено в тусклых холодных белых карликах – это 2% от «темного вещества» галактического гало [604], – а также в МАСНО (массивных, компактных объектах гало планетарных масс), выявляемых методом микролинзирования: в тот момент когда они затмевают внегалактические звезды (галактик Большое Магелланово Облако и М22), те ненадолго ярко вспыхивают, становясь видимыми по обе стороны от затмевающего МАСНО, привлекая наше внимание (МАСНО гравитационно стягивают лучи света от них, выполняя роль маленькой гравитационной линзы, откуда происходит название метода микролинзирования) [133; 376; 665; 666]; cp. [224]. Можно упомянуть еще гипотетические космические струны
– сверхмассивные, протяженные объекты замкнутой, петлистой фактуры, сохранившиеся от первой секунды жизни Вселенной, когда они застыли как пограничные состояния ее фазовых переходов (наподобие прожилок на замерзшем стекле, только длиной в световые годы) [22; 132; 167].
В небарионном (бескварковом) «темном веществе» предполагаются аксионы (см. разд. 1.1) [332; 586; 705] и вимпзиллы
[360; 376]. Вимпы
представляют собой слабо взаимодействующие массивные частицы в 106 ГэВ (гигаэлектронвольт), а вимпзиллы (т. е. вимпы-годзиллы) – это слабо взаимодействующие сверхмассивные частицы в 1012 ГэВ, которые еще предстоит открыть. Сюда же относятся гравитино (см. разд. 1.1) [332; 371, с. 39] и хиггсино (см. разд. 1.1) [371, с. 39; 706]. Называют также нейтралино
(суперпартнер фотона со спином 1/2, т. е. аналог фотино, в 10–1000 масс протона) [760] и само фотино в 100 масс протона (см. разд. 1.1) [332; 371, с. 39; 705; 706; 725]. Наконец, популярны снейтрино [371, с. 39], таунейтрино с массой в 17–17,2 кэВ или 15–10000 ГэВ [303; 343; 376; 513; 528; 530; 673; 706; 739] и электронное нейтрино с массой от менее 7 эВ до 30 эВ [740; 760] (см. разд. 1.1).
Перечисленные микрообъекты занесены в состав «темного вещества» как бы наудачу, в ожидании наблюдательных открытий [725]. Между тем обобщенный подход имеется в теории суперструн, где «темному веществу» соответствует «теневое вещество» Е
8' из калибровочной группы Е
8 × Е
8', в которой группа Е
8 описывает мир обычного, «светлого вещества» [9, с. 520; 122, с. 583; 472]. Напомним, что калибровочные группы служат математической основой для принятия «к производству» не произвольного набора объектов и их свойств, а их некой математически обоснованной матрицы вроде таблицы умножения. Поскольку вездесущим фоном нашего мира «светлого вещества» является известное реликтовое микроволновое излучение, наполняющее космос и состоящее из фотонов с температурой в 2,726 ± 0,01 К [705; 760], то фоном невидимого мира «темного вещества» могло бы оказаться фотино, и без того присутствующее среди кандидатов на эту грозную роль.
Причину сокрытости от нас «холодного темного вещества» можно понять. Допустим, оно действительно состоит из суперчастиц (в том числе фотино), для которых единодушно предполагается значительная индивидуальная масса. Тогда сопоставимое с числом «светлых» частиц количество «темных» суперчастиц должно быть в несколько раз массивнее, что объясняет тот факт, что оценочная масса «темного вещества» достигает 90–99% массы всей Вселенной, в то время как на долю «светлого вещества» приходится 1–10%.
Следует также отметить, что, помимо гравитонов, «темное вещество» не испускает иных излучений, в том числе электромагнитных, световых, откуда происходит его мрачное название. Причина может состоять в следующем. Суперчастицы очень массивны, и радиусы их взаимодействий крайне малы (см. разд. 1.1). В силу этого обстоятельства суперчастицы не в состоянии взаимодействовать со «светлым» веществом, образовывать с ним комбинации, склонные излучать фотоны (как свойственно всякому «светлому веществу»), обнаруживать себя оптически или же путем магнетизма, т. е. слабых и сильных взаимодействий. Поэтому, даже находясь рядом с нами, «темное вещество» останется неощутимым.
Таким образом, версия «темного вещества», состоящего из суперчастиц, представляется наиболее простой и убедительной, поскольку органично встраивается в современную научную картину физического мира. Другое дело, что природа «темного вещества» не облегчает задачу его эмпирического обнаружения, так как экспериментальное проникновение в мир суперчастиц с малыми радиусами взаимодействий требует больших энергий.
Зная отношение вселенской массы «светлого вещества» к массе «темного» (ок. 0,1–0,01), мы в состоянии предположить порядок масс отдельных суперчастиц (см. рис. 1). В принципе, их массы могут колебаться от 0,7–7,0 эВ у сэлектронного снейтрино до 1780–17800 ГэВ у истинного скварка. По-видимому, подобные объекты не в силах образовывать привычные нам формы вещества (нуклоны, атомы, молекулы), так как требуют очень высоких энергий для своего сближения, а в современной холодной Вселенной таких энергий нет. Если же они появятся, мощи сильного взаимодействия не хватит, чтобы удерживать суперчастицы в ансамблях, вследствие чего они и не сложились на заре горячей Вселенной. Из сказанного следует, что мир «темного вещества» чужд миру «светлого» в силу отсутствия организации, отчего «темный мир» представляется этаким холодным древним хаосом, о присутствии которого во Вселенной догадывались уже первобытные охотники.
Разделение громадного «темного» и крохотного «светлого» миров началось 13,(3) млрд лет назад, когда чередование событий измерялось мельчайшими планковскими квантами времени, меньше которых не существует единиц измерения времени по квантово-механическим причинам, – это 5,4 × 10–44 с. На заре нашего мира Вселенная пребывала в форме сингулярности (см. разд. 1.3). Затем она распалась на суперточки (см. разд. 1.1), которые превратились в трепещущие суперструны, что явилось первым проявлением термодинамики во Вселенной. Термодинамика – это наука о динамических состояниях макроскопических систем, и на заре времен начало им было положено хаотичным биением юных суперструн, рождающих закон неубывания энтропии (см. разд. 1. 4).
Отталкиваясь друг от друга, суперструны начали расходиться во все стороны, а новорожденная Вселенная стала расширяться и, соответственно, остывать. До момента 10–43 с после Большого Взрыва четыре основных физических взаимодействия (гравитационное, слабое, электромагнитное, сильное) проявлялись на одинаковых дистанциях и были неразличимы по эффектам (т. е. пребывали в состоянии Великого Объединения). Затем рост дистанций между суперструнами обособил гравитацию. По той же причине в момент 10–35 с после Большого Взрыва отделилось сильное взаимодействие, и Вселенная перестала склеиваться глюонами, переносчиками самого мощного – сильного – взаимодействия. В результате Вселенная взрывообразно вздулась. Эта стадия раздувания Вселенной (инфляция) заняла 10–35/10–34 – 10–32 с после Большого Взрыва, когда Вселенная выросла в 1050 раз [34; 67; 68; 373; 388; 389; 390; 512; 532]; cp. [448]. (Поясним при этом, что разделение фундаментальных физических взаимодействий по мере расширения и остывания Вселенной происходило потому, что для каждого из них характерны определенные дистанции проявления: в крохотной Вселенной они неразличимы, а в расширившейся заявляют о себе во весь голос.)
В эпоху 10–32 – 10–6 с после Большого Взрыва Вселенную наполнял кварк-электронный бульон. Через 1 с после Большого Взрыва состоялось разделение электромагнитного и слабого взаимодействий, и началось образование протонов. Спустя 100 тыс. лет после Большого Взрыва Вселенная все еще выглядела как смесь вещества и излучения (фермионов и бозонов). С возникновением нейтронов появились атомы (гелия, дейтерия, лития – 0,5 млн лет после Большого Взрыва [547]). Наступило разделение вещества и излучения – Вселенная стала прозрачной, как в настоящий момент. Из затравок, обусловленных колебаниями плотности вещества, зародились протосверхскопления, которые через 200 млн лет после Большого Взрыва [547] превратились в сверхскопления галактик. Разделенные колоссальными пустотами, они стали каркасом ячеистой крупномасштабной структуры Вселенной [41]. Обособление «темного» вещества от «светлого» произошло еще в ходе раздувания Вселенной, когда щупальца взаимодействий тяжелых суперчастиц перестали доставать до юрких, легких элементов «светлого» вещества.
Схема нынешней Вселенной видится следующим образом (рис. 2). В ее центре висит Великий Аттрактор (Абелль 3627), окруженный шестилучевым надсверхскоплением галактик, часть которого является агломератом Рыб – Кита (куда входят сверхскопления галактик Волосы Вероники, Гидра – Кентавр, Персей – Рыбы, Рыбы – Кит; поперечник этого образования достигает 1,2096 млрд светолет, масса 1017 – 1018 солнечных масс [759; 783]). Другая часть надсверхскопления связана с волокном Персей – Пегас протяженностью 912,8 млн светолет [177; 759]. Во все стороны от агломерата Рыб – Кита разбегаются пустоты того же поперечника, по десятку в каждом направлении. В их стыки вкраплены надсверхскопления галактик вроде нашего агломерата Рыб – Кита, только сильнее разреженные и менее массивные. Речь, разумеется, идет об идеализированной схеме – в реальности она причудливо искажена, разнообразится пустотами меньшего масштаба [411] и т. д.
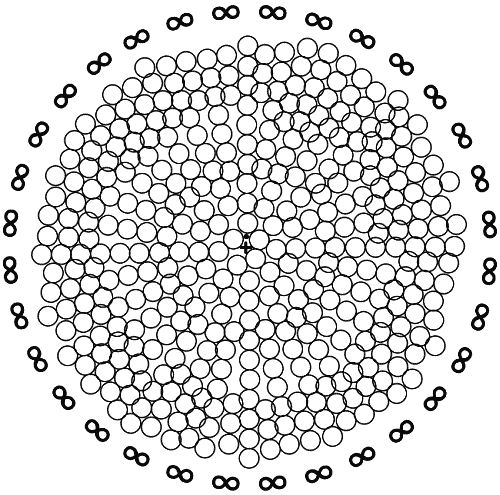
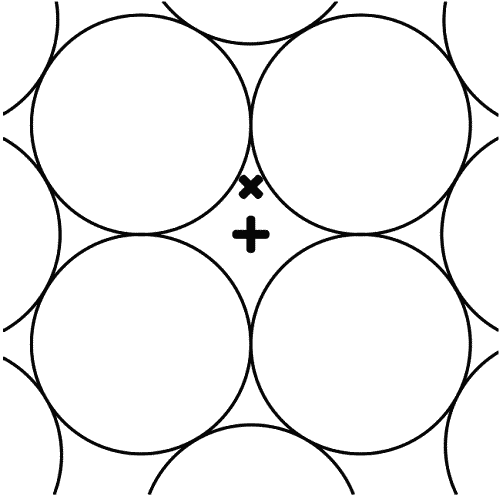
Рис. 2.
Принципиальная схема строения анизотропной Вселенной, имеющей расширяющуюся периферию и центр, совпадающий с древним эпицентром Большого Взрыва. Условно обозначены: + центр Вселенной, отвечающий Великому Аттрактору, или скоплению рентгеновских галактик Абелль 3627; его окружает ромбическая область агломерата Рыб – Кита, где находится × Галактика Млечный Путь с нашей Солнечной системой; поперечник агломерата Рыб – Кита достигает 1,2 млрд световых лет; О – пустоты, лишенные вещества, из которых в основном состоит Вселенная; вещество имеется лишь на границах пустот и в ромбических пазах между ними; поперечник одной пустой полости в среднем составляет 1,2 млрд светолет, что предполагает радиус Вселенной в 13,(3) млрд светолет; × Галактика Млечный Путь лежит на 303 млн светолет к «северу» от Абелля 3627; ∞ – окружающий Вселенную гипотетический Вечно Изменяющийся Мир, породивший ее 13,(3) млрд лет назад
Идея анизотропной Вселенной, располагающей конкретным центром, выглядит революционной лишь на первый взгляд. В действительности никакой другой Вселенная быть и не могла. В самом деле, общепризнанная космогония исходит из того, что Вселенная родилась в Большом Взрыве. Но всякий взрыв располагает эпицентром. Несомненно, он имелся и у Большого Взрыва. В таком случае расширение Вселенной происходило относительно этого эпицентра, который и является полноценным центром Вселенной. В сущности, мы всего лишь конкретизировали его местонахождение, отождествив таковое с небесными координатами Великого Аттрактора.
Представленная выше картина макромира позволяет единообразно разрешить самые острые парадоксы новейшей астрофизики. К ним относятся нарушения закона Хаббла удаленными квазарами и сверхновыми звездами, откуда в конечном счете следует, что либо Большого Взрыва никогда не было, а следовательно нет и закона Хаббла, либо Вселенная в десять раз древнее, чем полагают, и тогда львиная доля космогонии (астрофизической науки о происхождении Вселенной) подлежит забвению… Попробуем детально разобраться с этими бедами.
Закон Хаббла подразумевает, что в расширяющейся Вселенной скорость удаления галактик, например от нашего Млечного Пути, пропорциональна расстоянию этих галактик от нас. Так происходит не потому, что Млечный Путь занимает исключительное место во Вселенной, а потому, что Вселенная повсеместно как бы раздвигается. Из-за этого одновременно растут расстояния между всеми галактиками, эти расстояния в наших глазах складываются, и нам закономерно кажется, что ближайшая галактика удаляется со скоростью Х, вторая по удаленности – со скоростью 2Х, третья – 3Х и т. д. Изящество и простота закона Хаббла покорила астрофизическую науку.
Связанная с ним сложность всегда состояла в том, что никак не удавалось однозначно определить абсолютную скорость, с которой расширяется Вселенная. Мы точно знаем, какая из галактик удаляется от нас быстрее, но с какой именно скоростью – не знаем. Это происходит потому, что астрономам нелегко определить точные расстояния до других галактик. В результате всякое очередное определение какого-либо из этих расстояний тут же приводит к оглашению новой величины для скорости расширения Вселенной, т. е. для постоянной Хаббла, о разбросе величин которой говорилось выше.
Можно задаться вопросом: имеет ли это обстоятельство мировоззренческое значение? Ответ состоит в том, что, не зная постоянной Хаббла, мы не узнаем возраста Вселенной. Не зная возраста Вселенной, мы не поймем, как она строилась. Не понимая, как она строилась, мы не поймем, что ее ждет. Для космической цивилизации, в которую постепенно превращается цивилизация человеческая, такое положение вещей неприемлемо. Кроме того, нам интересно было бы узнать, как устроен небесный мир…
Методика расчета центра Вселенной (см. выше) привязывает постоянную Хаббла к дистанции от этого центра. В радиусе до 0,6 млрд светолет от вселенского центра Хабблова постоянная убывает пропорционально кубу расстояния от 105 км/с на мегапарсек (в 33 млн свето-лет от центра Вселенной) до 39,8 км/с на мегапарсек. Это укладывается в разброс постоянной Хаббла 39–105 км/с на мегапарсек, полученный для измеряемой части Вселенной, которая оценивается в 0,5 млрд све-толет [605; 657; 830]. С удалением в 5 млрд светолет от центра Вселенной H
0 убывает до 19,6, т. е. наполовину против своих минимальных эмпирических величин, и начинает нарушать закон Хаббла, требующий, чтобы с удалением постоянная Хаббла росла, а не падала. Однако наши теоретические выводы (см. приложение 7) согласуются с фактическими данными.
Для исследования космических глубин Вселенной используются своего рода космические маяки: сверхновые звезды и квазары (квазизвездные объекты). Сверхновая звезда – это колоссальная звездная вспышка, когда звезда выгорела, перестала разбухать от излучения и температуры, стремительно сжалась и катастрофически взорвалась от скачка гравитационной энергии или от термоядерной реакции в своем вырожденном углеродном ядре (исследования в этой области еще продолжаются). Максимальный блеск взрыва у сверхновых звезд типа I удерживается около недели, а у сверхновых типа II – около 20 дней. Квазар же – это ядро чрезвычайно активной галактики с «черной дырой» (сколлапсировавшей звездой) в центре (исследования продолжаются). Использующие гравитационную энергию своих «черных сердец» квазары исключительно ярки.
Зона распространения квазаров лежит в 0,25–12 млрд световых лет от нас. Поэтому половина квазаров находится в области нарушения закона Хаббла согласно нашим представлениям. Удаляющиеся от нас объекты имеют искажения в своих спектрах, называемые красным смещением: чем выше скорость удаления, тем больше красное смещение. Статистический анализ красных смещений квазаров показывает, что они наполовину не подчиняются закону Хаббла [591; 592]. Удаленные сверхновые звезды типа Ia, чей возраст превышает древность Земли (4,51 млрд лет), также нарушают закон Хаббла [615; 645]. Проще говоря, удаленные квазары и сверхновые звезды улетают от нас гораздо медленнее, чем ожидалось. При этом их удаленность говорит о древнем возрасте, поскольку свет от них идет к нам в среднем около пяти млрд лет.
Это открытие привело исследователей к прямолинейной мысли о том, что в древности скорость расширения Вселенной была заметно ниже нынешней (т. е. свойственной окрестностям нашей Галактики), а теперь она увеличилась под влиянием некой «отталкивающей силы» [356–359; 516; 610], обусловленной загадочной «темной энергией» [200; 302; 474; 475; 617; 789]; cp. [837]. Сторонники этой концепции экстраполируют торможение в расширении Вселенной в глубь времен и автоматически приходят к выводу, что Вселенная старше 150 млрд лет.
Подобный вывод неприемлем, поскольку за такой огромный срок сменяющие друг друга поколения звезд могли успеть десятикратно переработать все исходное вселенское вещество на тяжелые элементы, так что мы жили бы в мире без водорода, гелия и прочих легких элементов, чего не наблюдается. Кроме того, с физической точки зрения представляется противоестественной картина, когда продукты Большого Взрыва поначалу разлетаются вяло, а потом ни с того ни с сего в нарушение законов сохранения набирают скорость. Физическая наука подобных взрывов не знает. По-видимому, концепция «отталкивающей силы» и «темной энергии» не продумана.
На наш взгляд, падение постоянной Хаббла на периферии Вселенной носит не временной, а пространственный характер. Поскольку объем Вселенной с удалением от вселенского центра растет как куб расстояния, разбегание ее вещества должно тормозиться в той же прогрессии. На самом краю Вселенной постоянная Хаббла опускается до 14,2 км/с на мегапарсек безо всякого влияния какой-либо дополнительной энергии, в том числе и «темной». По той же причине местная плотность вещества там (Ωloc = ~ 0,226) сокращается не потому, что Вселенная «открыта», а в силу того, что она подчиняется элементарным требованиям стереометрии. Эти обстоятельства выявляются именно в анизотропной модели Вселенной (с центром и периферией). Изотропная (однородная) модель таких возможностей не дает.
Завершая обсуждение анизотропной модели Вселенной, мы вынуждены добавить несколько «апокалиптических» штрихов. Если агломерат Рыб – Кита уже целиком захвачен гравитонами Великого Аттрактора (а это вероятно), то процесс стяжения агломерата к центру Вселенной должен был начаться еще 605 млн лет назад, когда наш мир достиг возраста 12,73 млрд лет. В то время скорость его расширения на периферии (14,4 км/с на мегапарсек) упала впятеро против центральных областей и перестала нейтрализовать их стягивающее гравитационное влияние. В результате начался Большой Коллапс Вселенной. (Заметим в скобках, что это обстоятельство отличает ее от испаряющихся «черных дыр» [79, с. 291–296], с которыми Вселенная сравнивается в теории «пространственно-временной пены», см. гл. 1.3.) Ожидаемое время жизни Вселенной в общей сложности займет 25,5 млрд лет, свыше половины которых пройдено (см. приложение 7). Таким образом, известному нам миру осталось каких-то 12,124 млрд лет существования…
1.3. Сингулярность
Породившая Вселенную космологическая сингулярность,
имея размерность порядка планковской длины, характеризовалась планковской энергией εPl
= 4,64 × 10114 эрг/см3 и кривизной пространства-времени |R
iklm
R
iklm
| ~ =
1,47 × 10131 см-4. Обладая плотностью не меньше планковской ρPl
= 5,16 × 1093 г/см3, этот объект развивал настолько мощное поле тяготения, что останавливал для себя течение времени. С ним ничего не происходило. Поэтому следует считать, что момент его рождения совпадал с моментом его гибели (т. е. с распадом в Большом Взрыве; см. разд. 1.7).
Конечно, всякому пытливому уму мало знать сухие цифры, характеризующие объект, породивший Вселенную. Как же выглядела сингулярность? Ответ на этот вопрос дать крайне трудно. Не имея биографии во времени (поскольку оно стояло), сингулярность провела свой моментальный век, чуть обозначившись. Со стереометрической точки зрения ее форма едва ли могла развиться дальше элементарной точки, о которой нечего сказать. Кроме того, она была невидима – это совершенно несомненно, и вот почему.
Сингулярность напоминала «черную дыру», звезду, провалившуюся внутрь себя за гравитационный радиус, чрезвычайно уплотнившуюся и не выпускающую из себя даже света (что делает ее невидимой). Эти вероятные черты сингулярности привели к идее так называемой пространственно-временной пены [405; 140; 373, с. 47], согласно которой мироздание представляет собой скопище «черных дыр», то вспухающих, как наша Вселенная в прошлом, то опадающих, как она же в будущем. Недостаток этой идеи состоит в том, что она полагает для необозримого мироздания частные особенности нашего мира, что не выдерживает критики. Не исключено, что ближайшие к нашей Вселенной области Мироздания и впрямь наполнены «пространственно-временной пеной», однако остается вопрос: что находится за ее пределами? Подобная философская сторона проблемы выходит за рамки физики, но не является неразрешимой (см. разд. 1.7).
Самая поразительная особенность сингулярности заключалась не в ее физических параметрах, которые вполне осознаваемы. Труднее вообразить себе акаузальность (независимость от причинности), несомненно присущую сингулярности. Поскольку время для сингулярности стояло, события не происходили, а наши физические законы не выполнялись, то и не было никаких условий для действия закона причинности. Для его осуществления требуются течение событий и принципы сохранения. В сингулярности последние господствовали, однако первое отсутствовало. Здесь сразу же возникает новый парадокс. Господство принципов сохранения не должно было допустить распада сингулярности по соображениям каузальности (причинности). Однако парадокс состоял в том, что в сингулярности причинно-следственный закон не действовал, и потому она беспрепятственно распалась.
Может показаться, что рассуждения о причинности лишь запутывают проблему происхождения сингулярности вместо того, чтобы ее прояснить. Однако далее (см. разд. 1.7) мы увидим, что все обстоит как раз наоборот, хотя отнюдь и не просто.
1.4. Энтропия
Сингулярность являлась предельно упорядоченным объектом. Данное утверждение требует отдельного комментария. Упорядоченность – это повторяемость в состоянии объектов (в их размещении, динамике, симметрии и т. д.). Иными словами, упорядоченность предполагает множественность объектов. Сингулярность же (от лат. singularis – «одиночный») по самому своему понятию мыслится как «единичный» объект. В данном конкретном случае о его упорядоченности приходится говорить на том основании, что сингулярность послужила архетипом (образцом) для сонма суперструн, образующих Вселенную. В них она повторяется. Во всяком случае, о беспорядочности сингулярности вообще не приходится говорить.
Следовательно, распавшись и превратившись в расширяющуюся Вселенную, предельно упорядоченная сингулярность перешла в менее упорядоченное состояние. Отсюда следует, что ее энтропия (проще говоря, хаотичность) как замкнутой системы возросла. Это стало исходным воплощением Второго начала термодинамики в нашей Вселенной, которое означает [15], что всякая энергия в природе стремится перейти из более организованного состояния в менее организованное (тепловое, хаотичное) и что энтропия любой замкнутой системы со временем увеличивается (вернее, не убывает).
Вселенная представляется замкнутой системой (см. разд. 1.2). Разойдясь на бульон из суперструн, былая сингулярность избавилась от своей сверхплотности и от соответствующих ей релятивистских ограничений на течение событий и времени. Вселенная зажила во времени, которое применительно к столь высокому уровню явлений определяется как мера энтропии; ср. [146, с 211; 263]. Данная дефиниция согласуется с предшествующей (время – дистанция между событиями; см. разд. 1.1), поскольку вселенское время разворачивалось по мере расширения Вселенной и роста в ней общей энтропии. Однако расширение Вселенной – это и есть расширение дистанции между вселенскими событиями, в соответствии с чем расширялись и масштабы времени – в полном согласии с дефиницией для квантового мира (см. разд. 1.1), где сжатие суперструн от больших скоростей сопровождается сокращением амплитуд их колебаний и приличествующим сокращением масштабов времени, т. е. его торможением. Проще говоря, элементарная частица сокращается – время тормозится, Вселенная расширяется – время ускоряется. Мы видим согласованную картину на уровнях микро– и макромира.
Таким образом, сразу после Большого Взрыва в младенческой Вселенной самопроизвольно зародилось одно из двух условий действия причинно-следственного закона: течение событий во времени.
1.5. Закон сохранения
Элементарные частицы (суперточки-суперструны), возникшие как копии точечной сингулярности, до сих пор пребывают в движении с релятивистскими скоростями. Это означает, что собственное время для каждой из них стоит (см. разд. 1.1). По этой причине суперструны сохраняют характерные энергии, массы покоя, барионные числа (это указание на принадлежность или непринадлежность частицы к миру кварков-барионов), электрические и цветовые заряды (последние позволяют кваркам избегать квантовых ограничений на кучность), спины (собственные угловые моменты количества движения) и другие фундаментальные качества. Следует иметь в виду, что при этом речь не идет о неотторжимости свойства от частицы, а о неуничтожимости свойства при обмене им частицами, что объясняется их релятивистским сном во времени.
В итоге мы приходим к существенному выводу, что законы сохранения действуют во Вселенной не по метафизическим (трансцендентным, потусторонним), а по физическим причинам: в силу того, что фундаментальные кирпичики мира, снующие по Вселенной с релятивистскими скоростями, попросту заморожены во времени и сохраняют свои качества потому, что вообще не меняются, пока не испытают воздействие извне. Более крупные материальные объекты (атомы, молекулы, физические тела) перенимают начала сохранения из микромира.
Здесь мы находим второе условие действия причинно-следственного закона, нуждающегося, помимо течения событий во времени, в законах сохранения на всех уровнях бытия.
1.6. Причинно-следственный закон
В первые мгновения жизни Вселенной сочетание законов сохранения с энтропийным течением событий ввело в наш мир каузальность. На ее крыльях по Вселенной заскользили многочисленные законы «второго эшелона», число которых велико, но не бесконечно. Вселенная вообще ограничена: число барионов (стабильных протонов) в ней исчерпывается 1078 [34, с. 61]. Ранняя Вселенная породила всего 4 или 5 базовых физических взаимодействий: гравитационное, сильное, электромагнитное, слабое и милли-слабое-сверхслабое (аксионы; см. разд. 1.1). Становление этих сил заняло первую секунду жизни Вселенной. Подобная скудость удивляет, поскольку степени свободы суперструн могли бы создать и более широкий спектр сил.
Запрет на их разнообразие был наложен крепнущим причинно-следственным законом, который благоприятствовал повторяемости явлений и сущностей и противодействовал их новациям. Устойчиво повторяющиеся связи явлений заблокировали расширение списка физических взаимодействий сверх 4 или 5 сил, успевших «пробиться» в первую секунду бытия. То же обстоятельство объясняет однообразную природу существующих физических взаимодействий, которые ограничиваются притяжением-отталкиванием и передачей фиксированного набора свойств, подчиняющихся законам сохранения (см. разд. 1.5). Добавим, что в первые 10–43 с жизни Вселенной все силы были неразличимы в рамках Великого Объединения, т. е. исходное разнообразие мира выглядело совсем скромно. Мы видим, что относительная бедность и значительное однообразие физического бытия не являются какой-то случайностью природы – напротив, эти универсальные особенности кропотливо «выкованы» причинно-следственным законом на заре нашего мира.
Словно задыхаясь в тисках закона причин и следствий, Вселенная создала над суперструнами надстроечный мир из нуклонов, атомов, молекул и физических тел, подчиненных законам классической механики и химии. Причинно-следственный закон возобладал и здесь. Поэтому число оригинальных фундаментальных зависимостей макромира тоже ограничено. Если говорить об обобщениях высшего порядка, то в химии можно назвать периодический закон Менделеева, учитывающий всего 94 заметно стабильных элемента (от водорода до плутония). В астрофизике закон всемирного тяготения обусловливает формирование звезд, звездных систем, галактик и их скоплений. В математике мы имеем Грандиозную теорему конечных простых групп [30; 31], обнимающую все мыслимые множества вещей в природе. Планетарная геология земного типа описывается глобальной тектоникой плит (см. разд. 2.2). История жизни подчиняется дарвиновской теории естественного отбора, а эволюция не чужда направленности (см. разд. 2.3). Развитие материальной и духовной культуры людей также подчинено единому началу (см. гл. 3).
Мы вправе сделать важный вывод относительно происхождения уровней организации материи. В самом деле, почему материальный мир не ограничился уровнем микромира? Физике известны механизмы, позволяющие суперструнам формировать последовательные уровни организации макромира. Но что заставляет их работать? Вопрос этот имеет философский характер и требует ответа.
Из всего вышесказанного следует одна вероятная версия. Мы привели доводы в пользу того, что причинно-следственный закон в силу самой своей природы довольно негативно влиял на разнообразие Вселенной с первых мгновений ее существования. Поэтому основы микромира крайне ограничены в наборе. Стихийно движущиеся суперструны «искали» разнообразия за пределами своего уровня и последовательно выстраивали атомный, молекулярный и макроуровневые этажи. Это происходило потому, что на каждом этаже причинно-следственный закон создавал «драконовские» ограничения на разнообразие, и суперструнам приходилось взбираться все выше и выше по уступам бытия. Это обстоятельство объясняет, почему наш мир вообще подвержен тенденции к прогрессивному развитию вверх, при том, что вширь он жестко ограничен. Мы имеем в виду тот факт, что, вопреки казалось бы естественному порядку вещей, окружающее нас бытие не бесконечно разнообразно, а совсем наоборот. В итоге мы можем видеть, что закон причинности не только регулирует системность в поведении Вселенной, но и обусловливает ее фундаментальную фактуру (уровни элементарных частиц, атомов, молекул, физических тел, астрофизических образований, а также живой природы и человеческого общества).
В дальнейшем (см. разд. 1.8) мы увидим, что под влиянием каузальности сами законы природы подчиняются определенной матрице архетипов, среди которых преобладают принципы двойственности, симметрии, цикличности (симметрии во времени), асимметрии, иерархии. И это не так абстрактно, как кажется на первый взгляд.
1.7. Мироздание за пределами Вселенной
Мы подошли к наиболее интересной странице обзора физического мира. Негласно всегда считалось, что вопрос о природе Мироздания за пределами нашей Вселенной лишен смысла. Такая позиция зиждилась на изотропной модели космоса. Действительно, если Вселенная повсеместно однородна, то она не может иметь границ и каких-то «запредельных территорий». Изотропная Вселенная мыслилась как замкнутое на себя образование наподобие поверхности шара, по которой можно блуждать бесконечно, но при этом нельзя достигнуть края. Предполагалось, что Вселенную так искажает собственная гравитация, и в свете положений теории относительности это вполне возможно.
В настоящее время положение дел изменилось. Выше мы привели ряд фундаментальных фактов, в свете которых привлекательной становится анизотропная модель Вселенной с центром, периферией и, соответственно, внешними границами. При этом пограничная плотность вещества Вселенной автоматически оценивается как низкая. Она не способна обеспечить гравитационную замкнутость нашего мира на себя в том смысле, о котором говорилось выше. Отсюда следует, что край нашей Вселенной представляет собой стандартные границы определенной физической системы, за пределами которых расположены иные физические системы. В результате мы сталкиваемся с необходимостью ответить на вопрос: что может лежать за этими границами, и от каких физических систем они нас отделяют? Наблюдательные данные на этот счет пока отсутствуют, и на первый взгляд кажется, что вопрос по-прежнему не имеет практического смысла. Однако пессимизм такого рода неоправдан. Если за пределами нашей Вселенной имеется некое Мироздание, то оно неизбежно должно было наложить определенный отпечаток на наш мир. Задача состоит в том, чтобы вычленить и истолковать этот отпечаток, и мы рискнем за это взяться, прибегнув к фактам и логике.
Сингулярность не знала причинно-следственного закона. Отсюда вытекает, что породившее ее Мироздание относилось к причинности «без сердца». Положение дел в нем было на тот момент произвольным, и оно где подчинялось, где не подчинялось каузальности. В результате свойства подобного Мироздания варьировались бы в неограниченных пределах. Это был бы мир вечных превращений, Вечно Изменяющийся Мир (ВИМ). Диалектические особенности нашей Вселенной надо расценивать как его реликты.
Этот вывод уже перспективен, поскольку, несмотря на то что Гераклит Эфесский открыл диалектические свойства мира 2500 лет назад [104, с. 176–257], о происхождении диалектики природы и по сей день не известно ничего. В основе гераклитовского учения лежало представление о том, что существующие формы вещей являются их крайними состояниями, а подспудно все без исключения варианты объектов способны плавно перетекать друг в друга, в том числе в противоположные себе варианты. Например, прямые и кривые линии с очевидностью противоположны друг другу. Однако в больших масштабах их отрезки неразличимы. Горячее становится холодным, сухое – влажным и т. д. Почему мир устроен именно так, не ясно. Однако, если диалектика природы является отголоском ВИМа, где из минусов могли бы беспричинно вытекать плюсы в силу необязательности строгой причинности, то все становится на свои места. Подчеркнем, что своими силами наша Вселенная, подчиненная причинно-следственному закону, никогда не произвела бы противоречивую диалектику. Значит, ее источник действительно лежит вне нашей Вселенной, и это косвенно подкрепляет гипотезу Вечно Изменяющегося Мира, пропитанного контрастами.
Обладая чересполосицей то сравнимых, то несравнимых свойств, ВИМ не мыслится как единое целое. В противоположность нашей однообразной Вселенной, он обязательно распался бы на враждебные области, домены, внутри которых господствовали бы собственные логики или алогичности, несопоставимые с соседскими. Поэтому общая структура ВИМа представляется ячеистой, а ячеистость нашей Вселенной – наследием, полученным от ВИМа. Эти допущения позволяют составить некоторые представления относительно доменов ВИМа, непосредственно породивших Вселенную. По-видимому, из ВИМа происходят краеугольные свойства мира, не находящие объяснения внутри нашей Вселенной (т. е. конкретная скорость света и постоянная Планка, необходимые для фундаментальных расчетов в квантовой механике).
У нас стремительность взаимодействий любых объектов ограничена потолком скорости света. Можно представить, что в Вечно Изменяющемся Мире, бесконечном по возможностям, нашелся бы домен с неограниченными темпами контактов своих объектов. В подобном домене все представимые события произошли бы моментально, и больше ничего не случалось бы. Тогда сложилась бы область вечного покоя, напоминающая нашу сингулярность до ее распада.
Мыслится и домен другого типа. В нашей Вселенной дробность размерностей, масс, единиц времени, плотности и энергии определяется постоянной Планка и не углубляется ниже ее пределов. Это обстоятельство объясняет, почему все объекты нашей Вселенной обладают относительно устойчивыми формами и не подвергаются бесконечному дроблению (ср. гл. I, 1 о «демиурге»), что можно присовокупить к списочному составу антропного принципа
[78], предполагающего фундаментальные константы нашего мира в вариантах, благоприятных для зарождения жизни во Вселенной.
В ВИМе мог бы существовать домен, в котором не существует ограничений на квантование вещей, а их составляющие способны быть бесконечно малыми. Поскольку реальная бесконечность недостижима, этот домен пребывал бы в процессе вечного дробления, которое никогда не завершится.
Теперь представим, что домен бесконечных скоростей соприкоснулся с доменом бесконечной дробности. В силу несопоставимости их свойств, взаимопроникновения областей не наступило бы, и они тут же отошли бы друг от друга, оставив точечный след контакта. Этот след обладал бы смешанными свойствами, извлеченными частью из одного, частью из другого домена, а также из самого их взаимодействия, осуществившегося в виде последовательного притяжения и отталкивания, что стало характерно для суперструн нашего мира, поскольку в описываемых событиях мы предполагаем его начало в виде точечной сингулярности.
Сингулярность как след доменного контакта пришла бы в колебательное движение, характерное для суперструн. Его амплитуда принялась бы расширяться, догоняя отступающие домены. Наследуя дробность одного из них, сингулярность колебалась бы с мириадами степеней свободы, расширяющихся в разных пропорциях. Самые убористые породили бы суперструны. Более пространные – вселенскую мировую поверхность виртуальных частиц. Промежуточные – образования крупномасштабной структуры Вселенной, несущие приметы развернутых измерений Первовселенной (см. разд. 1.1). Крохотные обитатели такого мира (т. е. люди) констатировали бы, что живут внутри расширяющейся Вселенной.
От родительских областей ВИМа Вселенной достались бы средние величины скоростей взаимодействия и дробности, которые мы связываем со скоростью света и постоянной Планка. Они должны содержать указания, почему через 12,73 млрд лет после Большого взрыва расширение стало сменяться сжатием под влиянием истончения мировой поверхности и побеждающей внутренней гравитации. Поясним причину, по которой нашей Вселенной достались средние величины скоростей взаимодействия и дробности материи.
Скорость света в пустоте ограничена противодействием, встречаемым фотонами со стороны виртуальных частиц (см. разд. 1.1): чем их больше на единицу объема пространства, тем ниже предельная скорость света. Количество виртуальных частиц зависит от частоты их спонтанного рождения, а та, в свою очередь, определяется планковским квантом времени, поскольку виртуальные частицы не могут возникать чаще, чем раз в планковский квант времени. Если последний почему-либо сократится, виртуальные частицы станут рождаться чаще, а скорость света упадет из-за их возросшего противодействия.
Величина планковского кванта времени положительно зависит от постоянной Планка (кванта действия): чем она меньше, тем меньше и планковский квант времени. Одновременно возрастает средняя частота спонтанного рождения виртуальных частиц, а скорость света, наоборот, снижается. Отсюда следует очевидный вывод, что скорость света в пустоте пропорциональна постоянной Планка, а их соотношение отвечает некой мировой константе, которую еще предстоит определить.
Высказанные предположения поддаются экспериментальной проверке. На стадии расширения и разуплотнения Вселенной сопротивление мировой поверхности фотонам падает, а скорость света закономерно возрастает. Напротив, на стадии сжатия и сгущения Вселенной сопротивление мировой поверхности нарастает, а скорость света, соответственно, снижается. Следовательно, долговременная серия точных определений локальной скорости света в окрестностях Земли может эмпирически показать, в какой Вселенной мы живем: в еще расширяющейся или в уже сжимающейся.
Поскольку агломерат Рыб – Кита, в котором находится Земля, явно сжимается вокруг Великого Аттрактора (см. разд. 1.2), можно предсказать, что локальная скорость света в агломерате будет снижаться. Пропорционально сократится и постоянная Планка. Эмпирическая оценка нашего предсказания – вопрос времени. При этом следует помнить, что за пределами агломерата Рыб – Кита Вселенная все еще по инерции расширяется, и там скорость света и постоянная Планка по-прежнему растут. Проще сказать, что там, где во Вселенной преобладает фиолетовое смещение в спектрах галактик, свидетельствующее об их сближении (агломерат Рыб – Кита, окрестности Великого Аттрактора), скорость света и постоянная Планка убывают, а там, где во Вселенной доминирует красное смещение в спектрах галактик, свидетельствующее об их удалении (Вселенная за пределами агломерата Рыб – Кита), скорость света и постоянная Планка, наоборот, прибывают.
Из сказанного, между прочим, следует, что в древности скорость света во Вселенной была ниже нынешней, а впоследствии возросла. Это должно создавать впечатление, что прежние расстояния между астрономическими объектами превышали теперешние, а красные смещения от удаляющихся галактик выглядели бледнее современных, что в сумме создало бы впечатление более медленного расширения Вселенной, нежели это присуще ей в наши дни. Такое положение вещей дает мнимые факты в пользу идей об ускорении расширения Вселенной с течением времени, об «отталкивающей силе» и «темной энергии» (см. разд. 1.2). Однако, как представляется нам, в основе перечисленных явлений лежат совсем другие причины.
Связь постоянной Планка с величиной скорости света сложилась, по всей видимости, на заре времен. Например, если бы названные величины проистекали от соприкосновения доменов ВИМа, то бесконечно большая скорость взаимодействий одного из них, столкнувшись с бесконечной дробностью материи и бесконечно малым квантом действия другого домена, – эта бесконечно большая скорость взаимодействий автоматически упала бы до конечных значений, так как стала бы измеряться в бесконечно малых величинах планковских длин. Это отвечало бы схеме, в соответствии с которой прежде скорость света была N бесконечно больших планковских длин в секунду, впоследствии грозила стать N бесконечно малых планковских длин в секунду, но закономерно сделалась N средних планковских длин в секунду, что идентифицируется нами как конкретная скорость света в пустоте (299 792 ± 0,4 км/с). Параллельно произошел бы автоматический рост планковской длины и планковского кванта времени от бесконечно малых величин до более существенных, что выразилось бы и в подъеме бесконечно малой постоянной Планка до заметных значений, идентифицируемых нами как наличный квант действия.
Эти зависимости, объясняя взаимосвязанную конечность скорости света и постоянной Планка, свидетельствуют в пользу нашего понимания генезиса Вселенной из Вечно Изменяющегося Мира в результате столкновения его доменов с указанными неограниченными свойствами (с бесконечно большой скоростью взаимодействий и с бесконечно малым квантом действия).
Разбег доменов ВИМа как причина расширения Вселенной – привычная нам условность. В реальности Вселенная испаряется по направлению к удаляющимся доменам ВИМа примерно в силу тех же причин, по которым ветер высушивает и охлаждает некую поверхность. Его дуновения вводят в контакт ненасыщенный и прохладный воздух, который поддерживает испарение с поверхности контакта и ее остывание. Окружающие нас домены ВИМа тоже обновляются ежесекундно: в одном случае из-за неограниченно быстрых взаимодействий, в другом – из-за неограниченно глубокого дробления своей фактуры.
Наследие, полученное от краткого контакта этих доменов, представлено в нашей Вселенной геометрией и основными константами. Два измерения вселенской геометрии – это площадь контакта доменов ВИМа, а третье измерение – направление движения наших границ вслед за расходящимися доменами, засасывающими нас и вызывающими вселенское расширение. Основные константы нашего мира – это средние величины бесконечно больших скоростей взаимодействий и бесконечно малых масштабов дробления материи. Они дали, соответственно, планковскую длину как поперечник площади контакта доменов ВИМа и скорость света как быстроту начального расширения Вселенной. Первое запечатлено в вероятной размерности сингулярности, а второе – в вероятной скорости разбегания границ Вселенной.
Первоначально было множество мод колебаний суперструн, однако причинно-следственный закон позволил уцелеть лишь некоторым, известным нам как наличные суперструны и их квантовые числа (свойства вроде масс, зарядов и пр.). Это обстоятельство служит одним из проявлений асимметричностей нашего мира (см. разд. 1.8).
Если Вселенная находится в недрах какого-то Вечно Изменяющегося Мира и ведет свое начало от его коллизий, то, как и прочие домены, она стоит там особняком. Отчужденность ВИМа исключает физическую связь с ним для нашей Вселенной. Единственное, что может обнаружить его реальность, – это расхождение скорости сжатия Вселенной со скоростью ее расширения, что указало бы на влияние внешнего Мироздания, ускоряющего процесс в случае близости домена неограниченных скоростей и замедляющего в случае соседства с доменом бесконечной дробности, препятствующей сплочению Вселенной.
1.8. Симметрия
Известные нам законы природы, на первый взгляд, чрезвычайно разнообразны, поскольку определяют жизнь весьма непохожих сфер действительности. Между тем, анализ выявляет их общее происхождение и гипотетически указывает на вневселенские корни. Законы природы делятся на динамические, осуществляющиеся однозначно, и статистические, выполняющиеся лишь в определенном проценте случаев, т. е. с известной вероятностью. Динамические законы можно рассматривать как те же статистические, реализующиеся с предельной вероятностью.
Статистические законы отражают альтернативность своих возможных проявлений и в этом смысле совпадают с диалектическим взглядом на мир как на единство противоположностей. Подобная двойственность действительности берет начало в бытии суперструн, которые в качестве волн имеют фазу и противофазу, а потому при взаимодействиях достигают либо ожидаемого, либо противоположного исхода (предсказуема лишь его вероятность). В свою очередь, волновое поведение суперструн повторяет фазу и противофазу контакта родительских доменов ВИМа, что выносит истоки диалектики и статистики за пределы нашего мира, как это видно и из других соображений (см. разд. 1.7).
Обладая фазой и противофазой колебаний, пара суперструн располагает четырьмя вариантами контактов между собой, происходящих с вероятностью в 25% (при учете девятизначных степеней свободы суперструн вариантов становится 324, однако мы опускаем эту громоздкую возможность). На деле может сложиться так, что суперструны случайно прореагируют между собой только одним мыслимым способом, обманув наши ожидания (т. е. нарушив математическое ожидание своего поведения). Теоретически подобная крайность не исключена.
Однако, если дать суперструнам время повторить взаимодействия множество раз (например, 10 тыс., или, так сказать, 100% в квадрате, как рекомендует теория вероятностей), они суммарно поведут себя в соответствии с математическим ожиданием (т. е. как предсказано). Это называется полной реализацией закона больших чисел, и дальнейшие эксперименты с суперструнами уже не принесут ничего нового.
Закон больших чисел является фундаментальной особенностью нашего статистического мира. Его природа обусловлена тем, что причинно-следственный закон не допускает крайностей. Он исключает их потому, что крайности нарушают исходный для него закон неубывания энтропии (см. разд. 1.4), который ведет Вселенную в целом к однородному (тепловому) состоянию, а не наоборот. Закон же больших чисел является математическим выражением того обстоятельства, что отклонения от среднестатистических результатов опыта маловероятны, как того и требует закон причинности, что позволяет выводить закон больших чисел из закона каузальности. Поясним сказанное примером. Допустим, мы 10 раз подбросили монетку. У нас, в принципе, может выпасть 9 «орлов» и 1 «решка». Однако, если повторить эту процедуру 10 тысяч раз, «орел» и «решка» наверняка выпадут у нас практически поровну. Это является классическим выражением закона больших чисел. Если вдруг выпадет 9999 «орлов» и 1 «решка», это будет вопиющим нарушением закона причинности, и он позаботился о том, чтобы ничего подобного не происходило. «Позаботился», конечно, в переносном смысле слова. Просто закон каузальности требует, чтобы в сходных условиях происходили сходные события, а отклонения от них исключались. Поэтому крайности при классических испытаниях закона больших чисел (множество сходных опытов) не наблюдаются.
Благодаря выполнению закона больших чисел крупные совокупности объектов ведут себя повторимо, устойчиво, что обеспечивает долговременную стабильность Вселенной и ее частей, где не наблюдается каких-то спонтанных отклонений от типичного положения вещей (что само по себе еще не обязательно). Это происходит потому, что поведение крупной совокупности объектов статистически равноценно поведению, скажем, пары объектов (или парного объекта), повторенному множество раз, где усматривается архетип проявления закона больших чисел (см. выше).
Недопущение крайних состояний в нашем мире (где даже замкнутые «черные дыры» испаряются) характерно для закона каузальности. Оно отражает гибридное происхождение Вселенной от столкновения пары доменов ВИМа, отличающихся крайностями, что в сумме дало нечто умеренное, наш мир, где крайнее, неповторимое «выпалывается» законом причинности, ибо его стихия – это устойчиво повторяющиеся связи вещей.
Там же усматривается происхождение естественного отбора, пронизывающего все сферы жизни нашей Вселенной. Естественный отбор есть та же дихотомия исхода опыта (по схеме «быть или не быть»), которая отличает и статистику: один вариант действительности выживает, а противоположный элиминируется. Естественный отбор разыгрался еще на заре физической Вселенной, когда взаимодействия в горячем бульоне суперструн вычеркнули из активной жизни суперчастицы и антивещество, а «темное вещество» изолировали от «светлого» во всех отношениях, за исключением сил тяготения. В этом отношении Вселенная напоминает живое существо; ср. [375]. Естественный отбор осуществляется везде, где присутствуют выбор и конкуренция. Он более всего известен в эволюционной биологии и экономике, но действует на уровне любых сущностей и явлений.
Двойственное устройство основ Вселенной предполагает, что каждому объекту в ней соответствует антипод или партнер, т. е. Вселенную пронизывают отношения симметрии. До момента 10–43 с после Большого Взрыва физические взаимодействия были симметричными, а нынешние частицы пребывали в окружении суперчастиц. Затем физическая симметрия пала. Однако она по-прежнему актуальна в сфере химии, где периодический закон построен на началах симметрии. В математике мы имеем Грандиозную теорему, чьи конечные простые группы выдержаны в духе симметрии. Это же наблюдается и всюду, где выдерживаются гармония и равновесие.
Симметричные взаимоотношения вещей распространились и на временну́ю сферу. Это выразилось в подчинении течения событий циклическим алгоритмам с повторами во времени, чему охотно благоприятствовал причинно-следственный закон, радеющий за повторяемость. Собственно, благодаря закону причинности во Вселенной сложились устойчиво повторяющиеся связи вещей, называемые законами природы, хотя ни на каких небесных скрижалях не написано, что это должно быть так – возможно, в иных доменах ВИМа связи вещей, законы выглядят иначе: например, как правила, запрещающие повторы. Как полагал еще Гераклит (см. Введение), цикличность распространилась на саму Вселенную, если та отвечает пульсирующей модели. Цикличным схемам подчинены геологическая тектоника плит, история климата и биоэволюция (см. разд. 2.2, 2.3), а также демографо-технологическая зависимость, действующая в истории социума (см. разд. 3.1). Эти подобия не случайны, ибо они созвучны каузальности.
Между тем, как отмечалось выше, сформировавшийся причинно-следственный закон стал систематически поддерживать часто повторяющиеся явления и искоренять их менее частотные противоположности. Это происходило естественным путем. Например, при остывании Вселенной до 1027К средние дистанции между суперструнами возросли, и тяжелые суперчастицы стали много реже участвовать во взаимодействиях, нежели обычные частицы. Первые вымерли явочным порядком, а вторые возобладали в результате естественного отбора. Во Вселенной восторжествовало асимметричное устройство физических частиц и полей. Сюда же относится элиминация антивещества и дискриминация «темного вещества». Асимметрия пронизывает все сферы бытия. В микробиологии мы имеем левоориентированные молекулы, причем асимметричность аминокислот, возможно, распространена космически, так как находит внеземные аналогии [252]. Вспоминается доминирование левого полушария головного мозга и правой руки у человека (см. разд. 3.2).
Асимметричные начала распространились на действительность через ее иерархическое устройство. Во-первых, бытие состоит из множества явлений, подчиненных относительно немногочисленным сущностям. Во-вторых, астрофизические объекты обращаются вокруг гравитационных центров и стремятся к Великому Аттрактору. В-третьих, в биологических и социальных сообществах рядовые индивиды группируются вокруг доминирующих особей. Если мы сравним структуру Вселенной с планом цивилизованного города, то в обоих случаях обнаружим сходное иерархическое устройство (см. рис. 2 и 10). Иерархия просматривается повсюду, где нечто множественное подчинено единому началу. Это правило не случайно – оно порождено причинно-следственным законом и является универсальным. Механика рождения иерархии из закона причинности состоит в следующем.
По преимуществу причинно-следственный закон стремится к унификации явлений «под одну гребенку». Поэтому в микромире поразительным образом господствуют типологически схожие суперструны, а в макромире – небесные тела двух-трех типов: звезды, планеты (газовые и твердые) и их обломки (кометы, метеориты и астероиды, к которым примыкают спутники планет). Химия зиждется на сотне элементов, а в социуме преобладает однородная толпа и т. д. В то же время на каждом уровне организации материи указанное однообразие создает тесноту, которая методом естественного отбора выдавливает явления на следующий уровень действительности, что и объясняет происхождение «многоэтажности» нашего мира (см. разд. 1.6). Так возникает наиболее общая форма иерархии: в данном случае – иерархия уровней организации материи. И на каждом уровне повторяется аналогичная процедура (а повторы – «любимая тема» каузальности). В результате иерархия опутывает отдельные этажи бытия. К вышеприведенным примерам можно добавить следующие. Биологические экосистемы организованы по модели пищевых пирамид (хищники – травоядные – растения), а человеческое общество «пропитано» иерархичностью. В результате причинно-следственный закон хитроумно сочетает поддержку однообразия с его иерархической организацией. И потому наш мир далеко не прост.
* * *
Как можно убедиться, современная физическая картина мира разительно отличается от представлений конца прошлого века. Радикальные изменения привнесены в нее открытием универсальной элементарной частицы (суперструны) и астрофизическими данными в пользу анизотропности Вселенной. Теоретическое объединение всех физических взаимодействий (Теория Всего Сущего) вот-вот исполнит мечту Калуцы и Клейна о построении единой Теории поля, но принять ее к сведению можно и нужно уже сейчас. Не за горами наступление астрофизиков на Метавселенную, предположительные пути которого мы не оставили своим вниманием. Высказывания о Вечно Изменяющемся Мире следует пока держать в резерве рабочих гипотез, и все же выдвинутые суждения о бытии наблюдаемой Вселенной основаны на фактах.
Глава 2
Земная жизнь
Судьбы человечества неотделимы от истории земной биосферы. Чтобы оценить свое место в мире, нам необходимо правильно понимать историю живого, однако в ней немало неясных и спорных страниц. Успехи эволюционной биологии и сопряженных с ней наук в прошлом веке заполнили множество «белых пятен» в летописи земной жизни, однако дискуссии по ее узловым моментам все еще далеки от завершения.
Мы рассмотрим здесь:
1) современные гипотезы относительно возникновения органической жизни и ее возможной связи с пребыванием Солнечной системы поблизости от центра Вселенной в эпоху биогенеза;
2) общие сведения о глобальной тектонике плит, суперконтинентальном цикле, его влиянии на историю климата, экологию и направленность биологической эволюции;
3) современные сведения об узловых пунктах биологической эволюции, массовых вымираниях и расцветах новых форм организмов, а также о просматривающихся за этим повторяющихся, закономерных чертах эволюционного процесса, который выражается в чередовании мобильных и маломобильных фаун под влиянием оледенений и межледниковий;
4) новейшие данные о возникновении предков человека как о закономерном эпизоде биологической эволюции на Земле, в которой наши пращуры, гоминины, выступили единственными представителями очередной маломобильной фауны млекопитающих, вызванной к жизни одним из кайнозойских потеплений.
2.1. Биогенез
Возраст нашей планеты достигает 4,55 млрд лет [805], а возраст Солнечной системы – 4,566 ± 0,002 млрд лет [395, с. 144]. В эпоху геогенеза (т. е. в период достижения Землей нынешней массы с металлическим ядром и примитивной атмосферой 4,51–4,45 млрд лет назад) расстояние Солнечной системы от центра Вселенной составляло 200,1 млн световых лет (при постоянной Хаббла в 57,4 км/с на мегапарсек и местной плотности Вселенной Ωloc= 0,918), а в эпоху гелиогенеза (т. е. в период формирования Солнца) расстояние нашего новорожденного светила от центра Вселенной не превышало 198,8 млн светолет (при H
0 = 57,5 км/с на мегапарсек и Ωloc= 0,92). Новорожденная Земля пребывала тогда в относительно близких и древних окрестностях центра Вселенной. Это астрофизическое обстоятельство весьма существенно для биогенеза.
Благодаря активному обмену веществом и энергией со средой живые системы устойчивы против Второго начала термодинамики. Они не только противодействуют росту энтропии внутри себя, но и генерируют порядок, т. е. создают негэнтропию. В этой связи встает вопрос: каким образом в расширяющейся Вселенной с растущей энтропией зародилась негэнтропийная жизнь? Проще говоря, живые существа не только умеют постоять за себя перед лицом множества стихий, но и воспроизводят себе подобных (генерируют порядок). Происхождение этих способностей предполагает некую «инкубацию» в тепличных условиях. Однако наша остывающая Вселенная никогда не обещала ничего комфортного в этом смысле.
Земной биогенез пока непостижим [681]. Возможно, жизнь принесена на Землю с другой планеты посредством странствующего небесного тела (кометы, астероида, метеорита) [282; 306; 754]; а также [189; 230; 251; 301; 562; 594]; cp. [525]. Паракодоновый генетический код изученных организмов старше классического [300], что может означать, что исходный код сложился вне Земли, а на Земле переадаптировался. Такой вариант развития событий мыслим, что не снимает вопроса о происхождении жизни, пусть и не на Земле.
На наш взгляд, не исключен следующий сценарий. На ранних стадиях эволюции Вселенной в ее центральных областях сохранялись реликты былой негэнтропии, оставшиеся от сингулярности, в то время как на периферии они рассеивались быстрее (параллельно падению постоянной Хаббла). Условия, благоприятные для зарождения жизни, отвечают среде горячих глубоководных источников с температурой 113° С и более [278; 342; 427; 446; 465; 484; 525; 594]; cp. [212; 339; 571; 614]. Подобная обстановка присуща подводным рифтам с высокими давлениями и кипящим химическим рассолом. Однако в наших океанах такие точки являются оазисами. Вероятность завоевания ими целой землеподобной планеты растет ближе к центру Вселенной, и вот почему.
В окрестностях центра Вселенной велика средняя плотность вещества (см. приложение 7) и, соответственно, значительны гравитационные силы, что сообщает землеподобным планетам данного региона более высокие давления атмосферы и гидросферы. Подобные условия поднимают температуру кипения воды. Внушительное планетарное тяготение сильнее разогревает недра планеты, вызывает бурную мантийную конвекцию (тепловое перемешивание) и активную тектонику плит с обильными рифтами (см. разд. 2.2). На планете с такими условиями формируется плотный океан, густо испещренный глубоководными рифтами, где химически насыщенные воды кипят при температурах, перекрывающих земные, что повышает шансы пребиотического твердофазового синтеза нуклеотидов и аминокислот на минеральных поверхностях [321; 427] – как полагают, существовал вариант самопроизвольного возникновения материала нуклеиновых кислот (дезоксирибонуклеиновых – ДНК ядра клетки, и рибонуклеиновых – РНК цитоплазмы) и аминокислот (строительных блоков для белков) благодаря каталитической помощи некоторых минеральных поверхностей. Планета указанного типа имела бы предпочтительные возможности для биогенеза.
Можно представить, как на заре Вселенной в звездных системах близ ядер галактик, превосходящих по возрасту шаровые скопления на 1–2 млрд лет [270, с. 25], или в самих шаровых скоплениях, поскольку ядра галактик с затравочной «черной дырой» в центре чересчур турбулентны, чтобы служить колыбелью жизни, – в какой-то планетной системе, парящей в центре Вселенной, на планете земной группы, отличающейся вышеописанными свойствами, зародилась органическая жизнь. Первоначально она могла существовать в виде «мира РНК» [353; 754], когда наследственные свойства организмов кодировались только рибонуклеиновой кислотой (РНК), а дезоксирибонуклеиновая кислота (ДНК) еще ожидала своего часа. В известном смысле подобный мир напоминал бы планету вирусов, обходящихся одной РНК: у нас они паразитируют в клетках более сложных существ – там питались бы химическими рассолами.
Метеоритный материал с такой планеты мог бы перенести тамошние вирионы (закуклившиеся в белковую оболочку спящие вирусы) в Солнечную систему и основать «мир РНК» на Земле. Панспермия (космический посев жизни) мыслится и на стадии ДНК-содержащих бактерий.
Попав к нам, живой инопланетный материал угодил бы в нищенскую среду, тем более что молодое Солнце в гадейской эре (4,5–3,8 млрд лет назад) давало лишь 70% нынешних света и тепла. В инфракрасной (тепловой) части спектра недостача восполнялась парниковым эффектом за счет аммония в гадее, а впоследствии за счет метана и особенно двуокиси углерода [208; 450; 601; 691]. Чужеродной теплолюбивой жизни пришлось бы укрыться в оазисах глубоководных горячих источников. Для живых существ это было бы равносильно переселению из более продуктивной (жаркой) в менее продуктивную (просто горячую) среду, что закалило гипотетических мигрантов и подготовило их к земной эволюции вполне определенной направленности, зависящей от геологической и климатической истории планеты.
2.2. Геология
Сущность господствующей в геологии глобальной тектоники плит [65; 77; 96] заключается в следующем. Земля состоит из железо-никелевого, в основном железного ядра (16% объема и 1/3 массы планеты), которое окружено промежуточной оболочкой, мантией (около 2/3 массы планеты), а ее разуплотненный верхний слой, астеносфера, подстилает твердую земную кору, литосферу. Последняя разбита на огромные базальтовые блоки, литосферные плиты. В некоторые из этих плит впаяны обширные, менее плотные гранитные образования, представляющие собой поверхностные континенты. Дно океанов образовано более плотными базальтами.
Разогреваясь от радиоактивного распада расщепляющихся элементов, содержащее их мантийное вещество разуплотняется и всплывает, приводя в движение астеносферу, остывает и вновь погружается в недра Земли (это называется мантийной конвекцией). Текущая астеносфера тянет с собой блоки базальтовой коры. В местах, где они расходятся, вещество астеносферы поступает наружу, формируя срединно-океанические хребты с рифтами (центральными впадинами) и наращивая отступающие базальтовые плиты (поэтому ближе к хребтам они всегда моложе). Происходит раздвижение (спрединг) океанического дна. Противоположные края плит вслед за конвективными течениями погружаются в астеносферу и тают в мантии – при этом плита ныряет под встречную (субдукция). В мантии тонут лишь тяжелые, базальтовые части плит, а гранитные (континентальные) из-за меньшего удельного веса остаются на поверхности планеты и даже наращиваются легкими изверженными породами в местах субдукции (островными дугами вроде Курильских островов).
По своему происхождению все материки являются пакетами спрессованных островных дуг, наслоившихся друг на друга в течение 4,4 млрд лет плитовой тектоники на Земле. Континенты вместе с плитами медленно блуждают по поверхности планеты (континентальный дрейф). По ходу движения они сминаются в гигантские складки (горы вроде Анд). Сталкиваясь между собой, материки тоже сминаются, образуя горы (вроде Урала). Они могут спаиваться в более крупные континентальные образования, способные объединить даже все гранитные массивы в единый материк (Пангею, Мегагею и т. п.), который впоследствии раскалывается под действием мантийной конвекции и разъезжается осколками в разные стороны.
По существующим представлениям [589; 590; 418], данный процесс носит циклический характер. Примерно раз в 500 млн лет материковые массивы, перемещающиеся на своих базальтовых плитах, сближаются над нисходящими токами мантийной конвекции, сталкиваются и спаиваются горными поясами в единый суперконтинент. Его окружает безбрежный океан, базальтовое дно которого все время обновляется вследствие субдукции и спрединга. Сверхматерик душит нисходящий ток конвекции под собой и существует около 100 млн лет. За этот интервал происходит перестройка конвекции в мантии, и под суперконтинентом начинает скапливаться тепло, образуя восходящий ток конвекции. Как следствие, над восходящим током конвекции в сверхматерике закладываются рифты (тепловые трещины), и спустя 40 млн лет (140 от начала цикла) единый континентальный массив раскалывается. Родившиеся из его осколков материки начинают дрейфовать в разные стороны. Между ними возникают новые, молодые океаны (вроде Атлантического) с новым океаническим дном базальтовой породы (220 млн лет от начала цикла). Спустя еще 100 млн лет (320 от начала цикла) континенты максимально удаляются друг от друга.
Однако тяжелая базальтовая океаническая кора не может более 200 млн лет удерживаться на плаву над астеносферой (океанической коры старше 200 млн лет в современных океанах не известно, в том числе в «вечном» Тихом океане [204]). Максимальный возраст присущ базальтовой коре вдоль тыльных границ расходящихся материков. От старости она проваливается в мантию, тянет континенты на себя, и они приходят в попятное движение, поскольку конвекция уже перестроилась под влиянием обрушившейся океанической коры (420 млн лет от начала цикла). Материки начинают сближаться и в конце концов сталкиваются, образуя новый суперконтинент спустя 500 млн лет от начала цикла (цифры здесь ориентировочны). Затем все возобновляется, и часто на месте спаек материков (хотя и не всегда) зарождаются новые рифты, раскалывающие суперконтинент… Таким образом в суперконтинентальном цикле по 200 млн лет уходит на рождение и субдукцию дна молодых океанов, плюс около 100 млн лет на жизнь сверхматерика (что тоже ориентировочно).
Около 250 млн лет назад образовался суперконтинент Пангея (потом он распался, так что современное человечество обитает на его осколках-материках). Однако ему предшествовали и другие сверхматерики, отмеченные 565 ± 5–549/543 млн лет назад (эдиакарская фауна экваториального суперконтинента [203; 258; 293; 381]), а также 1–1,1; 1,5–1,7; 2–2,1; 2,5–2,7 млрд лет назад – последняя дата отвечает рубежу протерозойского (2,5–0,544 млрд лет назад) и архейского (3,8–2,5 млрд лет назад) эонов. Отметим, что перемещающиеся по поверхности планеты континентальные породы существовали уже в раннем гадее 4,404 ± 0,008 млрд лет назад [805]; а также [204; 223; 395]; cp. [419]. Следовательно, в столь отдаленное время (всего спустя ок. 145 млн лет после образования Земли) имели место материковый дрейф и тектоника плит; ср. [459; 460; 478]. Старейшие орогены (горообразования) датированы св. 3,0; 2,8–2,6; 2,0–1,9; 1,90–1,88; 1,88–1,84; 1,78–1,72; 1,67–1,63 млрд лет назад [418; 459]. Часть этих орогенов связана с образованием сверхматериков, часть – с эпизодом расхождения континентов. Заметим, что суперконтинентальный цикл существенен для истории земного климата.
В периоды суперконтинентального цикла Земля переживает по две пары эпизодов с крайними гидростатическими состояниями минимальных и максимальных уровней Мирового океана. В эпоху существования сверхматерика и в эпоху предельного расхождения континентов (как в настоящее время) на планете имеются наибольшие площади глубоководного океанического дна. В результате суша вздымается, уровень Мирового океана падает, а на Земле образуются максимальные по площади материковые поверхности. Последние же, в отличие от поверхности вод, обладают высоким альбедо (т. е. отражательной способностью) и мешают нагреванию планеты Солнцем. Если при этом суша попадает в околополярные области, на ней развиваются покровные оледенения [683; 70, с. 17–18].
Напротив, после раскола суперконтинента, когда начинается дрейф материков друг от друга, и после развития встречных движений континентов, когда они сближаются, океаны между ними мелки, уровень Мирового океана повышается, суша на окраинах материков затопляется, а планетарное альбедо снижается, позволяя Солнцу разогревать Землю. Наступает теплый и влажный климатический период – межледниковье. Иными словами, за время суперконтинентального цикла Земля закономерно переживает два оледенения и два межледниковья. Оледенения случались со средней периодичностью в 150 млн лет [42, с. 24, табл. 1; с. 30–35].
В эпоху оледенения на Земле устанавливается умеренный сухой климат, и такая обстановка не способствует высокой биопродуктивности земной экосреды. Растительность в среднем становится скудной и разреженной в пространстве. Напротив, в эпохи межледниковья климат становится теплым и влажным, что благоприятствует высокой биопродуктивности экосреды; растительность, как правило, богата и скучена в пространстве.
Когда при оледенениях растительность скудна, разрежена, и вообще пищевые ресурсы бедны и рассеяны, животным приходится собирать корм с больших пространств. В такие периоды всем организмам эволюционно выгодно быть подвижными, высокомобильными, хотя это сопряжено с большими потерями энергии на передвижение. В противоположность этому, при межледниковьях растительность изобильна и скучена, как и все пищевые ресурсы, так что животным не надо искать пищу по обширным территориям. В такие эпохи животным и всем организмам вообще ни к чему быть высокомобильными, поскольку эволюционно не выгодно тратить энергию на лишние передвижения.
Проще говоря, наиболее успешные живые организмы при оледенениях становятся высокоподвижными, а при межледниковьях – наоборот. Подобная закономерность представляется схематичной, однако ее можно проиллюстрировать. Допустим, на Земле стоит ледниковый период, в нем господствуют подвижные животные, а на вершине пищевой пирамиды царят высокомобильные хищники, которые, не считаясь с энергозатратами, без устали патрулируют свою экологическую нишу. Пища там скудна и ее необходимо потреблять без остановки, чтобы она не досталась конкурентам. При этом в тени господствующих высокомобильных существ обычно прозябают малоподвижные организмы, на которых никто не обращает внимания.
Но вот оледенение заканчивается, становится тепло и влажно. Растения множатся и скучиваются в пространстве (как ныне свойственно им в тропиках и субтропиках). Наступает эпоха обильных кормовых угодий, и больше не требуется обшаривать значительные территории в поисках еды. Однако подвижные господа прежней, холодной эпохи в силу закона Долло о необратимости эволюции уже не в силах изменить свою физическую и нейрофизиологическую организацию и продолжают взад-вперед сновать по эконише, без толку тратя энергию на суету. Пока эти трудяги прочесывают один край биома, на другом краю «расправляют крылья» недавние аутсайдеры ледниковых времен – малоподвижные организмы. Они не торопясь и экономя энергию потребляют обильную пищу, так что, когда из рейда по экосистеме возвращаются высокомобильные существа, им питаться уже нечем. Пока они удивляются превратностям судьбы, ситуация повторяется в других уголках экосреды. Господа эпохи оледенения приходят в упадок, а воцаряются их малоподвижные конкуренты.
Когда оледенение возвращается вновь, картина воспроизводится с противоположным знаком. Пища вновь скудна и редка. Ее недостает малоподвижным господам межледниковья, а осваивать большие пространства они не в силах. Уцелевшие остатки высокоподвижной фауны, коротающие теплые времена на задворках экосреды (как млекопитающие мезозоя; см. разд. 2.3), поднимают голову, обнаруживают, что снова стали актуальны, и приступают к подвижному патрулированию биоты. Недавние малоподвижные господа теплолюбивой жизни частью не выдерживают нового направления конкуренции и вымирают, а частью уходят в тень – до будущих теплых времен. Многочисленные палеонтологические примеры сказанному мы приводим ниже (см. разд. 2.3).
Таким образом, можно увидеть определенную тенденцию земной эволюции, которая направляется экологией, та, в свою очередь, климатом, а он – дрейфом материков, т. е. циклическим геологическим фактором. Продемонстрируем на фактах, что подобная эволюционная тенденция действительно реализовалась в истории земной жизни.
2.3. Эволюция
Археозой (4,55-2,5 млрд лет назад)
Согласно концепции абиогенеза (о химической эволюции жизни из неорганического материала), процесс начался в глубоководных гипертермальных источниках 4,2–4,0 млрд лет назад, а 4,0–3,7 млрд лет назад достиг поверхностных вод [525]. Жидкая вода возникла на Земле 4,3 млрд лет назад [575]. Следы ископаемых организмов появляются в палеонтологической летописи в позднем гадее св. 3,85 млрд лет назад [422], а также [156; 225; 282; 465; 574; 580; 600; 677; 678; 788]. Отметим, что по молекулярно-генетическим данным возраст первых организмов оценивается приблизительно в 1,8 млрд лет [578], что вдвое уступает палеонтологическому возрасту; по-видимому, 1,8 млрд лет назад – это возраст эволюционного расхождения тех организмов, чьи прямые потомки дожили до наших дней, поскольку молекулярно-генетический возраст, например, нескольких родственных видов живых существ определяется до момента разветвления эволюционных путей их предков.
Под влиянием мобилизующих колебаний продуктивности среды древние микроорганизмы развили пищевую автономность (автотрофность), чтобы не зависеть от внешних ресурсов в неурожайные (ледниковые) периоды. Около 3,8–3,5 (или 3,5; 3,465; 3,416) млрд лет назад цианобактерии
освоили фотосинтез, т. е. способность синтезировать углеводы, аминокислоты, белки, пигменты и другие соединения под действием солнечного света [232; 788; 677; 678; 753]; а также [399; 465; 600]. Побочным продуктом фотосинтеза является свободный кислород. Однако поначалу он практически весь уходил на окисление железа, так что 2,8 млрд лет назад свободного кислорода в атмосфере почти не было [465, с. 2112]. Лишь 2,32 млрд лет назад наметился его атмосферный рост [181].
Чтобы добиться еще большей независимости от пищевых ресурсов среды, микроорганизмы путем симбиотического включения в свои тельца других одноклеточных существ обзавелись св. 2,8 млрд лет назад митохондриями [470], служащими внутренними источниками энергии для клеток, что подобно аккумуляторам повышало их мобильность. Митохондрии поначалу были самостоятельными микроорганизмами, а впоследствии поселились в тельцах других микроорганизмов и стали служить им «энергетическими подстанциями».
Примерно тогда же, св. 2,7 млрд лет назад, прокариоты (организмы без клеточного ядра) путем аналогичного симбиоза поселили в своих тельцах другие микроорганизмы, которые стали играть для них роль клеточных ядер. В результате прокариоты стали эукариотами, организмами с клеточными ядрами [213; 294; 322; 465; 718]. Этим способом содержащийся в клеточном ядре геном приобрел как бы скафандр в виде материнской клетки, что повысило его независимость от условий среды и ее перепадов, а следовательно подняло его мобильность. Поэтому хронологическая близость возникновения митохондрий и клеточных ядер может отвечать климатическому пессимуму, возможно связанному с оледенением, ознаменовавшим расхождение материков 2,85 млрд лет назад. Отметим, что в силу той же логики появление белковой оболочки (вириона) у РНК-вирусов тоже могло быть откликом на климатический или другой пессимум в их истории, однако палеонтология вирусов не развита.
Протерозой (2,5-0,544 млрд лет назад)
После распада позднеархейского суперконтинента Мегагеи, образовавшегося и расколовшегося 2,7–2,5 млрд лет назад, имели место три Гуронских ледниковых периода (Рамсей-Лейк, Брюс и Гоуганда) в интервале 2,5–2,2 млрд лет назад. Они, вероятно, связаны с эпизодом сильного расхождения материков (см. разд. 2.2). Падение продуктивности среды при III палеопротерозойском низкоширотном оледенении (III Гуронском) 2,222 ± 0,013 млрд лет назад [181; 313; 465] подстегнуло тягу жизни к самостоятельности вообще и автотрофности в частности. Поэтому жизнь ок. 2,2 млрд лет назад овладела способностью фиксировать азот [593], что ослабило ее зависимость от стационарных природных источников этого элемента, необходимого для поддержания существования живых организмов. Здесь мы имеем дело с одним из ранних фактов повышения мобильности организмов под влиянием оледенения, точнее под воздействием вызванной им убыли продуктивности среды.
Другим способом повысить мобильность организмов выступила многоклеточность. Современные бактерии способны образовывать колонии с высокоупорядоченным строением, в которых бактерии дифференцируются на клетки различных типов. Колонии некоторых миксобактерий (например Chondromyces crocatus) формируются в виде многоклеточного плодового тела со скоплениями одноклеточных спор на концах ответвлений. Это едва ли не прообраз многоклеточных растений, переживший свое время. Другие миксобактерии (например Myxococcus xanthus) образуют сферичные колонии из миллионов клеток. Такие колонии способны двигаться, захватывать и переваривать жертвы. Подобные хищные сферы в чем-то напоминают многоклеточных животных [115]. Многоклеточность последнего типа нацелена на подвижный сбор разреженной пищи, т. е. на ледниковые условия.
В соответствии с такими представлениями о назначении многоклеточности первые макроскопические многоклеточные водоросли появились в палеонтологической летописи 2,110 ± 0,052 млрд лет назад ([399; 698]) в Даспортскую ледниковую эпоху (2,2–1,95 млрд лет назад), вызванную образованием следующего сверхматерика (2,1–2,0 млрд лет назад). Наступившие вслед за Даспортским оледенением условия климатического оптимума расширили экологические ниши. Это произошло потому, что мягкие условия способствовали подъему биопродуктивности среды, а ее богатство сопровождалось ростом разнообразия природных условий и, соответственно, раздвижением экониш, что в свою очередь благоприятствовало умножению и экспансии жизни.
Судя по всему, простейшие организмы 1,2 млрд лет назад освоили новые для себя наземные водоемы [423]. Одновременно произошла крупная радиация (всплеск разнообразия) многоклеточных организмов, выразившаяся в разделении предков растений, грибов и животных [294; 322]; cp. [312; 780]. По молекулярно-генетическим данным, процесс продолжился отделением от животных губок, затем гребневиков, медуз и морских анемон, далее – плоских червей [312], а потом – дивергенцией (разделением) хордовых животных (к которым принадлежат и люди) и моллюсков (1,225 млрд лет назад), разделением хордовых и кольчатых червей (1,204 млрд лет назад), расхождением хордовых и членистоногих (1,173 млрд лет назад) и дивергенцией хордовых и иглокожих (1,001 млрд лет назад) [824]; cp. [294; 322; 470]. Дальнейший путь развития животных отмечен так называемой преэдиакарской биотой 980–900 млн лет назад [259, с. 220, рис. 1], которая связана с теплым эпизодом, благоприятным для умножения экониш и расцвета фауны.
Вслед за распадом еще одного сверхматерика возраста 1,1–1,0 млрд лет назад наступали ледниковые периоды Гнейсе, 900–850 млн лет назад, и Стёрт, 760–740 млн лет назад, отвечавшие состоянию разреженности материков. Очередной суперконтинент сформировался 675–600 млн лет назад, что повлекло за собой низкоширотное оледенение Марино-Варангер ок. 680–606 ± 3,7/ – 2,9, 602 ± млн лет назад [313; 381; 465; 807]. Низкоширотность (тропичность) оледенений Гурон III и Варангер, определенная палеомагнитным методом (когда по намагниченности ископаемых пород рассчитывают положение магнитных полюсов в древности), объясняется дрейфом магнитного полюса Земли относительно географического полюса. В протерозое этот дрейф превосходил современный и заметно искажал регистрацию палеоширот по намагниченности ископаемых пород.
Выгодность мобильности в ледниковых условиях оживила подвижных многоклеточных животных, представленных норами червей 620 млн лет назад [464; 682]. Последовавшее потепление эдиакарского периода (ок. 600–544 млн лет назад [312; 381]) создало множество экониш и вызвало дивергенцию бесчелюстных рыбообразных животных (круглоротых вроде миног и миксин) и челюстноротых (предков рыб) 599 млн лет назад [824; 479]. В результате развернулась богатая эдиакарская фауна возрастом в 565 ± 5–549/543 млн лет назад [203; 258; 381], отмеченная в финале своего расцвета молекулярно-генетической дивергенцией позвоночных животных (наших предков) 546 ± 18 млн лет назад [479; 838].
Палеозой (544-251 млн лет назад)
Развившееся хищничество одело мягкотелых эдиакарцев в твердые наружные покровы [258], поэтому кембрийский период (544–510/505 млн лет назад) ознаменовался широким распространением животных с защитным наружным скелетом, а также хордовых животных, чей внутренний скелет позволял создавать в воде локомоторную волну (толкающую животное вперед [124]) и повышал спасительную подвижность. Хищники производят эффект снижения биопродуктивности среды, поскольку мешают консументам (в данном случае – нехищным потребителям) беззаботно кормиться. Поэтому усвоение последними упомянутой спасительной подвижности типологически близко реакции на падение биопродуктивности среды вследствие похолодания. Следом за жертвами в локомоции усовершенствовались и хищники, примером которых может служить подвижный нижнекембрийский Anomalocaris (45 см), напоминавший плоскую сигару с боковыми плавниками и парой хватательных придатков возле рта. Организмы, отставшие от описанной тенденции, пришли к массовому вымиранию в раннем кембрии [184; 674, с. 431–437].
Целым «пакетом» в нижнем кембрии отмечены такие животные, как членистоногие [85, с. 41; 211; 239]; ракообразные [712]; иглокожие [700]; полухордовые (век атдабан, ок. 525 млн лет назад) [237; 701; 703; 727]; оболочники [699]; хордовые (век ботом) [237]; черепноподобные (530 млн лет назад) [238; 308]; миногообразные и миксинообразные (ок. 530 млн лет назад) [702; 838]; в верхнем кембрии – конодонты (родня позвоночных; 515 млн лет назад) [669; 210; 325; 437] и другие позвоночные (514 млн лет назад) [479]. Так выглядел древнейший эволюционный взрыв в истории Земли, обусловленный теплом и высокой биопродуктивностью эдиакарского и кембрийского периодов.
В следующем, ордовикском, периоде (510/505–438 млн лет назад) материки максимально отдалились друг от друга, что вызвало в позднем ордовике (458–438 млн лет назад) оледенение, условия которого обострили спрос на мобильность животных. Из числа головоногих моллюсков, появившихся в нижнем кембрии (например, наутилоидеи, потомки которых из рода Nautilus, живые ископаемые, и поныне здравствуют на юго-западе Тихого океана), особый расцвет в ордовике испытали представители подкласса эндоцератит [38, с. 356–359; 83, с. 173–204]. Их тело помещалось в длинно– и короткоконических раковинах прямых очертаний, игравших роль гидродинамических обтекателей. В раковине имелось образование – сифон, выполнявший гидростатические функции (он имелся и у других раковинных головоногих моллюсков и работал, как плавательный пузырь рыб, т. е. помогал своему обладателю менять общий удельный вес тела). Гидродинамическая раковина вместе с гидростатическим сифоном (он наполнял соответствующие камеры раковины азотом, аргоном и жидкостью, как у современного наутилуса, и тем самым регулировал удельный вес тела и его частей) делали эндоцератит идеальными пловцами, т. е. высокомобильными животными, типологически напоминающими по устройству подводные лодки на реактивной тяге.
В связи с похолоданием, поощряющим мобильность, эндоцератиты расцвели, стали владыками морей и породили самых крупных среди известных раковинных беспозвоночных, до 9,5 м длиной [38, с. 356; 83, с. 191]. Казалось бы, такие успешные монстры должны были бы процветать долго, однако с концом позднеордовикского оледенения они неизбежно пришли в упадок. Представители ведущего отряда, эндоцератиды, просуществовали весь ордовик и вымерли – возможно, один род данного отряда (Humeoceras) дожил до середины силура (421 млн лет назад) [38, с. 358]. Парадокс этот вполне объясним, если учесть, что высокомобильные формы вроде эндоцератит были узко приспособлены к скромной экосреде окололедникового времени и лишь в ней были сильны, а с подъемом биопродуктивности среды в силуре перестали ей соответствовать и угасли.
Прохладный ордовик был периодом радиации жизни [297], в особенности холодолюбивой – точнее, годной к низкой биопродуктивности среды. Холодолюбивые существа активно замещали ушедших в небытие теплолюбивых хозяев кембрия. Давление в пользу мобильных организмов привело на сушу наземную флору. Среди пионеров были наземные грибы (конец нижнего ордовика, ок. 460 млн лет назад), которые, по молекулярно-генетическим данным, возникли еще в конце Варангского оледенения ок. 600 млн лет назад [636], т. е. уже были испытаны суровыми условиями. Бок о бок с ними по суше распространялись наземные растения, отмеченные как в нижнем ордовике (475 млн лет назад), так и в верхнем (458–438 млн лет назад) [346, с. 481; 794]. В морях, судя по молекулярно-генетическим данным, появились подвижные лучеперые рыбы (450±35,5 млн лет назад) [86, с. 323–484; 479], а также акулы [468]. Лучеперые рыбы и акулы составляют свыше 95% современной ихтиофауны, захватившей моря благодаря кайнозойскому похолоданию. Их успех в современных условиях объясняется происхождением из позднеордовикского оледенения (см. далее).
Акулы, помимо завидной подвижности, во многих случаях развили способность к яйцеживорождению и живорождению с подобием плацентарного вскармливания эмбрионов (как у холодолюбивых млекопитающих). Это является эволюционным признаком мобильности, поскольку потомство выводится не из стационарно отложенной икры, а на ходу, что повышает его шансы на распространение по экосреде. Добавим, что и яйцекладущие акулы по-своему мобильны. Они откладывают яйца (икринки) в твердой оболочке, чем уподобляются, например, рептилиям, у первых представителей которых способность откладывать яйца в твердой скорлупе послужила одной из основ сухопутной подвижности.
Лучеперые рыбы и акулы не успели продвинуться в ордовике так сильно, как эндоцератиты, а потому благополучно перенесли позднеордовикское вымирание, более суровое к остальной биоте [184; 239; 836]. Объяснение парадоксального уклонения лучеперых рыб и акул от позднеордовикского вымирания благодаря отставанию от эндоцератитов мы приведем ниже (вместе с рядом аналогичных случаев).
Силурийский (438–408 млн лет назад) и девонский (408–354 млн лет назад) периоды были временем сближения материков и, соответственно, эпохой прибывающего и отступающего потепления. По мере его углубления биопродуктивность среды возрастала, и суша превращалась в притягательную эконишу для консументов, потребителей органики, благо сухопутная флора к тому времени процветала уже не один миллион лет.
В верхнем ордовике под влиянием отступления оледенения сушу освоили норные животные, в которых подозревают членистоногих многоножек [643]. Затем, в позднем силуре (век пржидоли, 414 млн лет назад), землю завоевали такие членистоногие, как паукообразные и непосредственно зафиксированные многоножки [440]. Позже, в нижнем девоне (век зиген, 401–394 млн лет назад), их состав пополнили насекомые [307; 483], чьи крылья размером до 2 см поначалу служили целям терморегуляции, а с повышением размера до 4 см стали применяться для полета [502]. Поскольку полет увеличивал мобильность насекомых, их крылья должны были служить органами движения в некий холодный период. Однако данных на этот счет пока нет.
Обзаведясь фауной беспозвоночных, суша привлекла позвоночных хищников, которые прибыли в новую для себя экосреду в раннем верхнем девоне (век верхний фран, ок. 370 млн лет назад) [128а; 129, с. 420; 130, с. 508]; а также [246; 253; 274]. Вышедшие из пресноводных водоемов [834], первые земноводные были шестипалыми [345]. Они происходили от кистеперых рыб [334], а те – от рыб вроде Styloichthys changae (век поздний лочлов (жедин), нижний девон Китая), связывающих предков бесчелюстных рыбообразных с предками четвероногих позвоночных (земноводных, пресмыкающихся и млекопитающих) [835]. Рожденные межледниковьем, земноводные были маломобильными существами, а потому держались водоемов, привязанные к ним способом размножения (когда икра откладывается в воду, и там же развивается молодь, как у современных лягушек). Можно было бы предсказать, что, попади земноводные под давление отбора на высокомобильность, они изменили бы способ размножения на независимый от водной среды, принявшись прочесывать сушу и обзаведясь для этого непроницаемыми покровами тела взамен слизистой голой кожи, служащей для дыхания. Все это осуществили пресмыкающиеся.
По мере сближения материков климат девона портился, биопродуктивность среды падала, и маломобильные организмы погружались в кризис, что выразилось в позднедевонском вымирании (рубеж веков фран/ фамен, 367 млн лет назад), которое, помимо всего прочего, вызвало гибель тропических коралловых рифов и убыль чистой биомассы [184; 239]. Это прямо отражало снижение палеотемператур (чего не переносили прикрепленные маломобильные коралловые полипы) и оскудение среды.
Вслед за теплым девоном наступили каменноугольный (карбон, 354–286 млн лет назад) и пермский (286–251 млн лет назад) периоды. В это время континенты все более сближались, чтобы образовать сверхматерик Пангея (ок. 250–180 млн лет назад). Эти события вызвали пермо-карбоновое, или гондванское (по южной половине Пангеи – Гондване), оледенение (310–260 млн лет назад). Под давлением отбора на мобильность в соответствии с вышеописанным сценарием земноводные превратились в амниот (от греч.
'αμνίον – околозародышевый пузырь), т. е. начали откладывать яйца в плотной зародышевой оболочке, позволяющей им развиваться на суше. Защищенная чешуей кожа открывала недавним амфибиям сухопутные просторы. Появились пресмыкающиеся (век бригант (визей), нижний карбон, ок. 338 млн лет назад) [716].
На протяжении пермо-карбонового оледенения пресмыкающиеся сохраняли четвероногость, которая обеспечивала им максимальную мобильность. Ошибочно думать, будто ранние пресмыкающиеся не были в состоянии усвоить какую-либо другую локомоцию. Напротив, в их среде появилась интересная ящерицеобразная рептилия Eudibamus cursoris (из числа Bolosauridae, ранняя нижняя пермь, век ассель, 290 млн лет назад) [187]. Она напоминала ряд современных ящериц малопродуктивных, открытых пространств, бегающих рысью, переходящей в бег на двух задних ногах (такова, например, австралийская плащеносная ящерица), что отличается от энергосберегающей шагающей двуногой локомоции динозавро-человеческого типа. Отсюда следует, что E. сursoris был порожден оледенением и не имел ничего общего с двуногими динозаврами и людьми (в смыле конвергентной, внешне похожей эволюции), но подтверждал способность ранних рептилий разнообразить свою походку.
Здесь необходимо остановиться на способах локомоции (передвижения), присущих высшим наземным позвоночным. За вычетом промежуточных вариантов вроде бега E. cursoris или продвинутых кроличье-лягушачьих прыжков кенгуру, тушканчиков и им подобных животных существуют два основных типа локомоции: четвероногая, как у подавляющего большинства современных млекопитающих и «холодолюбивых» динозавров, и шагающая двуногая, как у «теплолюбивых» динозавров, людей и некоторых других существ.
Четвероногость весьма расточительна энергетически, сопряжена со значительным сцеплением животного с субстратом (из-за четырех конечностей), однако обеспечивает ему исключительную выносливость, проходимость, значительные скорости и дистанции передвижения. Подобная адаптация крайне полезна в прохладных условиях низкой биопродуктивности среды, когда скудную пищу приходится собирать с больших пространств.
В противоположность этому шагающая двуногость весьма выгодна, экономна энергетически, не сопряжена с большим сцеплением тела с субстратом (благодаря опоре на две конечности), однако не дает своему обладателю выносливости и не позволяет осваивать значительные территории. Такой способ передвижения выгоден в теплые эпохи высокой биопродуктивности среды, когда пища богата, скучена в пространстве и не требует больших путешествий для своей добычи. При этом двуногое существо очень выигрывает, экономя энергию на передвижении. С конкретными примерами такого рода мы познакомимся ниже.
Пермокарбоновое оледенение отмечено еще двумя перспективными достижениями биосферы. По молекулярно-генетическим данным, в это время возникли цветковые (покрытосеменные) растения. В конце нижнего карбона (век намюр, 319 ± 35 млн лет назад) они разделились на однодольные и двудольные, а в перми (276 ± 33 млн лет назад) двудольные растения претерпели радиацию [545]. Покрытосеменные растения были родственны растениям из рода Glossopteris, преобладавшему в перми и отличавшемуся простым способом размножения, как у цикадовых и гинкго [595]. Происходя из холодного ордовика, все наземные растения имели мобильные средства размножения (споры, семена) и совершенствовались в этой части. Возникнув в холодном карбоне, цветковые растения сделали здесь очередной шаг вперед: благодаря различным приспособлениям их семена распространялись на большие расстояния лучше, чем у голосеменных (например хвойных), хотя последние уступили преобладающее положение не сразу [88, с. 21–382].
Другое достижение касается холодолюбивой фауны, в рамках которой пресмыкающиеся котилозавры породили в верхнем карбоне звероподобных рептилий, господствовавших в перми, постепенно убывающих в триасе, вымирающих в средней юре, однако отмеченных в поздней юре и даже в позднем палеоцене (век ранний тиффан, ок. 60–57,3 млн лет назад) [25, с. 110–117; 87, с. 230–298; 250; 328; 655, с. 173–186]. От этих четвероногих рептилий в перми произошли зверозубые пресмыкающиеся – чрезвычайно прогрессивные существа: некоторые, по-видимому, стали теплокровными, покрылись шерстью и усвоили разнозубость. Зубы прочих пресмыкающихся обычно однородны (за вычетом динозавра гетеродонтозавра из позднего триаса или ранней юры (ок. 201 млн лет назад), у которого самцы имели клыки [37, с. 46–47]; гетеродонтность динозавра Eoraptor lunensis из верхнего триаса (ок. 230 млн лет назад) далека от разнообразия зверозубых рептилий и млекопитающих, cp. [689, с. 64, рис. 1]). У продвинутых же зверозубых пресмыкающихся зубы дифференцированы на резцы, клыки и коренные (как у их потомков – млекопитающих). Это требовалось для эффективного использования небогатой и разреженной пищи ледниковья. Казалось бы, перед прогрессивными зверозубыми рептилиями открыт весь мир. Между тем, их постепенно смело с эволюционной сцены.
С концом пермского похолодания грянуло позднепермское вымирание, неблагосклонное к мобильным организмам. Однако оно лишь потеснило цветковые растения и зверозубых рептилий. Это вымирание осуществлялось в две стадии: гваделупа, ок. 256 млн лет назад, истребившая 58% родов морских организмов, и татарий, ок. 251 млн лет назад, уничтоживший 67% родов морских организмов [723; 825 a]; cp. [35, с. 87, рис. 2; 184; 239; 311; 471; 639; 674, с. 437–439; 802].
Сухопутная биота понесла сходные потери, отвечающие эволюционно-экологической направленности естественного отбора, связанного с потеплением. Закономерно были утрачены небольшие подвижные рыбоеды и насекомоядные существа. Их участь разделили средние и крупные травоядные, привыкшие собирать скудные корма с обширных пространств. Высшие хищники потянулись следом. Даже спустя 15 млн лет этот урон среди пресмыкающихся еще не восполнился [185].
Легко понять, что избирательное двухступенчатое пермо-триасовое вымирание подчинялось определенной эволюционной логике, а не уничтожало живых существ без разбора, как свойственно стихийным бедствиям вроде падений астероидов или извержений вулканов. Как и в случае с мелтретичным вымиранием (см. далее), углубление исследований рисует эти биокатастрофы все менее окказионально (т. е. все менее случайно).
Мезозой (251-65 млн лет назад)
Наступил триасовый период (251–201 млн лет назад), начало которого ознаменовалось «метановым потеплением» (вследствие парникового эффекта от вулканических газов, включая метан) [742] и морской трансгрессией (подъемом уровня Мирового океана) [74, с. 219, рис. 7.1; 401]. Естественный отбор принялся поощрять маломобильных животных. Еще в верхней перми от рептилий-эозухий, близких прогрессивным котилозаврам-капторинам и низшим зверообразным пеликозаврам (заложившим черты молекулярно-генетического родства своих потомков-млекопитающих и людей с потомками текодонтов, т. е. птицами), произошли пресмыкающиеся текодонты (псевдозухии). Под влиянием потепления и связанного с ним роста биопродуктивности среды, они усвоили экономичную шагающую двуногость человеческого типа, длительное время процветали, однако не перенесли похолодания (падения биопродуктивности среды) в конце триасового периода и вымерли. Однако до того, в середине триаса, морская трансгрессия и потепление обострили давление отбора в пользу маломобильных животных. В результате от текодонтов монофилетично (из одного корня) произошли двуногие динозавры двух отрядов (ящеротазовые и птицетазовые), 227,8 ± 0,3 млн лет назад (см. рис. 3) [87, с. 494, 525; 271; 324; 329; 653; 655, с. 138–141; 688–690].
Судя по своим поздним потомкам мелового периода и подобно ближайшим родичам, птицам и крокодилам, динозавры имели четырехкамерное сердце. Аналогично птицам они были покрыты перьями [148; 441; 747; 827–829], возможно, отличались постоянной температурой тела (гомеотермией) [173]; cp. [660] и даже спали в птичьей позе [826]. Они были сходны с птицами в особенностях роста челюсти [172], но расходились с ними в скоростях общего роста [310], что можно объяснить повышенным метаболизмом (обменом веществ) у последних вследствие летающего образа жизни. Кроме того, двуногим динозаврам, даже их верхнемеловому венцу, хищному колоссу тираннозавру, были присущи маломобильность [430] и равнозубость, ориентированная на расточительное потребление богатой и скученной пищи, – то и другое гармонировало с высокой биопродуктивностью среды.
Длительное триасовое потепление раздвинуло существующие экониши и вовлекло в них маломобильных существ, что привело пресмыкающихся в водную среду и дало известных ихтиозавров (средний триас – поздний мел), плезиозавров (тогда же) и других водных ящеров [25, с. 18, 22, 33]. Похолодание позднего триаса (норий-рэтийская морская регрессия, падение уровня Мирового океана) привело к упадку и частичному вымиранию маломобильных ихтиозавров. Выжили те их формы, которые лучше освоили живорождение, являющееся вариантом неявной мобильности.
То же похолодание отпочковало от динозавров подвижных летающих птиц, засвидетельствованных спорными остатками Protoavis texensis («первоптицы из Техаса», поздний триас, ок. 225 млн лет назад [141]; cp. [240; 326]), ископаемыми птичьими следами (поздний триас, века норий-рэт, св. 212,5 ± 7 млн лет назад [565]) и синхронным молекулярно-генетическим расхождением птиц с крокодилами (222 ± 52 млн лет назад [479]), что, впрочем, не исключает обособления предков птиц от крокодилов еще в составе динозавров (т. е. сначала от крокодилов отделились динозавры, а впоследствии, много позже, от динозавров обособились птицы вроде археоптерикса, которые положили начало современной авиафауне, что представляется нам сомнительным, см. ниже).

Рис. 3. Ставрикозавр
(Staurikosaurus pricei, «ящер созвездия Южного Креста») из семейства ставрикозаврид подотряда теропод (звероногов) отряда заурисхий (ящеротазовых динозавров) был ловким двуногим хищником и охотился в среднем триасе, не подозревая, что в конце этого периода его отсталые родичи прозауроподы встанут на четвереньки и дадут начало крупнейшим наземным животным планеты, зауроподам. [37, с. 30–31]
Позднетриасовое вымирание (210 млн лет назад [184; 239; 518]) затронуло малоподвижных животных, но оживило четвероногих, что выразилось в оформлении млекопитающих как класса позвоночных животных (век норий, св. 200 млн лет назад [330]). Однако затем рэтийская регрессия ненадолго сменилась парниковым потеплением на 3–4° вследствие роста вулканического диоксида углерода 205,7 ± 4 млн лет назад [560], что спровоцировало вымирание мегафлоры, ориентированной на падение биопродуктивности среды.
Наступивший юрский период (201–144,2 млн лет назад) открылся очередной регрессией (похолоданием) и потряс двуногих динозавров – произошло их ограниченное нижнеюрское вымирание [239, с. 49]. Если целурозавры и карнозавры всегда оставались удачливыми, а потому эволюционно негибкими двуногами, то неуклюжие двуногие прозауроподы были готовы ко всему новому (о причинах чего см. далее) и охотно перешли к четвероногой локомоции, превратившись в гигантских зауропод (нижняя юра, век поздний лейас – верхний мел) (см. рис. 4, 5).
Отметим впечатляющую парадоксальность эволюционного процесса: предки динозавров (текодонты) сначала «встают» с четырех ног на две, порождают успешных двуногих динозавров, а затем вновь «опускаются» на четыре конечности (как произошло в случае с зауроподами и некоторыми другими динозаврами, см. далее). В рамках нашей версии эволюционной направленности этот нонсенс находит естественное объяснение – климато-экологическое. Возможно, обсуждаемое нижнеюрское похолодание развило у гетеродонтозавров разнозубость, являющуюся адаптацией к низкой биопродуктивности среды, свойственной похолоданиям.
После метанового потепления (ок.183 млн лет назад [742]) в середине средней юры (век поздний байос) палеотемпературы упали с наступлением антарктического микрооледенения, о чем свидетельствуют тиллиты (ископаемые морены, ледниковые отложения) юрского времени в Антарктиде [42, с. 33; 74, с. 220, 222], однако затем тепло стало нарастать к максимуму трансгрессии в середине верхней юры (конец века кимериджа). Средне-верхнеюрское потепление пагубно сказалось на четвероногих звероподобных рептилиях, которые почти полностью вымерли, оставив по себе молодых отпрысков, млекопитающих. То же самое произошло с четвероногими зауроподами, испытавшими с потеплением первый эпизод вымирания [25, с. 56]. Их судьбу повторили четвероногие птицетазовые динозавры сцелидозавры из ранней юры, когда их четвероногость была закономерна. Во второй половине юрского периода они дали четвероногих же стегозавров (середина юры – ранний мел), а через сирмозавров (конец раннего мела) – панцирных анкилозавров (ранний – поздний мел) [25, с. 79]. Иными словами, эволюция этих существ шла как бы «ракетным способом», когда в моменты потеплений отбрасываются «отработанные» (устаревшие и вымирающие) ступени.

Рис. 4.
Платеозавр
(Plateosaurus engelhardti, плоский ящер Энгельгардта) из семейства платеозаврид инфраотряда прозауропод (проящероногов) из подотряда теропод группы зауроподоморфов (ящероногоподобных) отряда заурисхий. Это неуклюжее двуногое животное питалось растительностью, но происходило от совершенных двуногих динозавров среднего триасового периода, хищных теропод. В позднем триасе под влиянием похолодания и падения биопродуктивности среды прозауроподы стали возвращаться к четвероногому способу передвижения, свойственному далеким «холодолюбивым» предкам двуногих текодонтов и динозавров [37, с. 34–35]
Нижнеюрское похолодание с его давлением в пользу мобильности выщепило из среды архозавров (родственников динозавров) летающих ящеров птерозавров, покрытых шерстью. Они просуществовали до верхнего мела и разделили судьбу динозавровой биоты [25, с. 106–109]. Их вымирание объясняется тем, что, в отличие от сноровистых, высокоподвижных птиц современного типа, они были планирующими, энергосберегающими созданиями, а не активными летунами, т. е. характеризовались ограниченной мобильностью [25, с. 106; 87, с. 593], в то время как маастрихтское (конца мела) и палеоценовое (после мела) похолодания требовали от животных более сильно выраженной подвижности.

Рис. 5.
Барапазавр
(Barapasaurus tagorsi, большеногий ящер) из подотряда зауропод (ящероногов) группы зауроподоморфов отряда заурисхий. Под влиянием ухудшения климата и падения биопродуктивности среды неуклюжие двуногие прозауроподы породили в раннем юрском периоде совершенных четвероногих зауропод. Они процветали и достигли размеров, рекордных для наземных животных [37, с. 42–43]
В поздней юре (150,7–144,2 млн лет назад) максимум трансгрессии сменился регрессией и похолоданием, что возобновило селекцию на подвижность и побудило динозавров вторично породить птицеподобных существ – в частности, известного археоптерикса [240, с. 349], который строением зрительной и слуховой коры головного мозга действительно напоминал современных птиц [136], что явилось результатом конвергенции, параллельной эволюции. Будучи предком энантиорнитинов, или «противоположных птиц» (от последовательности совмещения костей стопы в ходе развития, противоположной современным птицам), археоптерикс напоминал миниатюрного велоцираптора или дромеозавра [697], тоже пернатых динозавров, и отличался от протоависа, предка орнитуринов, давших современных птиц. Динозавров, археоптерикса и энантиорнитинов объединяли примитивные признаки эктотермности, т. е. непостоянной температуры тела [660].
Меловой период (144,2–65 млн лет назад) ознаменовался нижнемеловой регрессией (век валанжин, 130 млн лет назад) и продолжением отбора на мобильность. В это время развивались упомянутые тупиковые энантиорнитины и перспективные орнитурины, в среде которых началась дивергенция современных птиц (средний нижний мел, век апт, 122,9 ± 0,3, 121,2 ± 0,3–121,1 ± 0,2 млн лет назад) [260; 747]. За тот же временной интервал пернатые динозавры породили еще одно птицеобразное создание – четырехкрылого Microraptor gui из Китая [829], все четыре конечности которого годились для планирования, повышая подвижность. Среди современников этого маленького дромеозавра – китайских птиц-энантиорнитинов – также встречался четырехкрылый вид [833], так что данная эволюционная тенденция проявлялась неоднократно.
Если зауроподы откликнулись на падение биопродуктивности среды мобильной четвероногостью, то орнитоподы, ящеротазовые четвероноги (игуанодоны) и двуноги (утконосые динозавры-гадрозавры, верхняя юра – верхний мел) отреагировали иначе: они освоили умение жевать, как млекопитающие [596], что повышало эффективность потребления редкой пищи соответственно похолоданию. В этом отношении названные представители динозаврового племени принципиально отличались от прочих архозавров, т. е. других динозавров, а также крокодилов. Последние, кстати, пережили роковой меловой период в силу того обстоятельства, что являлись ловкими наземными бегунами-четвероногами, т. е. мобильными животными, что открывало им путь в холодный кайнозой, где они, впрочем, вскоре «замерзли» и перешли к водному образу жизни в тропических и субтропических реках, не выдержав конкуренции с крепнущими наземными четвероногими млекопитающими.
Рубеж нижнего-верхнего мела (105 млн лет назад) прошел с трансгрессией, а века турон – коньяк (92–86 млн лет назад) подверглись вулканическому диоксид-углеродному потеплению [750]. В конце мела, в маастрихте (71–66 млн лет назад) регрессия сопровождалась заметным похолоданием [526], когда тропические температуры в океанах упали ниже современных [288]. К этому времени изнеженные теплым и обильным мезозоем (251–65 млн лет назад) динозавры находились на вершине своего расцвета, о чем свидетельствует сам факт существования крупнейшего наземного хищника – королевского тираннозавра (76–65 млн лет назад) (см. рис. 6). Сопряженная с ним мегафауна стала совершенной и высокоспециализированной, а потому могла лишь углубляться в аппетитах, но никак не ограничивать себя, чего требовала оскудевающая обстановка.
Вымирание динозавров происходило постепенно и пропорционально прогрессу млекопитающих (по модели цветковых растений и костистых рыб, см. далее). В веке кампан (76–73 млн лет назад) на пик расцвета динозавров приходилось 30 их родов. С приближением маастрихтского похолодания (71–66 млн лет назад) гордые животные ощутили эволюционно-экологическое затруднение: они любили много потреблять и мало двигаться, а теперь среда благоволила к сноровистым аскетам типа четвероногих млекопитающих.

Рис. 6.
Тираннозавр
(Tyrannosaurus rex, королевский ящер-тиран) из семейства тираннозаврид инфраотряда карнозавров (хищных ящеров) подотряда теропод отряда заурисхий. Это был крупнейший наземный хищник всех времен, чуть уступавший лишь хищному гигантозавру начала верхнего мела (ок. 100 млн лет назад), который, будучи южноамериканцем, закономерно превосходил массой североамериканского собрата, как свойственно рептилиям с приближением к экватору (у млекопитающих – наоборот). Тираннозавр был совершенным хищником (и падальщиком, как современные львы), узко приспособленным к среде, богатой пищей. С ухудшением климата и падением биопродуктивности среды королевский ящер угас и вымер вместе с подавляющим большинством остальных динозавров. По современным данным, он был оперенным существом [37, с. 98–99]
За весь маастрихт и две трети дания (72–65,3 млн лет назад) число родов динозавров упало до 12. Затем вымирание ускорилось, и за период 65,3–64, 96 млн лет назад прошло пять убывающих стадий: 12–12–11–10–7 родов уходящих динозавров. О том, что биосфера отреагировала на коварный удар иридиевого астероида 65 млн лет назад, не может быть и речи. Параллельно со ступенчатым падением динозавров шел уверенный прогресс копытных млекопитающих, представленных в тех же временных точках 0–1 – 3–5 – 8 родами. Так что за 40 тыс. лет, истекших с момента падения астероида, млекопитающие зримо окрепли, а динозавры еще не вымерли окончательно, являя редкий случай послемеловых, палеоценовых динозавров, останки которых обнаружены выше мелтретичной иридиевой аномалии, оставленной упавшим болидом [714; 713]; cp. [222; 642; 696].
Приведенные количественные данные по динозаврам и копытным млекопитающим за период 65,3–64,96 млн лет назад происходят из формации Хелл Крик в Монтане, США (северо-запад страны). Монтана лежит на том же континенте, что и мексиканский кратер Чикхулуб, оставленный ударным небесным телом 65 млн лет назад. Монтанская биота совершенно не отреагировала на это событие: млекопитающие набирали эволюционные обороты, а динозавры сокращались в разнообразии в соответствии с тенденцией, наметившейся за 7 млн лет до вторжения астероида. Не избежали их судьбы и обитатели других сред.
Морские пресмыкающиеся (ихтиозавры, плезиозавры, мозозавры и пр.), страдавшие низкой мобильностью [35, с. 205–206], вымерли в верхнем меле: более подвижные ихтиозавры – в его теплом начале, остальные – ближе к холодному концу [25, с. 18, 33]. Господствовавших на море раковинных головоногих моллюсков аммонитов и белемнитов постигла та же участь.
Показательна в этой связи эволюционная история теплолюбивых аммоноидей (нижний девон – верхний мел) [83, с. 243–425; 84, 15–144]. Появившись в начале раннего девона (век жедин, 408–401 млн лет назад), они продолжили линию других головоногих моллюсков – бактритов, имевших прямую или слабосогнутую раковину, т. е. являвшихся более-менее хорошими пловцами. Однако «возниковение аммоноидей было связано с образованием вначале согнутой, а затем криоконовой раковины, состоявшей из несоприкасающихся оборотов, свернутых в одной плоскости» [38, с. 376]. Это делало аммоноидей слабыми пловцами и приводило в соответствие с теплым, высокобиопродуктивным девоном, когда была выгодна умеренная мобильность.
В эпоху пермо-карбонового оледенения ситуация изменилась, и малоподвижные аммоноидеи закономерно отступили в эволюционную тень, поскольку оледенение благоприятствовало более энергичным созданиям. Зато теплый мезозой востребовал их качества, и в кампане – маастрихте (76–66 млн лет назад) они выдвинули род Pachydiscus, у представителей которого свернутая раковина достигала рекордных 2 м в диаметре [84, с. 111]. Напомним, что скоростное плавание обеспечивала прямая раковина (см. выше об эндоцератитах). Свернутая же в диск раковина, наоборот, скоростей не давала. При этом под влиянием климатических колебаний в меле аммониты то пытались слепо распрямить раковину, чтобы повысить свою мобильность и попасть в унисон с похолоданиями, то сворачивали ее обратно, чтобы убавить подвижность и совпасть с потеплениями. В силу закона Долло о необратимости эволюции эти попытки были обречены на провал и лишь наводняли меловые моря уродливыми созданиями.
Избирательность верхнемелового вымирания прослеживается в судьбе двустворчатых моллюсков. Их прикрепленные формы вымирали заметно интенсивнее, нежели свободноживущие [434]. Это объясняется тем, что прохладное дыхание наступающего кайнозоя готовило перспективу лишь подвижным существам – в данном случае свободноживущим двустворчатым моллюскам.
С мезозойскими воздухоплавателями происходило то же самое. Отряды подвижных современных птиц дивергировали еще при похолодании в раннем меле, и 22 их филетические линии успешно пересекли мелтретичную границу, не побоявшись зловещего астероида [260]; ср. [252]. Однако сам факт биологической принадлежности к современным птицам отнюдь ничего не значил. В частности, могучие (более метра в высоту) нелетающие гесперорнисы были водоплавающими птицами и в силу своей маломобильности не перенесли верхнемелового отбора. Маломобильным планирующим птерозаврам тоже не повезло, равно как и «противоположным птицам» энантиорнитинам, которые летали хуже тогдашних орнитуринов, предков неорнитов, нынешних птиц [697].
Между тем, финал мелового периода не был эпохой тотального вымирания [35, с. 89, рис. 3; 184; 239; 394; 407; 674, с. 439–441]; cp. [444]. Эволюционная история меловой биоты выглядела как пара сообщающихся сосудов, в одной из емкостей которых убывали многочисленные маломобильные господа мезозоя (динозавры, птерозавры, ихтиозавры, плезиозавры, аммониты, белемниты и др.), а в другой прибывали скромные высокомобильные господа будущего кайнозоя (покрытосеменные растения, млекопитающие, современные птицы, костистые рыбы, акулы, внутреннераковинные головоногие моллюски и др.).
Будучи родом из пермо-карбонового оледенения (см. выше, а также [284; 407; 638]), цветковые растения положительно отреагировали на ухудшающуюся климатическую обстановку нижнего мела и в середине мела (кон. века альба, 105 млн лет назад) произвели революцию флоры. С верхнего мела к ним перешло флористическое господство, которое укрепилось в кайнозое и наиболее ярко представлено сейчас [88, с. 423].
Через своих рептильных звероподобных предков млекопитающие тоже ведут начало из пермо-карбонового оледенения. Оттуда происходит их четвероногая мобильность и эффективная разнозубость, выгодные в условиях скудной биопродуктивности среды. Как класс, они выщепились в холодное время норий-рэтийской регрессии. В поздней юре или раннем меле из их пула выделились однопроходные звери (ок. 144,2 млн лет назад) [149; 466]. По молекулярно-генетическим данным, то же самое произошло с сумчатыми млекопитающими еще раньше (в средней юре, 173 ± 12,3 млн лет назад) [479], хотя палеонтологически они известны лишь с позднего мела (84–65 млн лет назад) [183]. По молекулярно-генетическим данным, пять линий высших плацентарных млекопитающих обособились св. 100 млн лет назад, т. е. в начале верхнего мела (в веке сеномане, 105–92 млн лет назад), и большинство их современных отрядов появилось до мелтретичной границы; палеонтологически плацентарные млекопитающие отмечены 85 млн лет назад [479]; а также [651; 655, с. 187–310]. Радиация сумчатых и плацентарных млекопитающих связана с теплыми периодами юры и мела, когда подавляющие их четвероногие динозавры испытывали эволюционную депрессию, и вакансии в соответствующих эконишах доставались млекопитающим. Последние были преимущественно мелкими формами с черно-белым зрением, поскольку вели скрытный, ночной образ жизни. Однако среди них встречались и более выразительные представители. В раннем меле Китая, 139–128 млн лет назад, существовал хищник из числа млекопитающих Repenomomus giganticus, достигавший свыше метра в длину и 12–14 кг массы, что позволяло ему успешно охотиться на детенышей динозавров (в частности, на пситтакозавров, один из которых сохранился у него в брюхе [425]). Млекопитающие, подвижные зверьки, настойчиво прогрессировали и, подобно птицам, уверенно пересекли мел-палеоценовую границу.
Параллельно цветковым растениям, млекопитающим и птицам меловую экосреду завоевывали костистые рыбы. Являясь отпрысками лучеперых рыб, предки которых зародились в период ордовикского оледенения, костистые пловцы были сильны в прохладной, малопродуктивной среде, поскольку лучше других рыбообразных создавали осевым скелетом двигательную локомоторную волну [124]. Недаром их современный рекордсмен, меч-рыба, развивает огромную скорость до 130 км/ч, превосходя по этой части наземного гепарда. Непреодолимо прогрессируя с раннемелового похолодания, они возобладали в морях с верхнего мела, легко пересекли мелтретичную границу и ныне составляют 95% ихтиофауны [86, с. 334; 655, с. 61–71]. Эта победа облегчалась малой мобильностью прочих мезозойских рыб [35, с. 205]. Эволюционная история акул, также имеющих ордовикские корни и отличающихся высокими ходовыми качествами, развивалась по образцу костистых рыб [86, с. 196–237] и тоже отмечена рекордами в кайнозое (см. ниже). Наконец, безуспешная борьба за жизнь медлительных аммонитов и белемнитов закончилась вытеснением их подвижными внутреннераковинными головоногими моллюсками: десятиногими кальмарами и каракатицами, а также и восьминогими осьминогами [35, с. 187; 84, с. 145–178].
С падением общеглобальной биопродуктивности среды разнообразие жизни на планете убывает, и проявления эволюционной направленности теряют разнообразие. Масштабы организмов тоже мельчают. Поэтому господствующие наземные животные мезозоя были крупнее кайнозойских, а раннекайнозойские – внушительнее позднекайнозойских современников нарастающего похолодания. Соответственно, примеры нашей версии эволюции для кайнозоя относительно скромны.
Кайнозой (65-0 млн лет назад)
В палеоцене (65–55 млн лет назад) высокомобильные организмы мелового периода продолжают разнообразиться, не меняя своей эволюционной направленности, поскольку вся эпоха отмечена регрессией и похолоданием. В начале эоцена (55–36 млн лет назад) началось метановое потепление [742], и регрессия сменилась трансгрессией. Следовало бы ожидать, что в установившихся теплых условиях появятся маломобильные формы мезозойского типа.
В соответствии с этим ожиданием в среднем эоцене распространяются уцелевшие потомки вымерших меловых белемнитов – байанотеутиды [35, с. 189–190; 38, с. 382, 388, рис. 217; 84, с. 161–162]. Тогда же возникают не связанные общностью происхождения нелетающие, бегающие птицы Ratidae, своего рода конвергенты (т. е. внешние подобия) двуногих динозавров: страусоподобные элеутерорнисы (средний эоцен, век лютет) и эпиорнисовые псамморнисы (эоцен) [35, с. 245–246]. Наконец, в раннем среднем эоцене Германии (50–48 млн лет назад) обнаруживается двуногое бегающее млекопитающее Leptictidium nasutum, подлинный конвергент двуногих динозавров со свойственным им и людям прямохождением (см. рис. 7) [118, с. 34]. Эоценовое потепление повысило биопродуктивность среды и раздвинуло рамки экониш, привлекая в них новых обитателей. В соответствии с этой новацией сухопутные парнокопытные произвели водных китообразных в нижне-среднем эоцене (ок. 52 млн лет назад [752]) или в конце среднего эоцена (в лютете, 47 млн лет назад [354]). В гармонии со своим происхождением в теплую эпоху китообразные (зубатые киты) развили чуждую млекопитающим равнозубость динозаврового типа.
В олигоцене (36–24 млн лет назад) трансгрессия прекращается и сменяется регрессией во второй половине эпохи. В связи с похолоданием высокомобильные четвероногие млекопитающие испытывают эволюционный подъем и порождают мегафауну, украшением которой является гигантский безрогий носорог Indricotherium transouralicum – самое крупное и высокое наземное млекопитающее, до 5 м высотой (средний олигоцен).

Рис. 7.
Легкобег носатый
(Leptictidium nasutum) возник в эпоху раннего среднего эоцена на островах нынешней Германии 50–48 млн лет назад, когда из-за потепления поднялась биопродуктивность среды, и естественный отбор дал шанс появлению маломобильных двуногих животных. «Носач» бегал на двух ногах, как двуногие динозавры и люди, а потому являлся их эволюционным конвергентом (подобием), таким же «теплолюбивым», как и они [118, с. 34]
В миоцене (24–5,3 млн лет назад) ранне-среднемиоценовое потепление (24–13 млн лет назад) положило конец олигоценовой мегафауне. Зато пробившийся отбор на маломобильность вызвал к жизни двуногую обезьяну «перелоапитека каталонского» (Испания, 13 млн лет назад), положившего начало целой плеяде двуногих гоминин (см. разд. 2. 4). Происходя от аборигенов высокобиопродуктивных тропических лесов, они органично несли в себе мезозойскую ориентацию на маломобильность и прожорливость, что в сочетании с эволюционной отсталостью создало гоминин, перспективных господ будущего, ставших лидерами именно в силу своей отсталости.
В плиоцене (5,3–1,6 млн лет назад) начало эпохи ознаменовалось трансгрессией. Затем последовал ряд гляциалов (оледенений), интергляциалов (межледниковий), стадиалов (пиков оледенений) и интерстадиалов (смягчений оледенений) выраженной ледниковой эпохи. Ее «лицом» можно назвать тридцатиметровую белую акулу Carcharodon megalodon, вымерщую в среднем плиоцене (3,3 млн лет назад), когда интергляциальная трансгрессия Астий вызвала ограниченное вымирание в среде мобильной биоты; cp. [680].
Наконец, в плейстоцене (1,6–0,0117 млн лет назад) развилась богатая приледниковая мамонтовая фауна, вымершая с началом голоценового потепления 11700 календарных лет назад, или 10200 лет назад по радиоуглероду 14С (поясним, что вследствие колебаний содержания тяжелого изотопа углерода 14С в древней атмосфере радиоуглеродные даты исследуемых образцов оказываются моложе своего реального, календарного возраста, который можно установить, например, методом дендрохронологии, когда высчитываются годичные кольца очень старых деревьев, до 8000 лет возраста, после чего итог сравнивается с радиоуглеродной оценкой; получается, например, что календарной дате 4650 лет назад отвечает радиоуглеродная дата 4050 лет назад).
Подобно динозаврам, мамонты и их спутники вымерли не сразу. Некоторые виды жили дольше: островные карликовые мамонты острова Сен-Пол в Беринговом море, 7908 ± 100 лет назад [391], островов Санта-Барбара в Калифорнии [480, с. 213], острова Врангеля на границе Восточно-Сибирского и Чукотского морей, 7620–3730 лет назад [515; 768]; большерогий олень в Западной Сибири, 6900 14C, или 7700 календарных лет назад [736]; возможно, шерстистый носорог в Чувашии, 921/922 гг. н. э. [54, с. 61–66, 139–140; 91, с. 72–74].
«Лицом» нынешней голоценовой эпохи – межледниковой, однако все еще не свободной ото льдов – можно считать гигантского кальмара Architeuthis longimana 19 м длиной (возможны тридцатиметровые особи), продолжающего линию внутреннераковинных головоногих моллюсков, которые сменили аммонитов в конце мелового периода.
Относительно упомянутых вымираний существуют неокатастрофические взгляды, касающиеся в первую очередь пермо-триасовых и мел-третичных событий. Согласно одному из них, массовые вымирания вызваны вулканической деятельностью планеты, которая обескислороживает океаны, отравляя их. Эти несчастья происходят с периодичностью в ~ 32 млн лет [632]. Они ответственны за массовую гибель биосферы на границе перми и триаса [227; 637; 639], а также мела и палеоцена [227; 598]; cp. [361, с. 368].
Показано, однако, что обескислороживание океанов господствовало на Земле на протяжении 250 млн лет в позднем неопротерозое ок. 800–543 млн лет назад [471; 802], однако на тогдашней жизни (например на морской эдиакарской фауне) это заметно не сказалось. Показано также, что обескислороживание океанов в самом начале триаса явилось следствием предшествующего позднепермского вымирания (кризиса радиолярий, морских микроорганизмов) [633], а никак не его причиной. Наконец, показано, что обескислороживание океанов имело место и в конце нижнего мела (в альбе, ок. 112 млн лет назад) [481], что не помешало костистым рыбам, акулам и прочим наследникам мезозоя уверенно набирать эволюционные обороты. В свете указанных расхождений с реальностью вулканическая гипотеза не может быть принята.
Согласно другим представлениям, Землю бомбардируют космические тела (астероиды и кометы), удары которых поднимают в атмосферу взрывной материал, затеняют им планету и создают эффект «ядерной зимы», которая умерщвляет биосферу. О падениях небесных объектов говорят геологические прослойки редкого металла иридия, встречающегося в метеоритном веществе, как, впрочем, и в земных породах, изверженных из мантии в результате вулканической деятельности. Последнее обстоятельство заставляет исключить ископаемую иридиевую аномалию, например ордовик-силурийской границы, из числа космогенных [804]. В разведанной истории Земли насчитывают до 140 болидовых ударов [347]; cp. [361, с. 368]. Среди них наиболее известны следующие:
• поздний плиоцен, ок. 2,15 млн лет назад [347] или ок. 2,3 млн лет назад [482] (астероид св. 1 км в поперечнике);
• средний плиоцен, 3,3 млн лет назад [680];
• поздний эоцен, 35,7 ± 0,2 млн лет назад [199];
• ранний эоцен, ок. 50 млн лет назад [347];
• мелтретичный, 65 млн лет назад [137; 138; 192; 195; 378; 415; 433; 461–463; 477; 554; 558; 561; 563; 649; 743; 745]; болид вызвал глобальные лесные пожары [414; 771];
• поздняя юра, 140 ± 20 млн лет назад [378];
• триас-юрский [603];
• поздний триас, 211 (220–200) млн лет назад [518, с. 46]; 5 кратеров, век норий, 214 млн лет назад [721; 786].
• пермо-триасовый [175]; (251,4 млн лет назад) [180]; cp. [311].
• верхний девон, рубеж веков фран-фамен, 367 млн лет назад [247].
• средний девон, ок. 380 млн лет назад [305].
• поздний докембрий, ок. 600 млн лет назад [365; 366; 806].
• ранний протерозой, 1,85 млрд лет назад [179; 588].
• верхний ранний архей, 3,470 ± 0,002 млрд лет назад [226].
В настоящее время доказано, что после триас-юрской иридиевой аномалии, свидетельствующей о падении болида, тогдашние динозавры отнюдь не вымерли, а, наоборот, расцвели менее чем через 10 тыс. лет после грозного события [603]. Выше мы привели факты, указывающие на то, что позднепермское вымирание состояло из двух крупных стадий, разделенных 5 млн лет, так что его космический источник нереален. Приведены также факты, рисующие позднемеловое вымирание как избирательное событие в духе дарвиновской теории естественного отбора – его космический источник излишен. Кроме того, хорошо известно, что с ранней юры до позднего мела холодные высокие широты Земли населялись полярными динозаврами. Они известны из ранней юры (век плинсбах) Антарктиды [397], из раннего мела (века валанжин – альб, 130–105 млн лет назад) тогдашней заполярной Австралии [644; 142], из позднего мела (века поздний кампан – ранний маастрихт, 76–66 млн лет назад) тогдашней заполярной Аляски [215]. Хорошо приспособленные к продолжительному холоду и тьме полярной ночи, они бы встретили «ядерную зиму» с энтузиазмом. Приведенные соображения делают биологическую сторону космической гипотезы неприемлемой.
Наконец, существует убеждение, что массовые вымирания последних 50 тыс. лет вызваны охотничьим нажимом первобытного человека. Так были истреблены гигантские нелетающие птицы моа Новой Зеландии 980 ± 40–900 ± 45 лет назад [421; 480, с. 222], предположительно мамонтовая приледниковая фауна Старого Света и вся мегафауна Нового Света на рубеже плейстоцена – голоцена 11700 календарных лет назад [480, с. 221–222; 735], мегафауна Австралии 50 ± 5 тыс. лет назад в результате вторжения людей 55 ± 5 тыс. лет назад [570].
Доказано, однако, что мегафауна Австралии вымерла ок. 46,4 (51,2–39,8) тыс. лет назад из-за крайней аридности (засушливости), вызванной гляциальным максимумом [647], т. е. холодным стадиалом Вюрм II C (46,5–39 тыс. лет назад), когда вместе с похолоданием резко обострилась сухость земного климата, поскольку масса воды, в том числе атмосферной влаги, была скована в обширных ледниках.
Кроме того, обнаружено, что Новый Свет был заселен ранее 33 тыс. лет назад, о чем свидетельствуют остатки первобытной культуры в Монте Верде (Чили, 33370 ± 530 лет назад) и Тока ду Бокейрану да Педра Фурада (Бразилия, св. 32 тыс. лет назад) [385; 500, с. 708], в силу чего заселение Америки не может быть увязано с вымиранием ее мегафауны 11 700 лет назад.
Указанные обстоятельства не позволяют принять антропогенную гипотезу массовых вымираний, т. е. древних массовых вымираний, вызванных человеком (за вычетом новозеландского эпизода с птицами моа, который, впрочем, не относится к числу примеров массовых вымираний). Следует добавить, что вулканическая, космическая и антропогенная гипотезы массовых вымираний полностью оторваны от закономерностей развития земной жизни и не проливают здесь никакого света. Вдобавок людям, в том числе и ученым, вообще свойственно переоценивать значение катастроф в истории природы и общества и хвататься за них, как за соломинку, когда что-либо не сразу поддается объяснению (к сказанному добавим неокатастрофическое объяснение происхождения самосознания Дж. Джейнсом, см. разд. 3.2). Между тем, реальное значение катастроф не так фатально. Например, катастрофическое извержение вулкана Кракатау (Индонезия, 1883 г., погибло 36 тыс. человек) или удар Тунгусского метеорита (Красноярский край, Россия, 1908 г., произошел вывал леса в радиусе 15–30 км вокруг эпицентра) не оказали на развитие биосферы никакого влияния.
В изложенной нами версии эволюции биосферы под влиянием эколого-климатического воздействия суперконтинентального цикла бросается в глаза то, что его крупномасштабная периодичность не объясняет более дробных колебаний климата. В реальности суперконтинентальный цикл создает лишь общие условия для развития оледенений или межледниковий. Их конкретные проявления подчиняются влиянию частных факторов астрономического, атмосферного, гидросферного характера, хронометраж которых пока не разработан [70, с. 20; 560; 635; 742; 750], если не считать климатических последствий от колебаний в орбитальном движении Земли с периодами в 25, 44, 100, 133 и 400 тыс. лет [602]. Хронометраж заметных вымираний приводит к выводу, что их пики отстоят друг от друга на 20–60 млн лет [184]: на 26 млн лет за последние 150 млн лет [239, с. 50; 361, с. 363; 410] или на 27 млн лет за последние 250 млн лет (8–12 пиков) [634]. Комментарии этих наблюдений пока преждевременны.
Если принять нашу версию направленной эволюции, то встает вопрос, как она согласуется с дарвиновской моделью случайного хода эволюции и каков генетический механизм направленной эволюции. Мы отвечаем на него следующим образом.
Земная биосфера испытывала климато-экологическое давление среды на протяжении более чем 3,85 млрд лет, и это давление как таковое являлось одним из внешних условий, к которым живые существа приспосабливались в результате случайного естественного отбора. Следовательно, их геномы давно должны были «обзавестись» наследственным механизмом ожидания климато-экологических последствий суперконтинентального цикла, т. е. фактически усвоить склонность к эволюционной направленности естественным путем. В сущности, мы остаемся в рамках дарвинизма, хотя это учение избегало представлений об эволюционной направленности.
Геномы организмов слепы: они лишены органов чувств. Между тем, уже в течение нескольких миллиардов лет незрячие геномы с успехом противостоят изменчивой среде, все больше совершенствуются и, похоже, умеют предвидеть изменения в природе. Как же им это удается? Ответ здесь может быть один. Слепые геномы ориентируются в окружающем мире на основе принципов теории игр.
Разумеется, крохотные геномы не сильны в теории. Однако практическая жизнь и безжалостный естественный отбор сохранили в их рядах только те образцы, которые действуют так, словно и впрямь знакомы с Дж. фон Нейманом (1903–1957, основоположником теории игр). Для разъяснения их «талантов» нам не понадобится высшая математика. Допустим, вы втянуты в азартную игру, а глаза у вас завязаны. Как вы построите свое поведение? По-видимому, пойдете с крупной карты. А если не получится, то с мелкой. Если не выйдет, ограничитесь чем-то средним. И будете так экспериментировать, пока не зацепитесь за удачу. Геномы ведут себя точно так же.
Они находятся в состоянии постоянного мутагенеза, т. е. подвергают свои гены тем или иным мутациям, происходящим с примерно одинаковыми скоростями (ср., однако, разд. 2.4). Вопреки бытующему убеждению, результаты этого процесса далеко не произвольны, не случайны. Дело в том, что генные мутации делятся на серии характерных типов: гиперморфные (они увеличивают проявление подчиненного себе признака, фена), гипоморфные (они уменьшают проявление признака), антиморфные (признак уничтожается вообще) и неоморфные (признак дополняется) [7, с. 42–49]. Иными словами, мутагенез носит количественный характер (как и ваше «слепое» поведение в мысленном эксперименте с азартной игрой). Изменения блоков генов и целых хромосом также подчиняются количественному принципу: они дуплицируются, т. е. удваиваются, умножаются, как и отдельные гены [82; 468; 522; 523; 613].
Что такой мутагенез может дать геному и его организму-хозяину? Уверенность в завтрашнем дне. Допустим, грядет похолодание. Организмы слепо, механически мутируют обрисованным количественным способом. Треть из них немотивированно мутирует гипоморфно и утрачивает, например, запасы жира – они замерзнут. Другая треть вообще воздерживается от мутаций и прозябает. Зато оставшаяся треть, не подозревая о своей гениальности, опять же слепо, механически предается гиперморфным мутациям и ни с того ни с сего наращивает подкожный слой жира. Приходит холод, эта треть не мерзнет и овладевает миром. Она побеждает автоматически, в силу законов теории игр. На Земле уцелели лишь те организмы, которые неукоснительно следовали описанному способу количественного мутагенеза. Он позволял слепо предугадывать климато-экологические последствия суперконтинентального цикла и гарантировал жизнь определенной части участников событий. Другая часть так же гарантированно «не попадала» в ход событий и беспомощно вымирала.
При таком положении дел в эволюции мы могли бы ожидать, что определенный процент организмов в истории жизни будет вымирать систематически и внешне беспричинно – в силу того, что эти организмы не угадали правильный ход в мутагенезе. Но так ли это? Это так. Палеонтологи действительно обнаружили в ископаемой летописи загадочную череду автоматически текущих вымираний, которую они окрестили Законом Постоянного Вымирания, или гипотезой Красной Королевы (персонажа из «Алисы в стране чудес» Л. Кэрролла) [182]. В свете вышеизложенного мы начинаем понимать, откуда в палеонтологической летописи взялся след Красной Королевы.
Теперь посмотрим, что дает версия количественного мутагенеза для понимания узловых моментов истории жизни. Можно было бы ждать, что организмы, угадавшие грядущие перемены в экосреде, окажутся приспособленными к ней наиболее точно и займут в ее ландшафте господствующие высоты. Такие приспособленные организмы называются специализированными. Их судьба незавидна. Углубившись в «подгонку» себя к господствующей среде, холодной (как зверозубые пресмыкающиеся) или теплой (как динозавры), они сожгут за собой эволюционные мосты и с переменой вектора развития среды (с похолодания на потепление или наоборот) окажутся беспомощными и вымрут (как угасли зверозубые рептилии и динозавры).
«Отыграть эволюцию назад» им не позволит закон Долло о необратимости эволюции, а эволюция необратима потому, что не является прямолинейной, и угадать в ней извилистые пути отступления в полном соответствии с путями наступления – нереально. Например, мы помним, что с похолоданием в ранней юре двуногие прозауроподы встали на четвереньки и сделались могучими зауроподами. Но переделать свою прожорливую мезозойскую природу они не смогли и с необратимым похолоданием в конце мела сошли с эволюционной сцены. То есть они сумели отыграть эволюцию назад лишь отчасти и поплатились за это местом под солнцем.
Напротив, организмы, воздержавшиеся от углубленного приспособления к наличной среде, не пережившие подходящих количественных мутаций, прозябают в ее условиях на экологических задворках. Такими были жалкие четвероногие млекопитающие на фоне яркого мезозоя. Подобных живых существ называют генерализованными, и их участь, вопреки первому впечатлению, завидна. Когда среда меняется, им не приходится искать длинных путей попятной эволюции, поскольку они не проходили пространных путей поступательного развития. В результате они относительно легко приспосабливаются к новым условиям, довольно быстро становятся новыми господами планеты и из разряда генерализованных организмов перекочевывают в пул специализированных существ. Такое превращение произошло, как мы показали выше, с тусклыми млекопитающими на протяжении прохладного палеоцена, в начале которого канули в Лету последние динозавры.
Сенсационная судьба этих колоссов объясняется их адаптацией. Вообще массовые вымирания не более невероятны, нежели мелкие [165]. Но вымирание динозавровой фауны закономерно перекрыло все допуски. Дело в том, что преобладающие организмы мезозоя были приспособлены к весьма высокой биопродуктивности среды, и ее падение в конце мелового периода смело господствующие династии животных подчистую, наводя ученых на мысли о вулканической или космической катастрофе. Напротив, звероподобные пресмыкающиеся времен пермо-карбонового оледенения ориентировались на скромный уровень биопродуктивности среды. Ее подъем в триасовом периоде сделал адаптацию зверообразных пресмыкающихся невыгодной, но не лишил их привычно обильной пищи, как это произошло с динозаврами в финале их бытия. Поэтому зверообразные пресмыкающиеся частью вымерли, частью ушли в эволюционную тень без обвального вымирания.
Это обстоятельство уберегло наших предков млекопитающих от кризиса в теплом мезозое. Они даже усовершенствовались по части повышения мобильности. Можно спросить, куда же дальше, если они и без того были четвероногими? Ответ на этот вопрос таков. Одной из форм повышения мобильности животных является живорождение, поскольку оно не привязывает животных к месту кладки яиц. Во время потепления средней юры четвероногие динозавры претерпели кризис и освободили некоторые экониши, в которые устремились сумчатые млекопитающие. Они были примитивно живородящими, а потому более подвижными по сравнению с собратьями по классу. То же самое произошло с более совершенными по живорождению плацентарными млекопитающими, что в нынешнее кайнозойское время вывело их на первые биологические роли.
Таким образом, можно убедиться, что наша версия стихийно направленной эволюции не состоит в противоречии с данными палеонтологии и эволюционной биологии. Достоинством этой версии мы считаем последовательное подчинение узловых эволюционных событий единообразной причинности, однотипным принципам развития. Нам кажется весьма существенным, что в описанной системе представлений предки людей – ископаемые гоминины – закономерно находят свое место, что проливает свет на стартовые условия антропогенеза.
2.4. Гоминизация
Судя по молекулярно-генетическим данным, приматы вместе с ближайшими древесными родичами, предками лесных планеристов шерстокрылов [438], отделились от прочих плацентарных млекопитающих в начале кампана (верхний мел), ок. 75 млн лет назад [809, с. 1090, табл. 1]. Под влиянием похолодания в середине эпохи палеоцена, ок. 60 млн лет назад, из состава приматов выщепились мобильные, летающие формы – крыланы, или летучие лисицы [363; 619], а древолазающие формы распались на полуобезьян (тупайи, долгопяты, лемуры) и антропоидов (обезьян) [670, 112, табл. 6–7]. Последние с раннего эоцена (55–50,5 млн лет назад) обнаруживаются в палеонтологической летописи Северной Африки [178; 543; 544; 709] и Юго-Восточной Азии [236; 245; 436]. На рубеже эоцена и олигоцена от узконосых обезьян Старого Света отошли широконосые обезьяны Нового Света, 36 ± 3 млн лет назад. Наконец, в начале миоцена, 22 ± 2 млн лет назад, узконосые обезьяны разделились на низших, церкопитекоидных (павианы, макаки и т. п.), и человекоподобных, гоминоидов (гиббоны, орангутаны, гориллы, шимпанзе, люди).
Вопреки расхожему убеждению, обезьяны не являются лидерами эволюционного процесса среди млекопитающих. Они – жертвы так называемого биогенетического закона, согласно которому темпы индивидуального развития животного (онтогенеза) задают скорость эволюции (филогенеза) всего вида, к которому принадлежит это животное. Причина подобной зависимости очевидна. Если индивидуальная биография особей замедлена, поколения их вида сменяются тоже медленно, и их становится меньше, чем у более активно развивающихся животных. А единицей эволюционного развития является именно поколение. Чем поколений меньше, тем медленнее протекает эволюция вида.
Из-за жизни на деревьях, где нелегко воспитывать многочисленное потомство, обезьяны были ограничены в количестве одновременно рождаемых детенышей [12, с. 50–51]. Избавленные от конкуренции между собой, малыши отличались замедленным индивидуальным развитием (онтогенезом), что в силу биогенетического закона тормозило филогенез приматов, обусловливая их эволюционную отсталость. Ее признаки – цветное зрение, как у земноводных и рептилий, обильные кожные железы людей, как у земноводных, а также встречающаяся у некоторых людей шестипалость, воспроизводящая аналогичную особенность самых первых земноводных (см. разд. 2.3). У других млекопитающих подобные анцестральные, предковые черты отсутствуют. Речь, разумеется, не идет о том, что приматы вообще и в особенности люди прямиком унаследовали и сохранили перечисленные особенности амфибий. Суть дела состоит в другом.
Рождая малое количество детенышей, древесные приматы затормозились в эволюционном развитии, в том числе в молекулярно-генетическом, в эволюции генов, хромосом, геномов. У людей в силу неотении эта эволюционная заторможенность усугубилась (см. ниже). В результате наш геном сохранил застойный облик – стоит его «толкнуть» и вызвать у него так называемые «обратные мутации» (т. е. изменения, восстанавливающие предковый облик отдельных генов) [82, с. 69–70], как появятся гены настолько древние, «земноводного облика», что среди млекопитающих, обладающих лишь черно-белым ночным зрением, появятся существа со зрением цветного типа, т. е. приматы. У некоторых из них вдобавок обнаружатся обильные кожные железы земноводных и даже (иногда) свойственная их ранним представителям шестипалость – это мы находим у людей, которые, таким образом, оказываются еще застойнее, нежели другие приматы, что эволюционно выгодно (парадокс направленной количественной эволюции, см. разд 2.3).
В силу этого эволюционного аутсайдерства приматы имели предпочтительные шансы успешно приспосабливаться к изменениям среды (в кайнозое – к потеплениям, губительным для продвинутых млекопитающих). Напомним, что, согласно современным представлениям, «примитивные наследуют Землю» (слоган основоположника эволюционной генетики С. Оно [82, с. 174]), что обещало нашим обезьяньим предкам нелегкую, но перспективную судьбу. Указанные диалектические обстоятельства заведомо делали эволюцию приматов запутанной. В результате ни среди африканских гоминоидов (человекоподобных обезьян) [144, с. 641; 344; 350; 790], ни среди eвроазиатских дриопитеков (их родни) [144, с. 642, 643; 383, с. 69–70] не найдено надежных предков гоминин. Это привело к афро-евроазиатско-африканской гипотезе происхождения искомых предков человека [726], которая в настоящее время видится так.
С началом ранне-среднемиоценового потепления, 22–13 млн лет назад, предки современных гоминоидов в предвкушении открывающихся на севере экониш (под влиянием тепла) мигрировали из Африки в Евразию, что вызвало расхождение церкопитекоидов (мартышек) и гоминоидов (предков людей). С началом похолодания среднего миоцена – раннего верхнего миоцена, 13–10 млн лет назад, исконные африканские гоминоиды (проконсулы, афропитеки, кениапитеки и т. п.) вымерли, а сородичи двуногого перелоапитека (Испания, 13 млн лет назад) вернулись из Евразии в Африку, где положили начало гоминидам (гориллам, шимпанзе, предкам людей и людям согласно систематике [383, с. 66–70], в которой люди отнесены к биологическому отряду приматов, семейству гоминид, трибе гоминин). Состав двуногих предков человека и их родственников, гоминин [131, с. 526, рис. 1; 231, с. 850, рис. 1; 144, с. 644; 383, с. 70; 821; 822], приводим ниже. Обратим внимание на то, что молекулярно-генетические даты для гоминин не всегда согласуются с палеоантропологическими датами. Они приводятся под литерой «М». Перерассчитанные с «неотеническим коэффициентом» 1,894 (мотивацию см. ниже), они даны с литерой «Н».
1. Pereloapithecus cataluniensis («обезьяна из Перелоа каталонская»). Перелоа, Каталония, Испания, 13 млн лет назад.
2. Нгорора, Баринго, Кения, 11,94–9,82 млн лет назад (ближе к нижнему пределу), 1 верхний моляр (коренной зуб) гоминида [191; 494, с. 174; 500, с. 109]. Расхождение человека с большими африканскими человекообразными обезьянами: 6,25 ± 5,75 млн лет назад (М) [143; 196; 289; 403; 445; 476, с. 25; 506; 517; 646; 670; 726; 756; 801; 809, с. 1092], или 11,84 млн лет назад (Н).
3. Sahelanthropus tchadensis («сахелец чадский»). Торос-Меналла, пустыня Джураб, северный Чад, 7–6 млн лет назад [220; 221; 775]; cp. [814]. Строение основания черепа позволяет предполагать у «сахельца» прямохождение [839].
4. Orrorin tugenensis («первобытный человек тугенский»). Лукейно, Туген Хиллс, запад Баринго, Кения, 6,2 ± 0,19–5,625 ± 0,5 млн лет назад [494, с. 174; 623; 687]. Табарин, Туген Хиллс, запад Баринго, Кения, 4,96 ± 0,03 млн лет назад [416] (?).
5. Ardipithecus ramidus kadabba («базисная обезьяна-корень патриарх»). Аса Кома, средний Аваш, Афар, Эфиопия, 5,77 ± 0,08–5,54 млн лет назад [393; 811].
6. Ardipithecus ramidus («базисная обезьяна-корень»). Ас Дума, Гона, Афар, Эфиопия, 4,51–4,32 млн лет назад [686]. Арамис, средний Аваш, Афар, Эфиопия, 4,387 ± 0,031–4,29 млн лет назад [797; 798; 812; 813]; cp. [145, с. 556; 687] (предок шимпанзе?).
7. Australopithecus africanus («южная обезьяна африканская»). Штеркфонтейн, Йоханнесбург, Трансвааль, ЮАР, 4,17 ± 0,14(0,35) – ок. 2,0 млн лет назад [607].
8. Australopithecus anamensis («южная обезьяна озерная»). Канапои и Аллия Бэй, Туркана, Кения, 4,17/4,12–3,89 ± 0,02 млн лет назад [492].
9. Australopithecus afarensis («южная обезьяна афарская»). Танзания. Эфиопия: Белодели и Када Хадар, 3,89 ± 0,02 – ок. 3,0 млн лет назад [467; 799; 36; 443].
10. Australopithecus bahrelghazali («южная обезьяна с реки газелей»). Бахр эль Газаль, Коро Торо, Борку-Эннеди-Тибести, Чад, ок. 3,5–3,0 млн лет назад [219].
11. Australopithecus garhi («южная обезьяна-сюрприз»). Хата, Боури, западный Аваш, Афар, Эфиопия, ок. 2,5 млн лет назад [155; 408].
12. Australopithecus habilis («южная обезьяна умелая»). Олдувай, Танзания, ок. 1,83 млн лет назад [442; 495, с. 32–41; 708; 720; 816].
13. Paranthropus aethiopicus («околочеловек эфиопский»). Западный берег озера Туркана, Кения, 2,53 ± 0,05 – ок. 2,45 млн лет назад [787; 283].
14. Parantropus boisei («околочеловек бойсов»). Танзания, Кения, Эфиопия, ок. 2,5–1,3 млн лет назад [284; 286; 556; 741].
15. Paranthropus robustus («околочеловек массивный»). Сварткранс и Кромдраай, ЮАР, ок. 1,8–1,5 млн лет назад [284; 556; 720; 737; 738]: имел способность к точному захвату, позволяющему искусно владеть орудиями? cp. [599; 758].
16. Kenyanthropus platyops («кениец плосколицый»). Обособление линии, ведущей к роду Homo, – 1,86 млн лет назад (М) по Х-хромосоме [403], или 3,52 млн лет назад (Н). Ломекви, западный берег озера Туркана, Кения, ок. 3,5–3,2 млн лет назад [493].
17. Kenyanthropus rudolfensis («кениец с озера Рудольфа», акселерированный австралопитецин). Чемерон и Кооби Фора, Кения, 2,43 ± 0,02 – менее 1,82 ± 0,04, 1,6 ± 0,05 млн лет назад; Олдувай, Танзания, 1,839 ± 0,005–1,785 млн лет назад; Чивондо, Малави, ок. 2,4 млн лет назад; Хадар, Эфиопия, типичный олдовай, ок. 2,3 млн лет назад [193; 417; 487; 493; 495, с. 32–41; 510, с. 419; 679; 818; 819].
18. Homo ergaster («человек-мастер», акселерированный ранний питекантроп). В Африке 1,9/1,8–1,43 ± 0,02/1,41 ± 0,02 млн лет назад [153, с. 732, табл. 1, 733, рис. 1; 216; 217; 281; 487; 708; 741, с. 490]. В Азии: Лонггупо, Китай, олдовай, 1,942–1,785 млн лет назад [272; 426; 823]; Моджокерто и Сангиран, Ява, Индонезия, 1,81 ± 0,04, 1,66 ± 0,04 млн лет назад [507; 744]; Дманиси, Грузия, олдовай, Homo georgicus («грузинский человек»), ок. 1,75 млн лет назад [337; 338; 770].
19. Homo erectus («человек прямоходящий», неотеничный питекантроп). В Европе: Шандалья I, Хорватия, ок. 1,6 млн лет назад [500, с. 943]; д’Орсе и Куэва Виктория, Испания, ок. 1,5 млн лет назад [352]. В Азии: Триниль, Ява, Индонезия, 0,71 млн лет назад [500, с. 1031, 1071; 487].
20. Homo heidelbergensis («гейдельбергский человек»). В Африке: Олдувай, Танзания; Данакиль, Афар, Эритрея; Даканихило, Боури, средний Аваш, Афар, Эфиопия, ранний ашель, ок. 1,0 млн лет назад; Бодо д’Ар, средний Аваш, Афар, Эфиопия, развитой олдовай, 0,64 ± 0,03 млн лет назад; Брокен Хилл, Кабве, Замбия, поздний ашель (санго), Homo rhodesiensis («родезийский человек»), 0,11 млн лет назад [126; 154; 249; 500, с. 138, 159]. Исход из Африки: 0,63 ± 0,21 млн лет назад (М) [749], или 1,19 млн лет назад (Н). В Европе: Гран Долина, Сьерра де Атапуэрка, Испания, Homo antecessor («человек-предшественник»), 0,783 млн лет назад [188; 229]; прогрессивный неандерталец, 0,126 млн лет назад [44]. В Азии: Мугарет-эль-Зуттие, Вади Амуд, Кинерет, Израиль, финальный ашель (ябруд), Рисс III, 0,148 млн лет назад [500, с. 1148]; Нармада, Индия, поздний ашель, не бол. 0,15 млн лет назад [454]; Джинню-шань, Китай, 0,3–0,21, 0,281, 0,237, 0,187 млн лет назад; Джяньде, Китай, 0,117–0,09 млн лет назад [214]; [755]; Нгандонг и Самбунгмакан, центральная Ява, Индонезия, Homo soloensis («человек с реки Соло»), 53,3 ± 4–27 ± 2 тыс. лет назад [746]; пещера Лианг Буа, Рутенг, провинция Манггараи, о. Флорес, Малые Зондские о-ва, Индонезия, Homo floresiensis («человек с острова Флорес»), 95/74–12 тыс. лет назад [218; 584 а; 585; 572], где Homo heidelbergensis понимается по-афроевропейски узко, а потому «флоресиец» трактуется как поздний, карликовый Homo erectus, хотя лоб у малыша (рост женщины – 106 см) вздут не меньше, чем у иных «гейдельбержцев», а обезьянья челюсть не перекрывает мауэрской классики (знаменитой челюсти гейдельбергского человека из Мауэра в Германии, ок. 700 тыс. лет назад [273]) (рис. 8).
21. Homo neanderthalensis («неандертальский человек», акселерированный поздний гейдельбержец). Европа: 125 тыс. лет назад [43; 190; 285;
731]; Виндия, Хорватия, 29 080 ± 400 лет назад [128, с. 30, табл. 1]. На Ближнем Востоке: Кебара 12, Израиль, 59,9 ± 3,5 тыс. лет назад [765]; cp. [733]; Шанидар, Курдистан, Ирак, 60–44 тыс. лет назад [500, с. 968]. Расхождение с Homo sapiens – 620 ± 70 тыс. лет назад (М) [476, с. 25], или 1,17 млн лет назад (Н); ср. [614а, с. 184].
22. Homo sapiens praesapiens («человек разумный праразумный»). Грот де Фонтешевад, Орждей, Шаранта, Франция, эвано, интер-Рисс, 281–219 тыс. лет назад (фрагмент лобной кости Фонтешевад I неотличим от соответствующей кости черепа современной европейской женщины) [500, с. 397; 480, с. 219; 767, с. 352].
Homo sapiens Omo I, Omo II («человек разумный с реки Омо», 2 образца). Формация Кибиш, южная Эфиопия, 195 ± 5 тыс. лет назад [559].
Homo sapiens idaltu («человек разумный старший»). Херто, Боури, средний Аваш, Афар, Эфиопия, ашель – средний каменный век, 160 ± 2–154 ± 7 тыс. лет назад [248; 796]; cp. [316; 732, с. 693].
Homo sapiens modernus («современный человек разумный»). По Y-хромосоме – 153,5 ± 116,5 тыс. лет назад (М) [157, с. 1364; 295; 335; 396; 793; 801; 820]. По митохондриальной ДНК – 200 ± 80 тыс. лет назад (М) [228, с. 31; 774; 801]. По ДНК – 502,5 ± 386,5 тыс. лет назад (М) [157, с. 1364; 335; 646; 781]. Среднее по темпам мутагенеза – 200 ± 80 тыс. лет назад (М), или 378,81 тыс. лет назад (Н). Расогенез по селективно нейтральной дерматоглифике (пальцевым узорам и ладонным линиям) [111], что согласно [202; 626]; cp. [782]:
Негроиды – 169,6 ± 10,5 тыс. лет назад (Н); ср. 117,5 ± 82,5 тыс. лет назад (М) [157, с. 1364; 228, с. 35; 403; 451; 612; 749; 756; 820]. Класиес Ривер Маут, ЮАР, 125–105 тыс. лет назад [349; 729]; Бордер Кейв, ЮАР, св. 105 тыс. лет назад [349], 80–70, 65–50 тыс. лет назад [386], 74 ± 5 тыс. лет назад [387].
Европеоиды – 130,1 ± 4,3 тыс. лет назад (Н). Ближний Восток: Кафзех, Израиль (непрерывное обитание 20–30 тыс. лет), 90 тыс. лет назад [349], 115 ± 15, 92 ± 5 тыс. лет назад [734; 766]; Эс Схул, Израиль, 101 ± 12, 81 ± 15 тыс. лет назад [734]. Европа: пещера Бачо Киро 11, Дряново, Габрово, Болгария, ориняк I (ольшев) типа Ишталошко (нижний слой), Вюрм II Мурсхофд, 50 + 9/ – 4 тыс. лет назад (1σ) [579]; Бокштайн-Тёрле, Германия, ориньяк, 46380 + 1360/ – 1170 лет назад [256]; Эль Кастильо, Испания, ориньяк, 40 ± 2 тыс. лет назад; Л’Арбреда, Испания, ориньяк, 39,9 ± 1 тыс. лет назад [728]; грот де Фе, Шательперрон, Франция, ориньяк, 39780 ± 390, 35540 ± 280 тыс. лет назад [366а]; Пештера ку Оасе, Румыния, ок. 35 тыс. лет назад [803 a, с. 332]. Калибровка радиоуглеродного возраста по методу NotCal04 дает следующие даты распространения ориньякского сапиенса: Бокер Таштит, Израиль, св. 49 тыс. лет назад; Бачо Киро, Болгария, 46 тыс. лет назад; Богунице, Чехия, 46 тыс. лет назад; Виллендорф, Австрия, 45,5 тыс. лет назад; и т. д. до Шательперрона, Франция, 41 тыс. лет назад [567 а].

Рис. 8.
Человек с острова Флорес (Homo floresiensis) обитал на одном из Малых Зондских островов в Индонезии 74–12 тыс. лет назад, когда всей планетой уже владел биологический вид Homo sapiens. «Флоресиец» представлял собой позднюю карликовую форму гейдельбергского человека и достигал всего 106 см роста (судя по женскому экземпляру). Он не являлся ни сказочным хоббитом («чертенком»), ни позднейшим питекантропом, а сделался крохой подобно другим карликовым островным млекопитающим (таким, как карликовые мамонты о-ва Врангеля или вымершие карликовые слоны о-ва Мальта) [23а]
Австралоиды – монголоиды – 83,9 ± 3,1 тыс. лет назад (Н). Азия: Люджянг, Китай, 67 тыс. лет назад [214]; Япония, 70–50 тыс. лет назад [160]. Австралазия: Ниах грейт кейв, Саравак, Калимантан, Малайзия, св. 41500 ± 100 лет назад; Хуон, Папуа – Новая Гвинея, 53–45 тыс. лет назад [382]; Матенкупкум, Новая Ирландия, северная Меланезия, 32700 ± 1550 лет назад [135, с. 709]; Малакунанджа II, западный откос плато Арнемленд, Австралия, 61 ± 13–45 ± 9, 52 ± 11 тыс. лет назад [648]. Америка: Монте Верде, Чили, 33370 ± 530 лет назад; Тока ду Бокейрану да Педра Фурада, Бразилия, св. 32 тыс. лет назад [385; 500, с. 708].
Представители рода Homo отличались строением тела, приспособленным к прямохождению. Они имели относительно короткие руки и относительно длинные ноги, как у двуногих динозавров. В отличие от прочих млекопитающих, наши предки обладали не слишком различающимися резцами, клыками и коренными зубами, т. е. были равнозубыми, как динозавры, что объясняется их ориентацией на высокобиопродуктивную среду, когда не требуется тщательно утилизировать пищу. Сходство усугублялось тем, что некоторые динозавры (ранние карнозавры и целурозавры) располагали хватательными верхними конечностями с отстоящим внутренним пальцем, аналогом нашего большого [25, с. 45–46, 49]. Однако, несмотря на подходящую физическую организацию, мы все еще не обладаем твердыми наследственными навыками прямохождения. Это доказывается судьбами homo ferus (детьми, выращенными животными), которые без человеческого воспитания к прямохождению не способны (ползают на четвереньках) [76, с. 47–49, 183–186; 576].
Следует добавить, что, хотя люди преодолевают стометровку не быстрее крохотной, но четвероногой крысы (ок. 10 с в обоих случаях), они все же кое в чем продвинулись как выносливые бегуны. Предки рода Homo были приспособлены к выносливому бегу гораздо хуже [207], что отвечает большей неотеничности. Способность к выносливому бегу появилась лишь у Kenyanthropus rudolfensis или Homo ergaster в результате акселерации (противоположной неотении, см. ниже). Человек современного вида перенес до трех акселераций, а потому оказался способен к марафону, хотя все еще десятикратно уступает в этом четвероногим гиенам или волкам.
Представители остальных родов гоминин, будучи тоже прямоходящими, сохраняли обезьянье строение тела. Они имели относительно длинные, питекоидные руки (Australopithecus habilis, Australopithecus afarensis, Ardipithecus ramidus [36; 145, с. 556; 442; 443]), а также обезьянье строение вестибулярного аппарата, существенного для прямохождения (Australopithecus habilis, Paranthropus robustus [720]). Иными словами, история прямохождения у гоминин, от самых ранних до современных, постепенна и не закончена до сих пор.
Прямохождение возникло именно у наших обезьяноподобных предков благодаря эволюционной отсталости приматов, делающей их податливыми для фундаментальных перемен. Среди млекопитающих эволюционно продвинуты не они, а мышевидные грызуны [82, с. 97]. Энергетически экономная двуногость явилась откликом на подъем биопродуктивности среды в связи с потеплением, когда подвижный поиск пищи стал неактуальным, раз она обильна и скучена в пространстве. Подобная кормовая база спровоцировала у гоминин «мутацию прожорливости». Они стали потреблять примерно в 3,8 раза больше пищевой энергии на единицу массы тела, нежели все остальные высшие млекопитающие. У последних соответствующий показатель (постоянная Рубнера) составляет 191 600 ккал на кг массы тела в течение жизни, тогда как у людей это 725 800 ккал на кг телесной массы [6, с. 73, 89; 661; 662]. Таким образом, метаболизм
(обмен веществ) у людей по сравнению с другими высшими млекопитающими завышен в 3,8 раза.
Неотения и акселерация
Вынужденные перерабатывать почти вчетверо больше энергии, чем равновеликие млекопитающие, гоминины стали пропорционально дольше жить [323] и, соответственно, формироваться. Их индивидуальное развитие затормозилось, растянулось во времени и привело к сохранению инфантильных (детских) и даже эмбриональных черт во взрослом состоянии, что называется неотенией
(от греч.
νεότης – юность) (рис. 9) [10; 12, с. 17–19; 402; 577; 584, с. 13–44]. Аналогичное явление (педоморфоз) известно у земноводного аксолотля [564]. У гоминин неотения выразилась в удержании эмбриональной обезволошенности тела и невыраженности лицевого скелета, в силу чего мозговой скелет, купол черепа, непропорционально вздулся. Это не связано с интеллектуальными нагрузками: у испытывающего их последние две с половиной тыс. лет сапиенса мозг ничуть не изменился. Известно, что вздутие купола черепа началось у Australopithecus afarensis до того, как усложнилась кортикальная организация его мозга (организация его мозговой коры) [315]. Проще сказать, у гоминин обезьяний по сути мозг механически разросся в силу неотении, не усложняясь как орган интеллекта. Отметим, что старший современник «афарца», Australopithecus africanus из Таунга в ЮАР, по стадиям прорезывания зубов напоминал не ребенка 5–7 лет (как следовало бы из его вероятного возраста), а детеныша человекообразной обезьяны 3–4 лет [257, с. 626] в силу того, что фенотипическая (телесная) неотения отставала у гоминин от генотипической. Аналогичным образом наша генетическая предрасположенность к прямохождению еще не гарантирует, что человеческий ребенок усвоит двуногость автоматически.

Рис. 9.
Человек и его предки гоминины возникли в результате эволюции путем неотении, когда организм во взрослом состоянии сохраняет детские и даже эмбриональные (зародышевые) особенности. У человека к ним относятся обезволошенность тела (эмбрионы человека и многих других млекопитающих безволосы), а также вздутый купол черепа. У эмбрионов человека и прочих высших млекопитающих он вздут, однако после рождения у животных сильно разрастается морда, и мозговой участок черепа выглядит убогим; у людей этого не происходит: лицо остается крохотным, а потому мозг кажется обширным. По общему (ошибочному) убеждению это объясняется большим интеллектом людей. Однако человеческий ум существенно развивался только у Homo sapiens, да и то лишь последние 2,5 тыс. лет. При этом мозг человека современного типа не изменился ни на йоту. Вдобавок наш мозг активно загружен не более чем на 10%. Здесь мы видим эмбриональные и зрелые формы черепа у собаки и у человека. Оба эмбриональных абриса похожи, но лишь у человека зародышевые очертания сохраняются во взрослом состоянии [12, с. 19]
Под влиянием неотении, замедлившей индивидуальное развитие наших предков, у женщин и самок гоминин месячный цикл растянулся, так что эпизоды способности к зачатию как бы слились, в результате чего эструс (аналог течки у самок животных) размазался и стерся, хотя у женщин сохраняется несколько ежемесячных дней, когда они повышенно восприимчивы к ухаживаниям.
Самки гоминин стали существенно дольше, чем равновеликие млекопитающие, вынашивать потомство, что вызывало накопление ими заметных жировых запасов, необходимых для питания плода [107; 402; 583; 628]. Начиная с питекантропа, изменились сроки прорезывания зубов у молоди наших предков [280]. На протяжении своей эволюции гоминины все заметнее уподоблялись эмбрионам, что выразилось у них в грацилизации, т. е. в утончении костей скелета и приобретении изящества. Ныне это явление усиливается благодаря цивилизованному образу жизни: он вызывает последствия, напоминающие эффекты одомашнивания животных, которое изнеживает, инфантилизирует наших питомцев [490, с. 357–358].
Неотеническое увеличение продолжительности жизни у гоминин имело эволюционные последствия. Темпы эволюции зависят от скорости смены поколений. Поэтому замедление онтогенеза влечет за собой торможение филогенеза вида. У гоминин он в 3,8 раза замедлен против равновеликих млекопитающих. Поэтому, сравнивая их молекулярную эволюцию с нашей, мы должны умножать хронометраж на своего рода «неотенический коэффициент» 1,894, чтобы привести к среднему знаменателю нормально текущую эволюцию с нашей, вчетверо заниженной. Как можно видеть из вышеприведенного хронометрированного списка гоминин, подобная коррекция молекулярно-генетических дат (М) при помощи «неотенического коэффициента» дает исправленные датировки (Н), совместимые с палеоантропологическими данными. Заторможенность молекулярного мутагенеза у человека как таковая выявлена эмпирически [506], однако она не дотягивает до заторможенности человеческого онтогенеза, что объясняется сложностью расчетов абсолютных количественных показателей для мира млекопитающих, где они зависят от размеров, физиологии и других особенностей животных. Так, продолжительности жизни кошки, человека и кита относятся примерно как 1:10:20, в то время как массы их тел относятся друг к другу как 1:10:1500, откуда видно, что рост продолжительности жизни у млекопитающих заметно отстает от увеличения их размеров.
Принятое нами отношение древности современного человека (378,81 тыс. лет) к возрасту гоминин (11,84 млн лет) составляет 1/31,25, в то время как ожидаемое достигает почти 1/25 [100, с. 12]. Данное противоречие объясняется тем, что современный человек перенес три акселерации против двух у своих предшественников (см. далее). Это должно было ускорить его молекулярную эволюцию в 1,5 раза – вышло 1,25, ибо последняя, текущая акселерация только началась. Проще говоря, в истории наших предков имелись эпизоды ускоренной эволюции (акселерации), отчего их молекулярно-генетический возраст оказался ниже ожидаемого (т. е. возраста ок. 474 тыс. лет).
В сравнении с армадой двуногих мезозойских динозавров прямоходящие гоминины выглядят сиротливо. Двуногие прыгающие сумчатые (например кенгуру), насекомоядные (например прыгунчики) или грызуны (например тушканчики) прибегают к энергоемкой скачущей локомоции. Она быстра, но не экономична. Выгодной ходящей или бегающей локомоцией ныне обладают лишь бегающие птицы. Она выгодна, но не проворна. Так, африканский страус при массе тела 70–90 кг и соответствующей мощи развивает скорость всего 70 км/ч, тогда как гепард с массой тела 50–65 кг способен на спринт до 112 км/ч. Четвероногие ходоки и бегуны легко покрывают огромные пространства (например волки 500–600 км за день), что благоприятно в нынешних условиях скромной, разреженной пищи. В мезозое ее плотность была в среднем выше, а потому отбор дал плеяду прямоходящих динозавров. Сейчас он поощряет четвероногих животных, а гомининам отводит роль исключения. Отсюда наша аномально высокая постоянная Рубнера и нестандартная эколого-эволюционная судьба.
Живя дольше равновеликих млекопитающих, гоминины накапливали в своих сообществах представителей разных поколений и демографически умножались, что продолжается по сей день. Организмы в биомах подчиняются популяционным волнам, когда при благоприятных условиях последовательно умножаются сперва растения, потом растительноядные потребители, а затем хищники. Далее растительноядные истощают кормовую базу, падают в числе и вызывают упадок хищников. Раз за разом процесс повторяется [80, с. 243–258; 97, с. 62–72]. Поэтому поголовье животных в биомах в среднем фиксировано. Медленно развиваясь и размножаясь, гоминины не успевали за популяционными волнами, а потому постепенно, но неуклонно росли в численности. Их демографическая история объясняет ряд новаций в эволюционном состоянии и образе жизни.
Напомним, что для видообразования требуется репродуктивная (по воспроизводству себе подобных) изоляция меньшинства от большинства [82, с. 93]. Напротив, высокие плотности населения затрудняют эволюционные изменения, отчего многочисленное современное человечество не эволюционирует [82, с. 96]. Прямохождение, высокий метаболизм и неотения возникли у наших предков в условиях низкой плотности населения, а затем фенотипически прогрессировали до наших дней. Условием для такого порядка вещей были умеренные плотности населения.
Между тем в силу неотении гоминины были склонны к повышению своей численности. При ее пиковых изменениях, демографических взрывах следовало бы ожидать процессов, обратных неотении. Неотения – это замедление развития. Поэтому при демографических всплесках приходилось бы ожидать явлений ускорения развития, т. е. акселерации.
Демография
Однако гоминины не могли выдерживать длительных периодов своей повышенной численности, поскольку их перенаселение истощало кормовую базу, подрывало равновесие со средой и угрожало им крахом, ведь они были эволюционно заторможены и не могли быстро изменяться, восстанавливая свою численность. Выжили те их сообщества, которые стихийно научились стабилизировать свою населенность.
К концу оледенения Бибер I (3,2–3,0 млн лет назад), чреватого для Африки аридностью (засушливостью и скудостью), гоминины должны были попасть в депрессию, а их сообщества изолироваться, что благоприятствовало видообразованию. В плювиальный (влажный и обильный) интерстадиал Бибер I/II (3,0–2,6 млн лет назад) гоминины размножились в новом видовом составе, так что на хронологической точке в 2,5 млн лет назад они представлены новыми видами Australopithecus garhi, Paranthropus aethiopicus и, возможно, Kenyanthropus rudolfensis. Хотя последний известен лишь с отметки 2,43 ± 0,02 млн лет назад, свойственная ему археологическая культура типичного олдовая началась еще св. 2,63 ± 0,5, 2,58 ± 0,23 млн лет назад.
Если предположить, что гоминины человеческой линии (т. е. Kenyanthropus) ощутили вышеуказанные эколого-демографические трудности, то естественным способом обойти их мог стать для «кенийцев» уход в другую экологическую нишу. Биомасса растений в биоме примерно в 10 раз превосходит биомассу растительноядных потребителей, а биомасса последних – в 5 раз биомассу хищников [80, с. 86–87]. Подобно близкородственным шимпанзе (в особенности шимпанзе-пигмею бонобо [667]), древние гоминины были всеядными существами с преобладанием растительной диеты [139; 277]. Если бы они обратились к хищничеству, их численность автоматически упала бы в 5 раз (условно), что нейтрализовало бы их неотеническое размножение, в 3,8 раза превышающее свойственное равновеликим млекопитающим. Мнение о том, что гоминины обратились к некрофагии (трупоядению) [13; 18, с. 180–195; 90, с. 107–113; 711]; cp. [501; 772], неприемлемо, поскольку экониша падальщиков в силу своей вторичности теснее экониши хищников, и такая конкуренция была бы не по силам преимущественно растительноядным гомининам, мало пригодным к борьбе за падаль, например, со львом (50%-ный падальщик) или гиеной (30%-ный падальщик). Проще сказать, активным охотникам доводится бороться лишь с добычей, которая не всегда способна постоять за себя, в то время как падальщикам постоянно приходится сражаться с другими некрофагами, среди которых имеются смертельно опасные фигуранты (по степени опасности в Африке это львы, гиены и леопарды, которые совмещают потребление падали с умелой активной охотой, а потому являются грозными соперниками). Кроме того, в эконише некрофагов попросту меньше пищи, нежели в эконише охотников за живой добычей. Вдобавок даже состоявшуюся падаль стремительно пытаются растащить и некрофаги скромных габаритов (шакалы, грифы, стервятники).
Шимпанзе (а также павианы) около 3% пищи добывают групповой охотой [18, с. 183; 59, с. 142–143, 147; 103, с. 180, 182, табл. 6.8; 194; 722]. Трудно сомневаться, что гоминины, известные впоследствии как великие охотники, на заре своей истории совпадали по этой части с шимпанзе. «Кенийцы», усугубив свое пристрастие к животной пище, угодили в эконишу хищников и сократились в численности. Возможно, их путем пошли и прочие гоминины, например Australopithecus africanus (любил листву и/или сочные фрукты [380], а также мясо [719]), отличавшийся от земляка Paranthropus robustus (всеяден [349], с пристрастием к грубой пище [380]).
Решение демографической проблемы посредством перехода в эконишу хищников оказалось временным, поскольку «кенийцы» продолжали размножаться и испытывать прежние эколого-демографические трудности, еще более опасные для хищников, нежели для травоядных (хищники малочисленнее и уязвимей). В разное время эта незадача привела к вымиранию прочих австралопитецинов. «Кенийцы» же развили питекоидные способности к производительному применению орудий, впервые в истории животных сделали их коллективными и положили начало демографо-технологической зависимости, связывающей плотность населения со сложностью технологии (см. разд. 3.1).
Осваивая орудия археологической культуры типичного олдовая (2,6 млн лет назад) и переживая неблагоприятный гляциал Бибер II (2,6–2,3 млн лет назад), «кенийцы» некоторое время пребывали в стабильном состоянии. Однако с наступлением мягкого интергляциала Бибер/Донау (2,3–2,0 млн лет назад) они вновь размножились, открыв первый в истории человечества демографический взрыв. Около 2,2 млн лет назад он выплеснул «кенийцев» из Африки в Евразию, где их типично олдовайские орудия отмечены в Диринге (Якутск, Якутия, 3,2/3,1–2,7/2,2 млн лет назад [75, с. 131]), в Сен-Валье (Дром, Франция, 2,2 млн лет назад [500, с. 935]) и в Сент-Эбле (Мон Купе, Франция, 2,5–2,2 млн лет назад [127, с. 28]). Высокая плотность населения вызвала в среде «кенийцев» акселерацию и увеличение размеров тела, отчего Kenyanthropus rudolfensis долгое время воспринимался в палеоантропологии как крупный вариант Australopithecus habilis [815].
После этого эпизода «кенийцы» до конца ледниковой стадии Донау I (2,0–1,9 млн лет назад) около 300 тыс. лет снова пребывали в эволюционном покое, поскольку разрядили демографическую обстановку предпринятой экспансией. Умеренная плотность населения вернула им полноту неотении, очередным этапом которой стало появление нового гоминина с обостренной неотеничностью. Это выразилось в увеличении его мозгового черепа, укорочении верхних конечностей (инфантильный признак) и усовершенствовании вестибулярного аппарата, обслуживающего прямохождение, а также в сапиентных темпах взросления, судя по стадиям прорезывания зубов. Иначе говоря, возник питекантроп.
Он сразу же испытал рост плотности населения, пережил второй демографический взрыв и 1,835 млн лет назад мигрировал в Евразию, где известен от Индонезии до Испании (исходим из того, что размеренное расселение от Восточной Африки до Индонезии занимало 25 тыс. лет [507], а в Индонезии «человек-мастер» появился 1,81 млн лет назад). Прохождение им Левантского коридора отмечено олдовайской индустрией в Эрк-эль-Ахмар (долина Иордана, Израиль, 1,95–1,77 млн лет назад) [656]. Под влиянием повышенной плотности населения он испытал вторую акселерацию и предстал как Homo ergaster, по существу классический питекантроп акселерированного типа. Его африканский представитель, образец Нариоко-томе III (Туркана, Кения, 1,60 ± 0,05 млн лет назад [216; 217; 708]), имея возраст 12 ± 1 год, уже достигал роста 1,68 м, что обещало в зрелости ок. 2 м. Его азиатский представитель из Сангирана (Ява, 1,66 ± 0,04 млн лет назад) тоже известен как гигант, «мегантроп». Наконец, ребенок из Моджокерто (Ява, 1,81 ± 0,04 млн лет назад) в 1 ± 0,5 года имел объем черепа 72–84% от взрослого питекантропа. Такое соотношение юных и зрелых особей присуще шимпанзе и на 15% опережает развитие человеческого ребенка [261], т. е. является акселерированным. Кстати, акселерированный Homo ergaster до 2 м роста – типа Нариокотоме III или «мегантропа» – представляется наиболее вероятным кандидатом на роль гипотетического «снежного человека», скрытно распространенного в горах Центральной Азии и лесах Северной Калифорнии.
Прямых данных о росте плотности населения у африканского Homo ergaster нет. Погибшее при наводнении сообщество Australopithecus afarensis в Хадаре (Эфиопия, 3,5–3,0 млн лет назад) насчитывало ок. 13 индивидов [36, с. 158, 225]. Носители олдовайской культуры в Олдувае (Танзания, ок. 1,8 млн лет назад) жили сообществами в 15 ± 5 индивидов. Спустя миллион лет этот показатель удвоился, и сообщество Homo heidelbergensis в Олоргесаилие (Кения, 0,74–0,60 млн лет назад) насчитывало 30 ± 20 индивидов [50, с. 87–88; 249]. Поскольку данных о демографических взрывах у «гейдельбержцев» нет (см. далее), можно думать, что часть этого удвоения населения приходилась на долю демографического всплеска у «человека-мастера», а другая часть обусловила медленное расселение «гейдельбержца» в Евразию ок. 0,783 млн лет назад.
Подчиняясь демографо-технологической зависимости, Homo ergaster в силу возросшей плотности населения породил ашельскую технологию (1,6–1,5 млн лет назад), после чего пришел в демографически-стабильное состояние, вернулся к неотеничному облику и предстал как классический Homo erectus, менее рослый, но более головастый. Углубление этого инфантильного облика переродило его в Homo heidelbergensis, известного в Африке ок. 1,0 млн лет назад.
Несколько ранее, в интерстадиале Гюнц I/II (1,27–0,93 млн лет назад) 1,19–1,17 млн лет назад (Н) на стадии позднего питекантропа или раннего гейдельбержца, разошлись пути предков сапиенсов и неандертальцев. В теплую межледниковую стадию Гюнц/Миндель II (790–770 тыс. лет назад) ок. 783 тыс. лет назад Homo heidelbergensis достиг Европы, где предстал в Испании как Homo antecessor. Около 300 тыс. лет назад он появился в Китае, освоил Индонезию и закончил свой путь на Яве как Homo soloensis уже в эпоху азиатских сапиенсов. Поскольку равномерное расселение покрывает расстояние от Восточной Африки до Явы всего за 25 тыс. лет [507], передвижения гейдельбержца нельзя назвать демографическим взрывом. Сопутствующих примет акселерации он также не обнаруживал.
В теплую межледниковую эпоху Рисс/Вюрм (144–110 тыс. лет назад) финальный гейдельбергский человек в Европе, «прогрессивный неандерталец», достиг таких плотностей населения, что принял акселерированную форму Homo neanderthalensis (классический неандерталец), обозначившуюся 125 тыс. лет назад. Этот гоминин был на 30% крупнее нынешнего среднего человека и даже на 24% превосходил массивных обитателей наших высоких широт [449, с. 125], что отражало ускорение онтогенеза у неандертальца [541]. Наши высокоширотные современники акселерированы в силу того, что у млекопитающих в суровых условиях поднимаются скорости основного метаболизма (обмена веществ) [291], в результате чего организм развивается быстрее (этот повседневный метаболизм не следует смешивать с пожизненным метаболизмом, способным растягивать биографию организма, см. выше.) Неандертальский человек был столь форсирован в развитии, что приобретал питекоидные черты черепа еще на пренатальной стадии, в утробе матери [627]. Его зубные коронки формировались на 15% быстрее, чем у современных людей, а зрелость наступала в 15 лет [658, с. 937]. По темпам акселерации он не уступал Homo ergaster из Моджокерто. О демографическом взрыве и территориальных конфликтах у неандертальцев свидетельствует каннибализм (cp. [163]).
Холодный интерстадиал Вюрм II B 1–2 (59–51тыс. лет назад) толкнул разросшуюся популяцию неандертальцев к миграции на юг, где 59 тыс. лет назад они соприкоснулись с ближневосточными сапиенсами. Подчиняясь демографо-технологической зависимости, европейские неандертальцы в эпоху Вюрм II/III (37,5–36 тыс. лет назад) создали прогрессивную верхнепалеолитическую культуру перигор 0, сменившуюся перигором I (шательперроном), обозначенным в эпохи Вюрм II/III и Вюрм III А 1 (37,5–32,5 тыс. лет назад). В конце холодного стадиала Вюрм III B (30–29 тыс. лет назад) неандертальцы, окруженные процветающими сапиенсами, прекратили свое существование.
Популяция Homo heidelbergensis из Бодо д’Ар в Эфиопии (640 ± 30 тыс. лет назад) в начале интергляциала Миндель/Рисс (388–333 тыс. лет назад) настолько углубилась в неотению, что породила так называемую популяцию «Евы», 378,81 тыс. лет назад (Н), представители которой отличались особо вздутым куполом черепа и, соответственно, относительно убористым лицом, что является характерными анатомическими признаками вида Homo sapiens. Он представлял собой, таким образом, этап инфантилизации гейдельбергского человека (в противоположность неандертальцу, который являлся акселерированной стадией гейдельбергского человека). В теплую интерстадиальную эпоху интер-Рисс (281–219 тыс. лет назад) ранние сапиенсы (пресапиенсы) посетили Европу, где оставили каменные орудия прогрессивной культуры эвано верхнепалеолитического облика. Их представители типа Фонтешевад I не снискали будущего в Европе.
В эпоху значительного оледенения Рисс III (219–144 тыс. лет назад) развившаяся пустыня Сахара отделила североафриканскую часть популяции «Евы» от субсахарской. Последняя положила начало негроидной расе современного человека, 170 тыс. лет назад (Н), представленной автралоидным Homo sapiens idaltu (160 ± 2–154 ± 7 тыс. лет назад) [732, с. 693] и монголоидообразными людьми койсанской (бушменской) расы из Класиес Ривер Маут и Бордер Кейв в ЮАР, 125–105 тыс. лет назад. Австрало-монголоидный облик, вероятно, был типичен для популяции «Евы», что проявилось в конце ее существования (см. ниже). В теплом Рисс/Вюрме (144–110 тыс. лет назад) она двинулась на север и, положив начало европеоидной расе на Ближнем Востоке 130 тыс. лет назад (Н), отбыла на восток. Ее отпрыски в Израиле представлены протокроманьонцами из Кафзех (115 ± 15–92 ± 5 тыс. лет назад) и Эс Схул (101 ± 12–81 ± 15 тыс. лет назад).
Достигнув Индии, популяция «Евы» распалась на австралоидов и монголоидов, разделивших ее анатомические черты 84 тыс. лет назал (Н).
Свидетельством ее пребывания на субконтиненте является полиморфизм (разнообразие) щелочной фосфотазы плаценты и других признаков у местного населения, поскольку он характерен для центра распространения биологического вида [92], который в случае с человеческими расами переместился из Восточной Африки в Индию. Как отмечалось выше, популяция «Евы» сочетала австралоидные и монголоидные признаки, которые обусловили облик Homo sapiens idaltu и представителей койсанской расы. Они же предрешили типажи австралоидной и монголоидной рас после расщепления популяции «Евы» в Индии. Можно допустить, что она отличалась и европеоидным началом, которое на различных стадиях распада протопопуляции сообщило сходные черты европеоидам и америндам (американским индейцам), в частности развитое строение носа, хотя у европеоидов оно основано на костной, а у америндов – на хрящевой ткани [2, с. 158; 3, с. 27].
Тем временем у европеоидов на Ближнем Востоке произошел демографический взрыв, и они вторглись в Европу, используя потепление интерстадиала Вюрм II Мурсхофд (51–46,5 тыс. лет назад). В силу демографо-технологической зависимости всплеск их народонаселения породил пластинчатую технологию ориньякской археологической культуры (50 тыс. лет назад). Освоив Европу, пребывающие на пике демографического подъема сапиенсы вытеснили неандертальцев из выгодных охотничьих угодий и положили конец их виду (29 тыс. лет назад). Причина эволюционного успеха сапиенсов и поражения неандертальцев коренится в том, что у первых демографический взрыв состоялся позже (ок. 50 тыс. лет назад), нежели у вторых (ок. 125 тыс. лет назад), и перекрыл результаты их размножения. В силу того же демографического подъема европеоиды вошли в фазу акселерации и дали популяцию абри де Кро-Маньон (Лез Эйзиде-Тейяк, Дордонь, Франция, ориньяк IV, 27 680 ± 270 лет назад [567, с. 463]). Эти кроманьонцы в узком смысле слова отличались средним ростом 185 см у мужчин, в чем не уступали современным акселератам.
Следующий, преднеолитический, демографический взрыв случился с европеоидами тоже на Ближнем Востоке 16 тыс. календарных лет назад с окончанием первой местной мезолитической культуры кебары (21,8–16 тыс. календарных, или 19–13,95 тыс. радиоуглеродных лет назад). В это время в Леванте (Израиль, Иордания, Ливан, Сирия) начался распад общности носителей шумерского, сино-кавказских и ностратических языков, т. е. почти всех известных языков белой расы, а также некоторых групп восточных и центральноафриканских негроидов, северных и дальневосточных монголоидов, северо-западных америндов [52, с. 65–73]. В соответствии с демографо-технологической зависимостью этот популяционный взрыв вызвал неолитическую технологическую революцию 11,7 тыс.
лет назад. Однако признаки акселерации у тогдашних левантийцев не известны. Они ожидались бы у носителей третьей мезолитической культуры региона, натуфийской (14,3–11,7 тыс. календарных, или 12,45–10,2 тыс. радиоуглеродных лет назад), созданной первыми афразийцами (предками древних египтян, евреев, арабов и пр.). Между тем, средний рост натуфийцев-мужчин не превышал 1,60–1,71 м [762, с. 50], что исключает акселерацию. Возможно, она проявилась в неолитическое или более позднее время, однако данных на этот счет нет. Можно лишь отметить, что исторические западные семиты из Передней Азии отличались большей рослостью, нежели, например, древние египтяне.
Наконец, последний демографический взрыв произошел в Западной Европе исторически недавно. Его первый этап относится к XI – середине XVI вв. [23, с. 67–75]. Умножение западноевропейского населения ознаменовалось крестовыми походами на Восток (1096–1270 гг.), германской экспансией и Великими географическими открытиями кон. XV – XVII вв. Благодаря демографо-технологической зависимости вслед за демографическими событиями последовала промышленная революция, открывшаяся мануфактурами XIV в. в Италии.
С кон. XVIII в. в Западной Европе развертывается акселерация роста и созревания населения [12, с. 423–427]. Во 2-й пол. XX в. эта тенденция отчетливо охватила Северную Америку, Австралию, Россию и Восточную Азию, т. е. приобрела глобальные масштабы. Данное биологическое явление представляется нормальной эволюционной реакцией человечества на рывок демографического подъема. Отметим, что плавное демографическое развитие Индии, Юго-Восточной Азии и Китая никогда не сопровождалось процессами акселерации и действием демографо-технологической зависимости. Исключение составляет эпоха тамошней неолитической революции (в технологическом плане), а также современная акселерация в Китае и Японии (см. разд. 3.1).
Красота
Неотения отразилась не только на биологической и технологической истории гоминин. Она затронула врожденные человеческие чувства прекрасного, связанные с оценкой физического облика себе подобных, т. е. с оценкой их физической красоты [53, с. 13–15]. Общеизвестно и подкреплено специальными исследованиями [616], что у людей физически привлекательны признаки здоровья, плодовитости и властности. Эти особенности привлекательны для особей противоположного пола у всех позвоночных и многих беспозвоночных животных. Следовательно, названные особенности не могут считаться специфически человеческими, и мы не станем на них останавливаться. Мы попытаемся раскрыть природу физической красоты, присущей лишь человеку и, вероятно, его гомининным предкам.
Вопреки существующему убеждению, природа человеческой красоты не является тайной. Она обусловлена биологически и воспринимается инстинктивно. Как мы видели, в своей истории гоминины неоднократно проходили неотенические и акселерированные стадии развития. Неотеничность отвечает эволюционной направленности гоминин. Поэтому она репродуктивно (сексуально) привлекательна. Однако чередование неотеничных и акселерированных эпизодов привело к тому, что представления о красоте раздвоились.
Красота лица неотенична, поскольку эволюционно перспективна. Красота тела акселерирована, поскольку акселерация создает механические трудности при деторождении. Проще говоря, в нормальной человеческой популяции присутствуют как низкорослые неотеничные индивиды, так и высокорослые акселераты. Они свободно скрещиваются. Поэтому некрупная неотеничная женщина может родить ребенка от акселерата. Однако женский таз – это жесткая конструкция. Он затрудняет производство акселерированного потомства. Поэтому женщины независимо от роста имеют большой объем таза.
У акселераток он органичен. У неотеничек – вынужден. В любом случае биологически выгодно быть первыми. Поэтому они инстинктивно оцениваются как гармоничные, красивые. Отсюда интуитивно (по привлекательности) выстраиваются кроманьонские параметры современных топ-моделей. Это высокорослые женщины ок. 180 см высоты, с объемами 90–60–90, что отражает характерные параметры акселерации – как нынешней, так и древней, кроманьонской. Высокорослые мужчины красивы в силу соответствия подобным женщинам. Мы не станем говорить о банальной выгодности высокорослости при охоте и т. п. – хотя бы потому, что все зависит от того, где и на кого охотиться. Как известно, в джунглях охотнику выгоднее быть пигмеем.
Происхождение притягательных черт лица гораздо проще. В результате неотении череп наших предков стремился во взрослом состоянии сохранять эмбриональную модель, т. е. быть по возможности сферичным. Для решения этой задачи лицевой отдел черепа ушел под лоб, надбровья выпрямились и наехали на глазницы, скулы, нос, челюсти и зубная дуга сократились. В результате сформировалась женская классическая или кукольная внешность. Ее образцами отличаются, например, Ким Уайлд (английская певица с весьма выразительным лицом) или Магда Конопка (западная актриса польского происхождения, типологически близкая Ким Уайлд и вдобавок носившая титул «леди Вселенная»). Мужская красота в своей основе та же, только массивнее в силу полового диморфизма (полового расхождения). Образцами красивой мужской внешности обладали, например, лорд Байрон (великий английский поэт) или Рудольф Валентино (знаменитый актер немого кино).
Можно разобрать черты лица в деталях. Красив высокий закругленный лоб – потому, что его форма оптимально вписывается в сферичную неотеничную модель. Все прочее представляется угловатым и негармоничным. Красивы вытянутые глаза в низких глазницах – потому, что в древности округлые обезьяньи глазницы наших предков в результате неотении сплюснулись у гоминин под наехавшим лбом, отчего стали низкими и визуально продолговатыми. Отсюда же притягательность прямых бровей, бровей вразлет и т. п. форм. Оптимален небольшой прямой или чуть вздернутый нос (у женщин), находящийся подо лбом. Он хорошо вписывается в сферичную, неотеничную модель. Миниатюрные, утопленные в лицо скулы очаровательны по той же причине, как и облегченные челюсти, подбородок и зубная дуга. Понятно, у мужчин все должно выглядеть рельефнее, «мужественнее».
Очерченные критерии физической красоты, в принципе, однотипны во всех расах. Однако следует помнить их специфику. Негроиды оформились в резулультате вздутия черепного свода (общего для всех рас), миниатюризации скул и носа. Челюсти отставали от процесса, и это называется прогнатизмом (сильное выступание тяжелых челюстей вперед). С европеоидами случилось то же, лишь уменьшились челюсти (ортогнатность, утопленность челюстей под скулы), а нос остался велик. Монголоиды отличаются плоскими, но широкими скулами, а австралоиды сохраняют общую грубость лица, впрочем усредненно сократившегося как целое. Неотеничные критерии красоты следует применять к расам с указанными поправками. Непоследовательность в формировании лиц по расам объясняется тем, что все они независимо произошли от исходной популяции «Евы», перемещавшейся из Восточной Африки в Индию.
Как можно видеть, магистральные пути гоминизации у наших предков укладывались в поступательную неотению или отклонения от нее (Homo ergaster, Homo neanderthalensis). Неотения, прямохождение и высокая постоянная Рубнера (а также связанная с ней прожорливость) одинаково вытекали из эволюционной направленности гоминин в сторону экономной маломобильности, что проявляется в склонности к гиподинамии и ожирению у современного человека.
Если генетическое обеспечение гоминизации состоялось на заре трибы гоминин, то фенотипическое воплощение заложенных в генетической программе тем не завершилось до сих пор. Это выражается в ряде примет количественной эволюции (см. разд. 2.3). У человечества продолжаются редукция (измельчание) зубов и сокращение зубной дуги, равно как и общее облегчение всего скелета (грацилизация). Углубление внешних проявлений неотении ведет ко все большей миниатюризации человеческого лица, отчего грубые лица среднему человеку инстинктивно кажутся несовременными, «пещерными», что в определенном смысле отвечает сути дела. Наконец, развитые (и не только) страны страдают от гиподинамии и ожирения населения, что объясняется развертыванием заложенной в нас генетической программы. Гиподинамия представляется опасным, но адекватным воплощением нашей эволюционной маломобильности, а ожирение – досадным выражением одного из последствий неотении (напомним, что ожирение у женщин обусловлено аномально растянутой беременностью; у мужчин же избыточный вес прибывает в силу генетической солидарности с прекрасным полом, обусловленной общим для обоих полов генотипом). Последняя пара примеров эволюционно чрезмерна, невыгодна. Однако в гигантском человечестве естественный отбор подавлен, а потому на свет беспрепятственно выходят такие наши генетические задатки, которые в небольшой, активно эволюционирующей популяции давно бы пресеклись.
Понимание природы гоминизации имеет мировоззренческое значение. Человек стремился и стремится познать самого себя, как призывал еще Хилон Лакедемонский, третий из семи греческих мудрецов [104, с. 92–93]. Осуществить эту мечту, наблюдая себя и современников, не представляется возможным. История технологического человечества занимает более 2,6 млн лет. За это время природные гоминины подверглись таким сложным превращениям, что истинная суть покрылась флером наивных мифов. Приведем красноречивый пример. Едва ли кто-то сомневается, что человек обязан интеллектом своему большому мозгу. При этом упускается из виду, что: homo ferus (дети, воспитанные животными), располагая большим мозгом, не имеют никакого интеллекта, человеческий мозг задействован не более чем на 10%, эпоха интеллектуального расцвета человечества (последние 2,5 тыс. лет) не сопровождалась изменениями головного мозга человека. Вывод очевиден. Размеры мозга не обусловливают интеллект. Они, скорее всего, развились у людей как побочный результат неотении. Без знания гоминизации этого вывода не сделать.
Тогда откуда же у людей интеллект? Чтобы ответить на этот вопрос, нам придется уяснить, какова природа технологического прогресса, чем полезен интеллект, помимо его изобретательности (присущей и животным), и как, наконец, людям удалось создать громаду земной цивилизации.
Глава 3
Человек
В данной главе мы предполагаем рассмотреть:
1) происхождение материальной культуры предков человека, основу которой составляли технологические средства поддержания жизни, способные к саморазвитию и обеспечению прогресса человечества, в то время как оно убеждено, что само продвигается вперед;
2) зарождение начал духовной жизни у наших предков (интеллектуального, религиозного, нравственного, эстетического), которые возвысили человечество над природой так искусно, что оно приписало это достижение самому себе;
3) становление цивилизации, кажущейся нам средством облагораживания человеческой жизни, а на деле стиснувшей человечество железными объятиями в интересах выживания социума.
3.1. Материальная культура
Технология
Единицы материальной культуры человека делятся на средства производительного и непроизводительного потребления, которое может быть индивидуальным и коллективным. Средство индивидуального производительного потребления – это, например, топор дровосека-одиночки, а средство коллективного производительного потребления – заводской конвейер или копья первобытного охотничьего коллектива, применяемые запланированно, «по номерам», в загонной или засадной охоте. Средство индивидуального непроизводительного потребления – это, например, личная одежда, а средство коллективного непроизводительного потребления – коллективное жилище.
Средства непроизводительного потребления не способны к самодвижению – они изменяются вместе с потребителем или по его воле, потому что сами по себе ничего не производят и не приспосабливаются к решению каких-либо задач. Средства производительного потребления меняются в зависимости от задач. Однако с их индивидуальными вариантами это происходит лишь по воле носителя. Средства коллективного производительного потребления ориентированы на усредненного носителя. Поэтому они менее эффективны, нежели индивидуальные средства, отличаясь от них, как серийные изделия от штучных.
Неповторимой чертой средств коллективного производительного потребления является их способность к самодвижению. Усредненно рассчитанные на способности коллектива, они не зависят напрямую от возможностей и воли отдельных его членов. При безразличии последних коллектив может меняться, вызывая перемены своих орудий, тогда как отдельные производители никак не меняются. Более того, им приходится подтягиваться вслед за уровнем развития своих коллективных орудий, и это наблюдается на протяжении всей истории человечества.
Представители нашего биологического вида заметно не меняются уже более 200 тыс. лет: их физическая организация и нейрофизиология все те же, что и на заре времен. Напротив, их предметный мир очень заметно менялся и перепрограммировал под себя собственных носителей. В этом проявилась способность средств коллективного производительного потребления к самодвижению.
Предметный мир, аналогичный материальной культуре человека, имеется и у животных, как позвоночных, так и беспозвоночных, независимо от уровня организации. Такие простейшие организмы, как амебы, одеваются в чехлы из песчинок (средство индивидуального непроизводительного потребления). Одиночная роющая оса Ammophila urnaria обустраивает норку камушком, точно молоточком, утрамбовывая им покрытие, т. е. применяя камушек в качестве средства индивидуального производительного потребления. Жук Salyarata приманивает высосанным термитом других термитов (то же средство индивидуального производительного потребления). Паук Dinopis ловит паутинным сачком муравьев (и это – средство индивидуального производительного потребления). Муравьи Aphaenogaster черпают и носят пищу кусочками листьев или почвы (опять же средства индивидуального производительного потребления). Галапагосские дятловые вьюрки обрабатывают и применяют веточки для извлечения личинок из-под коры деревьев (классическое средство индивидуального производительного потребления, присущее животным и вдобавок произведенное ими самими). От них не отстают «наивные юные вороны», тоже умеющие работать палочками и веточками. Изобретательные африканские стервятники ловко раскалывают камнями неподатливые страусиные яйца, а калифорнийские каланы применяют и сохраняют камни для расщепления раковин двустворчатых моллюсков (опять же это – средства индивидуального производительного потребления) [114, с. 217–218; 400; 458].
Муравейники, термитники, осиные и шмелиные гнезда, гидротехнические сооружения бобров относятся к средствам коллективного непроизводительного потребления. Таковы и подземные жилища сурикатов или сурков.
Шимпанзе обрабатывают и применяют прутики для ужения термитов, раскалывают камнями орехи (средства индивидуального производительного потребления). Продукты деятельности последнего рода (40 кг ореховой скорлупы и 4 кг камня, аналога режущих орудий ранних гоминин), подобно «археологической культуре», раскопаны в пункте Панда 100 леса Таи, Кот-д’Ивуар [568]. Орудийность шимпанзе сравнивается с уровнем культуры типичного олдовая [825]. В неволе шимпанзе-пигмей Кэнзи не только освоил язык символов, но и научился под руководством Н. Тота (специалиста по олдоваю и раннему ашелю) производить олдовайские отщепы [508, с. 38]. Как и павианы, шимпанзе способны камнями и палками коллективно отгонять хищников. Они умеют коллективно охотиться, правда без применения орудий [59, с. 38, 75, 172–174, 150, 199–202; 348; 362, с. 83–85; 508, с. 41; 517; 778; 800].
Однако ни у шимпанзе, ни у других животных коллективные орудия (средства коллективного производительного потребления) не известны [39; 58; 73; 110; 117]. Это обстоятельство вполне можно понять. Как отмечалось, коллективные орудия уступают индивидуальным в эффективности. Поэтому естественный отбор никогда бы их не разрешил, что и наблюдается в животном мире. Между тем, наши предки каким-то образом обошли запрет.
Легко представить, как гоминины разгоняли камнями и палками стада копытных, чтобы добраться до их телят. Это могло бы положить начало средствам коллективного производительного потребления (коллективным охотничьим орудиям). Однако, будь эти средства изначально выгодны, естественный отбор оснастил бы ими и других животных, склонных к орудийности и коллективности. Следовательно, первоначальная ценность коллективных орудий для гоминин состояла не в производственной эффективности.
Производительность коллективных орудий отвечает усредненному носителю. Поэтому она измеряется не качеством отдельного орудия, а эффективностью всего инвентаря, поделенного на количество носителей. В силу этого обстоятельства совокупный парк инструментов человечества, соотнесенный с его численностью, во все времена оставался примерно постоянной величиной. По мере роста населения прогрессировала сложность применяемых человечеством орудий.
Количественная связь народонаселения с технологией объяснима. Допустим, некий коллектив применяет технологию той или иной степени сложности и при этом доволен результатом. С сокращением коллектива сложность технологии становится избыточной, а при его росте – недостаточной. В результате при данной технологии коллективу выгодно поддерживать свою численность на постоянном уровне. По этой причине небольшие коллективы обычно пользуются простыми орудиями, а крупные – сложными, что наблюдается исторически в общечеловеческом масштабе (см. разд. 2.4).
Можно сформулировать своего рода демографо-технологическую зависимость. Между степенью сложности технологии и численностью практикующего ее коллектива существует общее количественное соответствие, характерное для уровня биопродуктивности данной коллективу экосреды [52, с. 25]. Оговорка насчет биопродуктивности среды нужна для объяснения нарушения зависимости к востоку от линии Мовиуса (см. далее).
В доступной анализу истории наши предки испытывали демографический рост и сталкивались с демографо-экологическими трудностями (см. разд. 2.4). Биология подталкивала их к слепому размножению. Поэтому ранним гомининам потребовалось искусственное средство координации своей численности. Те их сообщества, которым ок. 2,6 млн лет назад посчастливилось овладеть коллективными орудиями, подчинили свою демографию соответствию с наличной технологией. Прочие гоминины с течением времени вымерли.
Феноменологически (т. е. внешне) утверждение средств коллективного производительного потребления могло выглядеть так. Гоминины (предположительно некий «кениантроп»), не оставляя традиционной всеядности, сделались последовательными хищниками. Как и у шимпанзе, основу их добычи составляли обезьяны, мелкие антилопы и их детеныши. Эти млекопитающие способны на отпор. Поэтому наши предки логично перенесли на них стратегию борьбы с враждебными хищниками. Они отгоняли взрослых особей палками и камнями, чтобы поживиться детенышами. Справедливости ради следует признать, что подобная модель совместима с некрофагической гипотезой становления плотоядности у гоминин (см. разд. 2.4). Они могли бы камнями и палками отгонять хищников от добычи. Однако, в отличие от охоты, такая практика ничем не засвидетельствована, и мы не видим оснований для ее признания.
Введя совместную орудийность в традицию, применив умение раскалывать орехи камнями для добычи костного мозга из трубчатых костей и головного – из черепов своих жертв и приступив к избирательному применению камней, что побуждало к их обработке, «кенийцы» запустили технологическую историю, однако столкнулись с обременительным и поначалу не окупаемым занятием. На это пошел только один размножившийся и акселерированный вид (предположительно Kenyanthropus rudolfensis). Созревая раньше своих предков, этот гоминин вступал в пору зрелости психологически инфантильным, а потому не держался за традиции и был эволюционно лабильным (нестойким) в поведении. У любого другого млекопитающего коллективная вооруженная охота была бы пресечена селекцией. У «кенийцев» это малопродуктивное занятие не способствовало расширению сообществ (избыток населения отселялся). Поэтому в данном конкретном случае отбор благоволил освоению средств коллективного производительного потребления.
Первые каменные орудия типичной олдовайской культуры [99; 491] были открыты в Када Гона, Хадар, Афар, Эфиопия, св. 2,63 ± 0,5, 2,58 ± 0,23 млн лет назад [500, с. 463]; в Оунда Гона, там же, 2,58–2,53 ± 0,15 млн лет назад [685]; в Гона, там же, 2,58–2,52 млн лет назад [684]; в Локалалеи, Начукуи, Туркана, Кения, 2,34 ± 0,05 млн лет назад [652], и в других местах. С временнóй отметки в 2,3 млн лет назад в Хадаре эти орудия связываются с Kenyanthropus rudolfensis [819]. Попытки приписать первую технологию Australopithecus garhi (ввиду наличия 3 порезов на сопровождающей его остатки челюсти четвероногого [408, с. 627]), а орудийный точный захват – Paranthropus robustus [737; 738]; cp. [398; 599; 758] не получили развития. Орудийность «кенийца» гармонирует с его человекоподобием (например, это очень выражено у черепа KNM-ER 1470, гладкий лоб которого указывает на неотеничное происхождение, а относительно крупные размеры – на акселерированность).
История «кенийца» повторилась у Homo ergaster ок. 1,9 млн лет назад. Первый человек перенес демографический взрыв и акселерацию, распространился по Старому Свету и ок. 1,6 млн лет назад в состоянии поведенческой податливости совершил ашельскую технологическую революцию. Орудия олдовая были обработаны односторонне. Обработанные с двух сторон орудия ашеля [491; 519] были технологически сложнее и разнообразнее, а потому отвечали новому демографическому состоянию. Ранний ашель начинается с Олдувая, Танзания, 1,6–1,5 млн лет назад [286; 684]; с Консо-Гардула, Эфиопия, 1,43 ± 0,02–1,41 ± 0,02 млн лет назад [153, с. 732, табл. 1, с. 733, рис. 1; 741, с. 490]; с Убейдия, долина Иордана, Израиль, ок. 1,4 млн лет назад [656; 823]. «Мастеру» же принадлежит освоение таких деликатных орудий, как зубочистки (Омо, Эфиопия, 1,84 млн лет назад [161]), и такой грозной стихии, как огонь (Кооби Фора, Кения, ок. 1,6 млн лет назад; Чесованджа, Кения, ок. 1,4 млн лет назад [557]; Сварткранс 3, ЮАР, 1,5–1,0 млн лет назад [206]).
Следует отметить, что ашель не изобретался, а, наоборот, долгое время игнорировался. Ядрища (нуклеусы), с которых скалывались олдовайские отщепы, в результате этой сырьевой процедуры абсолютно непреднамеренно принимали облик будущих двусторонних орудий ашеля, бифасов. Применительно к олдоваю их называют протобифасами [491]. Показано [99], что протобифасы были не самостоятельными орудиями, а подручным материалом и лишь потому переносились с места на место. Иными словами, готовый ашель всегда находился у «кенийцев» и «мастеров» под руками, однако те совершенно не осознавали, что уже владеют революционной технологией. В соответствии с демографо-технологической зависимостью она пошла в серию лишь с демографическим взрывом у раннего питекантропа.
Продвижение африканских культур в Европу никогда не встречало препятствий. Однако в восточном направлении они останавливались на линии Мовиуса, которая разграничивала по северо-восточным рубежам Индии ашельский Запад и олдовайский Восток [587, с. 100–104; 730, с. 36]. Предполагалось [630], что вместо бифасов на Востоке применялись орудия из бамбука. Отмечено, однако [55, с. 101–102], что в олдовайской Сибири бамбук не растет, а потому технологическая застойность за линией Мовиуса должна иметь какое-то другое объяснение.
С точки зрения демографо-технологической зависимости, при оценке плотности населения надо учитывать биопродуктивность наличной среды. Ашельцы, мигрируя из Африки в Европу, теряли исходную плотность населения, однако попадали в менее продуктивную среду, богатство которой в расчете на душу поредевшего населения выглядело привычным. Поэтому при переносе африканских культур в Европу демографо-технологическая зависимость выдерживалась без нарушений.
Напротив, при транспортировке африканских культур из Передней Азии на восток плотность населения мигрантов падала по дороге, в районе Индии они попадали в тропическую среду, где биопродуктивность в расчете на душу населения начинала мнимо превосходить африканскую и в соответствии с демографо-технологической зависимостью переставала нуждаться в сложной технологии. Поэтому мигранты, проходя Индию и по инерции оставаясь там еще ашельцами, утрачивали ашель на северовосточных рубежах и появлялись в Индокитае и Китае как олдовайцы. Заселение Сибири осуществляли дальневосточные монголоиды, имевшие индийское происхождение (см. разд. 2.4). Согласно изложенной модели, они должны были прибывать в Сибирь как носители простых технологий. То же самое происходило с индийскими по происхождению австралоидами, которые закономерно прибывали в Индонезию как носители «допотопных» (галечных) орудий – например, на остров Калимантан, в пещерное местонахождение Ниах грейт кейв, св. 41 500 ± 100 лет назад. Между тем, с ростом населения в Южном Китае, когда биопродуктивность среды в расчете на душу населения упала до африканских норм ашеля, там развилась собственная ашельская культура (бассейн р. Боcе, Южный Китай, 803 ± 3 тыс. лет назад [424]).
Прецедент «кенийцев» и «мастеров» возобновился с демографическим взрывом у европеоидов современного вида на Ближнем Востоке ок. 50 тыс. лет назад (см. разд. 2.4). Они произвели верхнепалеолитическую технологическую революцию, пустив на поток пластинчатую индустрию ориньякского типа. Тонкие ножевидные пластины изобильно снимались с подготовленного призматического нуклеуса и превращались в великолепные инструменты, отличающиеся не только красотой, но и производительностью, поскольку ориньякская культура была сложнее и богаче предшествующих.
О сознательном изобретении пластинчатой технологии также не может идти речи. Пластины известны со времен позднего «кенийца». Две пластинки представлены в местонахождении Омо 123 К, Эфиопия, 2,06/ 1,99–1,93 млн лет назад [500, с. 774]. Пластины входили постоянным элементом в состав мустьерской культуры [198; 566], датирующейся в Европе 333–32,5 тыс. лет назад. Они были присущи среднему каменному веку формации Каптурин, Баринго, Кения, 240 тыс. лет назад [392], откуда, как представляется, перекочевали в ближневосточный преориньяк, 144–110 тыс. лет назад (однако, как ни странно, их связь с ближневосточными протокроманьонцами пока не установлена: они пользовались мустьерской индустрией).
Европейский верхний палеолит рухнул вместе со своим предметом труда – холодолюбивой мамонтовой фауной, не перенесшей потепления в начале голоцена (см. разд. 2.3). Поясним, что с социально-философской точки зрения животные, служащие объектом охотничьего промысла, считаются предметом охотничьего труда. Вместе с крупными приледниковыми животными ушло блистательное искусство наскальной живописи (см. разд. 3.2). В регионах, где фауна не была столь приспособлена к низкой биопродуктивности среды, аналогичных потрясений не случилось, хотя охотники нашего вида промышляли с европейской жадностью, что доказывается примером исторически моментального истребления гигантских птиц моа в Новой Зеландии (см. разд 2.3). Автоматизм гибели верхнепалеолитической культуры косвенно свидетельствует в пользу ее автоматического формирования.
Примерно такое же автоматическое возникновение технологии произошло в эпоху неолитической революции производящего хозяйства 11,7 тыс. календарных лет назад, последовавшей за демографическим взрывом 16 тыс. календарных лет назад (см. разд 2.4). Неолитическая революция ознаменовалась одомашниванием сельскохозяйственных животных и растений. Это выдающееся технологическое достижение не должно считаться изобретением.
Мезолитические аборигены Австралии разволят ямс, дающий крупные съедобные клубни [11, с. 76], а южноамериканские индейцы порой выращивают детенышей диких съедобных животных. Лошадь и северный олень, возможно, приручались еще в верхнепалеолитической Европе [66, с. 28–33]. По молекулярно-генетическим данным, предки всех домашних лошадей имеют возраст ок. 320 или даже ок. 630 тыс. лет [776]. Однако бесспорные археологические свидетельства верховой езды относятся к древнехалколитической (древнемеднокаменного века) культуре Средний Стог (Украина – юго-запад России, 7180–6260 лет назад) [121]. Молекулярно-генетическая древность домашних собак достигает 135 тыс. лет [777]. Столь почтенный возраст объясняет происхождение собак динго, явившихся в Австралию под водительством первых аборигенов 61±13 тыс. лет назад (см. разд. 2.4). Близкие к китайским волкам, эти животные отмечены в Восточной Азии ок. 40 или ок. 15 тыс. лет назад, в Германии – 14 тыс. лет назад, в Израиле – 12 тыс. лет назад [582; 671].
Из сказанного следует, что навыки доместикации животных и растений имелись у людей и до неолитической революции, однако хозяйственно не применялись. Преднеолитический демографический взрыв востребовал их в соответствии с демографо-технологической зависимостью. Примерно в то же время демографический подъем происходил в Восточной Азии, где производящее хозяйство зародилось автономно, т. е. до прибытия с запада носителей сино-кавказских языков (чеченский, кетский, надене и др.), положивших начало сино-тибетским (китайский, бирманский и другие языки Центральной, Юго-Восточной и Восточной Азии).
Западноевропейская промышленная революция, развернувшаяся с XIV в., происходила по описанному сценарию. Характерные для нее машины были известны еще античным грекам, которые применяли их в театре. Герон Александрийский создал паровой двигатель в I в. Раймунд Луллий (ок. 1235 – ок. 1315) построил первую «логическую машину», зачаточный прообраз компьютера. Эти изобретения ни для кого не являлись тайной, однако принципиально родственные им машины «пошли в тираж» лишь после начала современного демографического взрыва в Западной Европе (XI – середина XVI вв.).
Густо населенная Азия (Индия, Китай, Япония) не случайно отставала от Западной Европы. Ее биопродуктивность превосходит западноевропейскую. Поэтому из-за расхождений биопродуктивности в пользу Южной и Восточной Азии последняя технологическая революция должна была начаться в Западной Европе раньше и с меньших плотностей населения, нежели в густонаселенных азиатских регионах. Там высокая биопродуктивность среды, поделенная на численность населения, опережала западноевропейский уровень, а потому не создавала условий для революционных перемен в технологии, как следует из предложенной нами демографо-технологической зависимости (см. выше). Проще говоря, в «райских» условиях можно и подождать с внедрением новой техники. Поэтому демографо-технологическая зависимость претворялась в Азии нестандартно: ашель пробился позже, чем на Западе; производящее хозяйство, правда, зародилось одновременно с ближневосточным неолитом, но вот начала промышленной революции привились лишь в ХХ в. (при этом передовые технологии – от производства бумаги до зачатков ракетной техники – были представлены еще в средневековом Китае, но не влияли на умы в смысле революционизации общего технологического состояния, как это было в Европе).
Демографо-технологическая зависимость действовала не только в среде гоминин человеческой филетической линии: 125 тыс. лет назад демографический взрыв охватил популяции европейского Homo heidelbergensis, превратив его прогрессивно-неандерталоидный вариант в акселерированного классического неандертальца (см. разд. 2.4). Демографический подъем потребовал усложнения технологии, и неандертальцы обнаружили, что давно производят в рамках своей мустьерской культуры изделия на пластинах, отвечающие более высокому уровню верхнепалеолитической техники. Соответствующие образцы были «пущены в серию», что привело к становлению археологической культуры перигор 0–1 (шательперрон), которая непосредственно связана с неандертальцами во французских местонахождениях Арси-сюр-Кюр (33 860 ± 250 лет назад) и Ле Котте (33 300 ± 500 лет назад) [569].
Демографо-технологическая зависимость позволяет объяснить природу ускоряющегося технического прогресса человечества. Каждая технологическая революция сопровождалась расширением орудийного инвентаря. В соответствии с законами технологии обогащение инструментария сопровождается сужением функций орудий, их специализацией. Специализированные орудия более производительны, чем неспециализированные. Поэтому всякая технологическая революция ознаменовывалась скачком производительности труда.
Технологические революции следовали за демографическими взрывами, а демографический рост происходил экспоненциально, с ускорением. Поэтому следующий за ним технологический прогресс шел также ускоренно. Творческое начало человечества играло при этом вспомогательную роль, хотя люди самолюбиво привыкли иначе оценивать ситуацию.
Социальность
Демографо-технологическая зависимость проливает свет на природу человеческой социальности. Происхождение кровнородственных отношений у людей имеет биологический характер. В среде высших коллективных животных действует закон Крука [265–267]. Согласно этому закону, структура сообществ высших коллективных животных зависит от биопродуктивности среды. Биопродуктивность эта минимальна, например, в пустынных высокогорьях. Поэтому у населяющих их обезьян гелад лишние самцы изгоняются из стада как ненужные рты, а победившие самцы-лидеры обзаводятся гаремами. То же бывает характерно и для людей, например, для автохтонов Аравийской пустыни, древних евреев и арабов, у которых издавна имелись гаремные семьи. Пустынные регионы порождают у обезьян патрилинейность, а у людей – патриархат, что в обоих случаях одинаково означает господство сильного пола.
В максимально биопродуктивных регионах, в тропическом лесу, пищи хватает всем. Поэтому второстепенные самцы не изгоняются, и доступ к самкам имеет все мужское поголовье. Там господствует промискуитет (свободная любовь), как, например, у шимпанзе [59, с. 135]. У обезьян ранг особи в стаде наследуется по материнской линии. Поэтому для лесных шимпанзе характерна матрилинейность (высокий статус самок, особенно приближенных к вожакам), а для человеческих сообществ в высокобиопродуктивных регионах – пережитки матриархального промискуитета при высоком значении женщин, особенно приближенных к вождям (например у древних эламитов на территории нынешнего субтропического Ирана). Это объясняет и существование пережитков матриархата в высокобиопродуктивных тропических джунглях Вьетнама: подобное социальное устройство представляет собой аналогию матрилинейным общественным связям шимпанзе в тропических джунглях Африки. В средах с промежуточной биопродуктивностью обезьянам и людям присущи переходные варианты кровнородственных отношений между матрилинейной свободой и патриархальной строгостью.
Сравнивая общественный образ жизни у людей и у других приматов, мы вовсе не предаемся биологизаторству. Научный подход к вопросу требует учитывать то элементарное обстоятельство, что человек является все же природным существом, пусть и связанным с колоссальной цивилизацией (см. разд. 3.3). Природные задатки в нас по-прежнему сильны, и мы продолжаем свой род вполне биологическим путем. Коль скоро это так, то подчинение закону Крука отнюдь не должно нас унижать.
В свете закона Крука поведение обезьян объясняет генезис широкого спектра кровнородственных отношений людей: от гаремной семьи у выходцев из Аравийской пустыни с ее минимальной биопродуктивностью до свободной любви (аналога промискуитета) в благополучных странах. Поведение обезьян объясняет и происхождение обусловленных кровнородственными связями подсознательных комплексов, выявленных З. Фрейдом (1856–1939) у современных людей.
Возьмем, например, эдипов комплекс, т. е. инстинктивное стремление молодого человека к близкородственным брачным отношениям (инцесту) с властной женщиной (матерью) при ненависти к отцу. Объяснение подобных побуждений лежит в повадках обезьян. В их стадах доминирующее положение самцов нередко передается по линии господствующей самки-матери, откуда возникает тяга к тесным (сексуальным) связям с ней, т. е. к инцесту. При этом молодежь мужского пола не прочь избавиться от лидирующих самцов старшего поколения, т. е. от своих отцов, чтобы занять их место в стадной иерархии. В сумме у молоди обезьян складывается благоприятная почва для эдипова комплекса, который достался нашим соплеменникам от далеких предков. У нормальных людей этот комплекс обуздан воспитанием – у невоспитанных дает о себе знать. Нейрофизиология подобных людей необязательно паталогична, так что помощь психоаналитика может оказаться небесполезной.
Помимо отношений, связанных с продолжением рода, людям свойственны производственные отношения, создающие каркас исторических обществ. Альтруистическое сотрудничество у людей не может быть результатом биологического группового отбора [317–320]. Групповой отбор – это разновидность естественного отбора, когда природа заботится не о выживании отдельной особи, а о сохранении генотипа, общего группе родичей. Во имя его выживания отдельная особь способна идти на существенные самоограничения и даже самоотверженно жертвовать жизнью ради своих родственников. Она погибнет, но ее генотип уцелеет у родственных собратьев. Заманчиво было бы считать, что человеческое трудовое сотрудничество зародилось в результате группового отбора родичей. Это, однако, упрощенный взгляд на вещи, поскольку для людей весьма характерно сотрудничество неродственных индивидов.
На наш взгляд, человеческая социальность имеет производственное происхождение. Демографо-технологическая зависимость, связывая людской коллектив с технологией, тем самым цементирует его технологически. Солидарность людей на почве демографо-технологической зависимости представляется основой социальности. Она имеет ту же древность, что и технология у гоминин. Проще говоря, демографо-технологическая зависимость сбивает тружеников в тесный коллектив. Его рамки ограничены не общественными инстинктами, как у других животных, а степенью сложности наличной технологии, что по определению является небиологическим фактором. Вот почему человеческая социальность отличается особой, не биологической, а технологической природой. Чем значительнее производственные технологии, тем крупнее и сложнее устроено общество. Это обстоятельство объясняет историческую последовательность трех основных типов социальной организации: первобытное общество (исходное), рабовладельческое и феодальное общество личной зависимости людей друг от друга (после неолитической революции), капиталистическое общество опосредованной, денежной зависимости людей от себе подобных (после промышленной революции). Изложенное понимание социальности избавляет нас от неконкретных споров о ее метафизической, философской природе, поскольку имеющиеся факты ставят проблему на вполне твердую почву.
Если человеческая социальность имеет технологическое происхождение, то она никогда не зависела от воли и сознания человека. Отсюда следует, что всякая социоинженерная деятельность людей может лишь подкорректировать автоматически складывающиеся человеческие отношения, но не сконструировать их произвольно, на что во все времена рассчитывали утописты.
3.2. Духовная культура
Основу материальной жизни общества составляют добыча средств к существованию, производство потомства и занятия, помогающие достижению этих целей. У шимпанзе, горилл и павианов, поведенчески близких нашим предкам, аналогичные задачи отнимают почти все активное время. У мезолитических бушменов Южной Африки и австралийских аборигенов, охотников и собирателей, сходным нуждам посвящено уже лишь 25% активного времени, а у неолитических папуасов Новой Гвинеи, примитивных земледельцев и скотоводов, – всего 10% [12, с. 491; 47, с. 151–153; 504; 810].
Уже 2,5 млн лет в основе социальности гоминин лежат материальные отношения, связанные с добычей средств к существованию. Это занятие мотивирует саму нужду в социуме. Между тем, есть основания считать, что каждая технологическая революция в истории сопровождалась подъемом производительности труда. Если наш предок, скажем Kenyanthropus rudolfensis, начинал свой исторический путь в добыче пищи с производительности, сопоставимой с таковой у шимпанзе, то после ашельской технологической революции (если не раньше), когда производительность труда повысилась, на создание средств к существованию у Homo ergaster стало уходить гораздо меньше времени.
Как следствие, часть активного времени сделалась праздной, и доля свободного времени в распорядке жизни у гоминин продолжала возрастать с каждым достижением в подъеме производительности труда. У наших предков неизбежно складывалось впечатление, что их напряженная социальность не требуется для выживания. Из обыденного опыта общеизвестно, что бестолковая праздность разлагает всякое человеческое существо и любое общество. В результате социальность наших предков оказалась под угрозой. Благополучнее всего ее пережили те сообщества гоминин, которые научились занимать праздное время общением непроизводственного типа. Менее удачливые сообщества распались и вымерли. Для создания непрагматических социальных связей у гоминин имелось немало биологических предпосылок, которые есть и у животных, но не развиваются без особой нужды.
Интеллект
У шимпанзе имеется набор жестов, гримас и поз, а также 23 вида различных голосовых сигналов, при помощи которых они общаются [50, с. 64]. Опыты с самкой шимпанзе Уошо показали, что эти приматы относительно легко усваивают человеческий язык жестов амслен, язык символов и арифметический счет [69; 340; 413; 549]. При этом усвоение идет лучше всего, когда пальцы обезьяне просто складывают в нужный жест. Легко представить, как орудийные «кенийцы», обучая друг друга технологическим приемам, оттачивали производственные жесты партнеров и тем самым невольно формировали первый словарь технологического языка жестов. Это достижение Kenyanthropus rudolfensis предопределило функциональное устройство коры человеческого головного мозга.
Человеческий мозг функционально асимметричен [28; 275; 364]. Правой рукой управляет моторная кора левого полушария головного мозга. Нижняя оконечность этой коры, руководящая пальцами руки, лицом и языком, разделяет прилегающие зоны Брока (спереди) и Вернике (сзади), которые командуют речью. Подобным соседством обусловлена наша назойливая жестикуляция при разговорах. Причина объясняется тем, что первоначально речевые зоны Брока и Вернике обслуживали движения правой руки и координировали язык жестов. Подобная функция закрепила за этими зонами пилотаж грамматических структур, которые были перенесены на зарождающийся голосовой язык, чья моторика обеспечивается нижней перемычкой между зонами Брока и Вернике [52, с. 43; 53, с. 181; 262].
Судя по образцу KNM-ER 1470, финальный Kenyanthropus rudolfensis уже обладал развитыми полями Брока и Вернике [18, с. 200; 831], а потому владел по меньшей мере языком жестов, но, быть может, и вербальной речью. Возможно, он был уже праворуким, а его потомок, ранний питекантроп, являлся таковым наверняка [99, с. 86; 205, с. 48]. Происхождение звуковой речи можно объяснить следующим образом.
После ашельской технологической революции у Homo ergaster выросла производительность труда и образовалось свободное время, которое требовалось заполнить общением непрагматического свойства, дабы поддержать целостность сообществ раннего питекантропа. Выжить посчастливилось тем, кому это удалось. Для выполнения данной задачи «человек-мастер» принялся скрашивать досуг обыденными разговорами – эта привычка характерна и для нас. Тематика таких бесед несущественна – лишь бы она находила слушателей, создавая эффект непроизводственного общения. Первые непринужденные разговоры (т. е. не посвященные производственным темам), надо полагать, были житейскими. Придерживаясь той же традиции, австралийские аборигены любят проводить время в сплетнях и скандалах [11, с. 260]. Современный цивилизованный человек, пожалуй, может поспорить в этом с австралийцами.
Внешняя непритязательность подобного занятия скрывает за собой важную социализирующую функцию. Кроме того, исходные бытовые разговоры послужили основой для формирования интеллекта человеческого типа. По сравнению с интеллектом животных его отличительная черта состоит в праздности. Животные интересуются тематикой, так или иначе важной для выживания. Человек же интересуется чем угодно и благодаря этому обладает универсальным мировоззрением. Происхождение столь «завидного» достижения коренится в первобытных сплетнях (мы, правда, несколько утрируем ситуацию).
Древнейшие разговоры родились как способ насыщения свободного времени общением. По этой причине исходный язык жестов сосуществовал со звуковым и дожил до верхнего палеолита европейских кроманьонцев, нередко оставлявших на стенах украшенных пещер отпечатки раскрытых ладоней или ладоней с поджатыми пальцами, символизирующих определенные высказывания языка жестов (см. приложение 1). По мере роста производительности труда праздного времени становилось все больше, и разговоры требовали все новой тематики. Она расширялась за счет всего, на чем останавливался взгляд гоминин, а он останавливался на множестве попадающихся на глаза объектов. Зрелые животные никогда бы не позволили себе такой расточительности, так как естественный отбор требовал от них концентрации внимания на биологически существенных предметах. История же гоминин сложилась так, что невозможные для нормальных животных праздные разговоры шли им только на пользу. Именно это социологическое, а не нейрофизиологическое (как ошибочно думают) обстоятельство обеспечило интеллектуальную пропасть между людьми и животным миром.
Условно говоря, человечество можно представить как живой суперкомпьютер, тысячелетиями набирающий базу произвольных данных. Его кругозор бесконечно превосходит животную память не в силу изощренного устройства, а в силу того, что борьба с праздностью требовала от гоминин впитывать из действительности любую сколько-нибудь приемлемую тему для беседы. Применительно к отдельному индивиду подобная картина выглядит преувеличением, но относительно социума с вековой историей она не так уж и далека от истины.
Животные способны к обобщениям прошлого опыта и предвидению будущего на его основе. Это умеют даже геномы организмов (см. разд. 2.3) – в противном случае они попросту не выжили бы в изменчивых условиях нашей планеты. Однако материал для обобщений у них жестко ограничен соображениями биологической выгоды, как бы широко ни понимать ее. Гоминины оказались в биологически парадоксальной ситуации, когда ради заполнения свободного времени естественный отбор позволил им выходить за рамки биологической целесообразности при накоплении информации. В результате социум как живой суперкомпьютер принялся механически открывать и предсказывать сущности вещей, далеких от нужд животного прагматизма. Следует сказать, что выявление сущностей вещей не является интеллектуальным священнодействием, доступным одним лишь рафинированным философам. Трудность эвристических задач здесь носит естественнонаучный характер. Поясним сказанное.
С точки зрения количественной теории информации [116], какое-либо количество информации содержится только в новых сведениях. Именно поэтому нам неинтересно выслушивать одно и то же. В силу той же причины отдыхом для нас служит смена деятельности. Теперь посмотрим, что представляют собой явления и сущности с информационной точки зрения.
Сущности выражают устойчиво повторяющиеся черты явлений, а всякое явление, воплощающее сущность, содержит случайные особенности, отличающие его от всех прочих родственных явлений. Иными словами, совокупность объектов данного класса (например бриллиантов) выражается соответствующим числом явлений и стоящих за ними сущностей. Но сущности при этом одинаковы (это бриллиант вообще), а явления различаются (по каратам, «воде», огранке и другим неповторимым мелочам). В соответствии с теорией информации мы воспринимаем все эти бриллиантовые (в данном примере) сущности как одно и то же, как одно сведение, как одну единицу информации. Неповторимые же явления мы видим каждое особо – как отдельную единицу информации. Получается, что для нас в любой совокупности объектов видится некоторое число явлений и только одна сущность. Неудивительно, что сущность теряется среди явлений, заслоняется ими и порождает наши философские представления о своем трансцендентном («потустороннем») бытии.
Последнее не соответствует действительности: на деле сущности пребывают в одном и том же измерении с явлениями, только в отличие от них по информационным причинам ускользают от нашего внимания. Когда за восприятие реальности берется крупный цивилизованный социум, всякая сущность становится достоянием большого числа наблюдателей, в силу чего приобретает несвойственную себе наглядную массовидность и выходит из тени множества явлений, поскольку идентичность массовидных сущностей (в противовес неповторимым явлениям) поражает человеческое воображение. Разумеется, социум в целом не занимается отслеживанием сущностей. Однако на заре науки он позволил их обнаружить и предоставил ученым методику «охоты» за ними – собственно философию и науку.
Толика данной способности доставалась и отдельным субъектам. Именно ее принято расценивать как специфически человеческий интеллект. Его генезис не был обусловлен непосредственной биологической целесообразностью – иначе интеллект человеческого типа широко распространился бы по животному миру. Коллективный разум гоминин позволял им сохранять свои сообщества от разрушительных угроз свободного времени и в этом отношении выполнял полезную функцию, чего не требовалось иным животным. В конце концов интеллект гоминин оказался вооруженным массой полезных внебиологических знаний, однако этот результат является побочным, и остальным представителям животного мира он недоступен.
Человеческая мысль связана с языком [72, с. 29]. Даже когда мы размышляем про себя, у нас неслышно колеблются голосовые связки. Эта реакция является пережитком тех времен, когда речь и мышление существовали только во внешнем варианте, были рассчитаны на слушателей, а потому обладали сугубо коллективной фактурой. Индивидуальное мышление современного типа оформилось лишь в эпоху цивилизации (см. разд. 3.3).
Механизм современного мышления [439] основан на собеседовании полушарий головного мозга: речь понимают оба, но разговаривает лишь левое (у правшей). Их разговоры создают впечатление присутствия у нас в голове второго Я. У первобытных людей навык внутренней речи не был развит. Поэтому, например, американские индейцы, готовясь к выступлениям на собраниях, проговаривали заготовленные речи вслух. Цивилизованные люди в преклонном возрасте тоже переходят на размышления вслух. Неслышная внутренняя речь и мысль были освоены только с приходом цивилизации.
Тогда появился правящий класс профессиональных администраторов. Руководя всем обществом разделенного труда, администраторы должны были управлять и собой, т. е. саморегулироваться. Направленные на самих правителей регулярные приказы послужили им моделью самосознания. Первоначально оно появилось у руководящей верхушки, а затем распространилось на другие слои общества, которые во все времена преданно заимствовали новации элиты.
Предметной формой самосознания послужила письменность. На Ближнем Востоке письменность шумерского круга (протошумерская иероглифика и шумерская клинопись) происходила от оттисков предметного письма в виде «жетонов» [675; 676]. Эти «жетоны» появились еще в IX в. до н. э. (Тепе Асиаб, Иран; Бельдиби, Турция) и распространились со временем от Джейтуна в Туркмении до Хартума в Судане и от Бельдиби в Турции до Чанху Даро в Пакистане. Они представляли собой глиняные геометрические фигурки разных форм, типы которых отвечали тем или иным товарам. Их запечатывали в полые глиняные шары (булы) и отправляли в качестве накладной с торговой партией. На наш взгляд, они служили предметной формой эсперанто (т. е. предметным письмом, понятным разным народам), что обеспечивало международную торговлю в обширном ближневосточном регионе с Месопотамией в центре. В раннем Шумере очертания их оттисков стали обслуживать на письме шумерский язык.
Самосознание поставило человека вне рамок животного мира, и вот почему. Поведение животных управляется инстинктами и рефлексами, побуждениями и эмоциями. Оно может быть изощренным, хитрым, альтруистичным, но всегда остается эмоциональным, а потому типологически одинаковым у всех представителей данного вида, ибо природа награждает их одними и теми же эмоциями. Все это свойственно и людям. Поэтому, например, эмоциональный настрой толпы очень однороден и носит название «стадного чувства».
Однако, обзаведясь самосознанием, цивилизованный человек попал в доселе не виданное положение среди животных. Теперь он получил возможность командовать самим собой при помощи слов внутренней речи, т. е. использовать свои ум и тело, свое второе Я как отдельное от сознания устройство. Слова можно складывать в любые приказы: и нелепые, и героические, и мудрые… По этой причине люди отличаются умением совершать не слыханные в животном мире глупости (например, ни с того ни с сего отписать все состояние чужим людям). Или, наоборот, способны совершать беспрецедентные героические поступки, недоступные животным.
Соответствующие факты известны уже в мире первых цивилизаций. Так, 5-й фараон XVIII династии Египта Тутмос III (1490–1437 до н. э.), освободившись от опеки властной тетки-мачехи, женщины-фараона Хатшепсут (1490–1469 до н. э.), в 1468 г. до н. э. осуществил военный поход в Палестину на объединившихся против него местных правителей. Дорога к городу Мегиддо шла через узкое ущелье, где колесницы могли следовать лишь гуськом и стать легкой добычей затаившегося на выходе противника. Вопреки уговорам осторожных военачальников Тутмос III приказал начать движение, поразил отвагой воображение врагов и разбил их наголову под стенами города Мегиддо. Битва осталась в памяти человечества под библейским названием Армагеддон («гора Мегиддо»), которое стало обозначением непримиримой схватки перед концом света.
От Тутмоса III не отстал 3-й фараон XIX династии Рамсес II Великий (1279–1213 до н. э.), который прославился беспрецедентной битвой при сирийском городе Кадеше (1274 до н. э.). Коварные хетты заманили фараона с телохранителями в ловушку, окружили 3 тысячами колесниц и предложили сдаться. Рамсес II сопротивлялся до тех пор, пока не подошли его войска.
Спартанский царь из династии Агиадов Леонид I (490–480 до н. э.) действовал в том же духе. С 300 телохранителями он до последнего защищал Фермопильское ущелье от персов и безо всякой надежды на успех не сдавался, пока не погиб вместе со всеми своими бойцами.
В животном мире подобное поведение исключено. «Загадочная человеческая душа» объясняется тем, что она управляется не только эмоциями, но и словами в рамках самосознания, чего лишены животные. Академик И.П. Павлов (1849–1936) не без основания видел наше расхождение с животными в наличии у нас 2-й сигнальной системы, словесной, хотя в его время и недоставало данных для правильного истолкования этого феномена.
В итоге человек, в принципе, способен на произвольное поведение, не ограниченное инстинктами, преломляющимися в голове как эмоции. Поэтому, с точки зрения наблюдающих нас животных, мы выглядим алогичными чудовищами, с которыми лучше не связываться. Именно по этой причине человек не входит в список добычи крупных хищников. На нас охотиться побаиваются (исключение составляют раненые или больные хищники, которым другая добыча не доступна и выбирать не приходится). Отсюда следует, что человек – это не «говорящий биологический автомат» [53], раз он способен отдавать себе и окружающим произвольные приказы, не отягощенные инстинктами.
В эпоху ранней цивилизации появился рафинированный интеллект, оформленный в виде институционализированной науки. Возникновение последней обусловлено двумя факторами. Правители, будучи деятелями умственного труда, нуждались в школах, готовящих новую смену и уподобляющих учеников друг другу в противовес последствиям их воспитания в различных подразделениях труда, как и в наше время.
Вопреки очевидности, происхождение общеобразовательной школы не столь банально, как может показаться. В самом деле, высшие животные обучают детенышей всему, что знают. Не удивительно, что, следуя той же модели, цивилизованный человек учредил для детворы общеобразовательную школу. Древнейшая общедоступная светская школа é-dub-(b)a переживала расцвет ок. 1800 г. до н. э. Она была основана в Шумере примерно в 3000 г. до н. э. [40, с. 61–63].
Для шумерского общества это было время состоявшегося разделения труда. В каждом подразделении бытовали свои цели, интересы и представления о действительности, которые, усваиваясь молодыми поколениями, все больше усугублялись. Проще говоря, подразделения труда становились все более непохожими друг на друга и все менее сочетаемыми в рамках единого общества. Это угрожало целостности социума, и он не мог этого терпеть. Сперва он запер общества разделенного труда в тесные города, чтобы они (общества) не развалились, и произвел их в ранг цивилизаций (см. разд. 3.3).
После этого на повестку дня был поднят вопрос о возможной унификации сознания цивилизованных людей. Для этих целей в ход пошла общеобразовательная школа. Обучая представителей различных подразделений труда одним и тем же общеобразовательным предметам, школа сближала их сознания вопреки узкопрофессиональным представлениям, усвоенным дома (представлениям конкретных подразделений труда). Тем самым школа цементировала общество своими средствами. Эта функция сохраняется за ней и поныне.
Следует понять, что программа общеобразовательной школы, не являясь специализированной, не может быть эффективной В обществе разделенного труда лучше всего учить молодые поколения профессиональным навыкам. Это, конечно же, так. Однако в этом случае общество настолько расслоится по профессиям, что станет внутренне несочетаемым и рассыплется. Именно поэтому выстояли те цивилизованные общества, которые пошли и идут по пути неэффективной общеобразовательной школы (неэффективной с точки зрения узких нужд подразделений труда). Поэтому, когда нам кажется, что многие школьные предметы нам в жизни не нужны, мы просто не понимаем истинное положение вещей, как оно видится с точки зрения социальной философии.
Цивилизованное общество обладало высокими плотностями населения (около 10 тыс. человек и выше). В столь многочисленных сообществах отчетливо проявляется действие статистического закона больших чисел, в соответствии с которым крупные совокупности объектов ведут себя предсказуемо (их проявления близки своему математическому ожиданию, вероятности).
Цивилизованное общество, став предсказуемым в социальном смысле, приобрело новые свойства как мыслящее целое. К нему пришла способность различать устойчиво повторяющиеся и случайные связи вещей (т. е. открывать и предсказывать сущности в обозримом времени). Это происходило независимо от пожеланий отдельных людей. Следуй наука их чаяниям, она стала бы прагматически ориентированной, предельно полезной людям. На деле ранняя наука отличалась непрагматичностью [40, с. 75]. К примеру, вместо насущных сельскохозяйственных наук и ветеринарии ранние ученые увлекались мифологической географией и магией. Так случилось потому, что наука складывалась стихийно, в духе накопления информации, пригодной для насыщения свободного времени, а этот процесс шел, как мы показали, неразборчиво.
Достижения первобытного общества были куда скромнее. Здесь накапливались сведения об окружающей действительности, однако они принципиально не отличались от информации, собираемой животными. Зато в сфере праздных разговоров наступили выдающиеся отличия. Систематические собеседования на обыденные темы породили предания, нарождающаяся религия оснастила их фантастическим флером, дав сказки, а становящееся нравственное сознание, оживив моралью, инициировало притчи. Тем самым были заложены основные составляющие художественной литературы: социальность, нравственность и фантазия. Впечатления от выдающихся переходов, охот и территориальных конфликтов сформировали эпос. Первоначально литература была устной, ориентировалась на слушателей, а потому наглядно служила целям социализации.
Религия
Становящаяся религия тоже имела биологические корни. В дождливую погоду, обещающую изобилие природы, шимпанзе приходят в возбуждение, приплясывают, размахивают ветвями, исполняют «танец дождя» [59, с. 48–49; 800]. Радостные встречи стад этих приматов в условиях изобилия среды (так называемые «карнавалы») сопровождаются картинными выражениями расположения человеческого типа с рукопожатиями, объятиями, похлопываниями по спине, поцелуями, уважительными касаниями рукой головы, «барабанной дробью» и другими человеческими элементами «протокола», свойственными, скажем, встречам государственных лидеров на высшем уровне [12, с. 117–118; 59, с. 72].
Первое мероприятие типологически напоминает исходный вариант для универсально распространенного в примитивных человеческих сообществах ритуала вызывания дождя [98, с. 28–29]. Второй проторитуал подобен обряду плодородия типа австралийского церимониала Кунапипи [11, с. 214–219]. Ввиду родства людей и шимпанзе можно допустить, что у ранних гоминин существовали свои «танцы дождя» и «карнавалы», послужившие основой их ритуального магического поведения, что подтверждает первичность магии среди человеческих верований [109, с. 39, 53–64].
Магия
– это ритуальные операции с особыми предметами в сопровождении музыки, танцев, песнопений и заклинаний, – операции, нацеленные на то, чтобы принудить потусторонний мир направить посюсторонние события в желательную сторону. Магические обряды коллективны. Феномен одиночных, подпольных колдунов – поздний. Он обусловлен враждебным религиозным окружением, как было с магами в средневековой христианской Европе. Подчеркнем, что приведенное определение магии отражает положение дел у мезолитических аборигенов Австралии и бушменов Южной Африки. Палеолитические предки человечества, вероятно, обходились магическими обрядами меньшей сложности.
С точки зрения нашей концепции происхождения духовной культуры первые ритуалы обслуживали свободное время гоминин с целью их социализации, поскольку являлись праздными коллективными занятиями. С течением времени они усложнялись и обогащались, подчиняясь определенной логике, ибо рост свободного времени у членов социума требовал все новых наполнителей.
С философской точки зрения орудия гоминин представляли собой предметы активного определенного класса (поскольку орудия составляли определенный набор и использовались активно). Имитация их применения в свободное время давала предметы активного неопределенного класса, в которых узнаются инструменты магии, могущие быть чем угодно (орудиями труда и охоты, минералами, животными и растительными материалами, жидкостями и другими предметами, призванными мнимо воздействовать на действительность и неограниченными в перечне, откуда, собственно, происходит квалификация их класса) [11, с. 224–254]. Присущие обрядам вызывания дождя неутилитарные кристаллы кварцы известны на стоянке синантропов в Нижней пещере Чжоукоудянь, Китай, 500–230 тыс. лет назад [214, с. 288].
Имитация предметов первобытного труда (промысловых животных) давала предметы определенного пассивного класса (это ограниченный круг предметов: ими не действуют – к ним обращаются с просьбой, мольбой). В них можно видеть кремневые статуэтки анималистической, животной мифологии, датируемые Рисс/Вюрмом позднего ашеля Богутлу в Армении, 144 тыс. лет назад. Поскольку эта изобразительная традиция неразрывно связана со знаковым творчеством, анималистическую мифологию можно датировать временем ок. 783 тыс. лет назад (Странска скала, Чехия, где обнаружен позвонок ископаемого слона с 7 знаковыми зарубками; см. приложение 1). В отличие от магии с ее наступательным началом [98, с. 37; 109, с. 39, 53–64], мифологию отличает пассивное, просительное отношение к предметам культа, что характерно для религии.
Мифология
– это исторические предания о кульминационных моментах в судьбе сообщества, как они видятся спустя века, в отсутствие точной информации и глазами людей, наделенных воображением. Первая мифология была космогонической и трактовала вопросы происхождения Вселенной, ограниченной в палеолите Землей и Небом (см. приложение 1). Мифологические предметы (скульптурные иллюстрации мифов в палеолите, их рисованные иллюстрации в среднем – верхнем палеолите, иконография наших дней) служили и служат объектами почитания, т. е. просительного, пассивного отношения.
Расширение мифологических инструментов до предметов неопределенного пассивного класса давало тотемные эмблемы, поскольку тотемами могли быть любые явления, хотя животные играли среди них видную роль (но использовались и растения, явления природы, солнце, луна и другие предметы неограниченного списка и пассивного, не действенного назначения, что задает их класс) [11, с. 162, 164, 170]. Древнейшая тотемная эмблема, выстроенные в линию кости слона, известна в Амброне, Испания, из древнего среднего ашеля Минделя II, 411–388 тыс. лет назад. Тотемизм
– это убеждение в родстве людей с объектами внешнего мира (животными, растениями, явлениями природы и др.), символизируемыми тотемными эмблемами, т. е. условными изображениями. Тотемная идея родилась в среде охотников, чьи профессиональные рассказы размывали грань между охотником и жертвой.
Особые тотемы противопоставляли первобытные сообщества друг другу. Это говорит о том, что тотемизм возник в эпоху конфликтов первобытных популяций, т. е. в период демографического взрыва у «человека-мастера». Его внучатый отпрыск, гейдельбергский человек, судя по находке в Амброне, уже располагал началами тотемизма, однако ни он, ни его предшественник, классический питекантроп (Homo erectus), не переживали демографических взрывов. Следовательно, тотемизм действительно освоил их предок, «человек-мастер», переживший демографический всплеск, хотя археологических свидетельств в пользу подобной точки зрения пока нет.
Пассивизация (выведение из активного применения) инструментов магии приводила к предметам аналогичного неопределенного пассивного класса, отвечающим фетишам (тоже способным быть чем угодно: нательными украшениями, орудиями и оружием, источниками и деревьями и другими объектами пассивного почитания [80, с. 210–211]), древнейшие из которых (полированные фрагменты кварцита) известны в древнем тейяке из интер-Рисса Бечова, Чехия, 281–219 тыс. лет назад. Фетишизм
– это вера в сверхъестественные силы разнообразных неодушевленных предметов, подлежащих почитанию, первоначально коллективному. Таково, например, почитание различных священных мест и примечательных элементов среды (родников, утесов и т. п.).
Логика развития предметных форм древних верований отвечала пассивизации и расширению их классов. Расширение следовало увеличению свободного времени, а пассивизация – непроизводственному назначению. Предельное развитие этой логики привело к появлению объектов верований, отнесенных к неопределенному пассивному классу беспредметного характера, т. е. существующему лишь на словах. Их беспредметность отвечает максимальному отрыву от мира практической жизни. В этих необычных объектах веры угадываются духовные существа анимизма (души живых существ и духи неодушевленных предметов) [95, с. 356–357]. Анимизм
– это убеждение в существовании нетленных душ живых существ и могущественных духов неживых предметов; бытующие в реальности лишь на словах, они являются объектами пассивного почитания. Вопреки мнению об их первичности по сравнению с фетишами [95, с. 337], происхождение этих абстрактных объектов культа видится нам следующим образом.
Духовным существам анимизма приписывается потустороннее бытие. Между тем, единственная реальная предметная форма потусторонней действительности соотносится с «тем светом» загробного мира, связанного с погребениями
. Проще говоря, никакого реального «того света», кроме могил, человечество не знает. Помещая в могилы вместе с покойниками различные приношения, в том числе фетиши, и обсуждая их дальнейшую судьбу, первобытные люди фактически наделяли их мнимой посмертной жизнью (на словах), отчего покойники с течением времени приобретали ранг «душ», а погребенные с ними предметы ранг «духов». В силу экспансии (расширения) объектов культа под влиянием роста свободного времени души и духи постепенно нашли воплощение и в предметах «этого света», став духами-хранителями очага, духами источников, деревьев и многого другого, выступив предтечами душ и духов классических религий и философского гилозоизма (учения о сплошной одушевленности природы). Они же послужили архетипом научного образа сущностей в философии и науке, а также философских представлений о трансцендентном бытии. Иными словами, религиозные и философские представления о потустороннем или трансцендентном мире имеют весьма древнее и вполне определенное происхождение. Судя по приведенным датированным примерам, их «изобрел» еще гейдельбергский человек.
Орангутаны и гориллы порой погребают своих мертвых, присыпая трупы листвой или землей [114, с. 167, 219], так что разумно допускать, что погребальная практика изначально присуща гомининам. Реальные же погребения зафиксированы археологически применительно к довольно позднему времени. Судя по следам преднамеренной посмертной практики на черепах Homo sapiens idaltu [248, с. 748], их подвергали погребальному обряду 160 ± 2–154 ± 7 тыс. лет назад. Погребения отмечены и у про-токроманьонцев Кафзеха 115 ± 15–92 ± 5 тыс. лет назад. Тогда же был погребен неандерталец в гроте Киик-Коба, Крым, Украина, 111 тыс. лет назад; cp. [290]. Поскольку неандертальцы были способны к членораздельной речи [150]; cp. [540], анимизм мыслится и для них.
Характер погребений задал нашим предкам локализацию потустороннего мира. При захоронении душа покойника вместе с телом убывала под землю. Отсюда выросли представления о ненасытном подземном мире. При кремации душа покойника вместе с дымом погребального костра отправлялась на небо. Это эмпирическое обстоятельство породило образ потусторонних небес, населенных душами различных небожителей. Представления о подземном и небесном мирах широко распространены по свету, а потому должны быть весьма древними, как и первые погребения.
У современных и первобытных людей религиозные и магические обряды, как правило, приурочены к определенным календарным датам. Лунный календарь [505; 535] и арифметический счет [108; 505] были известны уже у кроманьонцев. Связанные с памятниками анималистического изобразительного искусства календарные отметки восходят к временам Homo heidelbergensis и датируются в Странска скала, Чехия, приблизительно 783 тыс. лет назад. Их можно рассматривать и как свидетельства интеллектуальных достижений гоминин.
Относительно времени возникновения первых религиозных верований ясности пока нет. В свете приведенных фактов можно думать, что тотемизм и фетишизм имелись уже у гейдельбергского человека, а для них характерно просительное отношение к объекту верования, т. е. элементарная форма религиозности. Определенные указания на время зарождения религии дают социально-психологические соображения.
Как отмечалось выше (см. разд. 2.4), в истории наших предков имело место постоянное чередование акселерированных и неотеничных разновидностей. Акселерированным формам была присуща психологическая незрелость, инфантильность и порывистость. С таким психотипом гармонировали наступательные, требовательные верования, т. е. магия. Отсюда вытекает вероятность, что, например, акселерированный «человек-мастер» был склонен к магическим убеждениям.
Напротив, неотеничным формам гоминин была присуща психологическая зрелость, взрослость, склонявшая их к взвешенным, рассудительным верованиям, т. е. к просительной религиозности (мифотворчеству, тотемизму, фетишизму). Напрашивается предположение, что начала религиозности зародились у неотеничных Homo erectus и Homo heidelbergensis. Разумеется, приведенные умозаключения дают нам лишь общий взгляд на проблему, и решающее слово остается за археологическими фактами, которых пока не так уж много.
Отталкиваясь от наблюдений за чередованием неотеничных и акселерированных разновидностей у наших предков, мы можем сделать предположения относительно возникновения обрядов инициаций у древних гоминин. Инициации
– это ритуальные испытания, которым в первобытных племенах подвергалась молодежь с целью выяснения степени ее физической и нравственной зрелости. Юношам назначались трудные, порой мучительные и опасные испытания (от выбивания зубов с условием, чтобы не выступало слез от боли, до задания в одиночку добыть, например, льва, что до сих пор практикуется у африканского племени масаев из Кении и Танзании в Восточной Африке). Инициации у девушек выглядели скромнее и связывались с обрядами наступления половой зрелости.
Ошибочно думать, будто инициации канули в Лету вместе с первобытным обществом. Напротив, они процветали и процветают в мире цивилизованных людей. В частности, мы имеем в виду систему экзаменов, которыми пронизана средняя и высшая школа, а также правила присуждения степеней бакалавров, магистров, кандидатов и докторов наук. Профессиональные разряды на производстве и на транспорте присваиваются также через прохождение экзамена. Что и говорить – без него не получить элементарных водительских прав. В Средневековье без определенного испытания нельзя было получить звания рыцаря (в Западной Европе) или самурая (в Японии). Все эти формы аттестации молодых людей ведут начало от первобытных инициаций.
Встает вопрос: в чем причина их возникновения? Нельзя поверить, что первобытные гоминины целенаправленно придумали экзаменационные сессии для своей молодежи. Инициации, скорее, возникли следующим образом.
Когда наши неотеничные первобытные предки на стадии «человека-мастера» (или раньше) вступили в фазу акселерации, умудренные жизнью неотеники обнаружили себя в окружении высокорослой инфантильной молодежи (мы рассуждаем здесь несколько фигурально, но картина могла напоминать современную, когда не слишком рослое старшее поколение людей сталкивается с легкомысленным рослым юношеством). Нетрудно представить, что психологически незрелая, акселерированная молодежь неуверенно вписывалась в жесткую структуру традиционной первобытной жизни с ее количественно замороженной технологией и отвечающим ей регламентированным досугом (см. разд. 3.1, 3.2).
Как представляется, неотеничное старшее поколение стихийно принялось подгонять хронически незрелую молодежь к своим нормам жизни и тиранически экзаменовать юношество в преддверии большой жизни на предмет усвоения неотеничных принципов жизни – проще говоря, технологических и культурных достижений неотеничных гоминин. Нам думается, что неотеники пользовались обычными для высших млекопитающих методами, воспитывая неотеничную же детвору, склонную к благополучному психологическому созреванию. Однако, получив в своих популяциях неуправляемую акселерированную поросль, готовую к вызывающему поведению, несчастные неотеничные родители были вынуждены прибегнуть к мерам, экстраординарным для приматов.
Высшие млекопитающие готовят своих отпрысков ко взрослой жизни, однако экзаменов им не устраивают. Неотеничные гоминины с появлением акселерированной молодежи развили эту педагогическую практику путем моделирования физических и нравственных трудностей, что вылилось в инициации. В эпоху первобытности они не отличались продуманной последовательностью, свойственной нынешним экзаменационным испытаниям. Это выдает стихийное, непреднамеренное зарождение инициаций.
Происхождение системы табу, свойственной первобытному обществу, вероятно, объясняется в том же ключе. Табу
– это немотивированные запреты, возбраняющие первобытному человеку определенные поступки. Табу бывают производственными, охотничьими, пищевыми и половыми (т. е. могут захватывать основные сферы человеческой жизнедеятельности), но могут быть и второстепенными. Производственные табу – это запреты на нетрадиционные технологии, грозящие расширить технологический инвентарь сообщества вопреки требованиям демографо-технологической зависимости (см. разд. 3.1). Подобные запреты подспудно действуют по сей день и называются производственным консерватизмом. Это, конечно, не табу, но внешне схожее с ними явление.
Охотничьи табу запрещают преследование почитаемых животных, в частности тотемных. Они запрещают и нетрадиционные способы охоты, в чем совпадают с производственными табу. Пищевые табу, вероятно, являются продолжением охотничьих запретов, поскольку, ограничивая выбор пищи, они ограничивают и выбор охотничьей добычи (например, аборигенам Аравии – иудеям и мусульманам – нельзя есть свинину; соответственно, нет смысла охотиться на кабанов; запрещение же охоты на кабанов, очевидно, проистекло из неприятия специальных методов их добычи, а те, в свою очередь, не допускались, чтобы не усложнять технологию, которая в условиях скудной Аравийской пустыни должна была отвечать немногочисленному населению, т. е быть по возможности простой). Половые табу возбраняют повышенную сексуальную активность как количественно, так и качественно (например, отвергая инцест – близкородственное скрещивание).
Нетрудно видеть, что в основе табу лежит стремление предотвратить любое нетрадиционное отношение к устоявшемуся образу жизни. Угроза неправильного поведения здесь появилась с приходом акселерированных поколений, легкомысленно готовых покуситься на традиции, обеспечивавшие единство первобытного социума, который не допускал наступления на свою целостность. Выжили те сообщества древних гоминин, которые сумели опутать акселератов системой табу и инициаций, препятствующих экспериментам с однообразием общественного образа жизни. Со временем табу зажили собственной судьбой, расширились в масштабе от суеверий до религиозных и нравственных запретов и перестали распознаваться как меры обуздания незрелой молодежи. Разумеется, первобытные гоминины тоже не понимали их адекватно. По мере роста благосостояния общества табу расширялись в интересах упорядочения досужего времени.
Нравственность
В нравственной сфере завоевания наших предков являются еще более древними, поскольку имеют более внушительные биологические предпосылки. У высших животных поведение распадается на эгоистическую и альтруистическую составляющие. Эгоистическое поведение шлифуется под влиянием индивидуального естественного отбора, а альтруистическое своим происхождением обязано групповому [39, с. 56–57, 339; 73, с. 235, 330; 123; 379; 707], который побуждает организмы, располагающие общим генотипом, самоотверженно помогать друг другу, так что кажется, будто они заряжены неким геном альтруизма (см. разд. 3.1).
Подчиняясь альтруистическому началу, возникшему вследствие группового отбора сородичей, позвоночные и беспозвоночные животные способны ради своей родни на большие жертвы независимо от уровня развития нервной системы и характера разума (индивидуальный у позвоночных, коллективный у общественных насекомых). Они готовы воздерживаться от размножения (термиты, муравьи, пчелы, осы, шмели;
некоторые птенцовые птицы; млекопитающие вроде сурикатов или гиен), воспитывать чужое, но родственное потомство, защищать его и родню, не щадя жизни, и, разумеется, оказывать различные услуги.
То обстоятельство, что общественные насекомые контролируются при этом коллективным разумом, а позвоночные животные – индивидуальным, не должно настораживать в смысле сравнимости примеров. Характер разума здесь диктуется не высшими психическими факторами, а довольно прямолинейными законами экологии количественного свойства. Согласно закону Линча – Конери [523], размеры организмов и их геномов обратно пропорциональны численности популяций соответствующих видов. Эта зависимость обусловлена довольно простым обстоятельством.
По экологическим причинам любой биотоп способен вместить лишь ограниченную биомассу живых существ. Если они мелки, как, например, микроорганизмы, то разрешенная общая биомасса позволяет разместиться в биоме огромному числу организмов. Многоклеточные существа крупнее, и их особей в биоме меньше. На всякую живую особь приходится определенный процент объема биома, к которому необходимо приспосабливаться на генетическом уровне. У микроорганизмов жизненное пространство невелико, поэтому мал и отражающий его геном. У многоклеточных существ жизненное пространство куда обширнее, поэтому отражающий его геном крупнее.
Перенося эту зависимость в сферу нервной деятельности, надо учесть, что организмы отражают жизненное пространство не только на генетическом, но и на фенотипическом уровне, т. е. психически. Поэтому нервная система мелких существ (например насекомых) проста, а у крупных (например у позвоночных) она сложнее. Когда коллективные насекомые оккупируют несвойственное отдельным особям обширное пространство, потребность адекватно отражать его понуждает этих небольших существ объединяться на химической основе феромонов. Эти летучие вещества, перенося информацию запахом и вкусом, связывают нервные системы отдельных особей в единую сеть, что помогает сообществам муравьев, пчел и других общественных насекомых уверенно справляться с большим для себя миром, захваченным жизненным пространством. Как видим, природа их коллективного разума, в принципе, подобна психике позвоночных животных. Его устройство лишь компенсирует незначительность физических размеров насекомых.
Подобно другим высшим приматам, гоминины были склонны как к эгоистическим, так и к альтруистическим нравам. Эгоистическая модель замкнута на особи и не разнообразит ее общение. Напротив, альтруистическая модель ориентирована на окружающих, а потому подпитывает отношения с ними.
Когда производительность труда у гоминин пошла вверх, им потребовались все варианты непроизводственного общения, способные укомплектовать свободное время. Альтруистическое поведение тут же вошло в оборот и стало раздуваться пропорционально объему праздного времени. Оно было построено на самоотверженности одних индивидов по отношению к другим. Потребность гоминин все шире и шире применять эту способность (по мере подъема производительности труда и развития свободного времени) привела к дифференциации проявлений самоотверженности. Как отмечалось, ее элементарная, животная форма сводится к поддержанию и защите жизни и здоровья патронируемой родни, хотя порой дело доходит до содействия неродственным особям и даже представителям других видов, что отмечено у дельфинов, бегемотов, волков и др. Праздных гоминин такая узость устраивала все меньше. Первым делом они принялись жертвовать в пользу собратьев не только наличными, но и будущими интересами, существующими лишь «в проекте». Это привело к запрету на ложь в отношениях с другими индивидами.
В любом сообществе особи физически и умственно не равноценны. Животные нравы, подпитанные естественным отбором, требуют предпочтительного положения для выдающихся экземпляров, что ведет к иерархической системе доминирования, обычной у коллективных животных (это – система всеобщего неравноправия, когда слабые особи ступень за ступенью подчиняются сильным). Раздувая свою самоотверженность, гоминины пришли к известному равноправию индивидов в сообществе независимо от своих природных задатков. Наконец, противоестественно раздутая в сообществах гоминин самоотверженность стала в известном смысле подавлять здоровый природный эгоизм наших предков, и у них возникла невиданная в животном мире тяга к самооценке вклада каждого в общую самоотверженность.
В итоге на почве экспансии альтруистического поведения у гоминин сложилась система норм, включающая самоотверженность, честность, равноправие и совесть (самооценку вклада личности в самоотверженность общества). Обсуждаемая в беседах, она стала основой нравственного сознания. Моральное поведение не являлось универсальной моделью. Оно сосуществовало и сосуществует с эгоистическими стереотипами, поскольку его задача состояла не в усовершенствовании наших предков, а лишь в заполнении их свободного времени социализирующими занятиями.
Проявления альтруизма у ископаемых гоминин ограничены характером материала. Считается, что больные или покалеченные, но выжившие особи своим спасением обязаны альтруизму соплеменников. Примеры включают самку Homo ergaster KNM-ER 1808 из Кооби Фора в Кении св. 1,5 млн лет назад [18, с. 199]; классического питекантропа I, первого Homo erectus из Триниля на Яве, Индонезия, 710 тыс. лет назад; классического Homo heidelbergensis из Мауэра, Германия, ок. 700 тыс. лет назад [273]; позднего гейдельбержца из Мугарет-эль-Зуттиех в Израиле, 148 тыс. лет назад; прогрессивного неандертальца из Шале в Словакии, св. 110 тыс. лет назад; классического неандертальца Шанидар I из пещеры Шанидар в Курдистане, Ирак, 46 900 ± 1500 лет назад.
Как можно убедиться, первые наличные данные о проявлении зачатков нравственности у наших предков относятся ко времени ашельской технологической революции у «человека-мастера». Тогда производительность первобытного ашельского труда возросла против развитого олдовая. Однако мы не удивились бы, если бы признаки вторичных общественных структур, отвечающих духовной культуре, обнаружились бы у Kenyanthropus rudolfensis, поскольку его типичный олдовай бытовал в течение почти миллиона лет и вполне мог поднять мастерство «кенийцев» до высот, не предусмотренных одной только каменной фактурой орудий. Это могло бы увеличить производительность труда.
Расширяясь, этическая сфера породила еще две области духовной жизни: поведенческий этикет и правовые нормы. Этикет – это общепринятые правила поведения, регламентирующие обыденную (а теперь и официальную) жизнь стандартным набором приемов принятия пищи, отправления гигиенических нужд, встреч и времяпрепровождения с себе подобными. Не имея интеллектуальной, религиозной, нравственной или эстетической нагрузки, этикет выдерживается подобно этическим нормам, а оценивается скорее эстетически в понятиях приличного (красивого) и неприличного (некрасивого) поведения.
Архетипы, образцы приличного поведения наших предков уходят в обезьяньи времена, судя по повадкам родственных нам шимпанзе и горилл. Так, шимпанзе умеют пользоваться листьями как салфетками (столовыми и в качестве туалетной бумаги), заедают мясную пищу (например, кусочки мозга) листьями (подобно тому, как мы едим жаркое с гарниром), приветствуют друг друга наклоном головы (мы тоже при этом кланяемся) или даже припадают телом к земле (как делают люди перед лицом Бога или монарха), приветственно протягивают руку для рукопожатия, обнимаются, целуются (чмокаются, как женщины или политические лидеры при встречах), уважительно касаются головы рукой (мы снимаем шляпу или козыряем, если носим форму), самцы ободряюще треплют самок и детенышей по подбородку (что делают и радушные мужчины) [59, с. 148, 177, 202].
От шимпанзе не отстают гориллы: у них раздраженные доминирующие, господствующие самцы с серебристой гривой (род седины) демонстративно прохаживаются на глазах у стада, скрестив руки на груди в наполеоновской позе (подобно тому, как наши маститые, седовласые начальники рассерженно расхаживают, скрестив руки на груди, на глазах у подчиненных). Детеныши у хвостатых приматов держатся за хвост матери, у бесхвостых – за шерсть (дети у людей держатся за юбку матери); все приматы любят носить детенышей на спине (люди сажают их на шею, откуда наше выражение «сидеть на шее у кого-либо»). Гориллы склонны украшать себя пучком листьев на голове в виде султана (откуда происходят плюмажи индейцев, эгейских, этрусских и римских воинов, а также человеческие головные уборы всех времен). Выражая подчинение, обезьяны подставляют шею для укуса (люди, соответственно, отвешивают земной поклон) [114, с. 166, 168, 178, 180, 182].
Когда у гоминин начался рост производительности труда и свободного времени, они не преминули увлечься обезьяньими началами этикета и возвести их в непременные нормы жизни, чтобы заполнить свободное время еще одной формой праздного общения: правилами приличия. Ближе к концу первобытности (в эпоху «варварства», сменившего эпоху «дикости» и предшествовавшего эпохе «цивилизации») нравственные нормы, расширяясь сообразно укреплению производительности труда и прибавлению досуга, отпочковали от себя такую область духовной культуры, как обычное право, что произошло не раньше неолита, поскольку у автралийских аборигенов и южноафриканских бушменов, пребывающих в условиях мезолита, обычного судебного права еще нет.
Уже в эпоху цивилизации обычное право было письменно кодифицировано, дав первые образцы юрисдикции. Наиболее ранние образцы законодательного права происходят из Шумера (территория нынешнего южного Ирака). Нам известны изложения уже модернизированных (т. е. не самых ранних) законов Энметены (2360–2340 до н. э.) и Уруинимгины (2318–2312 до н. э.), которые являлись, соответственно, пятым и девятым царями I династии города Лагаша. Известны также своды законов Ур-Намму (2112–2094/2093 до н. э.) и Шульгира (2093–2046 до н. э.), которые, соответственно, были первым и вторым царями III династии города Ура. Эти документы считаются древнейшими памятниками писаного права [45, с. 208–210, 274–275]. Примечательно, что судебник Ур-Намму местами выходит за рамки обычных шумерских норм, а это указывает на то, что шумерские законники уже обладали профессиональным воображением и правовым самосознанием (в приличествующих эпохе скромных рамках).
Нормы обычного и кодифицированного права типологически напоминают нравственные установки. Те и другие регламентируют жизнь личности в интересах окружающих, одновременно оберегая ее от последних. Поэтому генетическая связь нормативов обоих типов не вызывает особых сомнений. Однако методы их утверждения в жизни различны: мораль основана на совести («нравственном законе», по И. Канту), право же опирается на насилие. Содержательные расхождения права и морали не велики: пожалуй, нет ни одного преступления, которое не порицалось бы и нравственно, что еще больше роднит обсуждаемые сферы общественного сознания. Их обособление друг от друга объясняется не самобытностью, а нуждой социума в новых социальных связях.
Когда жизнь цивилизованного общества обогатилась взаимоотношениями государств, строящимися (в идеале) по лекалу нравственного и правового сознания, социум обзавелся политическим сознанием, особенностью которого являются методы примирения моральных и правовых принципов с беспринципностью реальной корыстной жизни. Несмотря на возмущение политикой со стороны лучших (хотя и недалеких) умов человечества, политика от века к веку только крепла, поскольку представляла собой добавочную ветвь общественного сознания, которая пришлась по душе социуму, ибо тот неизменно поощрял любые формы украшения досуга общением непроизводственного свойства, в чем политическое сознание преуспевает не хуже прочих областей духовности.
Выше мы имели в виду внешнюю политику. Однако внутренняя политика обладает аналогичными социализирующими качествами. Отличие ее от внешней политики состоит лишь в том, что внутренняя политика ведет начало от изощренных интриг в стадах обезьян (например у шимпанзе), где мы находим все особенности «искусства компромисса» [53, с. 117–124]. Так, обезьяны умеют снискать благосклонность сильного лидирующего самца и науськать его на неугодных собратьев, а затем жить и скрытно царствовать у него «под крылом». При этом многоступенчатая, далеко идущая интрига здесь не редкость. Не редкость и коварное объединение слабых особей против сильных с целью произвести революцию. Заговор реализуется либо прямым силовым нажимом (модель путча), либо путем хитроумного внесения разногласий в среду мощных, но бесхитростных вожаков (модель «бархатной революции»). Перечень примеров можно продолжить.
Согласно изложенной гипотезе, нравственное поведение и моральное сознание зародились не для того, чтобы «облагородить» наших предков, а скорее для того, чтобы занять полезным праздным общением досужее время, образовавшееся у гоминин после ашельской технологической революции. Избранный объективистский («незаинтересованный») подход к проблеме генезиса нравственности позволяет объяснить, почему она возникла только у «человека-мастера» и не сложилась ни у одного другого живого существа, хотя, казалось бы, заслуживала большего распространения среди высших млекопитающих.
Искусство
Единственная форма духовной культуры, не имевшая у гоминин биологических предпосылок – искусство. Положим, у наших предков имелись некоторые зачатки эстетических чувств, а некоторые проявления художественного творчества опирались на биологическую подоплеку. Совсем как у людей, самки и подростки горилл тщательно заботятся о внешности и любят украшать себя пучками листьев на голове или кусками мха на загривке [114, с. 178, 180], испытывая, надо полагать, те же эстетические чувства, что и женщины, украшающие себя плюмажами. Те и другие кокетничают, гордясь своей красотой.
Показано [71], что величина пятен на шкуре африканских хищников семейства кошачьих положительно зависит от массы животных, превращаясь у взрослых львов в сплошной фон. Если первые гоминины-охотники маскировались на охоте какой-нибудь раскраской, то при массе 55 кг (Kenyanthropus rudolfensis) они разрисовывались бы мелкими пятнами, как у гепарда, а при массе 70 кг (Homo ergaster) – пятнами покрупнее, как у леопарда (ср. [103, с. 160–161, табл. 6.1; 231, с. 851, табл. 1], где приняты устаревшие рамки видов). Из этой практики мог возникнуть обычай покрывать тело красителями, татуировкой и нательными украшениями вроде наших драгоценностей. Красящий гематит известен на стоянке позднего африканского питекантропа BK II в Олдувайском ущелье, Танзания, ок. 1,2 млн лет назад.
Высокое искусство у гоминин имело сакральное происхождение. Производя каменные отщепы, наши предки подметили в некоторых из них сходство с силуэтами мифологических животных [53, с. 21–30, 211–221]. Повторяя это подобие методом воспроизводства технологических образцов, древние мастера «запустили в тираж» кремневую пластику, эстетические качества которой были предельно низкими. Лишь пройдя в верхнем палеолите художественные стили I – IV (см. приложение 1, рис. 11–18), доисторические художники достигли изящного «фотографического реализма». Судя по их настойчивости, в течение длительной эпохи аудитория одинаково восхищалась как исходным примитивизмом, так и финальным «фотографическим реализмом». Отсюда следует, что первоисточником эстетических чувств были не художественные образы, а их сакральный смысл.
Статистический анализ франко-кантабрийского искусства показывает, что оно являлось мифологическим, причем в центре соответствующей мифологии находилась триада Коня-Солнца, Быка-Земли и Оленя-Неба, и это объясняет сакральный смысл одного из погребений в пещере Кафзех, где подростка сопровождала на «тот свет» часть черепа лани с рогами (покойнику покровительствовала Лань-Небо, предположительно отвечающая за доступ в потусторонний мир, см. приложение 1).
Охотничья по происхождению, первобытная мифология была космогонической. Поэтому можно представить, насколько богатые чувства охватывали ее носителей при виде уродливых, схематических статуэток мифологических животных. Отпечатавшись в психике, эти религиозные эмоции обрели самостоятельную жизнь и дошли до нас в виде священного чувства прекрасного, метафорическая характеристика которого на деле содержит глубокий забытый смысл.
Рассчитанное на зрителей первобытное искусство, как и современное, служило мощным поводом для общения в свободное время. С точки зрения прагматических нужд социума оно имело несомненное положительное значение для сохранения общества в целостности.
Сказанное справедливо и для искусств других жанров. В упомянутых выше «карнавалах» шимпанзе радостно приплясывают, вскрикивают и производят «барабанную дробь» по корням деревьев, т. е. занимаются в зачаточном виде танцами, вокалом и музицированием. Трудно сомневаться, что у гоминин эти формы творчества пошли из аналогичного источника.
Приплясывающие «колдуны» изображены на стенах французских гротов де Лабастид, 14 260 ± 440 лет назад, и де Труа-Фрер-2, 13 900 ± 120 лет назад, итальянских гротта ди Леванцо, 11 800–10 800 лет назад, и гротта делл’Аддаура I, 10 800–10 200 лет назад, на Сицилии. Показано, что в украшенных пещерах верхнего палеолита имелась хорошая акустика – там пели и играли на барабанах, флейтах и свистульках [672]. Костяные «ударные инструменты» обнаружены в Межириче на Украине, 15 245 ± 1080 лет назад. В Труа-Фрер-2 есть изображение музыкального лука; сюда же добавим «охотника с луком под мышкой» из Ле Пеш-Мерль-2, 16 500–15 500 лет назад, который, возможно, не охотился, а музицировал. Свистульки из фаланг животных открыты в Бокштайншмиде, Германия, 144–110 тыс. лет назад, и в других местонахождениях. Иными словами, ударные, щипковые и духовые музыкальные инструменты возникли еще в среднем – верхнем палеолите.
От мероприятий типа «карнавалов» шимпанзе через ряд опосредований происходит театральное искусство. «Карнавалы» легли в основу сперва магических, а потом религиозных ритуалов. Последние в эпоху Архаического Ура в Шумере, Ирак, 2900–2615 до н. э., уже стали основой мистерий «священного брака», которые проводились в сопровождении арфисток и певчих [45, с. 178].
В Древней Греции у истоков классического театра оказались дионисийские мистерии и хоровая песнь дифирамб, в жанре которой творил хоровой лирик Арион ок. 600 до н. э. Представление усложнил Феспид, дополнивший в 534 до н. э. хор актером-декламатором, комментирующим зрелище. Введение второго актера (Эсхил, 525–456 до н. э.) и третьего (Софокл, 496–406 до н. э.) создало драматическое представление, не зависимое от хора. Современные ценители видят в театре «храм искусств» и переживают там чувства, близкие к священным. Исторически это обусловлено тем, что изначально прототеатральные ритуалы действительно заряжали участников сакральными эмоциями, которые унаследовались мистериями, а впоследствии – театром. Скрестившись с фотографией, театр породил кинематограф и обзавелся массовой аудиторией, которая боготворит кино в соответствии с первобытной моделью сакральных чувств, хотя содержание массовой кинопродукции никах не назовешь возвышенным.
Вкратце изложенная версия происхождения духовной культуры таит в себе гораздо бóльшие объяснительные возможности. Возьмем, например, физическую культуру. Иным критически настроенным людям исключительная популярность спорта кажется не во всем оправданной, поскольку занятия физической культурой по определению не преследуют целей рафинирования интеллекта или утончения эстетических пристрастий. Между тем скептическое отношение к спорту недальновидно, и дело здесь не только в ценности здорового образа жизни.
С социально-философской точки зрения физкультура и спорт являются средствами заполнения свободного времени социализирующим, но непроизводственным путем, и в этом состоит их ценность для целостности и выживания социума. В самом деле, что такое спортивные состязания? Это прежде всего огромные скопления людей, общающихся по поводу спортивного зрелища. Ситуация полностью отвечает положению дел, например, в театре (подчеркнем, что речь идет о сугубо социально-философском видении темы). Страстного театрала такое отождествление озадачит. Однако для слепого социума решительно все равно, чем связывать людей в досужее время: философическим «Гамлетом» или грубым боксом. Главное, чтобы без общения у людей не протекало ни минуты свободного времени (в идеале).
Страшно сказать, но общие празднества и даже коллективные злоупотребления алкоголем или наркотиками (что можно только осуждать) подчиняются той же модели социализации свободного времени непроизводственным путем. Причина того, что социум, поощряя здоровый образ жизни (например коллективный спорт), одновременно мирволит некоторым разрушительным увлечениям людей, если они коллективны, состоит, конечно, не в его циничности, а в том, что он слеп и неантропоморфен, т. е. не обладает человеческими представлениями о добре и зле.
Его интересуют лишь собственная целостность и физическое выживание, и, как мы видим, первое подчас входит в противоречие со вторым, поскольку нездоровые коллективные увлечения людей не способствуют физическому здоровью социума. Однако, прежде чем лечить незрячий социум, следует адекватно оценить его природу, чему и посвящена данная глава.
Неубывающий технологический прогресс и пропорциональный рост досужего времени привели духовную культуру к невиданному размаху. В этой связи умение создавать и поддерживать вторичные социальные связи приобрело в глазах гоминин жизненно важное значение (поясним, что к первичным социальным связям относятся отношения в сфере материальной жизни, прежде всего производственной, а к вторичным – культурные отношения в духовной жизни).
Умение создавать и поддерживать культурные связи зависит от творческих и речевых способностей людей. Половозрастное разделение труда возложило соответствующие обязанности в основном на зрелых мужчин. Эти обязанности важны для выживания общества. Поэтому осуществляющие их мужчины имеют привлекательный социальный статус. Вот почему женщины «любят ушами». Им мало, чтобы мужчина был здоров, крепок, неотенически красив (см. разд. 2.4) и пригоден к общественному лидерству. Необходимо, чтобы он был речист и, по возможности, способен создавать духовные ценности (интеллектуальные, религиозные, моральные, художественные, спортивные). Вот почему женщины, грубо говоря, любят болтунов. Здесь прекрасный пол как раз и проявляет свою прозорливость.
Напротив, на хрупкие плечи слабого пола в силу естественных причин возложены биологические обязанности по продолжению рода. В этом случае соответствие нормам неотеничной эволюции и физическая неотеничная красота выступают на первый план. Поэтому мужчины «любят глазами». Ни в женских, ни в мужских пристрастиях нет никакой злокозненной дискриминации. Эти пристрастия мудро сложились в ходе естественно-исторического развития социума. Поэтому идеология феминизма является скорее дополнительным камушком в здании духовности людей, нежели руководством по восстановлению социальной справедливости.
Глубокая древность основных ветвей духовной культуры давно создала видимость их самодостаточности, поскольку они подчинены собственным законам. Язык и интеллект стремятся отражать все более глубокие сущности вещей, что ведет к созданию вербального дубликата действительности различных уровней. Религия строит особый воображаемый мир для аутсайдеров обыденного социума. Нравственности свойственно конкурировать с правилами естественного отбора и поощрять у слабых людей иллюзию свободы среди сильных. Наконец, искусство творит свой дубликат реального мира, где моделируется его дисгармония. Объективно все эти искусственные варианты самостоятельной действительности помещают людей в часы досуга в общеупотребительные правила поведения, что объединяет социум и отвечает древнейшей функции духовной культуры.
3.3. Цивилизация
Неолитическая революция рубежа плейстоцена – голоцена (11 700 календарных лет назад) не сразу преобразила ближневосточное общество. Поначалу ростки производящего хозяйства сосуществовали в нем с охотой и собирательством предшествующей мезолитической эпохи, а социальная структура оставалась первобытной.
Несмотря на признаки индивидуального разделения труда, мезолитическое общество (например, у бушменов пустыни Калахари) является глубоко однородным, эгалитарным [457]. Роли индивидов в подобном социуме обусловлены биологически. Они зависят от половозрастного фактора, физических и умственных задатков, что характерно, в принципе, для всех сообществ высших коллективных животных. Иными словами, естественный отбор на ранних стадиях развития производящего хозяйства не позволял ему сказаться на структуре первобытного общества.
Причина такого консерватизма вытекает из природы социума. Фигурально выражаясь, он жив до тех пор, пока нерушима его целостность. С точки зрения слепого социума, все, что угрожает ей, ничуть не лучше физической гибели. Сам социум, разумеется, ничего не решает. Однако нормы общественной жизни, выработанные за тысячелетия его истории, автоматически противятся всему, что расчленяет общество. Люди, подчиненные подобным нормам, подвержены мощным коллективным эмоциям. Это впечатление подчеркивает непреодолимость ситуации.
Между тем теоретически прогресс мыслим и здесь. Человеческое общество издавна находится в состоянии непрерывного демографического роста (см. разд. 2.4). Пока население отдельных сообществ не превышает 2,5 тыс. человек, они являются совокупностями объектов, в которых не действует статистический закон больших чисел. Точнее, он выполняется, но в том смысле, что поведение этих сообществ не отвечает своему математическому ожиданию, т. е. выглядит непредсказуемым.
Говоря отвлеченно, положение дел выглядит так. Допустим, мы имеем группу из сотни человек и, исходя из человеческой природы, знаем, каких поступков ждать от любого индивида. Мы в состоянии предсказать, как часто каждый из них будет делать одно, как часто другое и т. д. В результате появляется возможность предсказать, в каком усредненном состоянии будет пребывать группа в очередной момент времени, т. е. мы предскажем вероятность ее состояний, или определим их математическое ожидание.
Однако эта относительно небольшая группа никогда не оправдает наших ожиданий. По сугубо статистическим причинам она будет вести себя не усредненно, а, наоборот, ориентированно то в одну, то в другую крайность, как монета, которую подбрасывают десять раз, ожидая по пять «орлов» и «решек», но получая перекосы в пользу одного из вариантов. Однако, если подбросить монету 10 тыс. раз, результат совпадет с ожиданием почти идеально. Так выразится закон больших чисел (см. разд. 1.8).
Сообщество, состоящее из 10 тыс. индивидов, тоже выглядит предсказуемым. Понятно, отдельные люди станут вести себя произвольно, но, поскольку набор их реакций не бесконечен, среднее состояние сообщества раз от раза окажется относительно одинаковым, т. е. предсказуемым. Когда исход испытания представим и его математическое ожидание известно, в простых случаях вероятность пропорциональна квадратному корню из числа опытов или численности испытываемой совокупности объектов. Поэтому предсказуемость поведения сообществ численностью от 1 тыс. до 10 тыс. индивидов будет колебаться где-то от 31,6% до приблизительно 100% и составит около 50% для сообщества с численностью 2,5 тыс. человек. Проще говоря, после этого «демографического Рубикона» плотность населения будет скорее способствовать проявлению закона больших чисел, нежели наоборот.
Неолитический социум придирчиво «следил» за поведением своих членов и «констатировал», что оно непредсказуемо в силу первобытной малочисленности. Поэтому предсказуемость навязывалась этому социуму искусственно, путем поддержания его первобытной однородности, несмотря на то что возникшее производящее хозяйство создавало очевидные предпосылки для дифференциации неолитического общества по профессиональному признаку. Специализированные профессионалы присутствовали, но профессионально специализированные слои населения пресекались, благо малочисленность тогдашнего общества сама по себе не создавала условий для его крупномасштабного расслоения.
Затем в одном из ближневосточных регионов численность населения перевалила «демографический Рубикон» в 2,5 тыс. человек на сообщество. Теперь с довольно высокой вероятностью 50% общее поведение подобного сообщества становилось предсказуемым, независимо от сохранения или несохранения им своей однородности. Ввиду этого обстоятельства социум снимал ограничения на дифференциацию. Сообщество исторически немедленно принималось расслаиваться на профессиональные страты, объединяемые в четыре традиционные группы. Границы между ними еще долгое время оставались условными, и все же с известными оговорками их можно определить.
Группа производителей пищи, строго говоря, не была монополистом в своем занятии. Например, в раннем Шумере сельским хозяйством занимались и ремесленники, не говоря о том, что сохранялись охота с рыболовством и собирательством. Однако с определенной вероятностью можно сказать, что в нашем 2,5-тысячном сообществе наметился слой земледельцев и скотоводов, профессия которых заявила о себе в ходе неолитической революции.
Выделение тружеников сельского хозяйства создало предпосылки для обособления ремесла и торговли, о прогрессе которых свидетельствовало международное распространение ближневосточных торговых «жетонов» в раннем голоцене (см. разд. 3.2). Наконец, еще с первобытных времен общество располагало администраторами и служителями культа (вождями и колдунами), которые не преминули образовать подразделение умственного труда. Сформировалась отчетливо иерархическая структура общества.
Казалось бы, вслед за неолитической технологической революцией пришла социальная революция общественно разделенного труда (т. е. разделенного по общественным слоям). Однако 2,5-тысячное сообщество балансировало на хрупкой грани населенности, отвечающей 50%-ному проявлению закона больших чисел. Колебания биопродуктивности среды и социальные коллизии легко могли нарушить его равновесие, снизить плотность населения и уничтожить статистические условия для общественного разделения труда. Поэтому 2,5-тысячное сообщество являлось эфемерным.
Относительная стабильность могла наметиться в сообществе с населением 5625 человек, которое подчинялось закону больших чисел с вероятностью 75%. В подобном сообществе разделение труда утвердилось бы надолго и стало бы прогрессировать. Последнее обстоятельство содержало в себе и социальную угрозу.
Подразделения труда представляли собой заметные совокупности лиц со своими интересами и целями, которые во многом не совпадали между собой. Это противоречие было способно расколоть и уничтожить общество разделенного труда. Слепой, но чуткий социум допустить этого не мог. Стихийным образом он «запер» общества разделенного труда в города, окружил их крепостными стенами и таким искусственным, принудительным путем уберег от распада и краха. Появилось городское общество, цивилизация, которую можно определить как предметную форму структуры общества разделенного труда, призванную уберечь его от распада [51, с. 143]. Подразделения труда вещественно привязаны к структуре цивилизованного города (собственно, иных городов и не бывает) (рис. 10), а потому город объединяет их, словно архитектурный обруч. Изложенная модель генезиса цивилизации отвечает фактам.
Неолитическая революция произошла в субтропиках Ближнего Востока не случайно, поскольку там биопродуктивность среды оптимальна для земледелия. В тропиках она еще выше, однако тропический круговорот веществ настолько мощен, что не оставляет земледелию плодородных почв. В умеренном поясе круговорот вял – он позволяет формироваться богатым, плодородным почвам (черноземам), однако там не высока общая биопродуктивность среды [20, с. 154–159]. В субтропических долинах рек сочетание уровня биопродуктивности с темпами круговорота веществ наиболее выгодно для природопользования.
В начале голоценового подъема биопродуктивности среды (11 700 лет назад) продолжающийся позднемезолитический демографический взрыв спровоцировал возникновение производящего хозяйства, отмеченного в Шанидаре и Зави-Чеми-Шанидаре, северо-восточный Ирак, протонеолит, 12 170 календарных, или 10 600 радиоуглеродных лет назад; в Гандж-Дарехе, западный Иран, докерамический неолит, 11 940 календарных, или 10 400 ± 150 радиоуглеродных лет назад; в Иерихоне, Иордания, докерамический неолит А, 11 830–10 010 календарных, или 10 300–8720 радиоуглеродных лет назад.
За это время ближневосточное население выросло десятикратно. В поселении Абу Хурейра, Сирия, поздний натуф, 12800 календарных, или 11150 радиоуглеродных лет назад, население составляло 250 ± 50 человек (всего 15,8% проявления закона больших чисел). В докерамическом неолите В того же поселения, 10 790–9190 календарных, или 9400–8000 радиоуглеродных лет назад, население достигало 2500 ± 500 человек [62; 503] (50% проявления закона больших чисел, т. е. «демографический Рубикон»).
При таких плотностях населения социум уже позволил бы общественное разделение труда и городскую цивилизацию. Это осуществилось в первом ближневосточном (и мировом) городе Иерихоне докерамического неолита А, созданном по фортификационной модели урбанизации (см. ниже) и отмеченном высокой каменной башней (7,75 м), фланкированной видной стеной (5,75 м), которые прикрывали гражданскую застройку из круглых, обычно однокомнатных домов из сырцового кирпича [500, с. 531]. Эта строительная техника ведет начало из натуфийской культуры поселения Бейда в Иордании, где открыто сооружение из сырцового кирпича [500, с. 115]. Как видим, и в области архитектуры социум придерживался методов оживления дремлющих достижений под влиянием демографического всплеска (см. разд. 3.1).
Рис. 10.
Обобщенная схема цивилизованного города, отражающая типичную структуру человеческих городов на протяжении последних 11 700 лет. Обычно значительный город окружен фортификационной системой, состоящей из крепостных стен и башен # (сейчас это системы противовоздушной обороны). В центре города находятся административные здания (резиденции правителей) + и святилища ×. Пространство между административным центром и фортификационными сооружениями занято жилыми постройками О, мастерскими (ныне предприятиями) & и торговыми местами (рынками, торговыми и меняльными лавками – ныне рынками, магазинами и банками) $. Город погружен в неразрывно связанные с ним сельскохозяйственные угодья ∞
. Первый город нашей планеты Иерихон (в современной Иордании) 11 700 календарных лет назад имел стену с башней. Второй по времени город Чатал-Хююк (в современной Турции) группировался вокруг святилищ «квартала жрецов» 9420 календарных лет назад. Современные города соответствуют их модели
Ближневосточный пример следует типичной модели демографического взрыва (см. разд. 2.4). Первоначальный подъем населения (16 тыс. лет назад) вызвал значительные миграции прашумеров, сино-кавказцев и ностратов из Леванта во все стороны света. Затем прирост населения стал обгонять темпы убыли мигрантов, и популяционная плотность подскочила вдесятеро за 2 тыс. лет (12,8–10,8 тыс. лет назад в Абу Хурейра). Демографический удар вызвал неолитическую и городскую революции.
В эпоху первогорода Иерихона население балансировало на грани «демографического Рубикона». Любые неблагоприятные изменения обстановки могли оборвать локальную урбанизацию. Поэтому городская эпоха в Иерихоне продлилась 11,8–10,0 тыс. лет назад и прекратилась. В окружающем регионе продолжалось углубление разделения труда и начал урбанизации.
В иорданском поселении Бейда на стадии среднего докерамического неолита В, 10160–9820 календарных, или 8850–8550 радиоуглеродных лет назад, появились пекарня, лавка мясника и мастерские. В турецком Хаджиларе эпохи керамического неолита, 8250–8030 календарных, или 7170–6990 радиоуглеродных лет назад, возникла гончарная мастерская. В турецком Мерсине 16, среднего халколита эпохи Амук D, 7410–6950 календарных, или 6450–6050 радиоуглеродных лет назад, отмечены начала фортификации.
Второй по времени город, Чатал-Хююк, Конья, Турция, ранний керамический неолит, 9420–8440 календарных, или 8200–7350 радиоуглеродных лет назад, строился по храмовой модели урбанизации (см. ниже), о чем говорит «квартал жрецов» с украшенными зданиями-святилищами, прототипами храмов, в отсутствиe фортификации. Отмечено орошаемое сельское хозяйство. Население оценивается в 5000 человек [500, с. 197–198] (70,7% проявления закона больших чисел). Просуществовав 980 лет в течение фазы Амук А, город прекратил свое существование, оставив по себе смежное поселение Чатал-Хююк-Запад. Оно продолжило последовательность обитания в течение фазы Амук В, 8670–7980 календарных, или 7550–6950 радиоуглеродных лет назад, продлив жизнь на равнине Конья до общей продолжительности в 1440 лет, т. е. несколько меньше, чем в Иерихоне, поскольку исторический процесс ускорялся с течением времени ([90, с. 26-37]; cp. разд. 3.1), и сроки сжимались.
Сравнение сроков жизни первогородов Иерихона (1820 лет) и Чатал-Хююка (1440 лет) не избавляет от впечатления, что они пресеклись автоматически, в силу статистических флюктуаций под влиянием каких-то конкретных пусковых факторов. Первой стабильной цивилизацией с известной демографией стал Шумер. Население Архаического Ура (ранне-династический период I, 2750–2615 до н. э.) составляло ок. 6000 человек по всей округе. Оно предполагало 77,5% проявления закона больших чисел, что переваливало за спасительный для цивилизации рубеж относительной стабильности в 75% (см. выше). Население округи Шуруппака, раннединастический период II, 2615–2500 до н. э., уже превышало 15–20 тыс. человек, что предусматривало свыше 100% выражения закона больших чисел. В Нгирсу, столице Лагаша, ~ 2340–2318 до н. э., обитало 17 500 ± 2500 жителей. В Мохенджо-Даро, Пакистан, 4710–4250 лет назад насчитывалось 40 тыс. жителей. Кносс, cтолица о. Крита, Греция, 1700–1580 до н. э., вмещал 100 тыс. человек [14; 45, с. 167, 174, 203]. За вычетом Мохенджо-Даро, пришедшего в упадок не без участия вторгнувшихся арийцев, остальные цивилизации дошли до исторической эпохи, не проявляя признаков внутренней нестабильности.
Рассуждая более общо, отметим, что прочные статистические основы цивилизации позволили ей дойти до наших дней без существенных перемен в предназначении и внешней форме. Современный цивилизованный человек полностью ангажирован цивилизацией. Он просыпается поутру не тогда, когда выспался, проголодался или по иному своему усмотрению, а тогда, когда будильник призовет его на работу или службу. Он отправится туда не собственным путем, а на общественном или личном транспорте, пользуясь дорогами, проложенными не им.
Рабочий день он посвятит занятиям, разработанным, как правило, до его рождения, подкрепляя силы в установленные не им перерывы. После трудового дня он отдыхает и развлекается не по велению души, а сообразно принятому цивилизацией распорядку часов досуга. Соответствующие зрелища, увеселения и забавы предлагаются специальными людьми тоже не по своему усмотрению, а в соответствии с принятыми традициями. Намаявшись, цивилизованный человек отходит ко сну не с заходом солнца, а по расписанию, приличествующему послушному члену общества. Ночью он мирится лишь с очерченным кругом снов, а при его расширении отправляется на прием к психоаналитику.
Подобный образ жизни, по существу, напоминает распорядок тюрьмы или зоопарка, а вовсе не крейсирование «хозяина жизни». Между тем переносимые ограничения нивелируют цивилизованных людей, превращая их в винтики слаженного механизма, далекого от разлада, невзирая на чрезвычайно углубившееся разделение труда. Нетрудно догадаться, что мы наблюдаем социально-интегративные функции цивилизации в действии. Сама она все та же, что на заре своих времен. В центре городов господствуют административные здания и храмы. В положенных местах учреждены торговые точки и производственные предприятия. Города прикрыты системами обороны и окружены сельскохозяйственными угодьями. Подобная инфраструктура ведет свое начало из эпохи ранней цивилизации. Устройство цивилизации и ее образ жизни весьма жизнеспособны, выгодны человеку и переносятся им с полным основанием в собственных интересах.
Отличительная черта цивилизованных обществ состоит в непременной иерархической стратификации патриархального характера, когда все социально значимые посты пирамидально подчинены друг другу и принадлежат мужчинам. Происхождение этой универсальной особенности объясняется действием этологического закона Крука (см. разд. 3.1). Цивилизованные люди, оказавшись запертыми в городах, подсознательно чувствовали себя, как все приматы в неволе. В зоопарке автоматически складывается патрилинейная иерархия (господство самцов в пирамидально устроенном сообществе) [840]. Это происходит потому, что, невзирая на регулярную и достаточную кормежку, животные драматически переживают отсутствие свободного доступа к пище, что инстинктивно воспринимается ими как пребывание в скудном, пустынном биотопе. В соответствии с законом Крука у них складывается патрилинейная иерархия. С людьми в городах происходит то же самое.
Формирование ранних цивилизаций подчинялось определенным закономерностям. К становлению городов привело самодвижение средств коллективного производительного потребления. Проще говоря, стихийное развитие человеческих технологий в подходящей демографической обстановке спровоцировало общественное разделение труда, с дезинтегративным влиянием которого социуму пришлось бороться путем формирования городов. Совершенствование технологий обусловлено стихийным развитием коллективных орудий, т. е. самодвижением средств коллективного производительного потребления.
Поскольку к общественному разделению труда и его угрозе для целостности социума привели средства коллективного производительного потребления, борьба с этими негативными последствиями тоже должна была осуществляться средствами коллективного, но непроизводительного потребления, поскольку производительная деятельность заинтересована в безусловном приросте технологий.
Из числа средств коллективного непроизводительного потребления самыми древними и органичными для людей являются жилища и их ансамбли, поскольку человеческие стоянки устроены наподобие таковых у шимпанзе [384]. В структуре поселения к числу наименее производительных относятся культовые и фортификационные сооружения, поскольку ни те, ни другие не предназначены для созидательного труда. Ожидалось бы, что они станут системообразующими началами цивилизации.
Инфраструктура цивилизации (например хараппанской) отражает и опредмечивает структуру общества разделенного труда: в раннем городе мы видим кварталы ремесленников, торговые места, административные и культовые здания [16, с. 5, 7; 119, с. 191–192]. Они материально увековечивают устройство общества и тем самым оберегают его единство. Однако сооружения, связанные с ремесленной, торговой или административной деятельностью, являются преимущественным достоянием соответствующих профессиональных групп. Только святилища и крепости одинаково принадлежат обществу в целом. Поэтому ранние цивилизации складывались внутри крепостных стен или вокруг храмов.
Фортификационная модель урбанизации присуща, как мы показали выше, иорданским Иерихону и Мерсину, а также древнейшим столицам древнеегипетстких номов (поначалу – малых царств, потом – областей Нижне– и Верхнеегипетского царств). Храмовая модель урбанизации свойственна Чатал-Хююку, древнейшим центрам шумерских округ, центрам древнеиндийской, хараппанской цивилизации (чья государственность отрицается вообще [119, с. 182–205]) и даже центрам мегалитической протоцивилизации Британских островов, где монументальные постройки ограничивались кругом культово-астрономических сооружений.
Принимая во внимание возраст и географическое положение Иерихона, Чатал-Хююка и других памятников ранней цивилизации, можно сказать, что фортификационная модель урбанизации тяготела к центру раннего цивилизованного мира, а храмовая – к его окраинам. Это объясняется тем, что в центре всякого популяционного ареала плотность населения выше, чем на периферии. Высокая плотность населения у людей издавна чревата территориальными конфликтами, что обусловливало нужду в фортификациях. По той же причине государственная власть «центральных» цивилизаций являлась военизированной, что отличало древнеегипетских фараонов и номархов. Напротив, власть у периферийных цивилизаций несла оттенок сакральности, что было не чуждо шумерскому, хараппанскому, минойскому (древнекритскому) и микенскому (доантичному древнегреческому) обществам, а также мегалитической протоцивилизации Британских островов.
В силу тех же отношений древнего центра и молодой периферии раннего цивилизованного мира хронологический примат цивилизованности и государственности принадлежит иорданскому Иерихону (ок. 11 700 календарных лет назад) и Нижнеегипетскому царству, датируемому изображением нижнеегипетской короны на сосуде амратской эпохи Нагада I (6600–6400 календарных, или 5744 ± 300–5577 ± 300 радиоуглеродных лет назад), против сакрального Чатал-Хююка (9420–8440 календарных, или 8200–7350 радиоуглеродных лет назад) и I династии Киша в Шумере (ок. 4900 календарных лет назад).
Цивилизация явилась закономерным итогом развития человечества. Как мы помним, в основе эволюции человека лежала неотения (см. разд. 2.4), которая в общем и целом побуждала людей к малоподвижному образу жизни. Исключения составляли эпизоды акселерации, когда люди активно расселялись по свету и даже устремлялись в космос (как сейчас). Однако тяга к перемене мест всегда являлась только лаком на эволюционно заложенной в нас тяге к малоподвижности (многие люди это знают по себе, в особенности женщины). Цивилизация сообщила нашим предкам глубоко оседлый, «принайтованный» образ жизни и тем самым «угодила» их глубинной эволюционной сути. Понятно, они об этом не знали – зато охотно повиновались воле социума и своим эволюционным задаткам, возводя города.
Анализ основ ранней человеческой цивилизации приводит к недвусмысленному выводу, что весь ход ее становления носил естественно-исторический, автоматический характер. Для уяснения закономерностей ее генезиса и раннего функционирования нам не потребовалось обращаться к сознательному человеческому фактору. Правда, нам постоянно приходилось апеллировать к социуму как сверхорганизму, однако это надчеловеческое образование всегда действовало слепо и стихийно.
Равнодушие к сознательно действующему человеку следует рассматривать как положительный момент исследования. В самом деле, поиск объективных закономерностей организации человеческой жизни позволяет лучше понять природу человека, в то время как гуманистический пилотаж предмета в интересах его ценностей обычно ведет к элементарной тавтологии. Например, на вопрос: как человеку удалось создать культуру и цивилизацию? – люди подсознательно отвечают: в силу того, что человек был умным. А как он стал умным? Потому что его воспитали культура и цивилизация. Подобные результаты не представляют научного интереса. Таким образом, нам удалось показать на фактах, что начальные формы культуры и цивилизации сложились сугубо стихийно и, лишь состоявшись, подтянули человека до уровня разумного существа. Став таковым и высокомерно забыв о своем происхождении (что для людей вообще типично), он, само собой, утверждает, что «сделал себя» сам. К таким убеждениям следует относиться с осторожностью.
Не претендуя на удачность данного конкретного исследования, отметим, что его замысел содержал известный положительный заряд. Попытка широкого охвата феноменов ранней цивилизации, предпринятая с объективистских позиций, способна не просто принести некий результат из сферы гуманитарного знания. Имеется определенная возможность получить сведения о закономерностях становления и развития цивилизации, сопоставимые с данными о живой, но внечеловеческой природе, и широком мире в целом.
В частности, мы нашли, что цивилизация, помимо общеизвестных антропных свойств, отличается рядом статистических особенностей, роднящих ее с физическим миром. На наш взгляд, она подчинена действию объединительных начал, что тоже универсально для обширного мира. Можно добавить, что цивилизация являет собой пример иерархически устроенного объекта, в чем она также совпадает с объективным миром (см. разд. 1.8).
Заключение
Рассмотрев физические, астрофизические, геологические, климатические, биологические и социальные основы нашего мира, мы находим, что его устройство повсеместно пронизано закономерностями различной степени общности. Это свидетельствует об универсальном распространении закона причинности в соответствии с версией о его происхождении на заре Вселенной (см. разд. 1.6). Лишь в первые мгновения бытия Вселенной однотипные закономерности могли охватить ее всю. Затем она стремительно раздулась в инфляционной фазе, и различные ее области потеряли «оперативную», взаимообразную связь между собой спустя 10-32 с после Большого Взрыва (см. разд. 1.2). С этого момента обмен законами природы между различными уголками Вселенной сделался проблематичным. Отсюда следует, что основополагающие закономерности нашего мира родились, едва он вышел из состояния сингулярности. Это позволяет понять, почему законы природы повсюду так однотипны и слажены (это называется принципом относительности Галилея (1564–1642), согласно которому все движущиеся системы – в данном случае различные области расширяющейся Вселенной – подчиняются одним и тем же законам природы) (см. разд. 1.8). В противном случае Вселенная давно бы развалилась на куски, и мы ничего бы не знали о ее удаленных частях. Дело не ограничивается затронутыми сферами бытия.
К примеру, мы не имели подходящего случая подробнее коснуться проблематики природы информации (см. разд. 3.2), тогда как она напрашивается сама собой в связи с обстоятельствами рождения причинно-следственного закона. Напомним, что он возник в результате сочетания законов сохранения с принципом неубывания энтропии, а последняя существенна для самого понятия информации. Согласно количественной теории информации [116], определенное количество информации содержится в сведениях, снижающих неопределенность выбора образа действия.
Если нам предстоит выбор из двух вариантов поведения, а полученное сообщение позволяет предпочесть один из них, мы получили 1 бит информации. Информация в более сложных случаях также поддается переводу в биты (байты).
Когда мы пребываем в состоянии неопределенности выбора образа действия, мы обречены на слепые, «броуновские» поступки, характеризующиеся высокой энтропией (беспорядочностью). Информация позволяет привнести в нее порядок. Его степень обратно пропорциональна свободе выбора и в идеале стремится к состоянию, присущему древней сингулярности (см. разд. 1.3). Следовательно, информация – это один из негэнтропийных реликтов сингулярности в свободно расширяющейся Вселенной. Эти негэнтропийные реликты сингулярности ответственны за нетипичные для Вселенной начала: формирование небесных тел и их систем, биогенез в невыясненных условиях (см. разд. 2.1), целесообразность живых организмов и разумных существ (людей). Мы называем эти феномены нетипичными для Вселенной потому, что в основе ее жизни лежит тенденция к общему расширению, а оно способствует росту отнюдь не упорядоченности, а, наоборот, общевселенской энтропии (см. разд. 1.4).
В силу своего количественного и качественного превосходства над животными в интеллекте (см. разд. 3.2) люди воспринимают как информацию сведения из различных (не только биологически значимых) областей действительности, поэтому массив целесообразности у них также незауряден. Эта особенность помогает ориентироваться в сложных условиях цивилизации, что создает иллюзию ее сознательного созидания.
Можно предсказать достижимую сложность человеческой цивилизации в земных условиях. В настоящее время планету населяют 30 ± 20 млн видов живых существ [555, с. 1447]. Каждый вид занимает особенную экологическую нишу, так что их численности эквивалентны. Размножающееся человечество стоит на пути абсолютного овладения планетой. Составной частью подобной задачи является освоение земных экониш. Для этой цели цивилизация со временем изыщет технические средства, которые когда-нибудь определят ее технологическую сложность, заведомо превосходящую 30 млн операций – того минимума сложности цивилизации, необходимого для освоения 30 млн земных экониш (заметим также, что прямолинейное истолкование демографо-технологической зависимости позволяет предполагать, что степень сложности человеческой технологии количественно близка численности человечества, так что уже сейчас при численности человечества в 6,5 млрд мы располагаем общепланетной технологией, состоящей из порядка 6,5 млрд операций, т. е. отдельных действий, необходимых для производства наличных инструментов, – действий, помноженных на число разновидностей этих инструментов).
Оседлая цивилизация вкупе с производящим хозяйством явились логичным результатом отбора, породившего в свое время маломобильных и прожорливых гоминин (см. разд. 2.4). В обоих случаях итоги предусматривали малоподвижность и обильное питание. Естественный отбор представляет собой воплощение причинно-следственного закона, способствующего удержанию повторяющихся явлений и схожих сущностей в ущерб спорадическим и неповторимым, которые хуже отвечают его природе. Благосклонность к шаблонам со стороны закона каузальности обусловила повторяемость лидирующих форм в биологической эволюции нашей планеты, чередующихся в поколениях высокомобильных и менее подвижных существ (см. разд. 2.3). Ввиду этого мы вправе ожидать, что и у человечества предпочтительным правом на существование пользовались прежде всего повторяющиеся явления. Это отражает не ограниченность нашего вида, а его выгодное соответствие требованиям закона причинности, обещающее долгую жизнь.
От таких медлительных существ, как наши предки, не следовало бы ждать тех эпизодов взрывной экспансии, о которых мы подробно говорили (см. разд. 2.4 и 3.1). Неотеничная природа не могла запретить гомининам тяги к обычным для живых организмов миграциям (пример – степенное распространение неотеничного Homo heidelbergensis по Евразии). Однако мгновенные (по историческим меркам) броски через континенты настораживают. Их причина состоит в том, что они совершались гомининами в состоянии акселерации, которая временно затушевывала их неторопливую неотеничную природу. Это хорошо видно в наше время, когда современный акселерированный человек мечтает об экспансии в космических масштабах и делает в этом направлении первые успешные шаги.
Другое воспроизведение архетипов былых эпох заметно в сфере социальной психологии. Акселерированные гоминины достигали зрелости поспешно, а потому не могли избежать психологической инфантилизации. Она помогла им отказаться от психологических стереотипов и произвести серию технологических революций. Современная эпоха позволяет рассмотреть процесс социальной инфантилизации детальнее.
Подобно прошлым стадиям акселерации, мы переживаем бурную научно-техническую революцию с агрессивной ломкой стереотипов, объясняющейся инфантилизмом участников, которым не жаль вековых традиций. Духовно омолаживающемуся человечеству эта научно-техническая сторона дела выгодна. Более спорна психологическая инфантилизация современной массовой культуры. Мы переживаем как бы второе детство человечества, визитной карточкой которого является вал однообразных небылиц, составляющих основу голливудской кинопродукции.
Если говорить о прошлом, то что-то подобное можно усмотреть в кощунственном появлении светского искусства после неолитической революции (до неолита исскуство было преимущественно религиозным, мифологическим). Акселерированные кроманьонцы Западной Европы отличились в верхнем палеолите тем, что перенесли свои святилища в глубокие, порой глубочайшие пещеры, что говорит скорее не о здравомыслии, а о юношеском задоре. Можно надеяться, что и современное упрощение художественного творчества выльется во что-то перспективное.
В своем исследовании мы позволили себе достаточно вольно обращаться с некоторыми устоявшимися мировоззренческими постулатами. Это происходило не только потому, что открылись какие-то новейшие сведения, революционизирующие науку, но и вследствие того, что иные мировоззренческие постулаты не во всем до конца осмыслены. Скажем, в астрономической науке и философии прочно утвердилось представление об изотропности, повсеместной однородности Вселенной. При всем удобстве такого подхода трудно не увидеть, что он плохо гармонирует с элементарными следствиями теории Большого Взрыва. Если когда-то произошел Большой Взрыв, то он имел определенный эпицентр, пусть и незаметный сейчас в силу большой давности. Это нехитрое умозаключение, на первый взгляд, ни к чему не обязывает. Однако совпадение расчетного расстояния Земли до центра Вселенной с эмпирическим расстоянием до Великого Аттрактора ставит все на свои места (см. разд. 1.2). При этом анизотропность Вселенной представляется не результатом совпадений, а следствием теории Большого Взрыва – следствием, до сих пор недооцененным.
Аналогичным образом обстоит дело с предложенными нами представлениями о направленности биологической эволюции. Дарвинизм исходит из того, что облик живых существ является не результатом претворения какого-то целенаправленного проекта, а делом случая, продуктом воздействия слепого естественного отбора. Мы исходим из того, что в изначально случайной Вселенной естественный отбор должен был истребить случайные, нестойкие формы организации материи и оставить в живых лишь устойчивые, неслучайные. Именно поэтому Вселенная состоит не из какой-то массы случайных форм, а из повторяющихся образцов в виде галактик, звезд, планет и их обитателей. В результате естественного отбора выжили не какие попало биологические виды, а только те, что отвечали колебаниям среды. Если колебания среды повторялись, то биологические виды, следуя тенденции, принимали принципиально подобные формы, конвергентные не только внешне, но и внутренне. Поэтому мы не видим расхождений с дарвинизмом в изложенной нами идее направленности биоэволюции.
Наши предки тоже возникли как результат определенной направленности развития живого. Нам глубоко чужда антропофильная («человеко-предвзятая») идея о том, что человек возник для воплощения высшего разума. На наш взгляд, она не отвечает не только научным фактам, но и здравому смыслу. Будь разум человеческого типа биологически выгодным, он давно утвердился бы у других животных, например у по-человечески двуногих динозавров, имевших хватательные верхние конечности с отстоящим «большим» пальцем, пригодным для развития точного захвата. Разум выгоден лишь социуму, притом не всякому, а именно человеческому (см. разд. 3.2). В то же время человек закономерен, однако происхождение его физической и психологической организации лежит не в сфере духа, а в области эволюционной экологии. Человек телеологичен (целесообразен) лишь в том смысле, что играет по правилам биосферы, а вовсе не парит над ней. На наш взгляд, его интеллектуальные победы в таком контексте впечатляют еще больше.
История человечества шла неравномерно. Она не просто ускорялась, но убыстрялась отчетливыми рывками во время технологических и интеллектуальных революций. Природа этого явления в свете изложенного (см. разд. 2.4 и 3.1) видится нам таким образом. В силу демографо-технологической зависимости в периоды демографических взрывов у людей усложнялись технологии и наступала акселерация. Последняя сопровождалась инфантилизацией психики и готовностью к отказу от традиций. Это не просто давало зеленый свет технологическим новшествам, но способствовало переработке на их основе социальной психологии.
В подобную модель укладываются узловые точки интеллектуальной и технологической истории человечества. Это позволяет понять происхождение культурной окраски человеческого прогресса. Нынешний его этап пронизан особенностями западной культуры. Причина этого заключена не в расово-культурных обстоятельствах, как могло бы показаться, а в более существенной подоплеке.
Западноевропейский демографический взрыв XI – сер. XVI вв. и его современное продолжение позволили очагу технологической революции разместиться в Западной Европе. Одновременно с началом промышленной революции (c XIV в.) открылась культурная революция эпохи Возрождения (с XIV в.), однако совершенно очевидно, что между этими событиями нет никакой причинно-следственной связи. Первые мануфактуры едва ли влияли на литературу и искусство Ренессанса, а Ренессанс вряд ли заглядывал на производство. Их связь была опосредована демографо-технологической зависимостью. Рост плотности населения вызвал к жизни сложные технологии, а зарождающаяся акселерация омолодила психику, что, собственно, и выразилось в Ренессансе. С тех пор Западная Европа испытала множество скачков в технологии и социальной психологии. Дело дошло до того, что революции в культуре вошли в моду и породили свои имитации: различные формы модернизма, которые ничего не революционизируют, а лишь по-детски разыгрывают взрыв новых форм. Различные манифесты модернистов – это естественное побуждение здоровой инфантильной психики, крепнущей в Западной Европе с конца XIX в.
Меньшими темпами, но по той же схеме технологическая и культурная революции разворачивались на Ближнем Востоке с начала голоцена (11 700 лет назад). Сначала произошел демографический взрыв (16 тыс. лет назад), затем возникли производящее хозяйство, цивилизация и непрагматическая наука. То обстоятельство, что цивилизованные люди старательно развивали систему во многом непрагматических знаний, выдает глубоко инфантильную природу их психики.
Древняя Греция находилась на периферии тогдашнего цивилизованного мира. Зародившаяся было Микенская цивилизация рухнула в ходе двухступенчатого движения «народов моря» (ок. 1200 до н. э.), разбросавшего участников по Восточному и Центральному Средиземноморью (от Палестины до Италии). Оправившаяся после «темных веков» Греция дала свою классическую культуру, и в ее рамках развернулось «греческое чудо». Непрагматичная ближневосточная наука была поднята на щит и дала такую игру ума, как философия. Ее рождение было верхом непрактичного инфантилизма, что давало повод хорошеньким и остроумным служанкам высмеивать первого философа Фалеса [104, с. 107, № 9]. Позже выяснилось, что в результате его отрешенных занятий произошел прорыв в сущность вещей.
В Индии и Восточной Азии ситуация выглядела сложнее. Демографический подъем и предпосылки производящего хозяйства имели там местное значение. Но в результате ближневосточного демографического взрыва туда устремились мигранты с запада: ностратические дравиды – в Индию, а синокавказцы (синотибетцы) – в Китай. Существуй в те времена оперативная связь между народами, тогдашняя эпоха прошла бы под эгидой Ближнего Востока от Средиземноморья до Тихого океана.
Верхнепалеолитическая технологическая революция ограничилась Европой (как основой) и Ближним Востоком (как периферией). В ней практически одновременно участвовали два вида гоминин: сапиенсы (50 тыс. лет назад, возраст ориньякской культуры) и неандертальцы (37 тыс. лет назад, возраст перигордийской культуры). Это доказывает, что расовые (здесь – видовые) различия не имели значения в деле прогресса. Неандертальцам не повезло потому, что они начали свою часть технологической революции позже сапиенсов.
Инфантилизм верхнепалеолитических сапиенсов состоял в том, что они с ненормальным энтузиазмом предавались непрагматичной духовной культуре, о чем свидетельствует размах их монументального наскального искусства, когда украшалось по одному святилищу за каждые 80 лет (см. приложение 4). Украшенные пещеры служили святилищами, поэтому их расцвет говорит не только о революции в художественном творчестве (отмеченной и в портативном, мобильном искусстве), но и о взрывном подъеме в мифологии, культе, религиозном церемониальном исполнительском искусстве (музыка, вокал, танец) и вообще в религиозной жизни. Сочетаясь с календарной арифметикой, т. е. с зачатками математики и астрономии, сакральный художественный подъем ознаменовал собой интеллектуальный расцвет в целом. Необходимая для этих успехов социальная сплоченность косвенно указывает на благоприятное положение дел в нравственной сфере духовной жизни. По совокупности признаков мы получаем культурную революцию с поправкой на примитивность времен.
Homo ergaster заслуживает тех же умозаключений. У него произошли демографический взрыв (ок. 1,9 млн лет назад) и ашельская технологическая революция (ок. 1,6 млн лет назад). В промежутке между этими событиями наступила акселерация (св. 1,81 млн лет назад, возраст акселерированного ребенка из Моджокерто), а вместе с ней и инфантилизация. Она выразилась в учреждении основ непрагматической духовной культуры, биологические предпосылки которой вышли из обезьяньего прошлого, а оформление в человеческих архетипах (речь – интеллект, магия – религия, нравственность и, отдельно, искусство) состоялось в эпоху «человека-мастера».
Kenyanthropus rudolfensis отличился тем же. Наши знания об этом гоминине довольно скромны. Обобщенно можно сказать следующее. Его предок, произошедший от гоминина Kenyanthropus platyops, под влиянием демографического роста сперва перешел в разряд хищников, а потом снова испытал демографический подъем и отозвался на это событие освоением средств коллективного производительного потребления, коллективных орудий, которые первоначально уступали индивидуальным в эффективности.
Подобное нерасчетливое достижение также могло быть следствием инфантилизма. Приблизительно 2,2 млн лет назад «кениец» пережил демографический взрыв и выплеснулся из Африки в Евразию. Сопутствующая акселерация вызвала увеличение его размеров, инфантилизацию и, возможно, усвоение такой непрагматичной формы общения, как язык жестов, обслуживающий другое не вполне выгодное достижение – коллективную орудийность.
Разумеется, духовная культура служила интересам выживания социума и в этом была выгодной. Однако развивающие ее гоминины не могли этого знать и предавались духовной жизни не из выгоды, а от души, т. е. на манер игры, по причине инфантильности. Аналогичным образом коллективные орудия в перспективе также являлись выгодным приобретением, так как после стадии усложнения и специализации (технологической революции) приобретали повышенную эффективность и становились выгодными. Однако «кениец» тоже не мог этого знать и погружался в орудийную деятельность с долей опрометчивости, т. е с той же инфантильностью.
Обрисованная нами модель человеческого прогресса во все времена подчинялась однотипным правилам, что выдает их дочеловеческую и догомининную древность. В сущности, история наших предков выглядела так. Неторопливые, неотеничные гоминины вели себя выдержанно и пользовались традиционными достижениями предшествующих эпох, не стремясь их совершенствовать специально (о чем свидетельствуют «спящие открытия», см. разд. 3.1). Затем их постигал демографический взрыв, популяция претерпевала акселерацию, и акселерированные гоминины производили в силу инфантильности технологическую и культурную революции. Плоды их трудов наследовали застойные неотеники, и ситуация повторялась на очередном уровне. Если говорить формально, прогрессивные акселераты в силу детской исторической наивности работали во благо исторически медлительным, т. е. примитивным, неотеникам. В итоге примитивные существа наследовали социум, обновленный по-детски бесхитростными акселератами.
Данный тезис напоминает положение вещей в эволюционной биологии (см. разд. 2.3): «примитивные наследуют Землю». Точнее говоря, прогрессивные, специализированные организмы виртуозно и необратимо приспосабливались к наличной среде, спаивались с ней навечно, а потом гибли при наступлении крупномасштабных перемен. Живущие в их тени менее конкретно приспособленные к обстановке, генерализированные организмы попадали под эволюционный удар в меньшей степени и поэтому автоматически наследовали изменившуюся среду. Оказавшись там «вне конкуренции», они выходили на оперативный простор, осваивали биоту, узко приспосабливались к ней и гибли при очередных крупномасштабных переменах. Их место доставалось более примитивным существам, до сих пор пребывавшим на задворках экосистем. Подобные чередования флор и фаун сопровождались массовыми вымираниями и повторялись неоднократно.
По этой причине общая типология развития биосферы тоже обнаруживает признаки наследования каких-то более широких принципов существования бытия, подобно тому как социальная жизнь унаследовала типологические основы развития биосферы для организации собственного мира. Источник принципов движения биосферы лежал в физическом мире, где своего рода примитивные объекты тоже наследовали Вселенную.
На заре Вселенной ее жаркое пространство было наполнено энергичными, преобладающими суперчастицами и вялыми, примитивными частицами современного типа. По мере остывания Вселенной дистанции характерных взаимодействий суперструн выросли, что стало губительным обстоятельством для массивных, близко взаимодействующих частиц. Их пул прекратил существование. С уходом суперчастиц Вселенная досталась скромным суперструнам современного типа, которые могут рассматриваться как примитивные объекты ввиду небольших масс и энергий, размах которых определял внутреннюю сложность суперструн (см. разд. 1.1). С известной условностью можно сказать, что примитивные формы суперструн унаследовали расширяющуюся Вселенную.
В области астрофизики происходило то же самое. При распаде единого взаимодействия суперструн в эмбриональной Вселенной первой обособилась гравитационная сила, которую по причине ее максимальной древности среди взаимодействий можно считать примитивной по определению. Ее фактическая примитивность вытекает из генерализованности. Гравитации подвержена вся материя Вселенной без исключения, даже таинственное «темное вещество» (см. разд. 1.1 и 1.2). Поэтому сила тяготения никак не специализирована, примитивна. Другие силы более специализированы. Сильное взаимодействие ориентировано на кварки. Электромагнетизм специализируется на заряженных частицах, кварках и электроноподобных лептонах. Слабое, милли– и сверхслабое взаимодействия тоже ограничиваются в основном узкой жизнью кварков. Вместе с сильным взаимодействием оно вообще ограничено микромиром. Проще говоря, негравитационные силы специализированы, а гравитация генерализована, примитивна.
Между тем, основные астрофизические объекты – планетные и звездные системы, галактики и их скопления, сама Вселенная, сохраняемая как целое тяготением, – все это зиждется на примитивных гравитационных полях и, следовательно, наследует Вселенную в силу собственной примитивности. Получается, что физический макромир, как и микромир, тоже подчиняется принципу «примитивное наследует Вселенную».
Более того, если справедливы изложенные представления (см. разд. 1.2) относительно уже начавшегося 605 млн лет назад коллапса Вселенной, то главенствующую роль в этом фатальном для нас процессе играет как раз гравитация, и, следовательно, именно она «задернет занавес» нашего мира, т. е. ненадолго унаследует его всецело. Можно спросить: откуда гравитация знала, что она примитивна и должна «закрыть» Вселенную? Она, конечно же, ничего о своей судьбе не знала. Просто в силу собственной примитивности тяготение меньше других взаимодействий пострадало от расширения Вселенной (в противоположность чему сильное и слабое взаимодействия замкнулись в микромире, а электромагнетизм сохранил силу только для 10% вещества Вселенной, оказавшись выдворенным за порог «темного вещества»). Поскольку тяготение пострадало меньше других взаимодействий, оно в соответствии со своей живучестью принялось дирижировать концом Вселенной. Здесь мы излагаем философское видение проблемы – для астрофизика здесь вообще нет проблемы, поскольку возможное гравитационное сжатие Вселенной является органичным положением теории пульсирующей Вселенной, в которой за расширением следует коллапс.
Это правило («примитивность наследует Вселенную»), универсальное для физического, биологического и социального миров, возникло в результате действия причинно-следственного закона. Он поощряет повторяющиеся явления и сущности и отбраковывает неповторимые. Прогрессивные, специализированные явления и сущности тонко организованы, индивидуальны. Поэтому закон причинности воспринимает их отрицательно и периодически «выпалывает» из бытия. Напротив, примитивные, неспециализированные явления и сущности больше похожи друг на друга в силу своей обобщенности, генерализированности. Закон каузальности воспринимает их с симпатией. Поэтому, к нашему удивлению и негодованию, именно они наследуют Вселенную и ее уголки после очередных ревизий со стороны причинности, хотя нам бы хотелось, чтобы прогрессивное прогрессировало дальше, а не гибло непонятно почему. Примеры бесчисленны. Так, великолепную Римскую империю, разложившуюся без видимых причин, унаследовали отталкивающие варвары, а некоторые их этнонимы (вандалы, гунны) стали для нас скверно нарицательными…
Биологический принцип естественного отбора и выживания объектов, наиболее приспособленных к условиям среды, можно переформулировать и распространить более широко, нежели это принято. Селекция со стороны закона каузальности сохраняет повторяющиеся явления и устраняет неповторимые, случайные. Объекты, отвечающие окружению, например биологической среде, вместе с обстановкой образуют повторяющуюся систему, а потому благополучно переносят естественный отбор, пока среда не изменится. Это происходит потому, что естественный отбор удовлетворяется их сиюминутной серийностью. Столь глубокая подоплека объясняет, почему физический, биологический и социальный миры подчиняются единым в своей основе законам.
Возможно, сфера их выполнения простирается еще дальше. Причинно-следственный закон возник вместе со Вселенной. Поэтому он мог столкнуться с наследием гипотетического досингулярного Мироздания.
Мы предположительно рассматривали сингулярность как результат столкновения доменов умозрительного Вечно Изменяющегося Мира (см. разд. 1.7). Один из этих доменов характеризовался бесконечной дробностью, а другой – неограниченными скоростями взаимодействий.
Свойства этих доменов могли достаться начальной Вселенной и тут же подпасть под селективное действие причинно-следственного закона. Такие крайности, как бесконечная дробность материи и неограниченные скорости ее взаимодействия, являются исключительными, беспрецедентными свойствами, которые ни в чем не найдут аналога, т. е. окажутся неповторимыми. Закон каузальности не мог этого принять. Поэтому от родительских доменов в нашей Вселенной утвердились средние нормы дробности материи и скоростей ее взаимодействия.
Средняя дробность в нашем мире претворилась в виде кванта действия (постоянной Планка) ħ
= h
/(2π). Средняя скорость взаимодействий отпечаталась на свойствах Вселенной как фиксированная скорость света c
. Поперечник контакта родительских доменов (наша сингулярность) дал предельную Планковскую длину l
Pl
. Первое же фундаментальное физическое событие – обособление гравитационной силы – обзавелось своей гравитационной постоянной G, которая определяется из формулы, связывающей три более ранние постоянные (G
= l
Pl
2c
3/ħ
). Иными словами, гипотеза происхождения нашей Вселенной из Вечно Изменяющегося Мира способна объяснить связь наших фундаментальных физических констант при условии, что их размерности определились под давлением причинно-следственного закона.
В этом случае он отсек неповторимые физические крайности точно так же, как отсекал их в микро– и макромире, создавая современную картину физической действительности. Аналогичным образом был разработан биологический естественный отбор и создана направленность земной биоэволюции. То же самое произошло с миром предков человека, сообщив ему определенный, закономерный ход истории с неотенией, акселерацией, технологическими и культурными революциями. Возможность увязать фундаментальные законы природы и общества в зависимость, основанную на действии причинно-следственного закона, представляется нам косвенным признаком небеспочвенности понимания их генезиса, представленного в нашей работе.
Конечно, мировоззренчески для человека существеннее другое: происхождение и природа нашего общества. Изложенная версия развития живой природы и социума наглядно демонстрирует цепочку причинно-следственной связи, выводящей человеческую цивилизацию из недр животного царства, а закономерности жизни последнего – из динамики неживой природы.
В самом деле, геологический суперконтинентальный цикл управляет земным климатом, колеблющимся между крайностями оледенений и межледниковий. Эти климатические состояния регулируют уровень биопродуктивности экосреды, а от нее зависит живое «лицо» планеты: мобильные или маломобильные организмы как преобладающие формы биосферы. В одно из потеплений на эволюционной сцене закономерно появились наши двуногие предки гоминины. Предавшись неотеничному развитию, они попали на конвейер демографического роста и экологических угроз, попытались спастись хищничеством, но остановились на коллективном применении орудий, невиданном в животном мире. Коллективные орудия зажили собственной жизнью и потянули человечество за собой по пути технологического прогресса. Его результатом стало опасное для социальности праздное время, которое пришлось заполнять непроизводственным общением, положившим начало духовной жизни. Технический прогресс и общественное разделение труда подвергли испытанию единство общества, и спасением послужил цивилизованный образ жизни, пришедший к нам закономерно.
Таким образом, от цивилизации к суперконтинентальному циклу действительно тянется нить причинности. Ее дальнейшие зримые истоки пока не просматриваются, поскольку материалы по планетогенезу ограничиваются Землей и Солнечной системой. Понятно, что в основе развития земной природы лежит причинно-следственный закон. Однако мы пока не имеем сравнительного материала, позволяющего выявить закономерности формирования планет земной группы и зарождения жизни на их поверхности. Поэтому любая современная картина мира окажется далека от законченности.
Приложение 1
Палеолитическое искусство
Наши представления о духовном мире палеолитического человека в значительной степени основываются на истолковании созданных им памятников изобразительного искусства. Долгое время их интерпретация оставалась бессистемной. Сталкиваясь с палеолитическими изображениями тучных женщин, ученые усматривали в них свидетельства культа плодородия. Изображения животных связывались с тотемизмом, а стреловидные изображения на их корпусах – с магическими обрядами [81; 171; 209]. Предполагалось, что полнота женщин и животных в изобразительном творчестве отражала состояние беременности и, соответственно, надежды на плодородие природы и прекрасной половины первобытных коллективов. При этом исследователи игнорировали тот факт, что в раннем стиле I и в позднем стиле IV (о них см. ниже) женщины изображались исключительно грацильными, стройными, а в стиле IV так же воплощались и животные. Может быть, на заре и в конце верхнепалеолитического искусства культ плодородия дремал? Это предположение – очевидная натяжка.
Неудивительно, что, как позже выяснилось усилиями А. Леруа-Гурана, признаки «плодородия» женских фигур и корпусов животных обусловлены каноном художественного стиля. Далее, анималистические (зооморфные) сюжеты верхнепалеолитического искусства отражали очень ограниченный и стабильный список животных, который в силу своей узости и одинаковой представленности у всех верхнепалеолитических общин Франции, Испании и Италии (а также Германии и России), по определению, не мог иметь отношения к тотемизму, иначе получилась бы невозможная картина, будто все первобытные общины названных стран имели одни и те же тотемы (символически родственных себе животных, которые на деле призваны отличать сообщества друг от друга, а не смешивать их между собой). И наконец, стреловидные изображения являлись элементом очень четкой знаковой системы, не связанной с магией. Революцию в интерпретации палеолитического искусства произвел французский археолог А. Леруа-Гуран [497, с. 77–152; 498].
Исследуя точно датированные памятники малых форм (так называемое мобильное, портативное, или переносное искусство) и некоторые археологически датированные монументальные памятники (наскальное искусство пещер и скальных убежищ), А. Леруа-Гуран обнаружил, что художественные каноны различных эпох заметно различались, и это наблюдение позволило обобщить характерные художественные приемы выражения сюжетов изобразительного искусства в представление о четырех последовательных верхнепалеолитических стилях, классически представленных во франко-кантабрийском регионе (Юго-Западная Франция и Северо-Западная Испания), но фактически распространенных гораздо шире в верхнепалеолитической Евразии (хронологию франко-кантабрийских стилей см. Приложение 4).
В древнейшем стиле I
фигуры животных предельно схематичны и с трудом поддаются отождествлению (рис. 11). Часто, но далеко не всегда передавались только головы. Этот изобразительный канон близок современному примитивизму: округлая, эллиптическая или более угловатая продолговатая фигура, символизирующая голову, изредка дополнялась геометрически начертанным туловищем, непропорционально большим по сравнению с головой и снабженным линейными конечностями. Стиль I является преимущественно геометрическим, т. е. символически представляющим свои денотаты (изображаемые модели). Немногочисленные детали (глаз, пасть, уши, рога) изображались также геометрически и не отражали точный облик соответствующих деталей реальных животных. Искусство стиля I было скорее понятийным, символическим, нежели изобразительным, однако его дальнейшая судьба показывает, что такой символизм являлся следствием низкого художественного мастерства, характерного для древнейшей стадии искусства.
В следующем стиле II
изобразительный канон существенно меняется (рис. 12). Основу силуэта животного образует извилистая шейно-спинная линия, обычно легко ассоциирующаяся с силуэтом заявляемого животного. Прочие части тела подвешиваются к шейно-спинной линии, что предопределяет некоторые характерные способы их передачи: «утиная голова», сильно провисшее брюхо и незаконченные конечности лошади, подковообразная линия брюха-конечностей мамонта и т. п. Детали (глаз, ухо, рога, хвост, шерсть на брюхе мамонта, на загривке бизона) редки и переданы весьма обобщенно. Примитивность изображений напоминает еще стиль I, однако говорить о геометрическом символизме уже нельзя: силуэты животных обычно ясно отличаются друг от друга и напоминают общие контуры денотатов (изображаемых объектов). Построение образа на основе шейно-спинной линии приводит к тому, что, как и в стиле I, корпуса животных оказываются непропорционально велики по сравнению с головами. Для женских изображений этот художественный принцип оборачивается тем, что женский торс запечатлевается очень массивным по сравнению с головой и конечностями. Отсюда происходят так называемые признаки плодородия у женских фигур и признаки беременности у фигур животных. Как видим, эти приметы культа плодородия имеют сугубо стилистическое происхождение и в действительности не отражают никаких сакральных представлений. В стилях I и II преобладающей техникой исполнения является гравюра (иногда раскрашенная), хотя встречаются красочные изображения знаков (рук, абстрактных символов) и изредка фигур животных.
Рис. 11.
Стиль I
. Изображение на каменном обломке со свода абри де Ла Ферраси во Франции, созданное в эпоху археологической культуры ориньяк I. Предположительный смысл произведения таков. «Лев-страж Преисподней – март месяц. Носорог-надзиратель Преисподней (передняя часть пещерного льва + двойной зигзаг. Голова шерстистого носорога)». Речь могла идти о времени украшения святилища или о месяце проведения ритуала, связанного с франко-кантабрийской мифологией [209, с. 307, рис. 558]
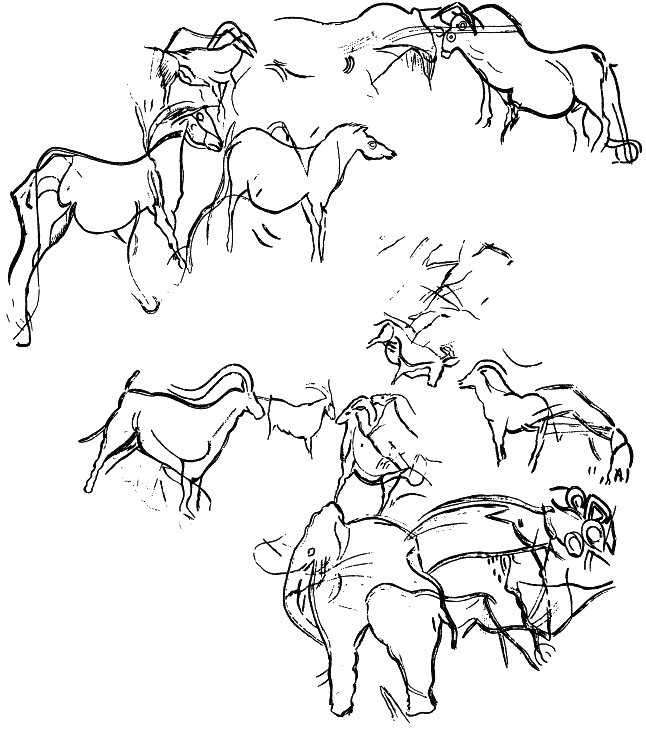
Рис. 12.
Стиль II
. Три наскальных панно во французском гроте де Пер-нон-Пер. Предположительный смысл изображений таков (слева направо). Представлена основная композиция франко-кантабрийского искусства: «2-е мая (вульва + двойная черта), Конь-Солнце (две лошади), Бык-Земля, Горный козел-дневное Небо, Пещерный медведь-проводник в Преисподнюю, Мамонт-ночное Небо, а также Горные козлы-дневное Небо, Большерогий олень-утреннее и вечернее Небо и другие мотивы». Запечатленный здесь космогонический миф о возникновении Солнца, Земли, Неба и Подземного мира известен в Древнем Египте, Древней Индии, у российских народов Севера и др. народов [209, с. 324–325, рис. 382, 383; 488, с. 260, рис. 7–9]
В стиле III
пропорции животных и женских фигур отвечают канону стиля II, однако построение фигур на основе шейно-спинной линии перестает быть господствующим принципом. Все части тела животного передаются равноценно, приблизительно отражая анатомические реалии, однако общий вид животного в стиле III древнем
весьма непропорционален: огромные корпуса снабжаются мелкими головами и конечностями (рис. 13). В стиле III позднем
анатомические пропорции начинают приближаться к реальным (рис. 14). Детали (глаз, уши, рога, хвост, копыта) встречаются часто. Обобщенный, синтетический образ стиля II начинает превращение в аналитический, а взятые в отдельности корпуса, головы и конечности животных гораздо лучше передают натуру, однако их конгломерат по-прежнему искажает анатомию денотата.
В стиле IV древнем
фигуры животных приобретают реалистические пропорции, а детали становятся многочисленными и точными, хотя имеются пережитки прежних стилей (например, контур брюшной области в виде М, идущий еще из стиля II, массивность крупов животных) (рис. 15). Общая тенденция реалистической передачи натуры распространилась и на антропоморфные изображения. Мужчины во всех стилях, как правило, передавались схематично, хотя имелись исключения в Лоссель (стиль II) и в гроте де Ла Марш (Люссак-ле-Шато, Вьенна, Франция, мадлен IIIa, стиль IV древний). Женские фигуры стиля IV становятся грацильными и лишенными, в отличие от предшествующих стилей, признаков «плодородия», хотя и здесь имеются исключения: в Ла Марш женские фигуры массивны, а женский барельеф из абри Пато (Лез Эйзи-де-Тейяк, Дордонь, Франция, слой III, перигор VI, 23 010 ± 170 лет назад, стиль II) грацилен. Схематичные женские фигуры стиля IV послужили источником для знаков в виде клавиформ (музыкальных ключей), весьма характерных для данного периода (заметим, что знаки клавиформ усматриваются в стиле II гротов Байоль № 17, № 22 и де Ла Бом-Латрон № 18).
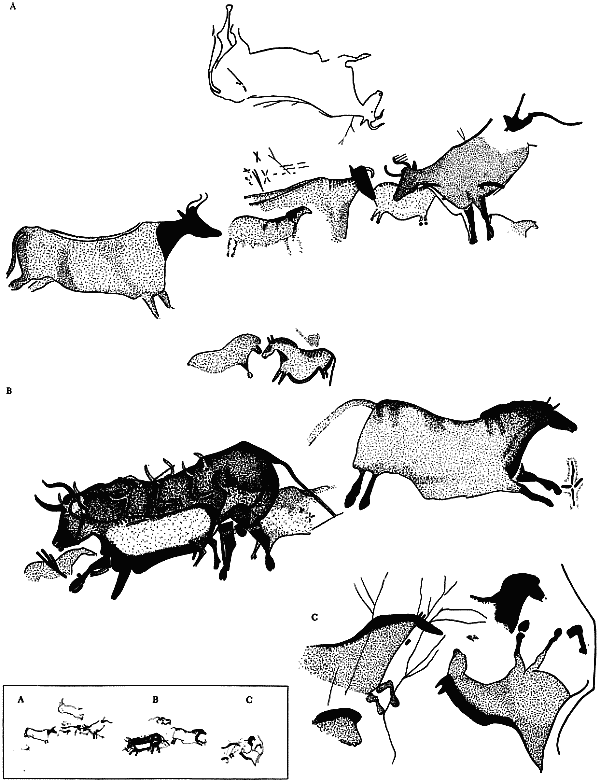
Рис. 13.
Стиль III древний
. Такими мы видим во французском гроте Ласко-1 лошадей, быков, ветвистые знаки, крест и пунктуации, которые предположительно значили «Конь-Солнце, Бык-Земля, 4 апреля, 3 ноября» (даты создания фигур или ритуалов в их честь). Огромные крупы животных, снабженные маленькими головами, являются отличительными признаками данного стиля. [488, с. 184]
Стиль IV поздний
показывает сложную картину эволюции художественного творчества. Сначала тенденции стиля IV древнего приводят к фотографическому реализму (Ла Мэри [488, с. 234–235, рис. 3], Леванцо [209, с. 400, рис. 524]) (рис. 16), а затем в связи с упадком палеолитического искусства наступает схематизация образов (Ла Рок; начало процесса отмечено в Аддаура I, где натуралистические изображения сосуществуют со схематическими). В количественном отношении вершина франко-кантабрийского искусства была достигнута в мадлене IV, однако в стилистическом, качественном отношении расцвет пришелся на мадлен V, сменившись стремительным закатом в мадлене VI. Следует отметить, что и здесь мы наблюдаем типичный для нашего мира ход незаурядного развития явления (в данном случае – верхнепалеолитического искусства), его расцвета, крайней специализации (в виде фотографического реализма) и катастрофического заката со сменой обстановки (с наступлением кризиса ледниковой мамонтовой мегафауны в связи с началом голоценового потепления, поскольку верхнепалеолитическое искусство зиждилось на этой фауне). Аналогичным образом угасли динозавры (см. разд. 2.3).

Рис. 14.
Стиль III поздний
. «Китайские лошади» грота де Ласко-2 во Франции – лучшие фигуры этого стиля. Три лошади – (здесь одна из них) добавлены к группе больших коров и большому благородному оленю более древнего стиля святилища Ласко-1, так что в сумме образовалась стандартная композиция лошадь – бык – олень [498, рис. 317]

Рис. 15.
Стиль IV древний
. Основная композиция во французском гроте де Труа-Фрер-2 («Трех Братьев») выполнена методом суперпозиции, когда изображения наслаиваются друг на друга, исключая полноценное эстетическое восприятие в нашем понимании. Мы видим рисованную передачу космогонического мифа о рождении Коня-Солнца – Бизона-Земли – Северного оленя-утреннего и вечернего Неба, Горного козла-дневного Неба – Медведя-проводника в Преисподнюю. Панно «читалось», а не созерцалось [488, с. 402, рис. 6]
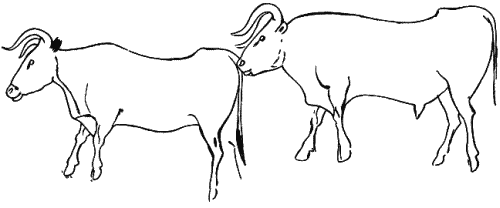
Рис. 16.
Стиль IV поздний
. Фотографически точно выполненный силуэт Быка-Земли следует за Коровой во французском гроте де Ла Мэри («пещере Мэрии»). Совершенство рисунка не уступает реалистической графике современности. Мифологическая иконография отвечает повседневным впечатлениям первобытных охотников, в чем они предвосхитили манеру художников Ренессанса [209, с. 315, рис. 375]; [488, с. 235, рис. 3]
Концепция названных художественных стилей привела к важным семантическим, смысловым открытиям в монументальном искусстве. До А. Леруа-Гурана наскальные пещерные изображения рассматривались как свидетельства различных эпох на том основании, что гравированные и живописные фигуры зачастую налегали друг на друга, образовывали своего рода наслоения вроде археологических и наводили археологов – прежде всего знаменитого аббата А. Э. П. Брейля (Брёйя, 1877–1961) – на естественные предположения, что нижние фигуры сделаны раньше, а верхние фигуры моложе. Соответственно, их датировка основывалась главным образом на археологическом методе суперпозиции (методе взаимоналоженных изображений). Если одна фигура покрывала другую, констатировалась их значительная эпохальная разновременность. А. Брейль создал целую концептуальную хронологию наскального искусства, основывающуюся на принципе суперпозиции, на особенностях техники исполнения (гравюра, живопись и их особенности) и очень мало – на датированных мобильных аналогиях. Поскольку в суперпозиции практически всегда (за редкими исключениями, например в Гаргас I) оказывались фигуры одного и того же стиля, благодаря усилиям А. Леруа-Гурана стало ясно, что принцип суперпозиции не может использоваться для крупномасштабной общей периодизации наскального искусства. Другой вывод говорил о том, что фигуры одного стиля в пещерных святилищах составляли целостные ансамбли, структура которых может многое сказать о представлениях древних художников.
Структура идеального святилища такова [497, с. 95–101; 498, с. 114–118, 441 рис. 763; 500, с. 76]. В его центре на главном панно находится основная композиция, состоящая из лошади («первое животное»), бизона или быка-тура («второе животное») и «третьего животного» (горный козел, благороднай олень, лань, мамонт, северный олень). У входа чаще всего присутствует «третье животное» (обычно благородный олень), которое встречается и в глубине. В этой связи вспомним, что в ближневосточной пещере Кафзех подростка-протокроманьонца сопровождал на «тот свет» череп лани с рогами, которая, по-видимому, каким-то образом отвечала за вхождение в потусторонний мир (см. разд. 3.2). В глубине – обычно «четвертое животное» (лев, медведь, носорог, а также более редкие млекопитающие, рыбы, птицы) и антропоморфы (человеческие изображения). Иногда эта структура стягивается в одно обширное панно (Альтамира-3, Ла Пенья-де-Кандамо-3, Руффиньяк и др.), на котором диспозиция животных сохраняется: в центре царят животные I, II, III, а на периферии – IV и антропоморфы. Изредка животные класса IV выступают в роли «третьего животного»: лев (в Ле Пеш-Мерль-1), мегацерос (в Рукадур, хотя в этом случае мы имеем дело пусть и с гигантским, но оленем, т. е. с «третьим животным»). Конечно, схема идеального святилища выдерживается в зависимости от топографии пещер, однако ее существование доказывается статистикой взаимного расположения животных. По всему пещерному святилищу также встречаются различные знаки, для которых А. Леруа-Гуран разработал свои закономерности распределения. Их принятие связано с трудностями, о которых речь ниже.
А. Леруа-Гуран проанализировал статистику 63 украшенных гротов из 110 известных в его время (1964 г.), а также большое число мобильных памятников [497, с. 91, 95]; [498, с. 444–445]. Из этой статистики следовала определенная частотная иерархия сюжетов, отраженная в понятиях животных I – IV. Мы произвели статистическую обработку 222 монументальных памятников франко-кантабрийских стилей из Франции, Испании, Италии и России (Капова пещера), результаты которой приведены в Приложении 2 (перечень и хронологию памятников см. в Приложении 4, где порядковые номера 5 новых добавленных памятников подчеркнуты; они специально не анализировались, однако их сюжеты идентичны прочим пещерным святилищам). Наш анализ полностью подтверждает основные положения А. Леруа-Гурана, лишь несколько уточняя их количественно и содержательно. Расчеты, сделанные нами, относят количество памятников, где встречается тот или иной сюжет, к общему количеству учтенных памятников. Первый столбец Приложения 2 отражает верхнепалеолитическое монументальное искусство франко-кантабрийских стилей, второй – мобильное искусство древнего и среднего палеолита, перечень и хронология памятников которого приведены в Приложении 3.
Мобильные памятники, по А. Леруа-Гурану [497, с. 116–122], образовывали портативную (переносную) параллель монументальным святилищам. Однако в портативных святилищах трудно установить характер первоначального ансамбля и степень фрагментарности дошедшего до нас материала, который иногда может быть огромным (например, более 800 гравированных пластинок в гроте д’Анлен, локализация и возраст которого те же, что и у Ле Труа-Фрер-2, см. Приложение 4). Поэтому монументальные святилища всегда доказательнее.
Франко-кантабрийские бестиарии (наборы животных) состояли из 39 определенных видов млекопитающих, птиц и рыб, а также из неопределенных змей (собственно, лучше говорить о 37 видах, поскольку № 28–29, барсук и бобр вызывают сомнения). Основу этих бестиариев составляли животные с частотами 66,2–9,5% (лошадь, бизон, бык, горный козел, благородный олень, лань, мамонт, северный олень, пещерный лев, медведь, шерстистый носорог). Среди них лидируют лошадь (66,2%) и бизон с быком (совместная частота 66,2%), что полностью подтверждает квалификацию их А. Леруа-Гураном как «первого» и «второго животного» основной композиции. Животные с частотами 44,1–16,7% (горный козел, благородный олень, лань, мамонт, северный олень) являются обычным «третьим животным» основной триады, а животные с частотами 17,1–9,5% – стандартными персонажами глубин святилищ и периферий основного панно (пещерный лев, медведь, шерстистый носорог). Таким образом, основу франко-кантабрийского бестиария составляли животные, выступающие в ансамблях монументального искусства в четырех статистико-топографических ролях. Проще говоря, чем чаще изображалось франко-кантабрийское животное, тем центральнее была его роль в наскальных композициях, что убеждает в справедливости их понимания А. Леруа-Гураном.
Следует добавить, что классическое франко-кантабрийское искусство, просуществовавшее 20 тыс. лет, всегда сохраняло абсолютное статистико-топографическое единообразие. Правда, святилища стиля I полностью разрушены, однако сохранились некоторые фрагменты каменных плиток, отслоившихся со стен и запечатлевших былое великолепие древнейших украшенных убежищ в виде фигур их бестиариев. Они показывают тождество с бестиариями стиля II, в котором основная композиция в законченном виде появляется начиная с Пер-нон-Пер. Самая поздняя основная композиция отмечена в Дриасе III (Аддаура I). Следовательно, в течение 20 тыс. лет западноевропейские художники изображали один и тот же ведущий сюжет: основную композицию, состоящую из лошади, бизона (или быка) и горного козла (или других животных класса III). На периферии этой композиции размещались животные класса IV и антропоморфы. Эти объективные статистико-топографические факты позволили А. Леруа-Гурану сделать вывод о том, что единственной идеологической формой, способной отразиться в столь стабильной иконографической системе, могла быть только мифология [497, с. 151–152]. Франко-кантабрийское искусство, таким образом, являлось религиозным по основному своему жанру. При этом декоративное мобильное искусство (резные копьеметалки, гравированные «жезлы начальников», каменные лампады с гравированными изображениями, снабженные гравюрами костяные стержни и мамонтовые бивни неясного назначения, костяные ложки, лопатки, кружки и просверленные диски, подвески, просверленные пластинки и другие украшенные предметы) – все это обыденное эстетическое богатство неизменно облагораживалось фигурами священных животных и сопутствующими им знаками, т. е. в духовном отношении всецело оставалось в сакральной сфере.
Конкретное содержание франко-кантабрийской мифологии остается загадкой. Основываясь на серии фигур из Ле Пеш-Мерль-2, демонстрирующей последовательное превращение женщины в бизона, и на том факте, что одна из скульптурных «Венер» местонахождения Лоссель держала в руках бизоний рог, А. Леруа-Гуран предположил, что «второе животное» (бизон) было символом женского начала, а все остальные животные – символами мужского. Подобное предположение противоречит фактам.
Мужчины с чертами бизонов (мужчины-бизоны) встречаются чаще (примеры имеются в Ле Габийю-1, Ле Габийю-2, Ла Пасьега С-1, Ле Труа-Фрер-2). Лоссельская «Венера», держащая рог бизона с 13 параллельными насечками, проще всего объясняется как символ лунного месяца с количественным пояснением к нему и обнаруживает ближайшую аналогию с клавиформой, сопровождаемой серией пунктуаций (Нио и т. п.). Вообще принцип интерпретации топографических классов животных при помощи категории пола плохо подкрепляется реалиями. Так, женщины-птицы (Альтамира-1, Ле Пеш-Мерль-2) соединяют в себе женское начало с «мужским», поскольку птицы в системе А. Леруа-Гурана относятся к мужскому классу. Бизоны, как отмечалось, могут быть и «мужскими», и «женскими» животными. «Мужскими» животными можно признать северного оленя и льва в силу того, что мужчина-северный олень известен в Ле Труа-Фрер-2, а мужчина-лев представлен в Ле Рок-о-Сорсье. Однако в свете сказанного разумнее воздержаться от сексуальной интерпретации классов франко-кантабрийских бестиариев.
Из статистики франко-кантабрийского бестиария вытекает только то, что лошадь символизировала верховное «божество», бизон-бык – «божество» второго плана, а «третье» и «четвертое животные» соотносились с «божествами», соответственно, третьего и четвертого планов. По ряду признаков франко-кантабрийская мифология отражала некую общечеловеческую мифологическую идеологему, и перспективным представляется поиск ее рефлексов (отражений) среди основных мифологем Старого Света, древнейшие из которых соотносились с мифами о Солнце, Земле и Небе. В этой связи заслуживает внимания интерпретация сюжетов верхнепалеолитического искусства отечественным археологом В. Е. Ларичевым [60].
На верхнепалеолитическом поселении Малая Сыя (Хакасия, Красноярский край, Россия, 34 500 ± 450, 33 060 ± 450, 33 060 ± 300 лет назад по 14С) среди произведений мобильного искусства В. Е. Ларичев открыл изображения композиций мамонт-бизон и мамонт-черепаха. Мамонт, стоящий на черепахе, по этнографическим параллелям интерпретирован Ларичевым как символ Неба, а черепаха (и замещающий ее в похожей композиции бизон) – как символ Земли. Кстати, подобная мифологема объясняет происхождение известных индуистских представлений о Вселенной как о слонах, стоящих на черепахе. Композиция бизон – мамонт (№ 9 в списке композиций А. Леруа-Гурана) представляет собой неполную триаду и, по нашим подсчетам, встречается в 10 гротах, в 6 из которых она является основной (Ле Шваль (Арси), Юшар, Капова, Ложери-От, Улан, Сен-Фрон). Эта композиция является частным случаем соединения «второго-третьего животных» без лошади, и такое соединение имеется в 26 гротах. Если предположить, что «второе животное» символизировало Землю, а «третье» – Небо, то франко-кантабрийская мифология предстанет как первая космогония.
Разумеется, эта первобытная космогония не являлась сциентистской, научно ориентированной по характеру мировоззрения. Мы вряд ли ошибемся, если допустим, что Мамонт-Небо, покрывающий Бизона-Землю или Черепаху-Землю, символизировал биологическое спаривание Неба и Земли, возможно в интересах плодородия последней. Подобное представление могло проистекать из наблюдений дождевого орошения Небом Земли, что вызывало урожайность последней. Натурализм первобытной мифологии является ее естественной особенностью.
Древнейшей аналогией, привлеченной В. Е. Ларичевым, послужила космогония Древнего Египта, старейшим вариантом которой в свою очередь, являлась космогония города Гелиополя [57, с. 82, 89]. Суть этой космогонии сводилась к тому, что из первобытных вод (Нун, Наун) возник Атум (Атама, «Всё», или «Совершенный»), который произвел пару Шу (Шау, «Воздух») и Тефнут (Тфини, «Влага»). От последних произошли Геб (Гиба, «Земля») и Нут (Ни, «Небо»). От Земли и Неба возникли Осирис (Усири) и Исида (Иси), а также Сет (Сута) и Нефтис (Нибтха) [57, с. 87–89; 46, с. 380]. При этом Осирис стал хтоническим (подземным) божеством и царем загробного мира [57, с. 89, 309].
В этой системе представлений имелись две модернизации. В «Текстах пирамид» (кон. V династии, фараон Уннас, – кон. VIII династии, т. е. 2423–2220 до н. э.), древнейшем источнике по религии Египта, Атум отождествляется с Ра (Риа, «Солнце») в ипостаси единого божества Ра-Атума (Риа-Атама, «Солнце-Всё»). «Тексты пирамид», судя по их враждебности к верхнеегипетским Осирису и Хору, а также по упоминаниям в них нижнеегипетского царя, восходят ко временам независимого нижнеегипетского царства, датирующегося изображением нижнеегипетской короны на сосуде амратской эпохи (она же Нагада I, 6600–6400 календарных, или 5744 ± 300–5577 ± 300 радиоуглеродных лет назад, или 4650–4450 до н. э.) и прекратившего существование с началом нулевой династии (3390 календарных, или 2700 радиоуглеродных лет до н. э.). После франко-кантабрийской эта мифология, таким образом, является древнейшей в мире.
С поправкой на «Тексты пирамид» гелиопольскую космогонию можно рассматривать следующим образом. Из первобытных вод (Наун) рождается Солнце-Всё (Риа-Атама), порождающее Воздух и Влагу (Шау и Тфини), а от последних происходят Земля и Небо (Гиба и Ни). Дальнейшая филиация (родословная) богов венчается рождением Усири, верховного божества Подземного мира.
Космогоническая линия Вода – Солнце – Земля – Небо – Подземный мир, в принципе, отвечает статистической иерархии франко-кантабрийских сюжетов Лошади (66,2%) – Бизона-Быка (66,2%) – Оленя (48,7%) – Пещерного льва (17,1%) как представителей классов первого, второго, третьего и четвертого животного. Лошадь можно отождествить с Солнцем, а Бизона-Быка – с Землей. С Небом совпадают Олень и остальные представители третьего класса животных: горный козел и мамонт. Судя по статистике (см. Приложение 2), олень символизировал утреннее и вечернее Небо, горный козел, 44,1%, – дневное Небо, а мамонт, 22,5%, – ночное Небо. Много позже представление о тождестве утреннего и вечернего Неба вылилось в открытие Пифагором (2-я пол. VI в. – нач. V в. до н. э.) тождества Вечерней и Утренней звезды (хотя мыслитель, вероятно, просто обнаружил принадлежность одному светилу вечернего и утреннего проявлений планеты Венеры) [104, с. 147, № 20].
С Преисподней логично связать Пещерного льва, медведя и шерстистого носорога. Последний не случайно наблюдает сцену убиения охотника бизоном в Колодце Ласко-3, где присутствует и птица. Медведь, в свою очередь, замечен в атаках на мужчин (как явствует из мобильных произведений гротов дю Пешиале, Дордонь, Франция, и дю Мас-д’Азиль, мадлен IV). Хтоническая же природа льва в первобытной мифологии согласуется с его загробной ролью в этрусской дисциплине (религии) и проясняет значение знаменитого египетского сфинкса-льва, стерегущего покой гробниц фараонов – величественных пирамид. Иконография IV животного наводит на мысль, что лев выступал своего рода стражем Преисподней, тогда как медведь служил проводником в загробный мир (но не в качестве мирного древнегреческого Харона, а скорее в роли агрессивного этрусского Хару, вооруженного смертоносным молотом). Носорог же выглядит надзирателем загробного царства. Его спутница-птица наводит на мысль, что фантом-полярная сова послужил прообразом кельтских призраков (душ умерших) вроде роковой супруги короля Артура Геневеры, чье имя значило «Белый призрак» (валлийск. Gwenhwyfar [25 a, с. 101, 253, пр. 2]). Гипотезе недостает лишь понимания смысла первобытных вод.
В наскальном франко-кантабрийском искусстве, помимо животных, антропоморфов и знаков, во всех стилях (кроме первого, святилища которого разрушены) имелся еще целый ряд изобразительных мотивов. К ним относятся совокупности кривых черт (19,8%), скобления (11,3%), макароны и меандры (9,9%), наброски пальцами (7,2%), хаотические черты (6,3%), пучки черт и линий (2,3%). Из этих беспорядочных изображений поддаются интерпретации макароны и меандры, представляющие собой серии волнистых (иногда прямых) линий, либо параллельных друг другу, либо нет. Архетипом подобного мотива служил мультиплицированный (повторяющийся) зигзаг.
Единственный памятник, где мотив зигзага находит точное объяснение, – это топографическая карта из Межирича (Канов, Черкассы, Украина, среднеднепровская культура, Вюрм IV Дриас I B, 15245 ± 1080 лет назад по 14С) [89, с. 135, 137 рис. 58; 29, с. 73], на которой параллельные зигзаги, покрывая очертания реки, символизировали воду. Представляется вполне вероятным, что панно макарон и меандров выражали то же самое.
В рамки подобных панно нередко включались фигуры животных, как бы рождающиеся из них. Так, в Ла Бом-Латрон из панно меандров № 5 возникала стандартная триада лошадь – бык – мамонт [298, с. 335–336]. Аналогичные примеры встречаются и в других гротах (Ардалес, Гаргас I-1, Ле Труа-Фрер-1, Ле Пеш-Мерль-2, Эрберюа и др.). Таким же окружением могли служить панно скоблений и хаотичных черт (Ла Рок). Можно предположить, что подобные панно отражали мифологическую линию рождения сюжетов из водного хаоса (меандров) или неопределенного хаоса (хаотичных черт).
Таким образом, древнеегипетская мифологема первобытные воды – Солнце – Земля – Небо – Преисподняя находит параллель во франко-кантабрийском изобразительном ряду панно меандров – первое – второе – третье – четвертое животные и позволяет ставить вопрос: не являлась ли франко-кантабрийская мифология древнейшей космогонией? Подобная мифологическая идеология могла бы содержать архетипы характерных для древнего мира культов: водного (божества морей, рек, источников), солярного (божество солнца – заметим, единственное во всех известных культах, а потому, вероятнее всего, восходящее к верховному божеству палеолита, Коню-Солнцу), хтонического (божества земли и подземного мира) и астрального (звездного, небесного с характерными божествами), а также соответствующих им первоэлементов (вода, огонь, земля, воздух). Излишне говорить, что учение философа Фалеса Милетского о воде как о первоэлементе восходит к древнеегипетским мифологическим представлениям (Фалес посещал Египет [104, с. 108, № 11]) и через них, вероятно, к описанной первобытной мифологии.
Соответствующее франко-кантабрийское идеологическое наследие, скорее всего, послужило одной из основ западноевропейской мегалитической культуры (культуры дольменов, гробниц из гигантских камней; менгиров, одиноко стоящих огромных камней – собственно, мегалитов; кромлехов, кольцевых сооружений из мегалитов типа знаменитого Стоунхенджа в Англии). Правда, мегалитическая культура начинается лишь в финальном мезолите, однако верхнепалеолитическое и мезолитическое население Западной Европы, по-видимому, было соплеменным и придерживалось одинаковых мифологических представлений. Мегалитическая культура примечательна своими астрономически выраженными лунно-солнечными культами, которые органично выводятся из духовного мира франко-кантабрийских художников, располагавших космогонией и лунным календарем.
Кроме бестиариев, франко-кантабрийское искусство располагало небольшим корпусом антропоморфных сюжетов, статистико-топографическое положение которых обычно связано с периферией основного панно или с глубокой периферией пещерного святилища в целом. Однако корпус антропоморфов не был однородным и распадался на две группы, различающиеся статистически, стилистически, композиционно, по тенденции к зооморфизации (к приобретению черт животных) и по отношению к системе знаков.
Встречаются изображения мужчин (14%), неопределенных антропоморфов (13,5%) и женщин (9,5%), причем неопределенные антропоморфы иконографически и статистически чужды женским изображениям; их можно рассматривать как крайнюю схематизацию мужского образа, к чему имелись существенные стилистические предпосылки. Наскальные женские фигуры подчинялись эволюционным стилистическим закономерностям, выражающимся в грацилизации (утончении) женского образа от стиля к стилю (II – IV). Напротив, мужской образ не подчинялся стилистическим закономерностям и во всех стилях был одинаково грацилен. Это объясняется тем, что, за вычетом копьеметателя из Лоссель, все мужские фигуры никогда не были реалистическими и всегда отличались большим или меньшим схематизмом. Стилистическая же эволюция затрагивала лишь натуралистические изображения женщин и животных, а потому не распространялась на мужские сюжеты. Именно преобладающая схематичность последних позволяет думать, что неопределенные антропоморфы являлись крайним случаем этой схематизации, т. е. относились скорее к мужским сюжетам, с которыми они имели идентичную частоту встречаемости.
Совместная частота неопределенных антропоморфов и мужчин достигала 22,5% (отдельные частоты при этом не складываются; учитываются памятники, где обнаруживаются неопределенные антропоморфы, мужские сюжеты или то и другое одновременно).
Таким образом, имелась мужская группа сюжетов (22,5%) и женская группа (9,5%). Они отличались статистически и, по-видимому, обладали различным смыслом. Поэтому сюжеты этих групп никогда не образовывали совместных композиций, если не считать случаев присутствия на одном панно, к которому стягивалась значительная часть сюжетов всего святилища (например в Масса). Мужские сюжеты могли составлять небольшие серии из похожих антропоморфов (Альтамира-3, Ле Комбарель I, Ла Пенья-де-Кандао-3, Руффиньяк); то же справедливо и для женских сюжетов (Ле Комбарель I, Лоссель, Ла Магделен, Ле Рок-о-Сорсье).
Таким образом, статистическая и стилистическая чуждость мужских и женских сюжетов дополняется топографической. Все это не подтверждает сексуальную концепцию А. Леруа-Гурана: представители разных полов явно не служили дополнением друг другу, а играли различные роли в наскальном искусстве.
Различна была и тенденция к зооморфизации мужских и женских образов: у мужских она иконографически и статистически несомненна (5,9%), у женских она статистически нерепрезентативна (0,9%). Имеются изображения мужчин-бизонов, что, как отмечалось, исключает женскую трактовку данного животного, которое вместе с быком символизирует женский пол у А. Леруа-Гурана, тогда как прочие животные отнесены к мужскому. Встречаются мужчины с рогами бизонов (Ле Габийю-1, Ле Габийю-2, Ла Пасьега С-1, Ле Труа-Фрер-2), а также мужчина с «хвостом бизона» вместо головы (Пергусе). Сюда же следует отнести антропоморфов с головами черепах из Лос Касарес, поскольку, по В. Е. Ларичеву, черепаха является композиционным эквивалентом бизона в паре бизон – мамонт, черепаха – мамонт. Однако черепаха – редчайшее животное во франко-кантабрийском искусстве (имеются мобильные изображения в абри дю Рок-де-Сен-Сирк, Сен-Сирк-дю-Бюг, Дордонь, Франция, и в Марсула), так что истолкование этого сюжета лишено статистики. Предположительно, мужчины-бизоны и мужчины-черепахи были причастны к хтоническому культу Земли (колдуны соответствующей специализации).
Можно также упомянуть еще мужчину-мамонта (Ле Комбарель I) и мужчину-северного оленя (Ле Труа-Фрер-2), в которых следовало бы видеть колдунов Неба. Упомянем мужчину-льва (Ле Рок-о-Сорсье, Ла Пенья-де-Кандамо-3, Лос Орнос-де-ла-Пенья-2 и, возможно, Альтамира-3, где три звероподобных антропоморфа изображены в «молитвенной» позе, как мужчины-львы в Ла Пенья-де-Кандамо-3 и Лос Орнос-де-ла-Пенья-2) и добавим мужчину-медведя (Комарк, Ле Комбарель I, Сен-Сирк-2). Они, должно быть, представляли колдунов Преисподней.
Присовокупим мужчину-птицу (Ле Пеш-Мерль-2, Ле Комбарель I). Последние, вероятно, тоже изображали колдунов Преисподней (поскольку птица присутствует в сцене гибели охотника от рогов бизона в Ласко-3; кроме того, сопоставляя женщин-бизонов из Ле Пеш-Мерль-2 и женщин-птиц из Альтамиры-1 и Ле Пеш-Мерль-2, мы вправе допустить, что птицы подобно бизонам являлись божествами, связанными с землей: если бизоны отвечали поднебесной Земле, то птицы, судя по всему, выражали ее недра, отчего их связь с ночной Луной кажется естественной, ибо ночные божества сродни подземным). Таким образом, мужские сюжеты были родственны животным II, III и IV. В женской группе имеются женщины-бизоны (Ле Пеш-Мерль-2) и женщины-птицы (Альтамира-1, Ле Пеш-Мерль-2), откуда можно лишь заключить, что сексуальная трактовка бестиария неприемлема.
В отличие от женских, мужские образы могли участвовать в сценах, которые в наскальном искусстве сводились к одной-двум фабулам. Во-первых, это конфликт мужчины с бизоном с последующим поражением мужчины (Ле Рок-де-Сер, где бизона замещает овцебык; Виллар, Ласко-3, Сен-Сирк-2, Су-Гран-Лак-2, Эль Пиндаль; в последнем случае фигурки двух поверженных бизоном мужчин [93, с. 252] можно трактовать и как схематическое изображение лошадиной головы). Возможно, сюда же следует отнести и фигуру копьеметателя из Лоссель, поскольку наиболее вероятной целью его атаки мог быть бизон (в мобильном искусстве, кроме конфликта мужчины с бизоном из Ложери-Бас, известны и конфликты мужчин с медведем: грот дю Пешиале, Дордонь, Франция, и Ле Мас-д’Азиль, мадлен IV).
Далее, в Су-Гран-Лак-2 мужчина, атакуемый рогом бизона (очень близкий к мужчине, атакуемому бизоном в Сен-Сирк-2), одновременно поражается тройкой стрел. В этом качестве он сближается с ранеными мужчинами из Куньяк и Ле Пеш-Мерль-2, пораженными дротиками.
Возможно, сюжеты мужчин, атакуемых бизоном, и раненых мужчин являются частью одного мифа. Следовательно, мужской образ являлся персонажем мифологии, чего нельзя сказать о женском.
Встает, конечно, вопрос о назначении женского образа, и здесь обнаруживается еще одно отличие мужских и женских персонажей. Мужские сюжеты франко-кантабрийского искусства никогда не выступали элементом знаковой системы, в то время как женский образ в стиле IV получил даже особое знаковое воплощение в виде клавиформы (знака музыкального ключа, схематического женского силуэта; имеются указания, что клавиформы появляются уже в стиле II гротов Байоль и Ла Бом-Латрон). Кроме того, женское изображение могло быть составной частью парного знака (например в Комарк), о значении которого речь пойдет ниже.
Высказанные наблюдения позволяют сделать вывод о том, что мужские сюжеты являлись элементом образной системы, выражающей суть франко-кантабрийской мифологии. Женские сюжеты, напротив, выступали преимущественно элементом знаковой системы, сопровождающей мифологическую иконографию.
Семантика мужских изображений составляет проблему. Как мы видели, во всех второстепенных классах животных (II – IV) имелись антропоморфные представители. И выглядит нелогичным, что подобный представитель не выступал от имени «первого животного» (лошади), хотя его ведущая роль в бестиарии, казалось бы, предполагала и максимальное антропоморфное представительство: проще говоря, непонятно, как это Конь-Солнце не располагал собственным колдуном.
Отсюда возникает предположение, что антропоморфы, не отмеченные чертами зооморфной специализации, возможно, представляли «первое животное». Это предположение проливает определенный свет на смысл сцены с мужчиной, поражаемым бизоном. Если незооморфный мужчина представлял «первое животное», а оно отождествлялось с Солнцем, то его конфликт с бизоном, символизирующим Землю, отражал некий миф, связанный с солнечным закатом или сменой сезона высокого стояния Солнца сезоном низкого, подчиненного стояния светила над горизонтом.
Типологически такой миф мог быть архетипом стандартных мифов древнего мира об умирающей и воскресающей природе. В Древнем Египте это был миф о знаменитом Осирисе (Усири), а в Шумере – миф о Думузи. Согласно шумерским представлениям, Солнце (Уту) уходило в преисподнюю еженощно, а Луна (Наннар) – раз в месяц, в период новолуния, когда Луны не видно. Если конфликт мужчины с бизоном выражал первое из этих представлений, то превращение бизона (Земли) в женщину-птицу (Луну) в Ле Пеш-Мерль-2, очевидно, передавало второе (поглощение Луны Землей; поясним, что сюжет в Ле Пеш-Мерль-2 можно понимать и как превращение женщины-птицы-Луны в женщину-бизона-Землю, т. е. как вероятное поглощение Луны Землей). Это допущение подкрепляется тождеством женского образа в знаковой графике с символикой Луны, точнее лунного месяца.
Революция в понимании франко-кантабрийской знаковой системы связана с работами А. Леруа-Гурана, А. Маршака [535] и Б. А. Фролова [108]. А. Леруа-Гуран, следуя своему принципу поиска в сюжетах франко-кантабрийского искусства признаков противопоставления полов (чего примитивные общества, судя по данным этнографии, не знали), разделил знаковую систему верхнепалеолитического искусства на два больших класса: тонкие (мужские) и широкие (женские) знаки [499, с. 302–303]. К первому классу он отнес фаллические изображения, одинарные и двойные палочки, скалиформы (знаки в виде «лестницы»), крюки, пенниформы («перообразные» знаки), ветвистые и зубчатые знаки (группа 1), а также пунктуации (ряды точек, группа 3) и ряды параллельных палочек (группа 4). Весь этот знаковый класс, по А. Леруа-Гурану, являлся стилизацией фаллического образа. Ко второму классу отнесены вульвы, стрелы, углы, треугольники, полуовалы, овалы, кресты, прямоугольные знаки, акколады (фигурные скобки), клавиформы (стилизованные женские силуэты) и т. п. Все эти знаки группы 2 возводились А. Леруа-Гураном к стилизациям вульв или женских профилей [497, с. 92, рис. 7; 498, с. 453, рис. 780, с. 454, рис. 781].
Следует признать, что треугольники, углы, полуовалы и овалы действительно могли быть стилизацией вульвы, а клавиформы – стилизацией женского профиля. Однако обнаружение нами доверхнепалеолитической знаковой системы франко-кантабрийского типа (см. далее) показывает, что основные типы знаков существовали задолго до верхнего палеолита, и, следовательно, строить историческую генеалогию стилизаций вульв и фаллосов в рамках материала верхнепалеолитического искусства (как это делал А. Леруа-Гуран) нельзя, поскольку эти мнимые «стилизации» родились до верхнего палеолита, когда изобразительных архетипов для них вообще не существовало. Проще говоря, основные группы знаков родились в нижнем – среднем палеолите сами по себе и при этом не являлись обязательно стилизацией признаков пола. Последние, конечно, были представлены в знаковой системе, но крайне ограниченно, сообразно своему месту в психике нормальных, работящих людей, какими были наши предки, не покладая рук возводившие фундамент здания материальной и духовной культуры, в котором мы пребываем ныне.
Гораздо важнее другое открытие А. Леруа-Гурана, состоящее в понятии парных знаков. По А. Леруа-Гурану, они представляли собой комбинацию знаков группы 2 (женское начало) со знаками групп 1, 3, 4 (мужское начало) [498, с. 454, рис. 782].
Как мы видели на примерах зооморфных антропоморфов, во франко-кантабрийском бестиарии принцип противопоставления полов не выдерживался. Основные типы знаков, появившись в законченном виде в нижнем и среднем палеолите, также исключают свое тотальное возведение к паре сексуальных символов. Действительная картина была не столь упрощенной. Б. А. Фролов показал, что пунктуации и ряды палочек не символизировали фаллический образ, а являлись хорошо выраженными счетными знаками (точка или палочка = 1, две точки или палочки = 2 и т. д.). С другой стороны, А. Маршак, проанализировав ряд мобильных памятников верхнего палеолита (например, из абри Бланшар-де-Рош I, ориньяк I; грот дю Таи, Сен-Назер-ан-Руайан, Дром, Франция, верхний мадлен), установил, что верхнепалеолитический человек вел регистрацию лунных фаз и, следовательно, обладал лунным календарем. Месячный лунный цикл, как известно, хронометрически близок женскому месячному циклу. Отсюда напрашивается вывод, что интервенция женского образа в знаковую систему франко-кантабрийского типа отражала лунно-календарный характер последней. Таким образом, в данной знаковой системе следует видеть комбинацию календарных и счетных, арифметических представлений, воплощенных в символике лунного месяца и количественного пояснения к нему.
В целом франко-кантабрийскую знаковую систему следует разделить на три класса изображений. Первый не имел никакой счетно-календарной окраски (меандры, хаотичные черты, скобления и т. п.) и, как показано выше, мог относиться к сюжетному кругу франко-кантабрийской мифологии. Два других класса имели отчетливый счетно-календарный характер. Во-первых, это чисто количественные знаки: одинарные и двойные черты, ряды черт и пунктуаций. Данные знаки служили количественными пояснениями к более сложным качественным знакам. Нередко количественные знаки фигурировали изолированно, но, очевидно, подразумевали известную художникам календарную природу (число дней текущего месяца или даже текущего года, поскольку иногда ряды пунктуаций отражали количество дней, превышающее длительность лунного месяца, например, в Эль Кастильо-1, Ле Труа-Фрер-2 и др.). Во-вторых, имелись качественные знаки, отражающие определенный образ (например, полуовал, овал, вульва, клавиформа, треугольник, угол, стрела, крест, прямоугольник, зигзаг и др.).
Вопреки А. Леруа-Гурану, все качественные знаки могли быть составной частью парных знаков, например: зигзаг + точка + линия из 14 точек (Масакулос и др.) или зигзаг + 13 палочек (Ардалес), ветвистый знак + пунктуации (Ле Кюзуль-де-Бракони, Марсула и др.) или пенниформа + палочки (Куньяк), анкориформа (полуовал) + полоса пунктуаций (Ла Меаса и др.), овал + палочка (Бернифаль, Лабастид, Оцозелайя-Аризтойя и др.), вульва, женщина + пунктуации или ряды палочек (Тито Бустильо, Комарк и др.), треугольник + 6 параллельных черт (Сомбр и др.), угол + черта (Фигье, Байоль, Улан и др.), стрела + палочки (Сен-Сирк-2, Капова и др.), крест + палочки (Ласко-1, Ле Габийю-2 и др.), прямоугольный знак + палочки или пунктуации (Лас Чименеас, Монгодье и др.). Таким образом, все названные качественные типы знаков играют равноценную роль в составе парных знаков (без «мужских» и «женских» особенностей). Иногда качественные знаки могут комбинироваться друг с другом: овал и треугольник (Сомбр), пенниформа + 2 вульвы (Гуй) и т. п., откуда следует, что разные качественные знаки обозначали различные месяцы. В этой связи встает вопрос о характере франко-кантабрийского календаря.
Сравнение календарных представлений древних кельтов с мегалитическим календарем показывает, что начало лета у палеоевропейцев приходилось на 5 мая, начало осени – на 6 августа, начало зимы – на 2 ноября, а начало весны – на 2 февраля (как следует из шестнадцатимесячного солнечного календаря мегалитической культуры в бронзовом веке, 1800 до н. э.). Приведенные даты сезонов в бронзовом веке Западной Европы (Британии) объясняют происхождение календаря у ирландских кельтов, у которых начало лета приходилось на 1 мая (праздник Бельтене в ночь на 1 мая, когда возжигался священный огонь), а начало зимы – на 1 ноября (праздник Самайн в ночь на 1 ноября, когда священный огонь тоже разводился). Последний праздник положил начало всем известному Хэллоуину, шотладскому (т. е. кельтскому) и американскому кануну Дня всех святых, приходящемуся на 31 октября и ведущему начало, по крайней мере, из бронзового века Британских островов. Более древний лунный календарь палеоевропейцев должен быть проще, однако нельзя исключать преемственности основных его понятий с мегалитическим календарем.
Изобразительные особенности и статистика позволяют выделить во франко-кантабрийском репертуаре фигур десять крупных самостоятельных классов знаков и предположить их календарную интерпретацию.
Первый класс знаков могут составить зигзаги и близкие к ним серпентиформы (змееобразные знаки). Исходя из топографической карты Межирича (см. выше), можно полагать, что зигзаг символизировал воду. Таким образом, месяц зигзага, вероятно, отвечал началу половодья и началу весны (т. е. нашему марту).
Второй класс знаков должны образовать тонкие качественные знаки. К ним относятся схожие зубчатые и ветвистые знаки, вилка – редуцированный ветвистый знак, крюк – редуцированный зубчатый знак, Т-образный знак как вариант вилки, пенниформа (перообразный знак) как вариант ветвистого знака, а также фаллос как элементарный аналог всех предшествующих типов (точка зрения А. Леруа-Гурана, пока что не подкрепленная доверхнепалеолитическими находками). Класс ветвистых знаков, предположительно, мог отражать вегетацию растений (весеннее распускание у них листьев) и обозначать месяц начала вегетационного периода, т. е апрель. В силу известных ассоциаций (в том числе мифологических представлений, скажем, о Приапе как фаллическом божестве растительного плодородия) сближение фаллического символа с месяцем начала вегетации растений представляется оправданным. Вопреки иконографической очевидности, сюда же приходится отнести знак скалиформы (лестницы), поскольку на топографической карте из Межирича именно скалиформами передаются деревья леса – по-видимому, их необходимо связать с прочей растительной символикой.
Сравнение верхнепалеолитических изобразительных знаков в виде кругов с вырезом (т. е вульв) и следов северного оленя на твердом снегу или льду показывает их тождество [573, с. 687, рис. 11]. Это обстоятельство заставляет связать иконографический знак «вульвы» (круга с вырезом) с месяцем весенней миграции северных оленей, т. е. с маем, когда олени движутся по отступающему снежному насту (и печатают вышеуказанные следы), а не уходят от него, как при осенней миграции (и не оставляют следов на снегу или льду). Как представляется, упрощенными вариантами знака «вульвы» (т. е. оленьего следа) являются знаки в виде овала, круга и кольца. Миграция северного оленя меняла промысловую ситуацию в приледниковой Европе и вызывала живой отклик у создателей верхнепалеолитического календаря.
С двумя следующими группами знаков мы попадаем в область смысловых догадок. Однако в общей системе верхнепалеолитического календаря этим знакам действительно нечему больше соответствовать, кроме месяцев июня и июля. Знак в виде полуовала, полукруга, дуги и т. п. ассоциируется с месяцем летнего солнцестояния и концом солнечного полугодия, т. е. с июнем. Знак в виде «кометы» (вертикальные линии, собранные в треугольный пучок) напоминает знойные испарения или марево. Столь прямолинейная трактовка допускается потому, что в Ле Труа-Фрер-2 раненый медведь изрыгает поток крови в виде аналогичного знака, только размещенного горизонтально и исходящего из пасти зверя. Подобные веерообразные изображения, сокращенные порой до тройки конвергентных черт (Пергусе) или до пары черт, сведенных углом, исходят из ноздрей животных и называются «дыханием» (Ле Пеш-Мерль-2, Ле Травер-де-Жануа, Лос Касарес). Отсюда проистекает предположение, что «кометы» выражали именно то, что в них видится. «Кометы» как символы знойного марева, а также их стилизации в виде «вееров», треугольников и скутиформ (щитообразных знаков) могли бы выпадать на месяц зноя, т. е. на июль.
Для первобытных людей, приверженных, помимо охоты, собирательству, женские изображения и их стилизованные силуэты-клавиформы, скорее всего, ассоциировались с месяцем урожая съедобных растений и плодов и, соответственно, с месяцем женского собирательства, т. е. с августом.
Седьмой знаковый класс связан со стрелообразными фигурами и дротиками. В Ла Пенья-де-Кандамо-3 имеется качественное изображение благородного оленя, пораженного шестью дротиками. Дротиками были ранены и мужчины в Куньяк и Ле Пеш Мерль-2. В Су-Гран-Лак-2 атакуемого рогом бизона мужчину поражает со спины тройка стрел. Похоже, что стреловидные знаки и дротики восходят к изображению наступательного оружия: стрела – это дротик с наконечником типа Ла Фон-Робер и т. п., а дротик – то же самое, но без наконечника. Данный знак входил в систему парных знаков (например, семь стрел под пунктуацией из 27 точек на панно отпечатка в Ласко-1, стрела и палочка в Каповой и т. п.). Как представляется, стрелы и дротики мыслились в качестве атрибутов подготовки в сентябре к большому охотничьему сезону.
К восьмому знаковому классу принадлежат изображения в виде углов, или «ран», когда они помещаются на корпусах животных. По-видимому, они выражают месяц массовой охоты на ведущих крупных четвероногих (октябрь), начинающийся вслед за их репродуктивным периодом (периодом размножения в мае – сентябре у лошади и бизона).
Девятый класс знаков состоит из крестообразных фигур (прямоугольный или косой крест), которые следует дополнить звездчатыми знаками (шести– и восьмиконечными астериксами и т. п.). В Ласко-1, Ле Габийю-2 и в других святилищах крест образует с палочками характерный парный знак в виде римских цифр XI, XIII и т. п. А. Н. Каландадзе предположил [49, с. 22], что крестовидная фигура символизировала скрещенные палочки для получения огня трением. В этом случае кресты и астериксы должны были символизировать огонь, очаг и т. п. Судя по древнеирландскому празднику Самайн (наступление зимы в ночь на 1 ноября), палеоевропейцы отмечали наступление зимы разведением священного огня. С учетом этого обстоятельства, знак креста, отвечающий огню, ассоциируется с ноябрем.
Последний знаковый класс объединяет знаки прямоугольных или близких очертаний: простые прямоугольники, прямоугольники, расчерченные на квадраты в виде шахматной доски (гербы), ромбы, решетки, сетки, барьеры, тектиформы (крышеобразные изображения, напоминающие хижины) и пектиформы (гребнеподобные изображения). А.Леруа-Гуран считал, что эти знаки развились в ходе стилизации символа женского пола в рамках франко-кантабрийского искусства, однако в классической форме они известны уже в нижнем палеолите (Бильцингслебен, три экземпляра, два из которых весьма отчетливы).
На топографической карте из Межирича, где нанесено селение на берегу реки, решетка-сетка и прямоугольники с вырезом (двугорбые тектиформы, обманчиво напоминающие перевернутые признаки женского пола) символизируют жилища. При этом прямоугольник с вырезом (собственно тектиформа) отвечает жилой части сооружения, а решетка-сетка – очагу перед его входом. На четыре жилища – очаг один, поэтому он, вероятнее всего, устроен перед «административно-сакральной» хижиной вождя и колдуна, которая реально известна в Межириче по археологическим данным – это т. н. жилище № 1, вход которого был отмечен черепом мамонта, украшенным ветвистым знаком месяца апреля. Данный очаг в виде решетки-сетки является священным, а потому при изолированном, знаковом воплощении, должно быть, символизирует конец зимы – начало весны, возможно, конец – начало года, когда возжигался священный огонь, как в начале лета и зимы. Иными словами, решетка-сетка приходилась на наш февраль.
Сказанное заставляет увязать данный знаковый класс с символикой жилья на зимовке. Месяц разбивки стационарных жилищ с начертанием на грунте их планов соотносится с прямоугольниками, гербами и ромбами, которые, таким образом, отвечают декабрю. Месяц собственно зимовки под домашним кровом, вероятно, отвечает тектиформам и родственным им барьерам с пектиформами, которые, соответственно, приходятся на январь. Высказанные предположения относительно трех групп знаков для зимних месяцев находят подтверждение статистикой.
Частоты знаков всех предыдущих классов колеблются в пределах от 14,9 до 30,6%, а их градиент (ступени расхождения частот близких по встречаемости знаков) достаточно плавен и не выходит за пределы 0,5–3,2%. У прямоугольных же знаков общая частота доходит до 39,6%, а отрыв от прочих классов составляет 9,0%, что не укладывается в рамки градиента для остальных знаков. Эта аномалия может объясняться тем, что класс прямоугольных знаков отвечал большему временному интервалу месяцев, нежели остальные классы. Действительно, если франко-кантабрийский календарь был лунным, то, в принципе, он должен был состоять не из 10 (как первоначальные этрусский и римский календари), а из 12 месяцев, при условии, что месяцы близки лунным. В солнечном году содержится 12,38 синодического лунного месяца, и примерно столько же следует отводить для франко-кантабрийской системы, если принимать ее как лунную.
Интерпретация решетки-сетки как февральского знака имеет археологическое подтверждение. В пещере Бломбос, ЮАР, 77 ± 6 тыс. лет назад, пара кусков охры несет на себе выгравированные знаки [412, с. 1279, рис. 2], представляющиеся календарными символами месяцев. Несколько выше залегал фрагмент с изображением зигзага, который ассоциируется с началом половодья и началом весны (т. е. с мартом). Несколько ниже, а значит раньше, был оставлен знак в виде косой сетки в прямоугольном контуре, напоминающий косую сетку в трапециевидном контуре из Бильцингслебена, отвечающую одному из трех зимних месяцев. Стратиграфия Бломбоса позволяет предполагать, что знак в виде косой сетки в четырехугольном контуре соответствовал февралю, предшествующему марту, отражаемому зигзагом (оговоримся, что в Южном полушарии сезоны были обратны европейским, но календарь ориентировался на сезонные явления природы, а потому мартовский зигзаг Европы мог отвечать сентябрьскому зигзагу Южной Африки, что не меняет сути дела).
Если разбить класс прямоугольных знаков на три группы, мы получим декабрьскую группу прямоугольников, гербов и ромбов (21,6%), январскую группу тектиформ, барьеров и пектиформ (15,3%) и февральскую группу решеток-сеток (17,1%). В этом случае их частоты укладываются в пределы, свойственные прочим классам знаков, а градиент различий между всеми классами снижается до 0–2,7%, что статистически благоприятно. Максимальная частота остается за вторым классом ветвистых знаков, а его отрыв от прочих классов (2,3%) является рядовым.
Клетки внутри гербов могли быть разноцветными (Ласко-1). В нижнем – среднем палеолите засвидетельствованы знаки типа прямоугольников (Бильцингслебен, Молодова I (2)) и, возможно, косая сетка в трапециевидном контуре (Бильцингслебен), а также барьер (Молодова I (2)). Иными словами, все три группы зимних знаков имеют доверхнепалеолитическое происхождение и не соответствуют гипотезе А. Леруа-Гурана относительно символики женского пола, якобы воплощенной в них.
Имеется еще по крайней мере две группы знаков, в отличие от предыдущих, не засвидетельствованных в нижнем – среднем палеолите. Во-первых, это знаки в виде акколад (фигурных скобок), некоторые из которых напоминают неполные гербы (Ле Габийю-2, Ласко-1 и, может быть, Ласко-2) или деформированные прямоугольные знаки (Эль Кастильо-1, Ла Пилета, Митро). Иные (собственно фигурные скобки и авиаформы, т. е. птицеобразные знаки), возможно, происходят от женских символов (Куньяк, Ле Кюзуль-де-Бракони, Леванцо, Улан, Ла Пасьега В, Ле Пеш-Мерль-2, Ле Портель-1, Ле Труа-Фрер-1, Виллар и др.). Таким образом, акколады образуют гетерогенную, разнородную группу, подгруппы которой находят место в вышеперечисленных знаковых классах.
Во-вторых, это знаки в виде отпечатков или контуров руки, иногда с поджатыми пальцами (Гаргас I-1, Тибиран-1, Мальтравьесо, Ласко-1). Их количество в одном сятилище может быть огромным (250 экземпляров в Гаргас I-1), а может быть удивительно невелико по сравнению с общим количеством фигур в святилище (несколько отпечатков рук с поджатыми пальцами в Ласко-1 при общем количестве фигур свыше 1 тыс.). Складывается впечатление, что количество рук в святилище обратно пропорционально количеству качественных календарных знаков (в Гаргас I-1 есть несколько крестообразных фигур, один круг, одна серпентиформа, одна вульва, один фаллос на 148 фигур животных, тогда как в Ласко-1 количество календарных знаков велико, и они разнообразны).
Иногда руки сопровождаются пунктуациями (Гаргас I-1, Тибиран-1), что наводит на мысль об их принадлежности к числу широких знаков (по А. Леруа-Гурану) или – к числу качественных календарных знаков (по нашему мнению). Последнее предположение объясняет обратное отношение рук и календарных знаков при условии, что руки выражали понятие месяца вообще (это значило бы, что, например, Гаргас I-1 украшался с марта по ноябрь в течение 11 лет). Поджатые пальцы, вероятно, означали прошедшие «недели» месяца, т. е. четверти лунного месяца, отвечающие фазам луны. Соответственно, рука с полностью раскрытой ладонью, должно быть, подразумевала полный месяц. В пользу календарного понимания рук говорит и следующее обстоятельство.
Франко-кантабрийские художники были европеоидами и говорили на языке, родственном шумерскому, сино-кавказским и ностратическим языкам. Сравнение же ностратических слов *man'u «пятерня, кисть руки» и *man'γä «месяц (небесный, календарный)» (суффиксальное образование на -γä) и учет доностратической древности этих основ у европеоидной расы, вытекающей из шумерских параллелей (ср. шумер. Nannar «Месяц» <*mannar по ассимиляции согласных), позволяет связать отпечатки рук (открытых ладоней, ладоней с поджатыми пальцами) в украшенных верхнепалеолитических пещерах с понятием календарного месяца. Возможно, речь идет о прежитках древнего языка жестов у наших предков (см. разд. 3.2).
Проанализированная нами знаковая система, возможно, отражала двенадцатимесячный календарь в уже законченной форме (рис. 17), и возникает предположение, что она имела определенную предысторию.
Число памятников мобильного искусства доверхнепалеолитического возраста достигает пяти-шести десятков, из которых мы учли 48. Описание некоторых памятников, например аккайской мустьерской культуры, не содержит точных хронологических привязок и может быть учтено лишь частично, когда ясен археологический слой находок. Древнейшим свидетельством изобразительной деятельности является позвонок слона из местонахождения Странска скала, на котором нанесено семь правильных радиальных зарубок. По-видимому, мы имеем дело со счетным знаком франко-кантабрийского типа, аналогичным знакам в виде рядов черт. Древнейший парный знак известен в местонахождении Вертешсёллёш: это зигзаг, сопровождаемый одинарной чертой (1 марта), на орудии из бычьей кости. Древнейшая скульптура происходит из местонахождения Берехат Рам. Это галька, чуть подправленная, чтобы создать на ее вершине ободок-шею. В этом произведении исследователи усматривают старейшую «Венеру», антропоморф женского пола. Древнейшая гравюра открыта в местонахождении Пронятин, где продольно расчлененная лучевая кость зубра несет на себе предельно схематичное очертание животного, в котором узнают лошадь (животное, право, больше напоминает павиана, но в тогдашней приледниковой Украине павианы не водились). Наконец, по имеющимся (но, вероятно, не окончательным) данным, на склоне горы Богутлу появляются первые образцы анималистической кремневой пластики, представляющие собой неутилитарные изделия, грубо подправленные для того, чтобы искушенный первобытный глаз разглядел в них абрисы любимых мифологических животных (на что ныне способен лишь опытный глаз археолога). Трудности идентификации первых произведений искусства находятся в согласии с нашими представлениями о нелегкой и неумелой заре художественного творчества – все начиналось с нуля.

Рис. 17.
«Надпись» из пещеры Ла Пасьега С-2
в Испании, 14800–14300 лет назад. Она означает, что пещерное святилище украшалось и служило ритуальным целям 4 января, 1 мая и в декабре месяце энного года (приблизительно в 12600 ± 250 до н. э.). Ее значение не следует преувеличивать. Буквально она переводится как календарная пометка «4 января, 1 мая, декабрь (тектиформа + 4 черты, следы животного + одинарная черта, квадратный знак)», а все остальное домысливается. Судя по изобразительной деградации знаков, способ чтения в данном случае был слева направо, поэтому, возможно, нет указания на число декабря месяца. На наш взгляд, здесь можно говорить лишь о самых элементарных началах письменности, поскольку полноценная письменная система должна обладать способностью универсально отражать мир. В палеолите графические системы существовали, однако не располагали указанным свойством универсального отражения действительности. Во всяком случае, нам не известны факты на этот счет, хотя франко-кантабрийская знаковая система представлена богато [498, с. 462, рис. 799]
Поэтому первый изобразительный стиль заслуживает наименования «нулевого» (рис. 18). В противоположность франко-кантабрийскому стилю I, в котором, например, костяная пластика была не лишена признаков художественной состоятельности, стиль 0
схематичен на 100%. Его произведения разгадываются, а не опознаются. За вычетом оленя из Молодовы I (2), у которого голова в виде закорючки снабжена рогами и поэтому легко узнается, произведения стиля 0 не содержат каких-либо деталей, если не считать схематичных конечностей упомянутых лошади и оленя. Иными словами, образцы стиля 0 всецело напоминали понятийные иероглифы, объемные или плоские, так что пользователи этого искусства сперва учились его читать (распознавать в нем мифологические сюжеты), а уж потом наслаждались увиденным сакрально (см. разд. 3.2). Превращение стиля 0 в стиль I произошло путем детализации произведений, позволяющей опознавать их без подсказки.
Отметим, что в искусстве нижнего – среднего палеолита представлено 9 месячных групп франко-кантабрийских знаков. Причем по крайней мере в 6 случаях образцы подобных знаков входят в состав парного знака, т. е. группируются с палочками или пунктуациями. Это справедливо для зигзага (Вертешсёллёш, Молодова I (2) и (4), Ле Пеш-де-л’Aзе II, Тата), ветвистого знака, вилки, скалиформы (Молодова I (2) и (4)), женщины (Берехат Рам, где о признаках парного знака сведений нет), угла (Молодова I (2) и (4), Тата, Эрмитаж), креста (Вилен), прямоугольника (Бильцингслебен, Молодова I (2)), ромба (Молодова I (2)) и сетки (Бильцингслебен).
В общей сложности парные знаки представлены в 8 местонахождениях (16,(6)%), а качественные календарные знаки разного рода – в 14 местонахождениях (31,(1)%; кроме названных, добавлены Бачо Киро, Пролом II (2), Турске Маштале, Цона). Счетные знаки (палочки, пунктуации) встречаются значительно чаще (62,5%), что в общем отвечает ситуации во франко-кантабрийском искусстве. Приведенные факты позволяют предполагать, что счетно-календарная система франко-кантабрийского типа существовала уже в нижнем – среднем палеолите и была унаследована верхнепалеолитическим искусством. Возраст этой системы неразрывно связан с возрастом счетных знаков и может достигать Гюнц/Минделя II (ок. 783 тыс. лет, Странска скала), хотя, строго говоря, качественные календарные знаки появляются лишь в Минделе I/II (Вертешсёллёш). Прототипы счетных знаков можно искать в параллельных чертах на кости из Сензель (Полиньяк, Верхняя Луара, Франция, верхний виллафранк, Гюнц I, 1,3 млн лет назад по калий/аргону), однако эти «графемы» могут оказаться следами снятия мяса с костей.
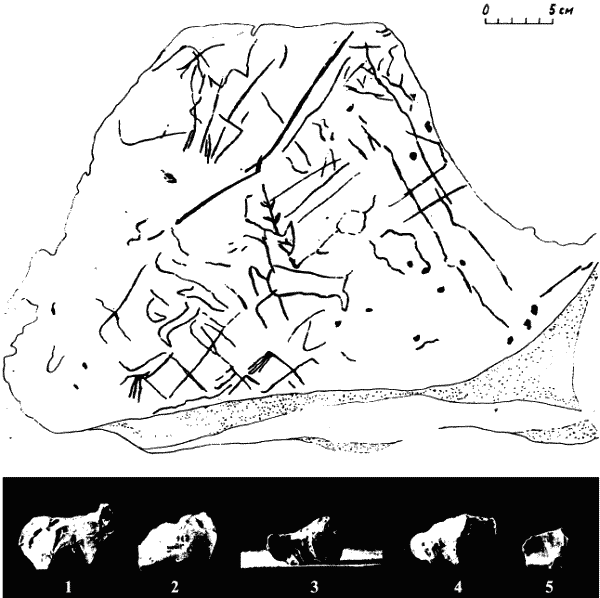
Рис. 18.
Стиль 0. Гравировки и рисунки на лопатке мамонта из мустьерского слоя 2 стоянки Молодова I на Украине. Предположительный смысл изображения мог быть таким («читаем» слева направо, сверху вниз): «1 марта (зигзаг + одинарная черта); 3 апреля (ветвистый знак + тройная точка); 9 апреля (скалиформа + 9 точек); 2 октября – Олень-утреннее и вечернее Небо (угол + двойная точка + большерогий (?) олень); декабрь (тройной ромбический знак)». Ждем новых прорисовок памятника. Кремневая пластика (смутные фигуры животных): 1, 2 – поздний ашель стоянки Богутлу в Армении; 3–5 – мустье стоянки Яштух в Абхазии и Ахштырской пещеры в Краснодарском крае России [61, с. 40; 113, с. 65, рис. 27]
Анималистическое искусство нижнего – среднего палеолита пока известно мало. Оно представлено в 27,1% соответствующих местонахождений. Его бестиарий, по-видимому, носил франко-кантабрийский характер. Представлены лошадь (Пронятин), олень (Молодова I (2)), медведь (Чокурча II), а также неопределенные четвероногие, запечатленные кремневой скульптурой, выявленной Э. Е. Фрадкиным среди археологических материалов из местонахождений Ахштырь, Богутлу, Иль, Келасури, Киик-Коба (6, 5), Лечкоп, Отхары, Староселье, Тешик-Таш, Яштух [105; 106; 61]. Учитывая генетические связи знаковой системы нижнего – среднего палеолита с франко-кантабрийской, можно думать, что нижне-среднепалеолитическая анималистическая система также состояла в генетической связи с франко-кантабрийской анималистической мифологией, что проливает свет на происхождение последней.
Наиболее древняя кремневая скульптура появляется в позднем ашеле Богутлу, что можно рассматривать как минимальный возраст анималистической мифологии франко-кантабрийского типа. На наш взгляд, эта мифология была неразрывно связана со счетно-календарной системой, что документально доказывается изображениями на лопатке мамонта из Молодовы I (2), где фигурка оленя сопровождается весьма богатой знаковой символикой. В Молодове I (4) со знаками соединена антропоморфная фигурка, которая также может рассматриваться как элемент мифологии франко-кантабрийского типа (и расцениваться в первом приближении как изображение колдуна Коня-Солнца). Не следует исключать и возможности того, что возраст анималистической мифологии достигал возраста счетно-календарной системы нижнего – среднего палеолита.
Как можно убедиться, статистический анализ палеолитического искусства дает основания считать его средством выражения анималистической мифологии, связанной со счетно-календарными представлениями. Возраст соответствующей системы взглядов достигал Гюнц/Минделя II (ок. 783 тыс. лет назад). Носителями доверхнепалеолитического искусства являлись гейдельбергский человек (Вертешсёллёш, Бильцингслебен), неандерталец (Киик-Коба (6, 5), Ле Мустье (J), Заскальная VI (3a), Ла Ферраси, Ла Кина, Пролом II (1), Джручула, Тешик-Таш) и человек современного вида (Ахштырь, Староселье), что указывает на их сходные способности к эстетической деятельности и мифологическому творчеству. Проще говоря, бытующие представления о том, что человек современного вида всегда был умнее и чувствительнее неандертальцев (которые якобы не были способными к художественному творчеству [90, с. 420]) или умнее и чувствительнее гейдельбержцев, не подкрепляются научными фактами.
Приложение 2
Статистика сюжетов палеолитического искусства
Учтено 222 наскальных франко-кантабрийских (их номера не подчеркнуты) и 48 мобильных доверхнепалеолитических памятников. Рассчитан процент встречаемости сюжетов: франко-кантабрийских (слева) и доверхнепалеолитических (справа).




Приложение 3
Местонахождения мобильного искусства нижнего и среднего палеолита (стиль 0)
Абсолютные даты даны от «наших дней», т. е. от 1950 г. н. э.
ГЮНЦ/МИНДЕЛЬ, 874 000–529 000
Гюнц/Миндель II, 790 000–770 000
1. Странска скала (Червени копец, пещера IV), Брно, Южная Моравия, Чехия. Олдовай. Гюнц/Миндель II, менее 783 000 (палеомагнетизм).
МИНДЕЛЬ, 529 000–388 000
Миндель I/II, 493 000–411 000
2. Вертешсёллёш, Татабанья, Комаром, Венгрия. Буда (прототейяк). Миндель I/II, менее 700 000 (термолюминесценция) – св. 350 000 (торий/уран). Homo heidelbergensis.
МИНДЕЛЬ/РИСС, 388 000–333 000
3. Тера Амата (С 1а), Ницца, Приморские Альпы, Франция. Древний ашель (II). Миндель/Рисс I, 380 000 (термолюминесценция).
4. Бильцингслебен (Штайнринне), Артерн, Эрфурт, Тюрингия, Германия. Древний тейяк (протошарант). Миндель/Рисс III, 350 000–300 000 (торий/уран). Homo heidelbergensis.
РИСС, 333 000–144 000
Рисс I, 333 000–281 000
5. Грот дю Пеш-де-л’Азе II (8), Карсак, Дордонь, Франция. Южный средний ашель (IV). Рисс I (Перигор II).
6. Маркклеберг, Лейпциг, Саксония, Германия. Поздний ашель (V). Рисс I (Дрента).
Рисс II, ок. 260 000 – ок. 240 000
7. Берехат Рам, Голанские Высоты, Израиль. Ашель, ок. 250 000 (800 000–230 000).
Рисс III, 219 000–144 000
8. Вилен, Лёррах, Карлсруэ, Баден, Баден-Вюртемберг, Германия. Мустье. Рисс II (= II, III, Варта).
РИСС/ВЮРМ, 144 000–110 000
9. Богутлу, Пирмалак, Верин Талин, Армения. Поздний ашель (V).
10. Грот д’Истюриц (М), Сен-Мартен-д’Арберу, Атлантические Пиренеи, Франция. Типичное мустье с топориками (экс-васкон). Рисс/Вюрм.
11. Пещера Кульна (11), Слоуп, Бланско, Южная Моравия, Чехия. Мустье шарантской традиции типа Таубах (Кина). Рисс/Вюрм.
12. Пещера Янкович (низ), Комаром, Венгрия. Микок типа Янкович (экс-задунайский селет, с конца Рисс/Вюрма).
13. Ильская (низ), Ильский, Краснодар, Россия. Мустье шарантской традиции типа Кина. Конец Рисс/Вюрма, 135 000 ± 25 000 (уран).
14. Грот Киик-Коба (6, 5), Тау-Кипчак, Симферополь, Крым, Украина. Зубчатое мустье. Рисс/Вюрм, 111 000 (коллаген). Homo neanderthalensis.
ВЮРМ I, 110 000–85 000
Вюрм I Амерсфорт, 105 000–104 000
15. Пещера Кульна (9 В), Слоуп, Бланско, Южная Моравия, Чехия. Мустье шарантской традиции типа Таубах (Кина). Вюрм I Амерсфорт.
Вюрм I Брёруп, 100 000–92 000
16. Тата (Кальвария-Берг), Комаром, Венгрия. Мустье шарантской традиции типа Таубах (Кина). Вюрм I Брёруп, 50000 ± 2500, 33600 ± 1100 (14 С), (95000 ± 10000, уран).
Вюрм I С, 92 000–85 000
17. Рипаро Тальенте, Верона, Венето, Италия. Мустье. Вюрм I C.
18. [Грот] Заскальная VI (4), Вишенное, Белогорск, Крым, Украина. Мустье ашельской традиции типа А (ак-кая, Вюрм I – II). Вюрм I C.
19. Грот Киик-Коба (4), Тау-Кипчак, Симферополь, Крым, Украина. Мустье ашельской традиции типа А. Вюрм I С, 86 000 (коллаген).
ВЮРМ II, 84 000–39 000
Вюрм
II A, 84 000–72 000
20. Бломбос Кейв, Южный мыс (Игольный), Капская провинция, Южная Африка. Средний каменный век. 77 000 ± 6000 (термолюминесценция).
21. Пещера Бачо-Киро (12), Дряново, Габрово, Болгария. Типичное мустье. Вюрм II A.
22. Пронятин, Тернополь, Украина. Типичное мустье (молодова, Вюрм II A – Вюрм II/III Хенгело, св. 45 600 – св. 40 300, 14 C). Вюрм II A.
23. Молодова I (4), Сокиряны, Черновцы, Украина. Типичное мустье (молодова). Вюрм II A, св. 44 000 (14 C).
24. Молодова I (2), Сокиряны, Черновцы, Украина. Типичное мустье (молодова). Вюрм II A.
Вюрм II Дюрнтен, 72 000–59 000
25. Чокурча II, Луговой (Чокурча), Симферополь, Крым, Украина. Типичное мустье. Вюрм II Дюрнтен.
26. Пещера Ахштырь (13), Адлер, Сочи, Краснодар, Россия. Зубчатое мустье (хоста, Вюрм II Дюрнтен – Вюрм II/III Ле Котте, св. 58 000 (коллаген) – 35680 ± 480 (14 С), 35 000 ± 2000 (уран)). Вюрм II Дюрнтен, св. 58 000 (коллаген). (Слои 8-7: Вюрм II/III – Ле Котте, 37 000 (коллаген) – 35 000 ± 2000 (уран). Homo sapiens).
Вюрм II B, 59 000–51 000
27. [Грот] Заскальная VI (3 a), Вишенное, Белогорск, Крым, Украина. Мустье ашельской традиции типа А (ак-кая). Вюрм II B 1. Homo neanderthalensis.
28. [Грот] Пролом II (2), Белогорск, Крым, Украина. Мустье aшельской традиции типа А (ак-кая). Вюрм II B 2 (св. 50 000 (14 C), Заскальная V (2), ак-кая).
29. [Грот] Заскальная VI (2), Вишенное, Белогорск, Крым, Украина. Мустье ашельской традиции типа А (ак-кая). Вюрм II B 2, св. 45 000 (14 C).
30. Грот де л’Эрмитаж (2), Люссак-ле-Шато, Вьенна, Франция. Мустье шарантской традиции типа Ферраси. Вюрм II.
31. Грант абри де Ла Ферраси (CD), Савиньяк-де-Мирмон, Дордонь, Франция. Мустье шарантской традиции типа Ферраси. Вюрм II. Homo neanderthalensis.
32. Нижнее абри дю Мустье (G), Ле Мустье, Дордонь, Франция. Мустье ашельской традиции типа А. Вюрм II В 1–2 (Перигор IV – VI), 55 800 ± 5000–50 300 ± 5500 (термолюминесценция).
33. Абри де Ла Кина (3), Ле Гард, Шаранта, Франция. Мустье шарантской традиции типа Кина. Вюрм II В 2 (Перигор VI). Homo neanderthalensis.
34. Грот де л’Эскишо-Грапау, Сент-Анастаси, Гар, Франция. Мустье шарантской традиции типа Кина. Вюрм II.
Вюрм II Мурсхофд, 51 000–46 500
35. Гротта ди Госто (DC), Монтанья ди Четона, Сартеано-Четона, Сиена, Тоскана, Италия. Мустье шарантской традиции типа Ферраси. Вюрм II Мурсхофд (Перигор VII), 48 000 ± 4000 (торий/уран).
36. Навес Староселье, Бахчисарай, Крым, Украина. Восточно-европейский микок. Вюрм II Мурсхофд (вне навеса: 41 000, 14 С). Homo sapiens.
Вюрм II C, 46 500–39 000
37. Грот дю Портель (в глубине сзади), Лубенс, Арьеж, Франция. Типичное мустье. Вюрм II С (Перигор VIII).
38. [Грот] Пролом II (1), Белогорск, Крым, Украина. Мустье ашельской традиции типа А (ак-кая). Вюрм II C. Homo neanderthalensis (?).
39. Пещера Джручула, Квемо-Зоди, Чиатура, Имеретия, Грузия. Типичное мустье (кударо, Вюрм I/II – Вюрм II C, 65 000 ± 5000 (уран) – 44 150 + 2400/ – 1850, 14 C). (Слой 12, Вюрм II A. Homo neanderthalensis).
40. Пещера Цона (5), Джава, Юго-Осетия, Грузия. Типичное мустье (кударо). Вюрм II Мурсхофд – Вюрм II C (?).
41. Пещера Тешик-Таш, Юкары-Мачай, Байсун, Сурхандарья, Узбекистан. Типичное мустье. Вюрм II C, 44 000 ± 1000 (уран). Homo neanderthalensis.
42. Нижнее абри дю Мустье (J), Ле Мустье, Дордонь, Франция. Типичное мустье. Вюрм II C (Перигор VIII), 40 300 ± 2600 (термолюминесценция). Homo neanderthalensis.
* * *
43. Яштух, Нижний Яштух, Сухуми, Абхазия, Грузия. Позднее зубчатое мустье.
44. Лечкоп, Сухуми, Абхазия, Грузия. Развитое мустье.
45. Келасури, Сухуми, Абхазия, Грузия. Развитое мустье. Верхний плейстоцен, после Рисс/Вюрм I, 144 000–127 000.
46. Отхары, Гудаута, Абхазия, Грузия. Мустье.
47. Гротта ди Тиволи, Рим, Лацио, Италия. Мустье шарантской традиции понтийского типа (Кина).
48. Пещера Турске Маштале, Тетин, Прага, Центральная Чехия, Чехия. Мустье.
Литература:
[17, с. 201, табл. XVIII, 10]; [49, с. 22]; [52, с. 223–226]; [56, с. 67–68, 107–109]; [61]; [81, с. 27]; [93, с. 124–125]; [94]; [102, с. 216]; [105]; [106]; [112]; [113, с. 6–102]; [147, с. 1451–1453]; [162, с. 346]; [171, с. 92]; [201, с. 180; 201]; [292, с. 93–94: Abb. 38–40]; [309, с. 37, 63]; [412]; [429, с. 217–231]; [496]; [500, с. 526, 588, 1001]; [533, с. 168–169, Abb. 146. № 3, с. 262–269]; [536]; [537]; [538]; [539]; [542]; [620]; [621]; [622]; [631]; [751, с. 261–264, 283]; [757]; [764]; [773].
Приложение 4
Местонахождения франко-кантабрийского наскального искусства
После порядкового номера святилища, помеченного звездочкой для точно датированных памятников (звездочка в скобках выражает проблематичность точной датировки) или вопросительным знаком для сомнительных (стилистически неопределенных), а также знаком «минус» для памятников с холодолюбивым бестиарием (мамонт, северный олень, шерстистый носорог, бескрылая гагарка; «минус» в скобках – для проблематичных случаев, когда о холодолюбивости бестиария свидетельствует лишь присутствие пещерного льва, медведя, полярной? совы, или фантомасовы, и тюленя), идут название пещеры (грот, куэва, кова, гротта) или убежища (абри, рипаро), которое для разрушенных памятников взято в квадратные скобки, а затем географическая привязка. Латинские цифры после названия памятника обозначают независимые одноименные гроты, арабские цифры – диахронические (разновременные) святилища в одном и том же гроте. Затем указана предположительно синхронная сопровождающая археологическая культура.
Иногда приводятся иллюстративные датированные мобильные памятники, близкие наскальным в стилистическом, археологическом или географическом отношении. Порядок памятников внутри стилей проблематичен. Памятники без бестиариев помечены указанием Зн
. (святилище знаков).
Хронологически сенсационный грот Шове отнесен к стилю II условно. Его стилистика близка стилю IV древнему, а датировка не считается окончательной [164]. В ее основу положено определение радиометрического возраста примененных красителей, а те могли преувеличенно состариться в силу неких нестандартных исторических ситуаций (в Альтамире, например, датированные красители страдали от исторических колебаний влажности [763, с. 69]).
Абсолютные даты даны «от наших дней», т. е. от 1950 г. н. э.
ПЕРВОБЫТНЫЙ ПЕРИОД (33 000–20 100)
СТИЛЬ I (33 000–32 400)
Вюрм III A (34 000–31 000)
Ориньяк I (33 000–30 600)
1. * – [Грант абри де Ла Ферраси], Савиньяк-де-Мирмон, Дордонь, Франция. Ориньяк I. Вюрм III A 2 (Перигор II A).
2. * [Абри Ларте], Валлон де Горж-д’Aнфер, Сирей, Лез Эйзи-де-Тейяк, Дордонь, Франция. Ориньяк I. Вюрм III A. Следы риcунков.
3. * – [Абри Кастане], Сержак, Дордонь, Франция. Ориньяк I. Вюрм III A. (Радиокарбон, калиброванный по методу NotCal04, – 40 500 [567 а]).
4. * [Абри Бланшар-де-Рош I], Сержак, Дордонь, Франция. Ориньяк I.
СТИЛЬ II (32 400–20 100)
5. * – Грот Шове, Валлон-Пон-д’Aрк, Ардеш, Франция; 32 400. (Радиокарбон, калиброванный по методу NotCal04, – ок. 36 000 [567 а]).
Перигор IV (древний граветт) (29 300–25 000) Интерстадиал Вюрм III Кессель (29 000–25 000) Ориньяк IV (29 000–22 500)
6. * – Гранд грот д’Арси-сюр-Кюр, Арси-сюр-Кюр, Йонна, Франция. 27 080 ± 410, 26 470 ± 390.
7. * Грот Коске, Марсель, Буш-дю-Рон, Франция. 27 000.
8. * Грот де Кюссак, Дордонь, Франция. 25 120 ± 120.
Вюрм III С 1 (25 000–23 500)
Перигор V (средний граветт) (25 000–22 500)
9. * – Грот де Пер-нон-Пер, Приньяк и Маркам, Жиронда, Франция. Перигор V A (и IV ?). Вюрм III C 1.
10. – Грот де Берну, Бурдей, Дордонь, Франция. Ориньяк (IV ?).
11. – Грот де Жовель, Ла Тур-Бланш, Дордонь, Франция.
12. – Грот де Ла Кроз-а-Гонтран, Сирей, Лез Эйзи-де-Тейяк, Дордонь, Франция (Перигор V C, Вюрм III C 1 (Перигор VI), абри Пато, Сирей, Лез Эйзи-де-Тейяк, Дордонь, Франция).
13. * – Грот де Гаргас I-1, Авантиньян, Верхние Пиренеи, Франция. Перигор V C.
14. ( – ) Грот де Тибиран-1, Тибиран-Жонак, Верхние Пиренеи, Франция.
15. – Грот де Рукадур, Темен, Ло, Франция. (Верхний перигор, грот дез Эскабас).
16. * – [Абри Лабатю], Сержак, Дордонь, Франция. Перигор V C.
17. * – Абри д’Орей-д’Анфер, Валлон де Горж-д’Aнфер, Сирей, Лез Эйзи-де-Тейяк, Дордонь, Франция. Перигор V B – C.
18. ( – ) Куэва де Ла Вента-де-ла-Перра, Карранса, Бискайя, Испания (Перигор V C, куэва де Болинкоба, Абадиано, Бискайя, Испания).
19. *( – ) [Абри де Лоссель], Марке, Дордонь, Франция. Перигор V A. Вюрм III C 1.
20. Абри дю Пуассон, Валлон де Горж-д’Aнфер, Сирей, Лез Эйзи-де-Тейяк, Дордонь, Франция. Перигор V C. Вюрм III C 1.
Интерстадиал Вюрм III Тюрсак (23 500–22 500)
21. Грот де Ла Мут-1, Сирей, Лез Эйзи-де-Тейяк, Дордонь, Франция.
22. (?) Грот дю Рок-де-Везак, Везак, Дордонь, Франция. Перигор V C.
23. Грот де Ла Грез, Марке, Дордонь, Франция. Перигор V A, C.
24. Куэва де Ла Пенья-де-Кандамо-1, Сан-Роман-де-Кандамо, Овьедо, Испания.
25. Куэва де Мальтравьесо, Мальтравьесо, Касерес, Испания. Зн.
.
26. Куэва де Ардалес, Алора, Малага, Испания.
Перигор VI (верхний граветт) (23 100–22 000)
27. Грот дю Кро-Марен, Монтиньи-сюр-Луэн, Фонтенбло, Сена и Марна, Франция (Перигор VI, О-ле-Рок, Монтиньи-сюр-Луэн).
Вюрм III C 2 (22 500–20 500)
Перигор VII (финальный граветт) (22 500–21 700)
28. * – Абри де Ложери-От, Сирей, Лез Эйзи-де-Тейяк, Дордонь, Франция. Перигор VII, 21 735 ± 250. Вюрм III C 2 (Перигор VIII).
Протосолютре (21 300–20 500)
29. – Грот Шабо, Эгез, Гар, Франция. Протосолютре. Вюрм III C 2 (Протосолютре, 20 650 ± 300, абри де Ложери-От).
30. – Грот дю Фигье, Сен-Мартен-д’Aрдеш, Ардеш, Франция. Протосолютре.
31. – Грот Байоль, Коллья, Гар, Франция.
32. – Грот де Ла Бом-Латрон, Рюссан-Сент-Анастаси, Гар, Франция (Нижнее солютре, 20 490, кова дель Парпальо, Гандия, Валенсия, Испания).
Интерстадиал Вюрм III Баньольс (20 500–20 300)
Нижнее солютре (20 500–20 100)
33. Грот де Мезеле, Валлон-Пон-д’Aрк, Ардеш, Франция. Солютре.
34. Грот де Ла Вашерес, Валлон-Пон-д’Aрк, Ардеш, Франция.
35. Грот де Ла Бом-де-Бушон, Валлон-Пон-д’Aрк, Ардеш, Франция.
36. Грот Сомбр, Сен-Мартен-д’Ардеш, Ардеш, Франция. Нижнее солютре.
Вюрм III C 3 (20 300–19 700)
37. – Грот д’Улан, Ле Гарн, Гар, Франция. Нижнее солютре, св. 19 710 ± 400, 19 360 ± 420.
38. – Грот Юшар, Сен-Мартен-д’Ардеш, Ардеш, Франция. Нижнее со-лютре.
АРХАИЧЕСКИЙ ПЕРИОД (22 600–15 500)
СТИЛЬ III ДРЕВНИЙ (22 600–16 500)
39. Гротта Паличчи, Риньяно Гаргано, Фоджа, Италия. Перигор V B. Вюрм III C 2, 22 600 ± 450.
40. * – Грот Майенн-Сьянс, Торинье-ан-Шарни, Майенн, Франция. Пе-ригор V (средний граветт), 22 600±380.
41. * Грот де Ла Тет-дю-Льон, Бидон, Ардеш, Франция. Вюрм III C 2, 21 650 ± 800.
42. – Грот де Гаргас I-2, Авантиньян, Верхние Пиренеи, Франция.
43. Куэва де Лос Орнос-де-ла-Пенья-1, Сан-Фелисес-де-Буэльна, Сан-тандер, Испания. Нижнее солютре.
Среднее солютре (20 100–19 500)
44. ( – ) Куэва де Ла Клотильде-де-Санта-Исабель, Торрелавега, Сантан-дер, Испания.
Верхнее солютре (19 900–16 500)
Интерстадиал Вюрм III/IV Ложери (19 700–18 500)
45. Грот де Труа-Фрер-1, Монтескье-Авантес, Арьеж, Франция.
46. Куэва де Ла Меаса, Комильяс, Сантандер, Испания. Зн.
.
47. Куэва де Кинтаналь, Бальмори, Овьедо, Испания (Среднее солютре, Куэто де Ла Минья, Овьедо, Испания).
48. Грот дю Рок-д’Алла, Сент-Андре-д’Алла, Дордонь, Франция.
Финальное солютре (18 900–18 300)
49. Грот де Пюи-Жарриж II, Брив, Коррез, Франция (Верхнее солютре, грот Фуйяд-дю-Рес, Брив).
50. ? Грот дю Мулен-де-Лагене, Шасто, Коррез, Франция. Зн.
.
51. Куэва де Лас Эррериас, Льянес, Овьедо, Испания. Зн.
.
52. Грот дю Канталь, Кабрере, Ло, Франция (Верхнее солютре, абри де Кабрере, Кабрере).
53. Грот Ле Кюзуль-де-Мелани, Кабрере, Ло, Франция.
Вюрм
IV Дриас
I A 1 (18 500–17 800)
54. ?( – ) Куэва де Кобрантес, Сан-Мигель-де-Арас, Сантандер, Испания (Верхнее солютре, 18 540 ± 320, куэва де Альтамира).
55. – Грот дю Пеш-Мерль-1, Кабрере, Ло, Франция.
56. *( – ) [Абри дю Фурно-дю-Дьябль], Бурдей, Дордонь, Франция. Верхнее солютре, Вюрм IV Дриас I A 1.
57. * – [Aбри дю Рок-де-Сер], Сер, Шаранта, Франция. Верхнее солютре.
58. – Грот де Фье, Мьер, Ло, Франция.
59. – Гранд грот де Сен-Фрон, Домм, Дордонь, Франция.
60. * – Грот де Пижоннье, Домм, Дордонь, Франция. Финальное солют-ре или бадегуль?
61. – Грот д’Эббу-1, Валлон-Пон-д’Aрк, Ардеш, Франция (Верхнее со-лютре, Ла Рувьер, Валлон-Пон-д’Aрк; верхнее солютре, 17 900, кова дель Парпальо).
Мадлен 0 (начальный бадегуль) (18 300–17 800)
62. * Грот де Касгро-1, Трантельс, Ло и Гаронна, Франция. Мадлен 0. Вюрм IV Дриас I A 1 (Мадлен 0, 17 960 ± 350, интерстадиал Ласко, абри Фрич, Пулиньи-Сен-Пьер, Эндр, Франция).
63. ( – ) Куэва де Ла Пенья-де-Кандамо-2, Сан-Роман-де-Кандамо, Овье-до, Испания.
Интерстадиал Вюрм IV Ласко (17 800–16 500)
Мадлен I (средний и финальный бадегуль) (17 800–15 900)
64. ? Куэва Чуфин, Риклонес, Сантандер, Испания. Верхнее солютре, 17 480 ± 120, интерстадиал Ласко.
65. Куэва де Лас Чименеас, Пуэнте Вьесго, Сантандер, Испания.
66. Грот дю Рок-Пуэнтю, Кастельс, Дордонь, Франция (Мадлен I, грот дю Плакар, Вийоннер, Шаранта, Франция).
67. Абри дю Шваль, Нуаси-сюр-Эколь, Сена и Марна, Франция (Мадлен I, Ле Борегар, Немур, Сена и Марна).
68. Грот дю Портель-1, Лубенс, Арьеж, Франция (Мадлен I, грот д’Ан-лен, Монтескье-Авантес, Арьеж).
69. Грот де Сен-Сирк-1, Сен-Сирк-дю-Бюг, Дордонь, Франция (Мадлен I-II, абри дю Рок-де-Сен-Сирк, Сен-Сирк-дю-Бюг).
Мадлен II (древний мадлен) (17 400–15 500)
70. Грот де Монгодье, Монброн, Шаранта, Франция.
71. Грот де Мерьер-Сюперьер, Брюникель, Тарн и Гаронна, Франция.
72. Грот де Су-Гран-Лак-1, Мейральс, Дордонь, Франция.
73. Грот дю Папетье, Сольяк, Ло, Франция.
74. Грот де Ла Мут-2, Сирей, Лез Эйзи-де-Тейяк, Дордонь, Франция.
75. [Грот де Ла Жюстис], Бутиньи-сюр-Эссонн, Эссонн, Франция.
76. * – Грот де Ласко-1, Монтиньяк, Дордонь, Франция. Мадлен II, 17 190 ± 140, 16 100 ± 500, интерстадиал Ласко.
Вюрм IV Дриас I A 2 (16 500–15 800)
77. ( – ) Грот де Габийю-1, Сурзак, Дордонь, Франция. Мадлен II.
78. – Грот де Фон-де-Гом-1, Сирей, Лез Эйзи-де-Тейяк, Дордонь, Франция.
СТИЛЬ III ПОЗДНИЙ (16 500–15 500)
79. – Куэва дель Кастильо-1, Пуэнте Вьесго, Сантандер, Испания; 16 850 ± 200.
80. Грот де Ласко-2, Монтиньяк, Дордонь, Франция. Мадлен II, 16 100 ± 500.
81. – Грот Ла Мартен, Домм, Дордонь, Франция.
82. – Грот де Фо-Моннейер, Бузьес, Ло, Франция.
83. – Грот де Куньяк, Пейриньяк, Ло, Франция.
84. – Грот дю Пеш-Мерль-2, Кабрере, Ло, Франция.
85. Грот Марсенак, Кабрере, Ло, Франция.
86. – Грот де Ла Сюдри, Виллак, Дордонь, Франция. Мадлен II (?).
87. – Грот де л’Альден, Сессера, Эро, Франция.
88. – Капова пещера, Белорецк, Башкирия, Южный Урал, Россия. Верхний палеолит.
89. – Грот де Габийю-2, Сурзак, Дордонь, Франция. Мадлен II.
90. – Грот Каррьо-1, Бузьес, Ло, Франция.
91. – Куэва де Альтамира-1, Сантильяна-дель-Мар, Сантандер, Испания.
92. – Куэва де Ла Пилета, Серрания-де-Ронда, Малага, Испания.
93. ( – ) Куэва де Ла Аса, Рамалес-де-ла-Виктория, Сантандер, Испания.
94. ( – ) Куэва де Атапуэрка, Ибеас-де-Хуаррос, Бургос, Испания.
95. ( – ) Грот де Мервей, Рокамадур, Ло, Франция.
Интерстадиал Вюрм IV Англь (15 800–15 500)
96. Грот Ле Травер-де-Жануа, Пенн, Тарн, Франция.
97. Грот дю Дерок, Валлон-Пон-д’Арк, Ардеш, Франция.
98. Куэва де Салитре, Аханедо, Мьера, Сантандер, Испания.
99. Куэва де Коваланас, Рамалес-де-ла-Виктория, Сантандер, Испания.
100. Куэва де Ла Пасьега С-1, Пуэнте Вьесго, Сантандер, Испания.
101. Куэва дель Кастильо-2, Пуэнте Вьесго, Сантандер, Испания.
102. Куэва де Ла Пасьега А, Пуэнте Вьесго, Сантандер, Испания.
103. Куэва де Ла Грьега, Педроса, Сеговия, Испания.
104. Куэва де Альтамира-2, Сантильяна-дель-Мар, Сантандер, Испания; 15 500 ± 700.
105. Грот Ле Кюзуль-де-Бракони, Блар, Ло, Франция.
106. Грот Нанси-1, Сирей, Лез Эйзи-де-Тейяк, Дордонь, Франция. Мадлен II (?).
107. Грот де Ла Калеви, Сирей, Лез Эйзи-де-Тейяк, Дордонь, Франция.
108. Грот де Виллар, Виллар, Дордонь, Франция.
Вюрм
IV Дриас
I B (15 500–14 800)
109. ( – ) Абри-су-рош де Ла Шер-а-Кальвин, Мутье-сюр-Боэм, Шаранта, Франция. До верхнего мадлена.
110. – Грот де Бара-Бао, Ле Бюг, Дордонь, Франция.
111. – Куэва де Охо-Гваренья, Паломера, Сотоскуэва, Бургос, Испания; 15 600 ± 230.
КЛАССИЧЕСКИЙ ПЕРИОД (15 500–12 700)
СТИЛЬ IV ДРЕВНИЙ (15 500–12 700)
112. – Грот де Ласко-3, Монтиньяк, Дордонь, Франция. Мадлен II, 15 516 ± 900.
Мадлен III (средний мадлен) (16 200/15 500–14 000/13 850)
113. Грот дез Эскабас, Темен, Ло, Франция.
114. Грот де Сасизилоага, Оссюрюк, Атлантические Пиренеи, Франция.
115. Абри де Пий-Бурс, Сен-Жермен-ла-Ривьер, Жиронда, Франция. Мадлен III, 15 300 ± 410, Вюрм IV Дриас I B.
116. – Грот де Пергусе, Сен-Жери, Ло, Франция.
117. – Грот де Ла Бигурдан, Сен-Жери, Ло, Франция.
118. – Грот де Сент-Элали, Эспаньяк, Ло, Франция. Мадлен IIIa (Мадлен IIIa, 15 200 ± 300, грот дю Камбу, Эспаньяк; 15 100 ± 270, грот дю Рос-синьоль, Эспаньяк).
119. – Грот дю Бурнету, Пенсак, Ло, Франция.
120. ( – ) Грот дю Пеш-Мерль-3, Кабрере, Ло, Франция.
121. * Грот Газель, Саллель-Кабардес, Од, Франция. Мадлен IV, 15 070 ± 270, Вюрм IV Дриас I B.
Интерстадиал Вюрм IV Пребёллинг (14 800–14 300)
122. Грот де Ла Магделен, Пенн, Тарн, Франция. Мадлен.
123. Грот Кристиан, Бузьес, Ло, Франция.
124. Грот Каррьо-2, Бузьес, Ло, Франция.
125. Грот дю Мулен, Сен-Сирк-Лапопи, Ло, Франция.
126. Грот де Касгро-2, Трантельс, Ло и Гаронна, Франция.
127. Грот Эчеберри, Каму-Сииг, Атлантические Пиренеи, Франция.
128. Куэва дель Кастильо-3, Пуэнте Вьесго, Сантандер, Испания. Мадлен III.
129. Куэва де Ла Соттариса, Хибаха, Сантандер, Испания.
130. Куэва де Пенчес, Вальделакуэва, Бургос, Испания.
131. Куэва де Ла Лоха, Эль Масо, Овьедо, Испания.
132. Куэва де Льонин, Льонин, Овьедо, Испания.
133. Куэва дель Буксу, Кангас-де-Онис, Овьедо, Испания.
134. Куэва де Ла Пасьега С-2, Пуэнте Вьесго, Сантандер, Испания.
135. Грот де Ла Мюзарди, Кампань-дю-Бюг, Дордонь, Франция. (Мадлен III, абри дю Рок-де-Сен-Сирк, Сен-Сирк-дю-Бюг, Дордонь).
136. Грот Нанси-2, Сирей, Лез Эйзи-де-Тейяк, Дордонь, Франция.
137. Куэва де Алькерди, Берроберрия, Наварра, Испания.
138. Куэва де Ла Пенья-де-Кандамо-3, Сан-Роман-де-Кандамо, Овьедо, Испания.
Вюрм IV Дриас I C (14 300–13 300)
139. ( – ) Грот де Синиколе-Ко-Карбия, Каму-Сииг, Атлантические Пиренеи, Франция.
140. *( – ) Абри Реверди, Сержак, Дордонь, Франция. Мадлен IIIa. Вюрм IV Дриас I C.
141. ( – ) Куэва де Ла Пасьега В, Пуэнте Вьесго, Сантандер, Испания.
142. ( – ) Куэва де Сантимаминье, Кортесуби, Герника, Бискайя, Испания. Мадлен III.
143. ( – ) Грот Оцозелайя-Аризтойя, Сен-Мартен-д’Арберу, Атлантические Пиренеи, Франция.
144. – Грот д’Эрберюа, Истюриц, Атлантические Пиренеи, Франция.
145. – Куэва де Лос Орнос-де-ла-Пенья-2, Сан-Фелисес-де-Буэльна, Сантандер, Испания. Нижний кантабрийский мадлен.
146. * – Куэва де Тито Бустильо, Риба-де-Селья, Овьедо, Испания. Мадлен V, 14 360 ± 300, 14 250 ± 300, 14 220 ± 180. Вюрм IV Дриас I C (или Дриас II ?).
147. * – Грот де Лабастид, Лабастид, Верхние Пиренеи, Франция. Мадлен IV, 14 260 ± 440.
148. * – Абри дю Кап Блан, Марке, Дордонь, Франция. Мадлен III. Вюрм IV Дриас I C (Перигор IV).
149. * – Абри дю Рок-о-Сорсье, Англь-сюр-л’Англен, Вьенна, Франция. Мадлен IIIa, 14 160 ± 80 (или интерстадиал Англь ?).
150. * Куэва де Альтамира-3, Сантильяна-дель-Мар, Сантандер, Испания. Мадлен III, 14 000 ± 400.
Мадлен IV (средний мадлен) (15 070/14 000–13 000/12 760)
151. Грот де Сен-Сирк-2, Сен-Сирк-дю-Бюг, Дордонь, Франция (Мадлен III, начало IV, абри дю Рок-де-Сен-Сирк, Сен-Сирк-дю-Бюг; низ мадлена IV, 13 890 ± 300, Вюрм IV Дриас I C (Перигор IV), абри де Ла Мадлен, Тюрсак, Дордонь).
152. Грот де Су-Гран-Лак-2, Мейральс, Дордонь, Франция.
153. * – [Абри де Ложери-Бас], Сирей, Лез Эйзи-де-Тейяк, Дордонь, Франция. Мадлен III. Вюрм IV Дриас I C (Перигор IV) (Мадлен IIIa, 13 850 ± 160, абри де Марсей, Сирей).
154. * – Грот де Марсула, Марсула, Верхняя Гаронна, Франция. Мадлен IV.
155. Грот де Гаргас I-3, Авантиньян, Верхние Пиренеи, Франция.
156. Грот де Гаргас II, Авантиньян, Верхние Пиренеи, Франция.
157. Грот де Тибиран-2, Тибиран-Жонак, Верхние Пиренеи, Франция. Средний мадлен.
158. Грот дю Шваль, Фуа, Арьеж, Франция. Мадлен IV.
159. *( – ) Грот де Масса, Масса, Арьеж, Франция. Мадлен IV.
160. Абри де «Фурольс О», Кампом, Восточные Пиренеи, Франция.
161. * – Грот де Бедейяк, Бедейяк, Арьеж, Франция. Мадлен IV.
162. Грот де Прадьер, Бедейяк, Арьеж, Франция. Зн.
.
163. Грот дез Эглиз, Усса, Арьеж, Франция.
164. Грот де Фонтане, Орнолак-Усса-ле-Бенс, Арьеж, Франция. (Мадлен IV), 13 810 ± 740.
165. – Грот де Монтеспан, Ганти-Монтеспан, Верхняя Гаронна, Франция.
166. Грот де Сент-Колом, Сент-Колом, Атлантические Пиренеи, Франция (Мадлен IV, грот де Сен-Мишель-д’Арюди, Сен-Мишель-д’Арюди, Атлантические Пиренеи; грот д’Эспалюнг, Сен-Мишель-д’Арюди).
167. * – Грот дю Мас-д’Азиль, Ле Мас-д’Азиль, Арьеж, Франция. Мадлен IV, св. 13 640 ± 110, 13 400 ± 1000, 13 200 ± 110.
168. – Куэва де Альчерри, Айя, Гипускоа, Испания (Мадлен IV, грот де Беф, Лепюг, Верхняя Гаронна, Франция).
169. – Куэва де Лос Касарес, Риба-де-Саэлисес, Гвадалахара, Испания.
170. – Грот дю Шваль, Арси-сюр-Кюр, Йонна, Франция (Средний мадлен (IV), 13 400 ± 400, 13 390 ± 300, абри де Ла Коломбьер, Невиль-сюр-Эн, Эн, Франция).
171. – Грот де Ла Мут-3, Сирей, Лез Эйзи-де-Тейяк, Дордонь, Франция.
172. – Грот де Курназак, Сирей, Лез Эйзи-де-Тейяк, Дордонь, Франция.
173. – Грот де Фон-де-Гом-2, Сирей, Лез Эйзи-де-Тейяк, Дордонь, Франция. Средний мадлен (IV ?).
174. – Грот де Комбарель I, Сирей, Лез Эйзи-де-Тейяк, Дордонь, Франция.
175. – Грот де Комбарель II, Сирей, Лез Эйзи-де-Тейяк, Дордонь, Франция. Средний мадлен (IV ?).
176. – Грот де Бернифаль, Мейральс, Дордонь, Франция.
177. – Грот дю Бизон, Мейральс, Дордонь, Франция.
178. – Грот де Руффиньяк, Руффиньяк, Дордонь, Франция.
179. Грот де Комарк, Сирей, Лез Эйзи-де-Тейяк, Дордонь, Франция. Мадлен IV, 13 370 ± 340, 12 760 ± 200 (среднее: 12 880 ± 170), конец Вюрма IV Дриас I C.
180. * – Грот де Труа-Фрер-2, Монтескье-Авантес, Арьеж, Франция (Мадлен IV, 13 940 ± 250, 13 900 ± 120, Вюрм IV Дриас I C; 13 400 ± 120, 12 900 ± 140, грот д’Анлен, Монтескье-Авантес).
181. * – Грот дю Тюк-д’Одубер, Монтескье-Авантес, Арьеж, Франция. Мадлен IV (поздняя фаза).
182. – Куэва дель Пиндаль, Пимианго, Риба-де-Дева, Овьедо, Испания.
183. ? Куэва де Масакулос, Пимианго, Риба-де-Дева, Овьедо, Испания. Зн.
.
184. Куэва де Ла Кульяльвера, Рамалес-де-ла-Виктория, Сантандер, Испания.
185. – Грот дю Буа-дю-Канте, Эспеш, Верхние Пиренеи, Франция. Мадлен V, 13 370 ± 270, 13 060 ± 430, конец Вюрма IV Дриас I C.
186. * Куэва дель Кастильо-4, Пуэнте Вьесго, Сантандер, Испания; 12 990 ± 200.
187. * – Грот де Нио (и Ле ресо Рене Кластр), Нио, Арьеж, Франция; 12 890 ± 160.
188. * – Грот дю Портель-2, Лубенс, Арьеж, Франция. Мадлен IV (12 760 ± 170, Бёллинг).
ПОЗДНИЙ ПЕРИОД (13 300–10 200)
СТИЛЬ IV ПОЗДНИЙ (13 300–10 200)
189. – Грот де Ла Форе, Тюрсак, Дордонь, Франция.
190. * – Грот д’Истюриц, Сен-Мартен-д’Aрберу, Атлантические Пиренеи, Франция. Мадлен IV, Вюрм IV Дриас I C.
191. – Куэва де Альтамира-4, Сантильяна-дель-Мар, Сантандер, Испания (Мадлен IV, грот дю Пюи-де-Лакан, Мальмор, Коррез, Франция).
192. – Куэва де Лас Монедас, Пуэнте Вьесго, Сантандер, Испания.
Интерстадиал Вюрм IV Бёллинг (13 300–12 300)
193. Грот Бланшар, Сен-Марсель, Эндр, Франция. Средний мадлен с «челноками» (IV/V), Бёллинг, ок. 13 000.
Мадлен V (верхний мадлен) (15 800/13 000 – менее 11 800)
194. Грот д’Эббу-2, Валлон-Пон-д’Арк, Ардеш, Франция. Мадлен V, 12 980 ± 220, начало Бёллинга.
195. ? Кова Бастера, Корнейя-де-Конфлан, Восточные Пиренеи, Франция (Верхний мадлен (V ?), Ле Тру Суффлер, Фюйя, Восточные Пиренеи). Зн.
.
196. Куэва де Лас Агвас-де-Новалес, Новалес, Сантандер, Испания.
197. Куэва де Сантиан, Пуэнте Арсе, Сантандер, Испания. (Мадлен V, абри де Ложери-Бас). Зн.
.
Вюрм IV Дриас II (12 300–11 800)
198. * – Грот де Ла Мэри, Тейжа, Дордонь, Франция. Мадлен V (и VI A ?).
199. Гротта дель Кавильоне, Бальци Росси, Гримальди, Вентимилья, Империя, Италия (финальный эпиграветт, Вюрм IV Дриас II, ок. 12 200 ± 400, гротта деи Фанчулли, Бальци Росси).
200. ? Гротта ди Флорестано, Бальци Росси, Гримальди, Вентимилья, Империя, Италия. Зн.
.
201. ? Рипаро Моки, Бальци Росси, Гримальди, Вентимилья, Империя, Италия. Финальный эпиграветт. Вюрм IV. Зн.
.
202. * – Куэва де Экайн, Дева, Гипускоа, Испания. Верхний мадлен (V ?), 12 050 ± 190, Вюрм IV Дриас II.
203. – Куэва дель Пендо, Эскобедо-де-Камарго, Сантандер, Испания. Верхний мадлен (V ?), Вюрм IV Дриас II/Аллерёд, ок. 11 800.
Вюрм IV Аллерёд (11 800–10 800)
Мадлен VI (верхний и финальный мадлен) (14 500/11 800–10 200)
204. * Абри Мюрат, Рокамадур, Ло, Франция. Мадлен VI A.
205. Грот Мюрат, Рокамадур, Ло, Франция. Зн.
.
206. ? Абри Фостен, Сессак, Жиронда, Франция. Мадлен VI A, 12 370 ± 220 (?). Зн.
.
207. ? Грот де Митро, Люгассон, Жиронда, Франция. (Мадлен VI B, грот де Фонтарно, Люгассон). Зн.
.
208. – Грот дю Коломбье I, Валлон-Пон-д’Aрк и Лабастид-де-Вирак, Ар-деш, Франция. Мадлен VI B, Вюрм IV Аллерёд (позже мадлена VI B, 12 350 ± 200, 12 320 ± 600, начало Аллерёда, грот де Дез-Авенс, Валлон, Ардеш).
209. Абри дю Коломбье II, Валлон-Пон-д’Арк, Ардеш, Франция. Мадлен VI B, Вюрм IV Аллерёд.
210. * – Грот де Гуй, Гуй, Приморская Сена, Франция. Мадлен VI B.
211. ? Грот дю Ренар, Ориваль, Приморская Сена, Франция. Зн.
.
212. * Гротта Дженовесе нелл’изола ди Леванцо, Фавиньяна, Эгады, Тра-пани, Сицилия, Италия. Финальный эпиграветт, 11 180.
213. Гротта Раккьо, Ла Кала-делла-Салинелла, Трапани, Сицилия, Италия. Финальный эпиграветт.
214. ? Гротта дель Джильо, Трапани, Сицилия, Италия. Зн.
.
215. * Рипаро дель Ромито, Папасидеро, Козенца, Италия. Романелли, 10 960 ± 950.
Вюрм IV Дриас III (10 800–10 200/10 000)
216. Гротта деи Пунтали, Монте Лунго, Палермо, Сицилия, Италия. Финальный эпиграветт.
217. Рипаро делла Дза Миника, Палермо, Сицилия, Италия.
218. Гротта ди Нишеми, Монте Пеллегрино, Палермо, Сицилия, Италия.
219. Гротта делл’Аддаура I, Монте Пеллегрино, Палермо, Сицилия, Италия. Финальный эпиграветт.
220. Гротта делл’Аддаура II, Монте Пеллегрино, Палермо, Сицилия, Италия.
221. Гротта делла Дза Миника, Палермо, Сицилия, Италия.
222. ? Рипаро Арметта, Палермо, Сицилия, Италия. Зн.
.
223. ? Гротта делла Монтаньола-ди-Санта-Розалия, Термини Имересе, Палермо, Сицилия, Италия. Зн.
.
224. ? Гротта делле Джументе, Чефалу, Палермо, Сицилия, Италия. Зн.
.
225. ? Гротта ди Сан-Теодоро, Сан-Фрателло, Мессина, Сицилия, Италия. Финальный эпиграветт. Зн.
.
226. * Гротта Романелли, Терра-д’Отранто, Лечче, Италия. Романелли, 10 640 ± 100–9880 ± 100 (11 930 ± 520–9050 ± 100 ?). Вюрм IV Дриас III.
227. Грот де Ла Рок, Ла Рок, Эро, Франция. Мадлен VI, 10 200±600, Вюрм IV Дриас III.
Литература:
[8]; [52, с. 227–238]; [93, с. 68, прим. 8]; [147, с. 1451–1452]; [159]; [168]; [169]; [170]; [209]; [367]; [368]; [485]; [488]; [497]; [498]; [499]; [500]; [608]; [625]; [710]; [761]; [763, с. 69 табл. 1]; [769]; cp. [164]; [279].
Приложение 5
Хронология кайнозойских оледенений
Абсолютные даты даны «от наших дней», т. е. от 1950 г. н. э.
Возраст изотопных стадий 18О шкалы Шеклтона [692; 694] рассчитан благодаря присутствию в керне Вема 28–238 на глубине 1200 см границы палео-магнитных эпох Матуяма/Брюнес (783 ± 11 тыс. лет назад [166, с. 357]). («Минусы» – холодные стадии, «плюсы» – теплые интерстадиалы).


Литература:
[52, с. 239–242]; [197]; [234]; [244]; [406]; [452]; [453]; [455]; [456]; [486, с. 222]; [489]; [500, с. 242, табл.]; [520]; [534]; [553]; [657]; [668]; [692]; [693]; [694].
Приложение 6
Классические археологические культуры палеолита
Абсолютные даты даны «от наших дней», т. е. от 1950 г. н. э.
Африка (нижний палеолит)
Типичный олдовай (Бибер I/II, св. 2 630 000 ± 500 000 (K/Ar), 2 580 000 ± 230 000 (Zirkon) – Донау III, 1 550 000).
Развитой олдовай А (Донау II/III, 1 790 000 – Донау/Гюнц, 1 360 000).
Развитой олдовай В (Донау IV, 1 500 000 – Гюнц II, 874 000).
Развитой олдовай С (Гюнц/Миндель II, 790 000 – Гюнц/Миндель III, 607 000).
Древний ашель (Донау III/Донау III/IV, 1 600 000/1 500 000 – Гюнц II, 874 000).
Средний ашель (Гюнц II, 930 000 – Рисс I, 281 000).
Поздний ашель (Миндель I/II, 493 000 – Рисс II/III, 219 000).
Финальный ашель (Рисс III, 219 000–144 000).
Европа
Нижний палеолит (Бибер/Донау, 2 300 000 – Рисс/Вюрм,110 000)
Типичный олдовай (преашель) (Бибер/Донау, 2 300 000 – Миндель I/II, 411 000).
Развитой олдовай (протоашель) (Гюнц II, 930 000 – Миндель II, 388 000).
Древний ашель I – III (Гюнц/Миндель II, 790 000 – Миндель II, 388 000).
Средний ашель IV (Миндель II, 411 000 – Рисс II, 240 000).
Поздний ашель V (Миндель I/II, 493 000 – Рисс/Вюрм, 110 000).
Клэктон (Гюнц/Миндель II, 790 000 – Рисс I, 281 000).
Прототейяк (Гюнц/Миндель II, 790 000 – Миндель I/II, 411 000).
Древний тейяк (протошарант) (Гюнц/Миндель IV, 561 000 – Рисс II, 240 000).
Эвано [аналог верхнего палеолита] (Рисс I, 333 000 – Рисс III, 144 000).
Средний палеолит (Рисс I, 333 000 – Вюрм III А 1, 32 500)
Эпиашель (Рисс I, 333 000 – Рисс III, 144 000).
Мустье шарантской традиции типа Кина (Рисс I, 333 000 – Вюрм II/III Хенгело, 37 500).
Мустье шарантской традиции типа Ферраси (Рисс II, 260 000 – Вюрм II/ III, 34 000).
Мустье типичное (Рисс II, 260 000 – Вюрм III А 1, 32 500).
Мустье ашельской традиции типа А (Вюрм I А, 110 000 – Вюрм II В 2 (Перигор VI), 51 000).
Мустье ашельской традиции типа В (Вюрм II А, 84 000 – Вюрм II/III Хенгело, 37 500).
Мустье зубчатое (Рисс I, 333 000 – Вюрм II/III Ле Котте, 34 000).
Мустье шарантской традиции типа Таубах (Кина) (Рисс/Вюрм, 144 000 – Вюрм II, 39 000).
Микок центрально-европейский (Рисс/Вюрм, 144 000 – Вюрм II, 39 000).
Группа Альтмюль (Вюрм II/III Хенгело, 39 000–37 500).
Верхний палеолит
(Вюрм II Мурсхофд, 51 000 – Вюрм IV Дриас III, 10 200)
Перигор 0 (Вюрм II/III, 37 500–36 000).
Перигор I (шательперрон) (Вюрм II/III, 37 500 – Вюрм III А 1, 32 500).
Ориньяк 0 (экс-перигор II) (Вюрм II/III Ле Котте, 36 000 – Вюрм III А 1, 32 500).
Ориньяк I (Вюрм II Мурсхофд, 50 000 + 9000/ – 4000 (или 46 000 [567 а ]) /Вюрм III А 1
(Перигор I D), 33 000 (или 41 000 [567 а]) – Вюрм III Арси (Перигор III А), 30 600).
Ориньяк II (Вюрм III А 2 (Перигор II В – С), 32 000 – Вюрм III В (Перигор IV А), 29 600).
Ориньяк III (Вюрм III Арси (Перигор III С), 30 300 – Вюрм III В (Перигор IV), 29 000/Вюрм III С 1 А (Перигор VI А), 24 500).
Ориньяк IV (Вюрм III Кессель, 29 000–25 000/Вюрм III Тюрсак, 22 500).
Перигор IV (древний граветт) (Вюрм III В (Перигор IV С), 29 300 – Вюрм III Кессель, 25 000).
Перигор V (средний граветт) (Вюрм III B, 30 000/Вюрм III C 1, 25 000 – Вюрм III Тюрсак, 22 500).
Перигор V А (Вюрм III Кессель, 28 000/Вюрм III С 1 (Перигор VI В – С), 24 500 – Вюрм III Тюрсак, 22 500).
Перигор V В (Вюрм III Тюрсак (Перигор VII А), 23 500–23 100).
Перигор V С (Вюрм III В, 30 000/Вюрм III С 1, 25 000 – Вюрм III Тюрсак, 22 500).
Перигор VI (поздний граветт, экс-перигор III) (Вюрм III Тюрсак (Перигор VII В – С), 23 100 – Вюрм III С 2 А (Перигор VIII), 22 000).
Перигор VII (финальный граветт, экс-протомадлен) (Вюрм III С 2 А, 22 500 – Вюрм III С 2 А/III С 2 В (Перигор IX), 21 500).
Ориньяк V (Вюрм III С 2 А/III С 2 В (Перигор IX), 21 700–21 300).
Протосолютре (Вюрм III С 2 В (Перигор Х), 21 300–20 500).
Древнее солютре (Вюрм III Баньольс, 20 500 – Вюрм III С 3 А (Перигор XII), 20 100).
Среднее солютре (Вюрм III С 3 А/III С 3 В (Перигор XIII), 20 100 – Вюрм III/IV Ложери, 19 590).
Позднее солютре (Вюрм III С 3 В (Перигор XIV), 19 900 – Вюрм IV Ласко (Перигор II), 16 500).
Финальное солютре (Вюрм III/IV Ложери, 18 900 – Вюрм IV Дриас I А 1 (Перигор I), 18 300).
Мадлен 0 (начальный бадегуль) (Вюрм IV Дриас I А 1 (Перигор I), 18 300–17 800/Вюрм IV Ласко (Перигор II), < 17 800).
Мадлен I (средний и финальный бадегуль) (Вюрм IV Ласко (Перигор II), 17 800–16 500/ Вюрм IV Дриас I А 2 (Перигор III А), 15 800).
Мадлен II (древний мадлен) (Вюрм IV Ласко, 17 400/ Вюрм IV Дриас I А 2, 16 500 – Вюрм IV Англь, 15 500).
Мадлен III (средний мадлен) (Вюрм IV Дриас I А 2, 16 200/ Вюрм IV Дриас I В, 15 500 – Вюрм IV Дриас I С, 13 850).
Мадлен IV (средний мадлен) (Вюрм IV Дриас I В, 15 070/ Вюрм IV Дриас I С, 14 300 – Вюрм IV Бёллинг, 12 760).
Мадлен V (поздний мадлен) (Вюрм IV Дриас I А 2, 15 800/ Вюрм IV Бёллинг, 13 300 – Вюрм IV Аллерёд, < 11 800).
Мадлен VI (финальный мадлен) (Вюрм IV Дриас I C, 14 300/ Вюрм IV Аддерёд, 11 800 – Вюрм IV Дриас III, 10 200).
[Азиль (мезолит) (Вюрм IV Аллерёд, 11 800 – Пребореал, 8800)]
Литература:
[52, с. 243–246]; [198]; [201]; [314]; [469]; [486]; [489]; [491]; [500]; [519]; [521].
Приложение 7
Расчетная убыль постоянной Хаббла к окраинам Вселенной
Исходя из убывания плотности вещества в расширяющемся объеме, а также из падения скорости его расширения от центра к периферии пропорционально кубу расстояния, можно приблизительно представить градиент убыли постоянной Хаббла по направлению к окраинам Вселенной. Поскольку Земля относительно недалека от ее центра (302,4 млн. световых лет), расчет можно вести от величин, характерных для окрестностей Местного Сверхскопления (Скопление Дева; Местная Группа, где пребываем мы). Основной массив определений постоянной Хаббла происходит из этих окрестностей. Полученные величины колеблются в пределах 39–105 ± 11 км/с на мегапарсек [151; 428]. Однако на расстоянии от Земли, сравнимом с половиной возраста Вселенной (около 5 млрд световых лет), постоянная Хаббла падает настолько, что удаленные квазары начинают нарушать закон Хаббла (в том, что касается пропорциональности скорости удаления объекта и его расстояния от нас; очень удаленные квазары улетают медленнее ожидаемого).
По той же причине очень удаленные сверхновые звезды наводят на мысли об «отталкивающей силе», поскольку они стары, удаляются медленно, а должны бы отодвигаться быстрее – значит, в древности скорость расширения была невелика, а впоследствии она возросла под влиянием «отталкивающей силы». В действительности скорости расширения в силу естественных физических законов падают к окраинам Вселенной, где она, правда, древнее, нежели поблизости от нас.
В силу тех же естественных физических законов плотность вещества в расширяющейся Вселенной тоже убывает от центра к периферии. В окрестностях Местного Сверхскопления плотность Вселенной оценивается как Ωloc = 0,8 ± 0,5 (0,3–1,3). Плотность Великого Аттрактора превосходит эту среднюю плотность вдвое. Градиент (ступени) падения постоянной Хаббла и Ω отвечает существующим данным. Приводим его, помещая в левом столбце расстояния от центра Вселенной, а в правом – расчетные величины постоянной Хаббла и местной плотности вещества во Вселенной (Цoc). В квадратные скобки помещены вероятные начальные условия вселенского расширения.


Как можно убедиться, в ближайших окрестностях Великого Аттрактора (в радиусе ок. 33 млн световых лет) расчетная местная плотность вещества (Ωloc = 1,68) действительно вдвое превосходит плотность, свойственную окрестностям Местного Сверхскопления (Ωloc = 0,8 ± 0,5), что позволяет говорить о согласии расчетов с эмпирией, поскольку они получены формально, независимо от оценок положения дел в Великом Аттракторе. О более глубокой сердцевине Вселенной данных нет.
Следует помнить, что ближняя Вселенная в радиусе 605 млн световых лет от Великого Аттрактора (куда входит Земля и измеренная нами Вселенная) уже охвачена гравитонами Великого Аттрактора, что выражается в стремящемся к нему потоке галактик Персея-Рыб – Гидры-Кентавра. Эта гравитационная обстановка должна искажать приведенный градиент постоянной Хаббла. Сложность состоит еще и в том, что исходные для его расчета данные извлечены из реальности, искаженной гравитацией. Поэтому репрезентативны свидетельства из очень удаленных областей, в которых наблюдаются неправильные квазары, древние сверхновые и видимость «отталкивающей силы», согласующиеся с приведенным градиентом.
Объем нашей Вселенной составляет 9929 млрд кубических световых лет, и на периферии она продолжает разбегаться, тогда как центральные области уже вовлечены в схлопывание. Поэтому ожидаемые высокие темпы расширения в окрестностях центра Вселенной ныне подавлены ее наметившимся сжатием, наблюдаемым как поток Персея-Рыб, стремящийся к Великому Аттрактору. В итоге признаки анизотропности Вселенной затушеваны.
Половина вселенского объема (4964,5 млрд кубических светолет) отвечает радиусу Вселенной (10,6 млрд световых лет), который соотносится с расчетной постоянной Хаббла (ок. 15,3 км/с на мегапарсек) и средней плотностью вещества (ΩUniv = ~ 0,245). Этот результат укладывается в модель открытой Вселенной. Однако он не имеет фатального мировоззренческого значения, поскольку при любой величине Ω Вселенная не в состоянии бесконечно расширяться по квантово-механическим причинам, так как не может «истончиться» сверх размерности планковской длины.
Библиография
1. Адэр Р. К. Дефект вселенского зеркала // В мире науки. 1988, № 4. С. 16–23.
2. Алексеев В. П. География человеческих рас. М.: Мысль, 1974. 351 с.
3. Алексеев В. П. О некоторых морфологических особенностях аборигенов Америки, важных для реконструкции процесса ее заселения // Исторические судьбы американских индейцев: Проблемы индеанистики. М.: Наука, 1985. С. 24–30.
4. Ансельм А. А., Уральцев Н. Г. Аксион // Физика высоких энергий: Материалы XVII Зимней школы ЛИЯФ. Л.: ЛИЯФ, 1982. С. 81–116.
5. Арефьева И. Я., Волович И. В. Суперсимметрия: Теория Калуцы – Клейна, аномалии, суперструны // Успехи физических наук. 1985. Т. 146. Вып. 4. С. 655–681.
6. Аршавский И. А. К проблеме продолжительности жизни человека в свете данных сравнительного онтогенеза // Вопросы антропологии. 1962. Вып. 12. С. 72–91.
7. Ауэрбах Ш. Проблемы мутагенеза. М.: Мир, 1978. 463 с.
8. Бадер О. Н. Каповая пещера. Палеолитическая живопись. М.: Наука, 1965. 34 с.
9. Барбашов Б. М., Нестеренко В. В. Суперструны – новый подход к единой теории фундаментальных взаимодействий // Успехи физических наук. 1986. Т. 150. Вып. 4. С. 489–524.
10. Берг Л. С. Эмбриональные черты в строении человека // Л. С. Берг. Труды по теории эволюции: 1922–1930. Л.: Наука, 1977. С. 339–341.
11. Берндт Р. М., Берндт К. Х. Мир первых австралийцев. М.: Наука, 1981. 447 с.
12. Биология человека / Харрисон Дж., Уайнер Дж., Тэннер Дж., Барникот Н., Рейнолдс В. М.: Мир, 1979. 611 с.
13. Блюменшайн Р. Дж., Кавалло Дж. А. Гоминиды-падальщики и эволюция человека // В мире науки. 1992, № 11–12. С. 176–183.
14. Бокиш Г. Дворцы Крита // Вестник древней истории. 1974, № 4. С. 88–97.
15. Больцман Л. О механическом смысле второго начала теории теплоты // Л. Больцман. Избр. труды. М.: Наука, 1984. С. 9–29.
16. Бонгард-Левин Г. М. Древнеиндийская цивилизация. Изд. 2-е, перераб. и доп. М.: Наука, 1993. 320 с.
17. Бонч-Осмоловский Г. А. Палеолит Крыма: Грот Киик-Коба. М.; Л.: Изд. АН СССР, 1940. Вып. 1. 226 с.
18. Бутовская М. Л., Файнберг Л. А. У истоков человеческого общества: Поведенческие аспекты эволюции человека. М.: Наука, 1993. 256 с.
19. Валантэн Л. Субатомная физика (ядра и частицы): В 2 т. Т. 1. Элементарный подход. М.: Мир, 1986. 272 с.
20. Вальтер Г. Общая геоботаника. М.: Мир, 1982. 264 с.
21. Весс Ю., Беггер Дж. Суперсимметрия и супергравитация. М.: Мир, 1986. 181 с.
22. Виленкин А. Космические струны // В мире науки. 1988. № 2. С. 36–43.
23. Вишневский А. Г. Воспроизводство населения и общество: История, современность, взгляд в будущее. М.: Финансы и статистика, 1982. 287 с.
23а. Вонг К. Самый маленький человек // В мире науки. 2005. № 5. С. 42–51.
24. Высоцкий М. И. Суперсимметричные модели элементарных частиц – физика для ускорителей нового поколения // Успехи физических наук. 1985. Т. 146. Вып. 4. С. 591–636.
25. Габуния Л. К. Вымирание древних рептилий и млекопитающих. Тбилиси: Мецниереба, 1969. 234 с.
25а. Гальфрид Монмутский. История бриттов. Жизнь Мерлина. М.: Наука, 1984. 286 с.
26. Гегель Г. В. Ф. Наука логики: В 3 т. М.: Мысль, 1970–1972. Т. 1. 501 с.; Т. 2. 248 с.; Т. 3. 371 с.
27. Генденштейн Л. Э., Криве И. В. Суперсимметрия в квантовой механике // Успехи физических наук. 1985. Т. 146. Вып. 4. С. 553–590.
28. Гешвинд Н. Специализация человеческого мозга // Мозг. М.: Мир, 1982. С. 219–239.
29. Гладких М. И., Корниец Н. Л., Софер О. Жилища из костей мамонта на Русской равнине // В мире науки. 1985. № 1. С. 68–74.
30. Горенстейн Д. Конечные простые группы: Введение в их классификацию. М.: Мир, 1985. 352 с.
31. Горенстейн Д. Грандиозная теорема // В мире науки. 1986. № 2. С. 62–74.
32. Грин М. Теории суперструн в реальном мире // Успехи физических наук. 1986. Т. 150. Вып. 4. С. 577–579.
33. Грин М., Шварц Дж., Виттен Э. Теория суперструн: В 2 т. Т. 1: Введение. Т. 2: Петлевые амплитуды, аномалии и феноменология. М.: Мир, 1990. Т. 1. 518 с.; Т. 2. 656 с.
34. Гут А.Г., Стейнхардт П. Дж. Раздувающаяся Вселенная // В мире науки. 1984. № 7. С. 56–69.
35. Давиташвили Л. Ш. Причины вымирания организмов. М.: Наука, 1969. 440 с.
36. Джохансон Д., Иди М. Люси: Истоки рода человеческого. М.: Мир, 1984. 295 с.
37. Диксон Д. Динозавры: Иллюстрированная энциклопедия. М.: Московский клуб, 1994. 144 с.
38. Друщиц В. В. Палеонтология беспозвоночных. М.: МГУ, 1974. 528 с.
39. Дьюсбери Д. Поведение животных: Сравнительные аспекты. М.: Мир, 1981. 480 с.
40. Дьяконов И. М. Научные представления на Древнем Востоке (Шумер, Вавилония, Передняя Азия) // Очерки истории естественнонаучных знаний в древности. М.: Наука, 1982. С. 59–119.
41. Зельдович Я. Б. Теория крупномасштабной структуры Вселенной // Крупномасштабная структура Вселенной. М.: Мир, 1981. С. 452–465.
42. Зимы нашей планеты: Земля подо льдом. М.: Мир, 1982. 336 с.
43. Зубов А. А. Неандертальцы: что известно о них современной науке? // Этнографическое обозрение. 1999. № 3. С. 67–83.
44. Зубов А. А. Новая интерпретация роли «Гейдельбергского человека» в эволюции рода Homo // Этнографическое обозрение. 2001. № 1. С. 91–111.
45. История Древнего Востока: Зарождение древнейших классовых обществ и первые очаги рабовладельческой цивилизации. Ч. I. Месопотамия. М.: Наука, 1983. 534 с.
46. История Древнего Востока: Зарождение древнейших классовых обществ и первые очаги рабовладельческой цивилизации. Ч. II. Передняя Азия. Египет. М.: Наука, 1988. 623 с.
47. Кабо В. Р. Первобытное общество и природа // Общество и природа: Исторические этапы и формы взаимодействия. М.: Наука, 1981. С. 149–158.
48. Казаков Д. И. Суперструны, или За пределами стандартных представлений // Успехи физических наук. 1986. Т. 150. Вып. 4. С. 561–575.
49. Каландадзе А. Н. Разыскания по археологии доантичной Грузии: Автореф. дисс. … д-ра истор. наук. Тбилиси: Ин-т истории, археологии и этнографии им. И. А. Джавахишвили АН Груз.ССР, 1969. 72 с.
50. Кларк Дж. Д. Доисторическая Африка. М.: Наука, 1977. 264 с.
51. Клягин Н. В. От доистории к истории: Палеосоциология и социальная философия. М.: Наука, 1992. 192 с.
52. Клягин Н. В. Происхождение цивилизации (социально-философский аспект). М.: ИФРАН, 1996. 252 с.
53. Клягин Н. В. Человек в истории. М.: ИФРАН, 1999. 238 с.
54. Ковалевский А. П. Книга Ахмеда Ибн-Фадлана о его путешествии на Волгу в 921–922 гг. Статьи, переводы и комментарии. Харьков: Изд-во ХГУ, 1956. 347 с.
55. Козинцев А. Г. Проблема грани между человеком и животными: антропологический аспект // Этнографическое обозрение. 1997. № 4. С. 96–104.
56. Колосов Ю. Г. Аккайская мустьерская культура. Киев: Наукова думка, 1986. 224 с.
57. Коростовцев М. А. Религия Древнего Египта. СПб.: Журнал «Нева»; «Летний Сад», 2000. 464 с.
58. Крушинский Л. В. Биологические основы рассудочной деятельности: Эволюционный и физиолого-генетический аспекты поведения. М.: Изд-во МГУ, 1977. 272 с.
59. Лавик-Гудолл Дж. ван. В тени человека. М.: Мир, 1974. 207 с.
60. Ларичев В. Е. Мамонт в искусстве поселения Малая Сыя и опыт реконструкции представлений верхнепалеолитического человека Сибири о возникновении Вселенной // Звери в камне: (Первобытное искусство). Новосибирск: Наука, 1980. С. 159–198.
61. Левитин К. Ископаемые концепции // Знание – сила. 1979. № 9. С. 39–43.
62. Легг Э. Дж., Роули-Конуи П. А. Массовая охота на джейранов в каменном веке на территории Сирии // В мире науки. 1987. № 10. С. 62–70.
63. Лем С. Звездные дневники Ийона Тихого. Путешествие двадцать восьмое // С. Лем. Собр. соч.: В 10 т. М.: Текст, 1994. Т. 7. С. 266–284.
64. Ленин В. И. Материализм и эмпириокритицизм. Критические заметки об одной реакционной философии // В. И. Ленин. Полн. собр. соч. 5-е изд. М.: Политиздат, 1973. Т. 18. 525 с.
65. Ле Пишон К., Франшто Ж., Боннин Ж. Тектоника плит. М.: Мир, 1977. 288 с.
66. Линдблад Я. Человек – ты, я и первозданный. М.: Прогресс, 1991. 264 с.
67. Линде А. Д. Новый сценарий раздувающейся Вселенной: возможное решение проблем горизонта, плоскости, однородности, изотропии и реликтовых монополей // Квантовая гравитация: Труды второго семинара «Квантовая теория гравитации». М., 13–15 окт. 1981. М.: ИЯИ АН СССР, 1982. С. 106–112.
68. Линде А. Д. Раздувающаяся Вселенная // Успехи физических наук. 1984. Т. 144. Вып. 2. С. 177–214.
69. Линден Ю. Обезьяны, человек и язык. М.: Мир, 1981. 272 с.
70. Лосев К. С., Подгорная Л. И., Ушаков С. А. Палеогляциология Антарктиды (с позиции тектоники литосферных плит). // Антарктика: Доклады комиссии. М.: Наука, 1980. Вып. 19. С. 16–22.
71. Мари Дж. Д. Отчего у леопарда пятна на шкуре // В мире науки. 1988. ¹ 5. С. 36–54.
72. Маркс К., Энгельс Ф. Немецкая идеология. Критика новейшей немецкой философии в лице ее представителей Фейербаха, Б. Бауэра и Штирнера и немецкого социализма в лице его различных пророков. // К. Маркс, Ф. Энгельс. Соч. 2-е изд. М.: Политиздат, 1955. Т. 3. С. 7–544.
73. Меннинг О. Поведение животных: Вводный курс. М.: Мир, 1982. 360 с.
74. Монин А. С., Шишков Ю. А. История климата. Л.: Гидрометеоиздат, 1979. 407 с.
75. Мочанов Ю. А. Древнейший палеолит Диринга и проблема внетропической прародины человечества. Новосибирск: Наука, 1992. 253 с.
76. Никифоров А. С. Этюды о разуме. М.: Советская Россия, 1981. 208 с.
77. Новая глобальная тектоника (тектоника плит): Сб. ст. М.: Мир, 1974. 471 с.
78. Новиков И. Д., Полнарев А. Г., Розенталь И. Л. Численные значения фундаментальных постоянных и антропный принцип // Проблема поиска жизни во Вселенной: Труды Таллиннского симпозиума. М.: Наука, 1986. С. 36–40.
79. Новиков И. Д., Фролов В. П. Физика черных дыр. М.: Наука, 1986. 328 с.
80. Одум Ю. Основы экологии. М.: Мир, 1975. 740 с.
81. Окладников А. П. Утро искусства. Л.: Искусство, 1967. 135 с.
82. Оно С. Генетические механизмы прогрессивной эволюции. М.: Мир, 1973. 227 с.
83. Основы палеонтологии. Справочник для палеонтологов и геологов СССР: В 15 т. T. 5. Моллюски – головоногие. I: Наутилоидеи, эндоцератоидеи, актино-цератоидеи, бактритоидеи, аммоноидеи (агониатиты, гониатиты, климении). М.: Изд. АН СССР, 1962. 438 с.
84. Основы палеонтологии. Т. 6. Моллюски – головоногие. II: Аммоноидеи (цератиты и аммониты). Внутреннераковинные. Приложение: кониконхии. М.: Госгеолтехиздат, 1958. 359 с.
85. Основы палеонтологии. T. 8. Членистоногие. Трилобитообразные и ракообразные. М.: Госгеолтехиздат, 1960. 515 с.
86. Основы палеонтологии. T. 11. Бесчелюстные, рыбы. М.: Наука, 1964. 522 с.
87. Основы палеонтологии. T. 12. Земноводные, пресмыкающиеся и птицы. М.: Наука, 1964. 722 с.
88. Основы палеонтологии. T. 15. Голосеменные и покрытосеменные. М.: Гос-геолтехиздат, 1963. 743 с.
89. Пидопличео И. Г. Позднепалеолитические жилища из костей мамонта на Украине. Киев: Наукова думка, 1969. 163 с.
90. Поршнев Б. Ф. О начале человеческой истории: Проблемы палеопсихоло-гии. М.: Мысль, 1974. 487 с.
91. Путешествие Ахмеда ибн-Фадлана на реку Итиль. Казань: Татарское книжное издательство, 1999. 112 с.
92. Спицын В. А. К проблеме происхождения и дифференциации человеческих рас в пространстве // Вопросы антропологии. 1977. Вып. 54. С. 3–22.
93. Столяр А. Д. Происхождение изобразительного искусства. М.: Искусство, 1985. 298 с.
94. Сытник А. С. Гравированный рисунок на кости с мустьерской стоянки под Тернополем // Пластика и рисунки древних культур. (Первобытное искусство). Новосибирск: Наука, 1983. С. 39–46.
95. Тайлор Э. Б. Первобытная культура. М.: Политиздат, 1989. 573 с.
96. Тарлинг Д., Тарлинг М. Движущиеся материки. М.: Мир, 1973. 104 с.
97. Тимофеев-Ресовский Н. В., Воронцов Н. Н., Яблоков А. В. Краткий очерк теории эволюции: 2-е изд., перераб. М.: Наука, 1977. 301 с.
98. Токарев С. А. Ранние формы религии и их развитие // С. А. Токарев. Ранние формы религии. М.: Политиздат, 1990. С. 13–374.
99. Тот Н. Первая технология // В мире науки. 1987. № 6. С. 80–90.
100. Уилсон А. К., Канн Р. Л. Недавнее африканское происхождение людей // В мире науки. 1992. № 6. С. 8–14.
101. Уэст П. Введение в суперсимметрию и супергравитацию. М.: Мир, 1989. 328 с.
102. Формозов А. А. Очерки по первобытному искусству: Наскальные изображения и каменные изваяния эпохи камня и бронзы на территории СССР. (Материалы и исследования по археологии СССР. № 165). М.: Наука, 1969. 255 с.
103. Фоули Р. Еще один неповторимый вид: Экологические аспекты эволюции человека. М.: Мир, 1990. 368 с.
104. Фрагменты ранних греческих философов. Ч. 1. От эпических теокосмого-ний до возникновения атомистики. М.: Наука, 1989. 576 с.
105. Фрадкин Э. Е. К вопросу о времени возникновения скульптуры из кремня (в связи с проблемой происхождения палеолитического искусства) // Краткое содержание докладов сессии Института этнографии АН СССР, посвященной столетию создания первого Академического этнографо-антропологического центра. Л.: Наука, 1980. С. 90–92.
106. Фрадкин Э. Е. О нескольких ранних произведениях изобразительного искусства // Советская этнография. 1985. № 2. С. 122–127.
107. Фриш Р. Э. Полнота и плодовитость // В мире науки. 1988. № 5. С. 56–64.
108. Фролов Б. А. Числа в графике палеолита. Новосибирск: Наука, 1974. 239 с.
109. Фрэзер Дж. Дж. Золотая ветвь: Исследование магии и религии. 2-е изд. М.: Политиздат, 1983. 703 с.
110. Хайнд Р. Поведение животных: Синтез этологии и сравнительной психологии. М.: Мир, 1975. 855 с.
111. Хить Г. Л., Кейта Б. Дерматоглифическая дивергенция основных расовых ветвей человечества // Расы и народы. Современные этнические и расовые проблемы. Ежегодник. М.: Наука, 1981. Вып. 11. С. 25–39.
112. Черныш А. П. О времени возникновения палеолитического искусства в связи с исследованиями 1976 г. стоянки Молодова I // У истоков творчества: (Первобытное искусство). Новосибирск: Наука, 1978. С. 18–23.
113. Черныш А. П. Многослойная палеолитическая стоянка Молодова I // Молодова I: Уникальное мустьерское поселение на Среднем Днестре. М.: Наука, 1982. С. 6–102.
114. Шаллер Дж. Б. Год под знаком гориллы. М.: Мысль, 1968. 246 с.
115. Шапиро Дж. А. Бактерии как многоклеточные организмы // В мире науки. 1988. № 8. С. 46–54.
116. Шеннон К. Математическая теория связи // К. Шеннон. Работы по теории информации и кибернетике: Сб. ст. М.: ИЛ, 1963. С. 243–332.
117. Шовен Р. Поведение животных. М.: Мир, 1972. 487 с.
118. Шторх Г. Млекопитающие островной Европы // В мире науки. 1992. № 4. С. 32–38.
119. Щетенко А. Я. Первобытный Индостан. Л.: Наука, 1979. 275 с.
120. Эйнштейн А. Собрание научных трудов: В 4 т. М.: Наука, 1965–1967. Т. 1. Работы по теории относительности 1905–1920 гг., 1965. 700 с. Т. 2. Работы по теории относительности 1921–1955 гг. 1966. 878 с.
121. Энтони Д., Телегин Д. Я., Браун Д. Зарождение верховой езды // В мире науки. 1992. № 2. С. 36–42.
122. Энтони С. Суперструны: всеобъемлющая теория? // Успехи физических наук. 1986. Т. 150. Вып. 4. С. 579–583.
123. Эфроимсон В. П. Эволюционно-генетическое происхождение альтруистических эмоций // Научная мысль. Вестник Агентства печати «Новости» (АПН). М.: АПН, 1968. С. 27–42.
124. Яковлев В. Н. Функциональная эволюция скелета рыб // Палеонтологический журнал. 1966. № 3. С. 3–12.
125. Abazov V. M. et al. A precision measurement of the mass of the top quark // Nature. 2004. V. 429, № 6992. P. 638–642.
126. Abbate E., Albianelli A., Azzaroli A., Benvenuti M., Tesfamariam B., Bruni P., Cipriani N., Clarke R. J., Ficcarelli G., Macchiarelli R., Napoleone G., Papini M., Rook L., Sagri M., Tecle T. M., Torre D., Villa I. A one-million-year-old Homo cranium from the Danakil (Afar) Depression of Eritrea // Nature. 1998. V. 393, № 6684. P. 458–460.
127. Ackerman S. European Prehistory Gets Even Older // Science. 1989. V. 246, № 4926. P. 28–30.
128. Ahern J. C. M., Karavanic´
I., Paunovic´
M., Jankovic´
I., Smith F. H. New discoveries and interpretations of hominid fossils and artifacts from Vindija Cave, Croatia // Journal of Human Evolutio № 2004. V. 46, № 1. P. 27–67.
128à. Ahlberg E., Clack J. A., Blom H. The axial skeleton of the Devonian tetrapod Ichthyostega // Nature. 2005. V.437, № 7055. P. 137–140.
129. Ahlberg P. E. Elginerpeton pancheni and the earliest tetrapode clade // Nature. 1995. V. 373, № 6513. P. 420–425.
130. Ahlberg P. E., Milner A. R. The origin and early diversification of tetrapods // Nature. 1994. V. 368, № 6471. P. 507–514.
131. Aiello L. C., Collard M. Our newest oldest ancestor? // Nature. 2001. V. 410, № 6828. P. 526–527.
132. Albrecht A., Brandenberger R., Turok N. Cosmic strings and cosmic structure // New scientist. 1987. V. 114, № 1556. P. 40–44.
133. Alcock C., Allsman R.A., Alves D. R., Axelrod T. S., Becker A. C., Bennett D. P., Cook K. H., Drake A. J., Freeman K. C., Geha M., Griest K., Keller S. C., Lehrer M. J., Marshall S. L., Minnitt D., Nelson C. A., Peterson B. A., Popowski P., Pratt M. R., Quinn P. J., Stubbs C.W., Sutherland W., Tomaney A. B., Vandehei T., Welch D. Direct detection of a microlens in the Milky Way // Nature. 2001. V. 414, № 6864. P. 617–619.
134. Allen D. A., Norris R. P., Staveley-Smith L., Meadows V.S., Roche P. F. A super-cluster of IRAS galaxies behind the Great Attractor // Nature. 1990. V. 343, № 6253. P. 45–46.
135. Allen J., Gosden C., Jones R., White J. P. Pleistocene dates for the human occupation of New Ireland, nothern Melanesia // Nature. 1988. V. 331, № 6158. P. 707–709.
136. Alonso P. D., Milner A. C., Ketcham R. A., Cookson M. J., Rowe T. B. The avian nature of the brain and inner ear of Archaeopteryx // Nature. 2004. V. 430, № 7000. P. 666–669.
137. Alvarez L. W., Alvarez W., Asaro F., Michel H. V. Extraterrestrial Cause for the Cretaceous-Tertiary Extinction // Science. 1980. V. 208, № 4448. P. 1095–1108.
138. Alvarez W., Asaro F., Montanari A. Iridium Profile for 10 Million Years Across the Cretaceous-Tertiary Boundary at Gubbio (Italy) // Science. 1990. V. 250, № 4988. P. 1700–1702.
139. Ambrose S. H., De Niro M. J. Reconstruction of African human diet using bone collagen carbon and nitrogen isotope ratios // Nature. 1986. V. 319, № 6051. P. 321–324.
140. Amelino-Camella G. A phenomenological description of space-time noise in quantum gravity // Nature. 2001. V. 410, № 6832. P. 1065–1067.
141. Anderson A. Early Bird Threatens Archaeopteryx’s Perch // Science. 1991. V. 253, № 5015. P. 35.
142. Anderson I. Diggers at Dinosaur Cove // New scientist. 1993. V. 137, № 1860. P. 28–32.
143. Andrews P. Improved timing of hominoid evolution with a DNA clock // Nature. 1985. V. 314, № 6011. P. 498–499.
144. Andrews P. Evolution and environment in the Hominoidea // Nature. 1992. V. 360, № 6405. P. 641–646.
145. Andrews P. Ecological apes and ancestors // Nature. 1995. V. 376, № 6541. P. 555–556.
146. Antoniou I. E. Time asymmetry // Nature. 1989. V. 338, № 6212. P. 210–211.
147. Appenzeller T. Art: Evolution or Revolution? // Science. 1998. V. 282, № 5393. P. 1451–1454.
148. Appenzeller T. T. Rex Was Fierce, Yes, But Feathered, Too // Science. 1999. V. 285, № 5436. P. 2052–2053.
149. Archer M., Flannery T. F., Ritchie A., Molnar R. E. First Mesozoic mammal from Australia – an early Cretaceous monotreme // Nature. 1985. V. 318, № 6044. P. 363–366.
150. Arensburg B., Tilier A. M., Vandermeersch B., Duday H., Schepartz L.A., Rak Y. A. Middle Palaeolithic human hyoid bone // Nature. 1989. V. 338, № 6218. P. 758–760.
151. Arnett W. D., Branch D., Wheeler J. C. Hubble’s constant and exploding carbon-oxygen white dwarf models for Type I supernovae // Nature. 1985. V. 314, № 6009. P. 337–338.
152. Arsuaga J-L., Martinez I., Gracia A., Carretero J-M., Carbonell E. Three new human sculls from the Sima de los Huesos Middle Pleistocene site in Sierra de Atapuerca, Spain // Nature. 1993. V. 362, № 6420. P. 534–537.
153. Asfaw B., Beyene Y., Suwa G., Walter R. C., White T.D., WoldeGabriel G., Yemane T. The earliest Acheulean from Konso-Gardula // Nature. 1992. V. 360, № 6404. P. 732–735.
154. Asfaw B., Gilbert W. H., Beyene Y., Hart W. K., Renne P. R., WoldeGabriel G., Vrba E. S., White T. D. Remains of Homo erectus from Bouri, Middle Awash, Ethiopia // Nature. 2002. V. 416, № 6878. P. 317–320.
155. Asfaw B., White T., Lovejoy O., Latimer B., Simpson S., Suwa G. Australopithecus garhi: A New Species of Early Hominid from Ethiopia // Science. 1999. V. 284, № 5414. P. 629–635.
156. Awramik S. M. New fossil finds in old rocks // Nature. 1986. V. 319, № 6053. P. 446–447.
157. Ayala F. J. Gene Lineages and Human Evolution // Science. 1996, V. 272, № 5266. P. 1363–1364.
158. Bahcall J. N., Glashow S.L. Upper limit on the mass of the electron neutrino // Nature. 1987. V. 326, № 6112. P. 476–477.
159. Bahn P. G. Ice Age drawings on open rock faces in the Pyrenees // Nature. 1985. V. 313, № 6003. P. 530–531.
160. Bahn P. G. Excavation of a palaeolithic plank from Japan // Nature. 1987. V. 329, № 6135. P. 110.
161. Bahn P. G. Early teething troubles // Nature. 1989. V. 337, № 6209. P. 693.
162. Bahn P. G. Age and the female form // Nature. 1989. V. 342, № 6248. P. 345–346.
163. Bahn P. G. Eating people is wrong // Nature. 1990. V. 348, № 6300. P. 395.
164. Bahn P. G. Charting the past // Nature. 2004. V. 432, № 7020. P. 952.
165. Bak P., Flyvbjerg H., Sneppen K. Can we model Darwin? // New scientist. 1994. V. 141, № 1916. P. 36–39.
166. Baksi A. K., Hsu V., McWilliams M. O., Farrar E. 40Ar/39Ar Dating of the Brunhes-Matuyama Geomagnetic Field Reversal // Science. 1992. V. 256, № 5055. P. 356–357.
167. Ball P. Cosmic beakers // New scientist. 1996. V. 151, № 2048. P. 46–50.
168. Balter M. New Light on the Oldest Art // Science. 1999. V. 283, № 5404. P. 920–922.
169. Balter M. Restorers Reveal 28,000-Year-Old Artworks // Science. 1999. V. 283, № 5409. P. 1835.
170. Balter M. Stone Age Artists – or Art Lovers – Unmasked? // Science. 2001. V. 294, № 5540. P. 31.
171. Bandi H.-G., Maringer J. Kunst der Eiszeit. Levantekunst. Arktische Kunst. Basel: Holbein-Verlag, 1952. 168 S.
172. Barreto C., Albrecht R. M., Bjorling D. E., Horner J. R., Wilsman N. J. Evidence of the Growth Plate and the Growth of Long Bones in Juvenile Dinosaurs // Science. 1993. V. 262, № 5142. P. 2020–2023.
173. Barrick R. E., Showers W. J. Thermophysiology of Tyrannosaurus rex: Evidence from Oxygen Isotopes // Science. 1994. V. 265, № 5169. P. 222–224.
174. Bartel N., Rogers A. E. E., Shapiro I. I., Gorenstein M. V., Gwinn C. R., Mar-caide J. M., Weiler K. W. Hubble’s constant determined using very-long baseline interferometry of a supernova // Nature. 1985. V. 318, № 6041. P. 25–30.
175. Basu A. R., Petaev M. I., Poreda R. J., Jacobson S. B., Becker L. Chondritic Meteorite Fragments Associated with the Permian-Triassic Boundary in Antarctica // Science. 2003. V. 302, № 5649. P. 1388–1392.
176. Battaner E., Garrido J. L., Membrado M., Florido E. Magnetic fields as an alternative explanation for the rotation curves of spiral galaxies // Nature. 1992. V. 360, № 6405. P. 652–653.
177. Batuski D. J., Burns J. O. A possible 300 megaparsec filamen of clusters of galaxies in Perseus-Pegasus // The Astrophysical Journal. 1985. V. 299, № 1, part 1. P. 5–14.
178. Beard K. C., Krishtalka L., Stucky R. K. First skulls of the Early Eocene primate Shoshonius cooperi and the anthropoid – tarsier dichotomy // Nature. 1991. V. 349, № 6304. P. 64–67.
179. Becker L., Bada J. L., Winans R. E., Hunt J. E., Bunch T. E., French B. M. Fullerenes in the 1.85-Billion-Year-Old Sudbury Impact Structure // Science. 1994. V. 265, № 5172. P. 642–645.
180. Becker L., Poreda R. J., Hunt A. G., Bunch T. E., Rampino M. Impact Event at the Permian-Triassic Boundary: Evidence from Extraterrestrial Noble Gases in Fulle-renes // Science. 2001. V. 291, № 5508. P. 1530–1533.
181. Bekker A., Holland H. D., Wang P.-L., Rumble III D., Stein H. J., Hannah J. L., Coetzee L. L., Beukes N. J. Dating the rise of atmospheric oxygen // Nature. 2004. V. 427, № 6970. P. 117–120.
182. Benton M. At the court of the Red Queen // New scientist. 1985. V. 108, № 1484. P. 52–54.
183. Benton M. J. First marsupial fossil from Asia // Nature. 1985. V. 318, № 6044. P. 313.
184. Benton M. J. Diversification and Extinction in the History of Life // Science. 1995. V. 268, № 5207. P. 52–58.
185. Benton M. J., Tverdokhlebov V. P., Surkov M. V. Ecosystem remodelling among vertebrates at the Permian – Triassic boundary in Russia // Nature. 2004. V. 432, № 7013. P. 97–100.
186. Bergh S. van den. The Age and Size of the Universe // Science. 1992. V. 258, № 5081. P. 421–424.
187. Berman D. S., Reisz R. R., Scott D., Henrici A. C., Sumida S. S., Martens T. Early Permian Bipedal Reptile // Science. 2000. V. 290, № 5493. P. 969–972.
188. Bermú
dez de Castro J. M., Arsuaga J. L., Carbonell E., Rosas A., Martinez I., Mosquera M. A Hominid from the Lower Pleistocene of Atapuerca, Spain: Possible Ancestor to Neandertals and Modern Humans // Science. 1997. V. 276, № 5317. P. 1392–1395.
189. Bernstein M. P., Dworkin J. P., Sandford S. A., Cooper G. W., Allamandola L. J. Racemic amino acids from the ultraviolet photolysis of interstellar ice analogues // Nature. 2002. V. 416, № 6879. P. 401–403.
190. Bishoff J. L., Julia R., Mora R. Uranium series dating of the Mousterian occupation at Abric Romani, Spain // Nature. 1988. V. 332, № 6159. P. 68–70.
191. Bishop W. W., Pickford M. H. L. Geology, fauna and palaeoenvironments of the Ngorora Formation, Kenya Rift Valley // Nature. 1975. V. 254, № 5497. P. 185–192.
192. Blum J. D., Page Chamberlain C., Hingston M. P., Koeberl C., Marin L. E., Schuraytz B. C., Sharpton V. L. Isotopic comparison of K/T boundary impact glass with melt rock from the Chicxulub and Manson impact structures // Nature. 1993. V. 364, № 6435. P. 325–327.
193. Blumenschine R. J., Peters C. R., Masao F. T., Clarke R. J., Deino A. L., Hay R. L., Swisher C. C., Stanistreet I. G., Ashley G. L., McHenry L. J., Sikes N. E., Merwe N. J. van der, Tactikos J. C., Cushing A. E., Deocampo D. M., Njau J. K., Ebert J. I. Late Pliocene Homo and Hominid Land Use from Western Olduvai Gorge, Tanzania // Science. 2003. V. 299, № 5610. P. 1217–1221.
194. Boesch C. First hunters of the forest // New scientist. 1990. V. 126, № 1717. P. 38–41.
195. Bohor B. F., Modreski P. J., Foord E. E. Shocked Quartz in the Cretaceous-Tertiary Boundary Clays: Evidence for a Global Distribution // Science. 1987. V. 236, № 4802. P. 705–709.
196. Bonis L. de, Bouvrain G., Geraadis D., Koufos G. New hominid skull màterial from the late Miocene of Macedonia in Nothern Greece // Nature. 1990. V. 345, № 6277. P. 712–714.
197. Bonnefille R. Evidence for a cooler and drier climate in the Ethiopian uplands towards 2,5 Myr ago // Nature. 1983. V. 303, № 5917. P. 487–491.
198. Bordes F. Essai de Classification des industries «moustériennes» // Bulletin de la Société Préhistorique française. 1953. T. 50. № 7–8. P. 457–466.
199. Bottomley R., Grieve R., York D., Masaitis V. The age of the Popigai impact event and its relation to events at the Eocene/Oligocene boundary // Nature. 1997. V. 388, № 6640. P. 365–368.
200. Boughn S., Crittenden R. A correlation between the cosmic microwave background and large-scale structure in the Universe // Nature. 2004. V. 427, № 6969. P. 45–47.
201. Bourdier F. Préhistoire de France. Paris: Flammarion, 1967. 412 P.
202. Bowcock A. M., Ruiz-Linares A., Tomfohrde J., Minch E., Kidd J. R., Cavalli-Sforza L. L. High resolution of human evolutionary trees with polymorphic microsatellites // Nature. 1994. V. 368, № 6470. P. 455–457.
203. Bowring S. A., Grotzinger J. P., Isachsen C. E., Knoll A. H., Pelechaty S. M., Kolo-sov P. Calibrating Rates of Early Cambrian Evolution // Science. 1993. V. 261, № 5126. P. 1293–1298.
204. Bowring S. A., Housh T. The Earth’s Early Evolution // Science. 1995. V. 269, № 5230. P. 1535–1540.
205. Bradshaw J. The impotance of being right– (or left-) handed // New scientist. 1985. V. 105, № 1440. P. 46–48.
206. Brain C. K., Sillen A. Evidence from the Swartkrans cave for earliest use of fire // Nature. 1988. V. 336, № 6198. P. 464–466.
207. Bramble D. M., Lieberman D.E. Endurance running and the evolution of Homo // Nature. 2004. V. 432, № 7015. P. 345–352.
208. Brandes J. A., Boctor N. Z., Cody G. D., Cooper B. A., Hazen R. M., Yoder Jr H. S. Abiotic nitrogen reduction on the early Earth // Nature. 1998. V. 395, № 6700. P. 365–367.
209. Breuil H. Quatre cents siècles d’art pariétal. Les cavernes ornées de l’Age du renne. Montignac (Dordogne): Centre d’études et de documentation préhistoriques, 1952. 417 P.
210. Briggs D. E. G. Conodonts: A Major Extinct Group Added to the Vertebrates // Science. 1992. V. 256, № 5061. P. 1285–1286.
211. Briggs D. E. G., Fortey R. A. The early Radiation and Relationships of the Major Arthropod Groups // Science. 1989. V. 246, № 4927. P. 241–243.
212. Brochier C., Philippe H. A non-hyperthermophilic ancestor for Bacteria // Nature. 2002. V. 417, № 6886. P. 244.
213. Brocks J. J., Logan G. A., Buick R., Summons R.E. Archean Molecular Fossils and Early Rise of Eukaryotes // Science. 1999. V. 285, № 5430. P. 1033–1036.
214. Brooks A.S., Wood B. The Chinese side of the story // Nature. 1990. V. 344, № 6264. P. 288–289.
215. Brouwers E. M., Clemens W. A., Spicer R. A., Ager T. A., Carter L. D., Sliter W. V. Dinosaurs on the North Slope, Alaska: High Latitude, Latest Cretaceous Environments // Science. 1987. V. 237, № 4822. P. 1608–1610.
216. Brown F., Harris J., Leakey R., Walker A. Early Homo erectus skeleton from west Lake Turkana, Kenya // Nature. 1985. V. 316, № 6031. P. 788–792.
217. Brown F. H., Feibel C. S. Stratigraphical notes on the Okote Tuff Complex at Koobi Fora, Kenya // Nature. 1985. V. 316, № 6031. P. 794–797.
218. Brown P., Sutikna T., Morwood M.J., Soejono R. P., Jatmiko, Saptomo E. W., Due R. A. A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia // Nature. 2004. V. 431, № 7012. P. 1055–1061.
219. Brunet M., Beauvilain A., Coppens Y., Heintz E., Moutaye A. H. E., Pilbeam D. The first australopithecine 2,500 kilometres west of the Rift Valley (Chad) // Nature. 1995. V. 378, № 6554. P. 273–275.
220. Brunet M., Guy F., Pilbeam D., Lieberman D. E., Likius A., Mackaye H. T., Ponce de Leô
n M. S., Zollikofer C. P. E., Vignaud P. New material of the earliest hominid from the Upper Miocene of Chad // Nature. 2005. V. 434, № 7034. P. 752–755.
221. Brunet M., Guy F., Pilbeam D., Mackaye H. T., Likius A., Ahounta D., Beauvilain A., Blondel C., Bocherens H., Boisserie L.-R., De Bonis L., Coppens Y., Dejax J., Denys C., Duringer P., Eisenmann V., Fanone G., Fronty P., Geraads D., Lehmann T., Lihoreau F., Louchart A., Mahamat A., Merceron G., Mouchelin G., Otero O., Campomanes P. P., Ponce De Leô
n M., Rage J.-C., Sapanet M., Schuster M., Sudre J., Tassy P., Valentin X., Vignaud P., Viriot L., Zazzo A., Zollikofer C. A new hominid from the Upper Miocene of Chad, Central Africa // Nature. 2002. V. 418, № 6894. P. 145–151.
222. Bryant L. J., Clemens W. A., Hutchison J. H. Cretaceous – Tertiary Dinosaur Extinction // Science. 1986. V. 234, № 4781. P. 1172–1173.
223. Buick R., Thornett J. R., McNaughton N. J., Smith J. B., Barley M. E., Savage M. Record of emergent continental crust ~ 3.5 billion years ago in the Pilbara craton of Australia // Nature. 1995. V. 375, № 6532. P. 574–577.
224. Burrows A., Liebert J. Probing dark matter // Nature. 1995. V. 373, № 6511. P. 191–192.
225. Byerly G. R., Lowe D. R., Walch M. M. Stromatolites from the 3300–3500-Myr Swaziland Supergroup, Barberton Mountain Land, South Africa // Nature. 1986. V. 319, № 6053. P. 489–491.
226. Byerly G. R., Lowe D. R., Wooden J. L., Xie X. An Archean Impact Layer from the Pilbara and Kaapvaal Cratons // Science. 2002. V. 297, № 5585. P. 1325–1327.
227. Campbell I. H., Czamanske G. K., Fedorenko V. A. Hill R. I., Stepanov V. Synchronism of the Siberian Traps and the Permian-Triassic Boundary // Science. 1992. V. 258, № 5089. P. 1760–1763.
228. Cann R. L., Stoneking M., Wilson A.C. Mitochondrial DNA and human evolution // Nature. 1987. V. 325, № 6099. P. 31–36.
229. Carbonell E., Bermú
dez de Castro J. M., Arsuaga J. L., Diez J. C., Rosas A., Guenca-Bescó
s G., Sala R., Mosquera M., Rodriguez X.P. Lower Pleistocene Homi-nids and Artifacts from Atapuerca – TD6 (Spain) // Science. 1995. V. 269, № 5225. P. 826–830.
230. Caro G. M. M., Meierhenrich U. J., Schutte W. A., Barbier B., Segovia A. A., Rosenbauer H., Thiemann W. H.-P., Brack A., Greenberg J. M. Amino acids from ultraviolet irradiation of interstellar ice analogues // Nature. 2002. V. 416, № 6879. P. 403–406.
231. Carroll S. B. Genetics and the making of Homo sapiens // Nature. 2003. V. 422, № 6934. P. 849–857.
232. Catling D. C., Zahnie K. J., McKay C. P. Biogenic Methane, Hydrogen Escape, and the Irreversible Oxidation of Early Earth // Science. 2001. V. 293, № 5531. P. 839–843.
233. Cayrel R., Hill V., Beers T. C., Barbuy B., Spite M., Spite F., Plez B., Andersen J., Bonifacio P., Franç
ois P., Melare P., Nordströ
m B., Primas F. Measurement of stellar age from uranium decay // Nature. 2001. V. 409, № 6821. P. 691–692.
234. Cerling T. E., Hay R. L., O’Neil J. R. Isotopic evidence for dramatic climatic changes in East Africa during the Pleistocene // Nature. 1977. V. 267, № 5607. P. 137–138.
235. Chaboyer B., Demarque P., Kernan P.J., Krauss L. M. A. Lower Limit on the Age of the Universe // Science. 1996. V. 271, № 5251. P. 957–961.
236. Chaimanee Y., Suteethorn V., Jaeger J.-J., Ducrocq S. A new Late Eocene anthropoid primate from Tailand // Nature. 1997. V. 385, № 6615. P. 429–431.
237. Chen J.-Y., Dzik J., Edgecombe G. D., Ramskö
ld L., Zhou G.-Q. A possible Early Cambrian chordate // Nature. 1995. V. 377, № 6551. P. 720–722.
238. Chen J.-Y., Huang D.-Y., Li C.-W. An early Cambrian craniate-like chordate // Nature. 1999. V. 402, № 6761. P. 518–522.
239. Cherfas J. Extinction and the pattern of evolution // New scientist. 1985. V. 108, № 1476. P. 48–51.
240. Chiappe L. M. The first 85 million years of avian evolution // Nature. 1995. V. 378, № 6555. P. 349–355.
241. Chippindale C. At home with the first Europeans // New scientist. 1986. V. 112, № 1529. P. 38–42.
242. Chown M. Double or quit // New scientist. 2000. V. 168, № 2260. P. 24–27.
243. Chown M., Gribbin J. Sizing up the Universe // New scientist. 1986. V. 109, № 1501. P. 30–34.
244. Ciesielski P. Biostratigraphy and paleoecology of neogene and oligocene silico-flagellates from cores recovered during Leg 28, Deep Sea Drilling Project // Initial Reports of the Deep Sea Drilling Project. Washington: US Government Printing Office, 1975. V. 28. P. 625–691.
245. Ciochon R. L., Savage D. E., Tint T., Maw B. Anthropoid Origins in Asia? New Discovery of Amphipithecus from the Eocene of Burma // Science. 1985. V. 229, № 4715. P. 756–759.
246. Clack J. A. Discovery of the earliest – known tetrapod stapes // Nature. 1989. V. 342, № 6248. P. 425–427.
247. Claeys P., Casier J.-G., Margolis S. V. Microtektites and Mass Extinction: Evidence for a Late Devonian Asteroid Impact // Science. 1992, V. 257, № 5073. P. 1102–1104.
248. Clark J. D., Beyene Y., WoldeGabriel G., Hart W. K., Renne P. R., Gilbert H., Defleur A., Suwa G., Katoh S., Ludwig K. R., Boisserie J.-R., Asfaw B., WhiteT. D. Stratigraphic, chronological and behavioural contexts of Pleistocene Homo sapiens from Middle Awash, Ethiopia // Nature. 2003. V. 423, № 6941. P. 747–752.
249. Clark J. D., Heinzelin J. de, Schick K. D., Hart W. K., White T. D., WoldeGabriel G., Walter R. C., Suwa G., Asfaw B., Vrba E., H.-Selassie Y. African Homo erectus: Old Radiometric Ages and Young Oldowan Assemblages in the Middle Awash Valley, Ethiopia // Science. 1994. V. 264, № 5167. P. 1907–1910.
250. Clark J. M., Hopson J. A. Distinctive mammal-like reptile from Mexico and its bearing on the phylogeny of the Tritylodontidae // Nature. 1985. V. 315, № 6018. P. 398–400.
251. Clark S. Polarized Starlight and the Handedness of Life // American Scientist. 1999. V. 87, № 4. P. 336–343.
252. Clarke J. A., Tambussi C. P., Noriega J. I., Erickson G. M., Ketcham R. A. Definitive fossil evidence for the extant avian radiation in the Cretaceous // Nature. 2005. V. 433, № 7023. P. 305–308.
253. Clé
ment G., Ahlberg P. E., Blieck A., BlomH., Clack J. A., Poty E., Thorez J., Janvier P. Devonian tetrapod from western Europe // Nature. 2004. V. 427, № 6973. P. 412–413.
254. Coles P. The pulling power of galaxy clusters // Nature. 1999. V. 398, № 6725. P. 288–289.
255. Coles P., Ellis G. The case for an open Universe // Nature. 1994. V. 370, № 6491. P. 609–615.
256. Conard N. J., Bolus M. Radiocarbon dating the appearance of modern humans and timing of cultural innovations in Europe: new results and new challenges // Journal of Human Evolution. 2003. V. 44, № 3. P. 331–371.
257. Conroy G. C., Vannier M. W. Dental development of the Taung skull from computerized tomography // Nature. 1987. V. 329, № 6140. P. 625–627.
258. Conway Morris S. The Search for the Precambrian-Cambrian Boundary // American Scientist. 1987. V. 75, № 2. P. 156–167.
259. Conway Morris S. The fossil record and the early evolution of the Metazoa // Nature. 1993. V. 361, № 6409. P. 219–225.
260. Cooper A., Penny D. Mass Survival of Birds Across the Cretaceous-Tertiary Boundary: Molecular Evidence // Science. 1997. V. 275, № 5303. P. 1109–1113.
261. Coqueugniot H., Hublin J.-J., Veillon F., Houë
t F., Jacob T. Early brain growth in Homo erectus and implications for cognitive ability // Nature. 2004. V. 431, № 7006. P. 299–302.
262. Corballis M.C. The Gestural Origins of Language // American Scientist. 1999. V. 87, № 2. P. 138–145.
263. Coveney P. Chaos, entropy and the arrow of time // New scientist. 1990. V. 127, № 1736. P. 49–52.
264. Crane P. R. Time for the angiosperms // Nature. 1993. V. 366. № 6456. P. 631–632.
265. Crook J. H. The Socio-ecology of Primates // Social Behaviour in Birds and Mammals. Essays on the Social Ethology of Animals and Man L., N. Y.: Academic Press Inc. (London) Ltd., 1970. P. 103–166.
266. Crook J. H. Social behavior and ethology // The Origin & Evolution of Man: Readings in physical anthropology, N. Y.: T.Y.Crowell Cº, Inc., 1973. P. 283–295.
267. Crook J. H., Gartlan J.S. Evolution of primate societies // Nature. 1966. V. 210, № 5042. P. 1200–1203.
268. Croswell K. The Galactic empire // New scientist. 1990. V. 128. № 1745. P. 54–58.
269. Croswell K. The constant Hubble war // New scientist. 1993. V. 137, № 1860. P. 22–23.
270. Croswell K. The quest for the cosmological constant // New scientist. 1993. V. 137, № 1861. P. 23–27.
271. Cruickshank A. R. I., Benton M. J. Archosaur ankles and the relationships of the thecodontian and dinosaurian reptiles // Nature. 1985. V. 317, № 6039. P. 715–717.
272. Culotta E. Asian Hominids Grow Older // Science. 1995. V. 270, № 5239. P. 1116–1117.
273. Czarnetzki A., Jakob T., Pusch C. M. Palaeopathological and variant conditions of the Homo heidelbergensis type specimen (Mauer, Germany) // Journal of Human Evolution. 2003. V. 44, № 4. P. 479–495.
274. Daeschier E. B., Shubin N. H., Thomson K. S., Amaral W. W. A. Devonian Tetra-pod from North America // Science. 1994. V. 265, № 5172. P. 639–642.
275. Damasio A., Bellugi U., Damasio H., Poizner H., Gilder J. Van. Sign language aphasia during left-hemisphere Amytal injection // Nature. 1986. V. 322, № 6077. P. 363–365.
276. Dar A. Astrophysics and Cosmology Closing in on Neutrino Masses // Science. 1990. V. 250, № 4987. P. 1529–1533.
277. Davis D. L. Paleolithic Diet, Evolution, and Carcinogens // Science. 1987. V. 238, № 4834. P. 1633–1634.
278. Day S. Hot bacteria and other ancestors // New scientist. 1994. V. 142, № 1920. P. 21–25.
279. Dayton L., McDonald M. The atomic age of cave art // New scientist. 1993. V. 137, № 1862. P. 34–37.
280. Dean C., Leakey M. G., Reid D., Schrenk F., Schwartz G. T., Stringer C., Walker A. Growth processes in teeth distinguish modern humans from Homo erectus and earlier hominins // Nature. 2001. V. 414, № 6864. P. 628–631.
281. Dean D., Delson E. Homo at the gates of Europe // Nature. 1995. V. 373, № 6514. P. 472–473.
282. Delsemme A. H. An Argument for the Cometary Origin of the Biosphere // American Scientist. 2001. V. 89, № 5. P. 432–442.
283. Delson E. Human phylogeny revised again // Nature. 1986. V. 322, № 6079. P. 496–497.
284. Delson E. Evolution and palaeobiology of robust Australopithecus // Nature.
1987. V. 327, № 6124. P. 654–655.
285. Delson E. One source not many // Nature. 1988. V. 332, № 6161. P. 206.
286. Delson E. One skull does not a species make // Nature. 1997. V. 389, № 6650. P. 445–446.
287. deMenocal P. B. Plio-Pleistocene African Climate // Science. 1995. V. 270, № 5233. P. 53–59.
288. D’Hondt S., Arthur M. A. Late Cretaceous Oceans and the Cool Tropic Paradox // Science. 1996. V. 271, № 5257. P. 1838–1841.
289. Diamond J. M. DNA-based phylogenies of the three chimpanzees // Nature.
1988. V. 332, № 6166. P. 685–686.
290. Diamond J. M. Were Neanderthals the first humans to bury their dead? // Nature. 1989. V. 340, № 6232. P. 344.
291. Diamond J. M. Quantitative design of life // Nature. 1993. V. 366, № 6454. P. 405–406.
292. Die paläolithischen Neufunde von Markkleeberg bei Leipzig / Baumann W., Mania D., Toepfer V., Eiß
mann L. (Veröffentlichungen des Landesmuseums für Vorgeschichte Dresden. Band 16). Berlin: VEB Deutscher Verlag der Wissenschaften, 1983. 280 S.
293. Donovan S. K. The fit of the continents in the late Precambrian // Nature. 1987. V. 327, № 6118. P. 139–141.
294. Doolittle R. F., Feng D.-F., Tsang S., Cho G., Little E. Determining Divergence Times of the Major Kingdoms of Living Organisms with a Protein Clock // Science. 1996. V. 271, № 5248. P. 470–477.
295.Dorit R. L., Akashi H., Gilbert W. Absence of Polymorphism at the ZFY Locus on the Human Y Chromosome // Science. 1995. V. 268, № 5214. P. 1183–1185.
296. Dressler A., Faber S. M. New measurements of distances to spirals in the great attractor: further confirmation of the large-scale flow // The Astrophisical Journal. 1990. V. 354, № 2, part 2. Letters. P. 45–8.
297. Droser M. L., Fortey L. A., Li X. The Ordovician Radiation // American Scientist. 1996. V. 84, № 2. P. 122–131.
298. Drouot É.
Grotte de la Baume-Latrone // L’art des cavernes: Atlas des grottes ornées paléolithiques françaises. Avant-propos d’Andre Leroi-Gourhan, membre de l’Institut. Paris: Ministère de la Culture, 1984. P. 333–339.
299. Duff M., Sutton C.
The membrane at the end of the Universe // New scientist. 1988. V. 118, № 1619. P. 67–71.
300. Duve C. de.
The second genetic code // Nature. 1988. V. 333, № 6169. P. 117–118.
301. Duve C. de.
The Beginnings of Life on Earth // American Scientist. 1995. V. 83, № 5. P. 428–437.
302. Dvali G.
Neutrino probes of dark energy // Nature. 2004. V. 432, № 7017. P. 567–568.
303. Efstathiou G.
Beyond cosmic ripples // New scientist. 1993. V. 138, № 1871. P. 26–30.
304. Ellis J.
Hopes grow for supersymmetry // Nature. 1985. V. 313, № 6004. P. 626–627.
305. Ellwood B. B., Benoist S. L., El Hassani A., Wheeler
C., Crick RE.
Impact Ejecta Layer from the Mid-Devonian: Possible Connection to Global Mass Extinctions // Science. 2003. V. 300, № 5626. P. 1734–1737.
306. Engel M. H.,
Macko S. A.
Isotopic evidence for extraterrestrial non-racemic amino acids in the Murchison meteorite // Nature. 1997. V. 389, № 6648. P. 265–268.
307. Engel M. S., Grimaldi D. A.
New light shed on the oldest insect // Nature. 2004. V. 427, № 6975. P. 627–630.
308. Enserink M.
Fossil Opens Window On Early Animal History // Science. 1999. V. 286, № 5446. P. 1829.
309. Eppel F.
Fund und Deutung: Eine europäische Urgeschichte. Wien, München: Verlag Anton Schroll & Co, 1958. 247 S.
310. Erickson G. M., Rogers K. C, Yerby S. A.
Dinosaurian growth patterns and rapid avian growth rates // Nature. 2001. V. 412, № 6845. P. 429–433.
311. Erwin D. H.
The Permo-Triassic extinction // Nature. 1994. V. 367, № 6460. P. 231–236.
312. Erwin D., Valentine
J., Jablonski D.
The Origin of Animal Body Plans // American Scientist. 1997. V. 85, № 2. P. 126–137.
313. Evans D. A., Beukes N.
J., Kirschvink J. L.
Low-latitude glaciation in the Palaeoproterozoic era // Nature. 1997. V. 386, № 6622. P. 262–266.
313a. Evans N.
Quarks on a gravitational string // Nature. V. 439, № 7079. P. 921–923.
314. Excavation in the Bacho Kiro cave (Bulgaria): Final report. Warszawa: Panstwowe
wydawnictwo naukowe, 1982. 178 P.
315. Falk D.
Hadar AL 162-28 endocast as evidence that brain enlargement preceded cortical reorganization in hominid evolution // Nature. 1985. V. 313. № 5997. P. 45–47.
316. Faupl P., R
chter W., Urbanek C.
Dating of the Herto hominin fossils // Nature. 2003. V. 426, № 6967. P. 621–622.
317. Fehr E., Fischbacher U.
The nature of human altruism // Natere. 2003. V. 425, № 6960. P. 785–791.
318. Fehr E., Gächter S.
Altruistic punishment in humans // Nature. 2002. V. 415, № 6868. P. 137–140.
319. Fehr E., Gächter S.
The puzzle of human cooperation // Nature. 2003. V. 421, № 6926. P. 911–912.
320. Fehr E., Rockenbach B. Detrimental effects of sanctions on human altruism // Nature. 2003. V. 422, № 6928. P. 137–140.
321. Ferris J. P., Hill Jr A. R., Liu R., Orgel E. Synthesis of long prebiotic oligomers on mineral surfaces // Nature. 1996. V. 381, № 6577. P. 59–61.
322. Field K. G., Olsen G. J., Lane D. J., Giovannoni S. J., Ghiselin M. T., Raff E. C., Pace N. R., Raff R. A. Molecular Phylogeny of the Animal Kingdom // Science. 1988. V. 239, № 4841. P. 748–753.
323. Finch C. E., Pike V. C., Witten M. Slow Mortality Rate Accelerations During Aging in Some Animals Approximate That of Humans // Science. 1990. V. 249, № 4971. P. 902–905.
324. Flynn J. J., Parrish J. M., Rakotosamimanana B., Simpson W. F., Whatley R. L., Wyss A. R. A Triassic Fauna from Madagascar, Including Early Dinosaurs // Science. 1999. V. 286, № 5440. P. 763–765.
325. Forey P., Janvier P. Evolution of the Early Vertebrates // American Scientist. 1994. V. 82, № 6. P. 554–565.
326. Forster C. A., Sampson S. D., Chiappe L. M., Krause D. W. The Theropod Ancestry of Birds: New Evidence from the Late Cretaceous of Madagascar // Science. 1998. V. 279, № 5358. P. 1915–1919.
327. Forward R. The power of negative matter // New scientist. 1990. V. 125, № 1708. P. 54–56.
328. Fox R. C., Youzwyshyn G. P., Krause D. W. Post-Jurassic mammal-like reptile from the Palaeocene // Nature. 1992. V. 358, № 6383. P. 233–235.
329. Fraser N. At the feet of the dinosaurs // New scientist. 1988. V. 120, № 1633. P. 39–42.
330. Fraser N.C., Walkden G.M., Stewart V. The first pre-Rhaetic therian mammal // Nature. 1985. V. 314, № 6007. P. 161–163.
331. Freedman W.L., Madore B.F., Mould J.R., Hill R., Ferrarese L., Kennicutt Jr R.C., Saha A., Stetson P.B., Graham J.A., Ford H., Hoessel J.G., Huchra J., Hughes S.M., Illingworth G.D. Distance to the Virgo cluster galaxy M 100 from Hubble Space Telescope observations of Cepheids // Nature. 1994. V. 371, № 6500. P. 757–762.
332. Frenk C.S., White S.D.M., Efstathiou G., Davis M. Cold dark matter, the structure of galactic haloes and origin of the Hubble sequence // Nature. 1985. V. 317, № 6038. P. 595–597.
333. Frenk C., White S. More missing matter mystery // Nature. 1985. V. 317, № 6039. P. 670–671.
334. Fritzsch B. Inner ear of the coelacanth fish Latimeria has tetrapod affinites // Nature. 1987. V. 327, № 6118. P. 153–154.
335. Fu Y.-X., Li W.-H. Estimating the Age of the Common Ancestor of Men from the ZFY Intron // Science. 1996. V. 272, № 5266. P. 1356–1357.
336. Fukugita M., Hogan C.J., Peebles J.E. The cosmic distance scale and the Hubble constant // Nature. 1993. V. 366, № 6453. P. 309–312.
337. Gabunia L., Vekua A. A Plio-Pleistocene hominid from Dmanisi, East Georgia, Caucasus // Nature. 1995. V. 373, № 6514. P. 509–512.
338. Gabunia L., Vekua A., Lordkipanidze D., Swisher III C.C., Ferring R., Justus A., Nioradze M., Tvalchrelidze M., Antô
n S.C., Bosinski G., Jö
ris O., Lumley M.-A.-de, Majsuradze G., Mouskhelishvili A. Earliest Pleistocene Hominid Cranial Remains from Dmanisi, Republic of Georgia: Taxonomy, Geological Setting, and Age // Science. 2000. V. 288, № 5468. P. 1019–1025.
339. Galtier N., Tourasse N., Gony M. A Nonhyperthermophilic Common Ancestor to Extant Life Forms // Science. 1999. V. 283, № 5399. P. 220–221.
340. Gardner R. A., Gardner B. T. Teaching Sign Language to a Chimpanzee // Science. 1969. V. 165, № 3894. P. 664–672.
341. Gaskell C. M. Direct estimate of Hubble’s constant from the double quasar // Nature. 1985. V. 314, № 6010. P. 402.
342. Gaucher E. A., Thomson J. M., Burgan M. F., Benner S. A. Inferring the palaeoenvironment of ancient bacteria on the basis of resurrected proteins // Nature. 2003. V. 425, № 6955. P. 285–288.
343. Gawiser E., Silk J. Extracting Primordial Density Fluctuations // Science. 1998. V. 280, № 5368. P. 1405–1411.
344. Gebo D. L., MacLatchy L., Kibyo R., Deino A., Kingston J., Pilbeam D. A Ho-minoid Genus from the Early Miocene of Uganda // Science. 1997. V. 276, № 5311. P. 401–404.
345. Gee H., Ahlberg P. E. Four legs to stand on… for Devonian vertebrates // Nature. 1989. V. 342, № 6251. P. 738–739.
346. Gensel P. G., Andrews H. N. The Evolution of Early Land Plants // American Scientist. 1987. V. 75, № 5. P. 478–489.
347. Gersonde R., Kyte F. T., Bleil U., Diekmann B., Flores J.A., Gohl K., Grahl G., Hagen R., Kuhn G., Sierro F. J., Vö
lker D., Abelmann A., Bostwick J. A. Geological record and reconstruction of the late Pliocene impact of the Eltanin asteroid in the Southern Ocean // Nature. 1997. V. 390, № 6658. P. 357–363.
348. Gibbons A. Chimps: More Diverse Than a Barrel of Monkeys // Scitnce. 1992. V. 255, № 5042. P. 287–288.
349. Gibbons A. Paleoanthropologists Launch A Society of Their Own // Science. 1992. V. 256, № 5061. P. 1281–1282.
350. Gibbohs A. New Study Points to Eurasian Ape as Great Ape Ancestor // Science. 1998. V. 281, № 5377. P. 622–623.
351. Gibbons G. Quantum gravity: the last frontier // New scientist. 1985. V. 108, № 1480. P. 45–47.
352. Gibert J., Sanchez F., Ribot F., Gibert L., Ferrandez C., Iglesias A., Gibert P., Gonzá
lez F. Restes humains dans les sédiments du Pléistocène inférieur de la région d’Orce et de Cueva Victoria (sud-est de l’Espagne) // L’anthropologie. 2002. V. 106, № 5. P. 669–683.
353. Gilbert W. The RNA world // Nature. 1986. V. 319, № 6055. P. 618.
354. Gingerich P. D., Haq M., Zalmout I. S., Khan I. H., Malkani M. S. Origin of Whales from Early Artiodactyls: Hands and Feet of Eocene Protocetidae from Pakistan // Science. 2001. V. 293, № 5538. P. 2239–2242.
355. Giovanelli R. Less expansion, more agreement // Nature. 1999. V. 400, № 6740. P. 111–112.
356. Glanz J. New Light on Fate of the Universe // Science. 1997. V. 278, № 5339. P. 799–800.
357. Glanz J. Exploding Stars Point to a Universal Repulsive Force // Science. 1998. V. 279, № 5351. P. 651–652.
358. Glanz J. Astronomers See a Cosmic Antigravity Force at Work // Science. 1998. V. 279, № 5355. P. 1298–1299.
359. Glanz J. No Backing Off From the Accelerating Universe // Science. 1998. V. 282, № 5392. P. 1249–1251.
360. Glanz J. Superheavy Particles From the Big Bang? // Science. 1999. V. 283, № 5405. P. 1095–1096.
361. Glen W. What Killed the Dinosaurs? // American Scientist. 1990. V. 78, № 4. P. 354–370.
362. Goodall J. Continuities between Chimpanzee and Human Behavior // Human Origins. Louis Leakey and the East African Evidence. A Staples Press Book. W.A. Benjamin, Inc., Menlo Park, Ca. – Reading, Mass. – L. – Amsterdam – Don Mills, Ontario – Sydney, 1976. P. 81–95.
363. Goodman B. Phylogeny! Did Bats Evolve Twice? // Science. 1991. V. 253, № 5015. P. 36.
364. Gorina D. P., Vaid J., Bellugi U. The Linguistic Basis of Left Hemisphere Specialization // Science. 1992. V. 255, № 5049. P. 1258–1260.
365. Gostin V. A., Haines P. W., Jenkins R. J. F., Compston W., Williams I. S. Impact Ejecta Horizon Within Late Precambrian Shales, Adelaide Geosyncline, South Australia // Science. 1986. V. 233, № 4760. P. 198–199.
366. Gostin V. A., Keays R. R., Wallace M. W. Iridium anomaly from Acraman impact ejecta horizon: impacts can produce sedimentary iridium peaks // Nature. 1989. V. 340, № 6234. P. 542–544.
366a. Gravina B., Mellars P., Ramsey C. B. Radiocarbon dating of interstratified Neanderthal and early modern human occupations at the Chatelperronian type-site // Nature. 2005. V. 438. № 7064. P. 51–56.
367. Graziosi P. Les industries paléolithiques à dos rabattu et le passage du Paléolithique au Mésolithique en Italie // Bulletin de la Société préhistorique française. 1951. T. 48, № 1–2. P. 55–61.
368. Graziosi P. L’arte preistorica in Italia. Firenze: Sansoni, 1973. 203 P.
369. Green M. B. Unification of forces and particles in superstring theories // Nature. 1985. V. 314, № 6010. P. 409–414.
370. Green M. Superstring Theories in the real world // New scientist. 1985. V. 107, № 1471. P. 35.
371. Gribbin J. In search of the missing mass // New scientist. 1986. V. 109, № 1490. P. 37–40.
372. Gribbin J. The structure of the Universe // New scientist. 1987. V. 116, № 1584. P. 58–62.
373. Gribbin J. An anniversary of some gravity // New scientist. 1987. V. 116, № 1586. P. 44–47.
374. Gribbin J. Famous for 14 minutes, 49 seconds // New scientist. 1993. V. 137, № 1864. P. 41–44.
375. Gribbin J. Is the Universe alive? // New scientist. 1994. V. 141, № 1908. P. 38–40.
376. Gribbin J. The Galaxy’s dark secrets // New scientist. 1994. V. 142, № 1920. P. 26–29.
377. Gribbin J., Wesson P. The fifth dimention of mass // New scientist. 1988. V. 119, № 1631. P. 56–59.
378. Grieve R. A. F. When will enough be enough? // Nature. 1993. V. 363, № 6431. P. 670–671.
379. Griffin A. S., West S. A. Kin Discrimination and the Benefit of Helping in Cooperatively Breeding Vertebrates // Science. 2003. V. 302, № 5645. P. 634–636.
380. Grine F. E., Kay R. F. Early hominid diets from quantitative image analysis of dental microwear // Nature. 1988. V. 333, № 6175. P. 765–768.
381. Grotzinger J. P., Bowring S. A., Saylor B. Z., Kaufman A. J. Biostratigraphic and Geochronologic Constraints on Early Animal Evolution // Science. 1995. V. 270, № 5236. P. 598–604.
382. Groube L., Chappell J., Muke J., Price D. A 40,000 year-old human occupation site at Huon Peninsula, Papua New Guinea // Nature. 1986. V. 324, № 6096. P. 453–455.
383. Groves C. P. A theory of human and primate evolution. Oxford: Clarendon press, 1989. XII. 375 P.
384. Groves C. P., Sabater P.J. From ape’s nest to human fix-point // Man. 1985. V. 20, № 1. P. 22–47.
385. Gruhn R. Linguistic evidence in support of the coastal route of earliest entry into the New World // Man. 1988. V. 23, № 1. P. 77–100.
386. Grü
n R., Beaumont P.B., Stringer C.B. ESR dating evidence for early modern humans at Border Cave in South Africa // Nature. 1990. V. 344, № 6266. P. 537–539.
387. Grü
n R., Beaumont P., Tobias P.V., Eggins S. On the age of Border Cave 5 human mandible // Journal of Human Evolution. 2003. V. 45, № 2. P. 155–167.
388. Gunzig E., Gé
hé
niau J., Prigogine I. Entropy and cosmology // Nature. 1987. V. 330, № 6149. P. 621–624.
389. Guth A. H. Inflationary universe: A possible solution to the horizon and flatness problems // Physical Review D. Particles and fields. 1981. V. 23, № 2. P. 347–356.
390. Guth A. H., Steinhardt P. J. The Inflationary Universe // Scientific American. 1984. V. 250, № 5. P. 90–102.
391. Guthrie R. D. Radiocarbon evidence of mid-Holocene mammoths stranded on an Alascan Bering Sea island // Nature. 2004. V. 429, № 6993. P. 746–749.
392. Gutin J. Do Kenya Tools Root Birth of Modern Thought in Africa? // Science. 1995. V. 270, № 5239. P. 1118–1119.
393. Haile-Selassie Y. Late Miocene hominids from the Middle Awash, Ethiopia // Nature. 2001. V. 412, № 6843. P. 178–181.
394. Hallam A. End-Cretaceous Mass Extinction Event: Argument for Terrestrial Causation. // Science. 1987. V. 238, № 4831. P. 1237–1242.
395. Halliday A. N. In the beginning… // Nature. 2001. V. 409, № 6817. P. 144–145.
396. Hammer M.F. A recent common ancestry for human Y chromosomes // Nature. 1995. V. 378, № 6555. P. 376–378.
397. Hammer W. R., Hickerson W. J. A Crested Theropod Dinosaur from Antarctica // Science. 1994. V. 264, № 5160. P. 828–830.
398. Hamrick M. W., Inouye S. E. Thumbs, Tools, and Early Humans // Science. 1995. V. 268, № 5210. P. 586–587.
399. Han T.-M., Runnegar B. Megascopic Eukaryotic Algae from the 2.1-Billion-Year-Old Negaunee Iron-Formation, Michigan. // Science. 1992. V. 257, № 5067. P. 232–235.
400. Hansell M. What’s so special about using tools? // New scientist. 1987. V. 113, № 1542. P. 54–56.
401. Haq B. U., Hardenbol J., Vail P.R. Chronology of Fluctuating Sea Levels Since the Triassic // Science. 1987. V. 235, № 4793. P. 1156–1167.
402. Hardy A. Was Man more aquatic in the past? // New scientist. 1960. V. 7, № 174. P. 642–645.
403. Harris E. E., Hey J. X chromosome evidence for ancient human histories // Proceedings of the National Academy of Sciences of the USA. 1999. V. 96, № 6. P. 3320–3324.
404. Harris W. E., Durrell P. R., Pierce M. J., Secker J. Constraints on the Hubble constant from observations of the brightest red-giant stars in a Virgo-cluster galaxy // Nature. 1998. V. 395, № 6697. P. 45–47.
405. Hawking S. W. Spacetime foam // Nuclear physics. Ser. B. 1978. V. 144, № 2–3. P. 349–362.
406. Hayes D. E., Frakes L. A. General synthesis, Deep Sea Drilling Project Leg 28 // Initial Reports of the Deep Sea Drilling Project. Washington: US Government Printing Office, 1975. V. 28.. P. 919–942.
407. Hecht J. Evolving theories for old extinctions // New scientist. 1988. V. 120, № 1638. P. 28–30.
408. Heinzelin J. de, Clark J. D., White T., Hart W., Renne P., WoldeGabriel G., Beyene Y., Vrba E. Environment and Behavior of 2.5-Million-Year-Old Bouri Hominids // Science. 1999. V. 284, № 5414. P. 625–629.
409. Hellemans A. Galaxy’s Oldest Stars Shed Light on Big Bang // Science. 1998. V. 281, № 5382. P. 1428–1429.
410. Henbest N. Geologists hit back at impact theory of extinctions // New scientist. 1989. V. 122, № 1662. P. 34–35.
411. Henbest N. Hot bucbles in space // New scientist. 1994. V. 142, № 1923. P. 28–31.
412. Henshilwood C. S., d’Errico F., Yates R., Jacobs Z., Tribolo C., Duller G. A. T., Mercier N., Sealy J. C., Valladas H., Watts I., Wintle A. G. Emergence of Modern Human Behavior: Middle Stone Age Engravings from South Africa // Science. 2002. V. 295, № 5558. P. 1278–1280.
413. Hewes G. W. Primate Communication and the Gestural Origin of Language // Current Anthropology. 1973. V. 14, № 1–2. P. 5–12.
414. Heymann D., Chibante L. P. F., Brooks R. R., Wolbach W. S., Smalley R. E. Fullerenes in the Cretaceous-Tertiary Boundary Layer // Science. 1994. V. 265, № 5172. P. 645–647.
415. Hildebrandt A. R., Boynton W. V. Proximal Cretaceous-Tertiary Boundary Impact Deposits in the Caribbean. // Science. 1990. V. 248, № 4957. P. 843–847.
416. Hill A. Early hominid from Baringo, Kenya // Nature. 1985. V. 315, № 6016. P. 222–224.
417. Hill A., Ward S., Deino A., Curtis G., Drake R. Earliest Homo // Nature. 1992. V. 355, № 6362. P. 719–722.
418. Hoffman P. F. Did the Breakout of Laurentia Turn Gondwanaland Inside-Out? // Science. 1991. V. 252, № 5011. P. 1409–1412.
419. Hofmann A. W. Early Evolution of Continents // Science. 1997. V. 275, № 5299. P. 498–499.
420. Hogan C. J. Cosmological conflict // Nature. 1994. V. 371, № 6496. P. 374–395.
421. Holdaway R. N., Jacomb C. Rapid Extinction of the Moas (Aves: Dinornithi-formes): Model, Test, and Implications // Science. 2000. V. 287, № 5461. P. 2250–2254.
422. Holland H. D. Evidence for Life on Earth More Than 3850 Million Years Ago // Science. 1997. V. 275, № 5296. P. 38–39.
423. Horodyski R. J., Knauth L.P. Life on Land in the Precambrian. // Science. 1994. V. 263, № 5146. P. 494–498.
424. Hou Y., Potts R., Yuan B., Guo Z., Deino A., Wang W., Clark J., Xie G., Huang W. Mid-Pleistocene Acheulean-like Stone Technology of the Bose Basin, South China // Science. 2000. V. 287, № 5458. P. 1622–1626.
425. Hu Y., Meng J., Wang Y., Li C. Large Mesozoic mammals fed on young dinosaurs // Nature. 2005. V. 433, № 7022. P. 149–152.
426. Huang W., Ciochon R., Yumin G., Larick R., Qiren F., Schwarcz H., Yonge C., Vos J. de, Rink W. Early Homo and associated artefacts from Asia // Nature. 1995. V. 378, № 6554. P. 275–278.
427. Huber C., Wä
chtershä
user G. Peptides by Activation of Amino Acids with CO on (Ni, Fe)S Surfaces. Implications for the Origin of Life // Science. 1998. V. 281, № 5377. P. 670–672.
428. Huchra J. P. The Hubble Constant // Science. 1992. V. 256, № 5055. P. 321–325.
429. Hugues C., Gagniè
re S., Rappaz O. Les Moustériens de Sainte-Anastasie (Gard) // Festschrift für Lothar Zotz: Steinzeitfragen der alten und neuen Welt. Bonn: Ludwig Röhrscheid verlag, 1960. P. 217–231.
430. Hutchinson J. R., Garcia M. Tyrannosaurus was not a fast runner // Nature. 2002. V. 415, № 6875. P. 1018–1021.
431. Ikebe Y., Ezawa H., Fukazawa Y., Hirayama M., Ishisaki Y., Kikuchi K., Kubo H., Makishima K., Matsushita K., Ohashi T., Takahashi T., Tamura T. Discovery of a hierarchical distribution of dark matter in the Fornax cluster of galaxies // Nature. 1996. V. 379, № 6564. P. 427–429.
432. Irion R. The lopsided Universe // New scientist. 1999. V. 161, № 2172. P. 26–30.
433. Izett G. A., Dalrymple G. B., Snee L. W. 40Ar/39Ar Age of Cretaceous-Tertiary Boundary Tectites from Haiti. // Science. 1991. V. 252, № 5012. P. 1539–1542.
434. Jablonski D., Raup D. M. Selectivity of End-Cretaceous Marine Bivalve Extinction. // Science. 1995. V. 268. № 5209. P. 389–391.
435. Jacoby G. H. The Univerce in crisis // Nature. 1994. V. 371, ¹ 6500. P. 741–742.
436. Jaeger J.-J., Thein T., Benammi M., Chaimanee Y., Soe A.N., Lwin T., Tun T., Wai S., Ducrocq S. A New Primate from the Middle Eocene of Myanmar and the Asian Early Origin of Anthropoids // Science. 1999. V. 286, № 5439. P. 528–530.
437. Janvier P. Conodonts join the club // Nature. 1995. V. 374, № 6525. P. 761–762.
438. Jasny B. R., Hines P. J. Genome Prospecting // Science. 1999. V. 286, № 5439. P. 365, 443.
439. Jaynes J. The Origin of Consciousness in the Breakdown of the Bicameral Mind. Boston: Houghton Mifflin Company, 1976. 467 P.
440. Jeram A. J., Selden P. A., Edwards D. Land Animals in the Silurian: Arachnids and Myriapods from Shropshire, England // Science. 1990. V. 250, № 4981. P. 658–661.
441. Ji Q., Currie P. J., Norell M. A., Ji S.-A. Two feathered dinosaurs from northeastern China // Nature. 1998. V. 393, № 6687. P. 753–761.
442. Johanson D. C., Masao F. T., Eck G. G., White T. D., Walter R. C., Kimbel W. H., Asfaw B., Manega P., Ndessokia P., Suwa G. New partial skeleton of Homo habilis from Olduvai Gorge, Tanzania // Nature. 1987. V. 327, № 6119. P. 205–209.
443. Johanson D.C., White T. D. A Systematic Assessment of Early African Hominids // Science. 1979. V. 203, № 4378. P. 321–330.
444. Johnson K. R. Extinctions at the antipodes // Nature. 1993. V. 366, № 6455. P. 511–512.
445. Jones J. S., Rouhani S. How small was the bottleneck? // Nature. 1986. V. 319, № 6053. P. 449–450.
446. Joyce G. Hydrothermal vents too hot? // Nature. 1988. V. 334, № 6183. P. 564.
447. Kaku M. Into the eleventh dimension. // New scientist. 1997. V. 153, № 2065. P. 32–36.
448. Kamionkowsky M., Jaffe A. New troubles for inflation? // Nature. 1998. V. 395, № 6703. P. 638–641.
449. Kappelman J. They might be giants // Nature. 1997. V. 387, № 6629. P. 126–127.
450. Kaufman A. J., Xiao S. High CO2 levels in the Proterozoic atmosphere estimated from analyses of individual microfossils // Nature. 2003. V. 425, № 6955. P. 279–282.
451. Ke Y., Su B., Song X., Lu D., Chen L., Li H., Qi C., Marzuki S., Deka R., Underhill P., Xiao C., Shriver M., Lell J., Wallace D., Wells R. S., Seielstad M., Oefner P., Zhu D., Jin J., Huang W., Chakraborty R., Chen Z., Jin L. African Origin of Modern Humans in East Asia: A Tale of 12,000 Y Chromosomes // Science. 2001. V. 292, № 5519. P. 1151–1153.
452. Keany J., Kennett J. P. Pliocene-Pleistocene radiolarian biostratigraphy and paleoclimatology at DSDP Site 278 on the Antarctic convergence // Initial Reports of the Deep Sea Drilling Project. Washington: US Government Printing Office, 1975. V. 29. P. 757–767.
453. Keigwin Jr L. D., Thunell R. C. Middle Pliocene climatic change in the western Mediterranean from faunal and oxygen isotopic trends // Nature. 1979. V. 282, № 5736. P. 294–296.
454. Kennedy K. A. R., Sonakia A., Chiment J., Verma K. K. Is the Narmada Hominid an Indian Homo erectus? // American Journal of Phisical Anthropology. 1991. V. 86, № 4. P. 475–496.
455. Kennett J. P. et al. Cenozoic paleoceanography in the southwest Pacific ocean, Antarctic glaciation, and the development of the Circum-antarctic current // Initial Reports of the Deep Sea Drilling Project. Washington: US Government Printing Office, 1975. V. 29. P. 1155–1169.
456. Kennett J. P., Vella P. Late cenozoic planktonic foraminifera and paleo-ceanography at DSDP Site 284 in the cool subtropical South Pacific // Initial Reports of the Deep Sea Drilling Project. Washington: US Government Printing Office, 1975. V. 29. P. 769–799.
457. Kent S. Sharing in an egalitarian Kalahari community // Man. 1993. V. 28, № 3. P. 479–514.
458. Kenward B., Weir A. A. S., Rutz C., Kacelnik A. Tool manufacture by naive juvenile crows // Nature. 2005. V. 433, № 7022. P. 121.
459. Kerr R. A. Plate Tectonics Goes Back 2 Billion Years // Science. 1985. V. 230, № 4732. P. 1364–1367.
460. Kerr R. A. Plate Tectonics Is the Key to the Distant Past // Science. 1986. V. 234, № 4777. P. 670–672.
461. Kerr R. A. Yucatan Killer Impact Gaining Support // Science. 1991. V. 252, № 5004. P. 377.
462. Kerr R. A. Extinction by a One-Two Comet Punch? // Science. 1992. V. 255, № 5041. P. 160–161.
463. Kerr R. A. A Second Crater Points to Killer Comets // Science. 1993. V. 259, № 5101. P. 1543.
464. Kerr R. A. Earliest Animals Growing Younger? // Science. 1999. V. 284, № 5413. P. 412.
465. Kerr R. A. Early Life Thrived Despite Earthly Travails // Science. 1999. V. 284, № 5423. P. 2111–2113.
466. Kielan-Jaworowska Z., Crompton A.W., Jenkins, Jr, F.A. The origin of egg-laying mammals // Nature. 1987. V. 326, № 6116. P. 871–873.
467. Kimbel W. H., Johanson D. C., Rak Y. The first skull and other new discoveries of Australopithecus afarensis at Hadar, Ethiopia // Nature. 1994. V. 368, № 6470. P. 449–451.
468. Kimura M. The neutral theory of molecular evolution. // New scientist. 1985. V. 107, № 1464. P. 41–46.
469. Klark J. D., Kurashina H. Hominid occupation of the East-Central Highlands of Ethiopia in the Plio-Pleistocene // Nature. 1979. V. 282, № 5734. P. 33–39.
470. Knoll A. H. The Early Evolution of Eukaryotes: A Geological Perspective // Science. 1992. V. 256, № 5057. P. 622–627.
471. Knoll A. H., Bambach R. K., Canfield D. E., Grotzinger J. P. Comparative Earth History and Late Permian Mass Extinction. // Science. 1996. V. 273, № 5274. P. 452–457.
472. Kolb E. W., Seckel D., Turner M. S. The shadow world of superstring theories // Nature. 1985. V. 314, № 6010. P. 415–419.
473. Kraan-Korteweg R. C., Woudt P. A., Cayatte V., Fairall A. P., Balkowski C., Henning P. A. A nearby massive galaxy cluster behinde the Milky Way // Nature. 1996. V. 379. № 6565. P. 519–521.
474. Krauss L. M. What is dark energy? // Nature. 2004. V. 431, № 7008. P. 519–520.
475. Krauss L. M., Chaboyer B. Age Estimates of Globular Clusters in the Milky Way: Constraints on Cosmology // Science, 2003. V. 299, № 5603. P. 65–69.
476. Krings M., Stone A., Schmitz R. W., Krainitzki H., Stoneking M., Pää
bo S. Neandertal DNA Sequences and the Origin of Modern Humans // Cell. 1997. V. 90, № 1. P. 19–30.
477. Krogh T. E., Kamo S. L., Sharpton V. L., Marin L. E., Hildebrand A. R. U-Pb ages of single shocked zircons linking distal K/T ejecta to the Chicxulub crater // Nature, 1993. V. 366, № 6457. P. 731–734.
478. Krogstad E. J., Balakrishnan S., Mukhopadhyay D. K., Rajamani V., Hanson G. N. Plate Tectonics 2.5 Billion Years Ago: Evidence at Kolar, South India // Science. 1989. V. 243, № 4896. P. 1337–1340.
479. Kumar S., Hedges S. B. A molecular timescale for vertebrate evolution. // Nature. 1998. V. 392, № 6679. P. 917–920.
480. Kurté
n B. The Age of Mammals, N.Y.: Columbia University Press, 1972. 250 P.
481. Kuypers M. M. M., Blokker P., Erbacher J., Kinkel H., Pancost R. D., Schouten S., Damsté
J.S.S. Massive Expansion of Marine Archaea During a Mid-Cretaceous Oceanic Anoxic Event // Science. 2001. V. 293, № 5527. P. 92–94.
482. Kyte F. T., Lei Z., Wasson J. T. New Ewidence on the Size and Possible Effects of a Late Pliocene Oceanic Asteroid Impact // Science. 1988. V. 241, № 4861. P. 63–65.
483. Labandeira C. C., Beall B. S., Hueber F. M. Early Insect Diversification: Evidence from a Lower Devonian Bristletail from Québec // Science. 1988. V. 242, № 4880. P. 913–916.
484. Lake J. A. Origin of the eukaryotic nucleus determined by rateinvariant analysis of rRNA sequences // Nature. 1988. V. 331, № 6152. P. 184–186.
485. Laming-Emperaire A. La signification de l’art rupestre paléolithique. Méthode et applications. Thèse de Doctorat. Paris: Éditions A. et J.Picard et Cie, 1962. 424 p.
486. La prehistoria / Leroi-Gourhan A., Bailloud G., Chavaillon J., Laming-Emperaire A. et al. Barcelona: Ed. Labor, 1987. XVI, 331 p.
487. Larick R., Ciochon R. L. The African Emergence and Early Asian Dispersals of the Genus Homo // American Scientist. 1996. V. 84, № 6. P. 538–551.
488. L’art des cavernes: Atlas des grottes ornées paléolithiques françaises. Avantpropos d’Andre Leroi-Gourhan, membre de l’Institut. Paris: Ministère de la Culture, 1984. 673 p.
489. Laville H. Les remplissages de grottes et abris sous roche dans le Sud-Ouest // La préhistoire française. Les civilisations paléolithiques et mésolithiques de la France. Publié à l’occasion du IXe Congrès de l’U.I.S.P.P., Nice, 1976. Paris: Éditions du Centre national de la recherche scientifique, 1976. T. I, part 1. P. 250–270.
490. Leach H. M. Human Domestication Reconsidered // Current Anthropology. 2003. V. 44, № 3. P. 349–368.
491. Leakey M. D. Cultural Patterns in the Olduvai Sequence // After the Australo-pithecines. Stratigraphy, Ecology, and Culture Change in the Middle Pleistocene. The Hague, P. : Mouton Publishers, 1975. P. 477–493.
492. Leakey M. G., Feibel C. S., McDougall I., Walker A. New four-million-year-old hominid species from Kanapoi and Allia Bay, Kenya // Nature. 1995. V. 376, № 6541. P. 565–571.
493. Leakey M. G., Spoor F., Brown F. H., Gathogo P.N., Kiarie C., Leakey L. N., McDougall I. New hominin genus from eastern Africa shows diverse middle Pliocene lineages // Nature. 2001. V. 410, № 6827. P. 433–440.
494. Leakey R. E. Hominids in Africa // American Scientist. 1976. V. 64, № 2. P. 174–178.
495. Leakey R. E. Human Origins. N.Y.: Lodestar books E.P. Dutton, 1982. 87 p.
496. Leonardi P. Incisioni pre-leptolitiche Europee // Annali dell’Universita di Ferrara. Sezione 15: Paleontologia umana e paletnologia. Ferrara, 1975. V. 2, № 8. P. 283–321.
497. Leroi-Gourhan A. Les religions de la préhistoire: (Paléolithique). Paris: Presses Universitaires de France, 1964. 155 p.
498. Leroi-GourhanA. Préhistoire de l’art occidental. Paris: Éditions d’art Lucien Mazenod, 1965. 482 p.
499. Leroi-Gourhan A. Arte y grafismo en la Europa prehistorica. Madrid: Istmo, 1984. 326 p.
500. Leroi-Gourhan A. Dictionnaire de la Préhistoire. Paris: Presses Universitaires de France, 1988. X, 1222 p.
501. Leutenegger W. Origin of hominid bipedalism // Nature. 1987. V. 325, № 6102. P. 305.
502. Lewin R. On the Origin of Insect Wings // Science. 1985. V. 230, № 4724. P. 428–429.
503. Lewin R. A Revolution of Ideas in Agricultural Origins // Science. 1988. V. 240, № 4855. P. 984–986.
504. Lewin R. New Views Emerge on Hunters and Gatherers // Science. 1988. V. 240, № 4856. P. 1146–1148.
505. Lewin R. Ice Age Art Idea Toppled // Science. 1989. V. 243, ¹ 4897. P. 1435.
506. Lewin R. Molecular clocks run out of time // New scientist. 1990. V. 125, № 1703. P. 38–41.
507. Lewin R. Human origins: the chellenge of Java’s skulls // New scientist. 1994. V. 142, № 1924. P. 36–40.
508. Lewin R. Birth of a tool-maker // New scientist. 1995. V. 145, № 1968. P. 38–41.
509. Li C.-W., Chen J.-Y., Hua T.-T. Precambrian Sponges with Cellular Structures // Science. 1998. V. 279, № 5352. P. 879–882.
510. Lieberman D. E. Another face in our family tree // Nature. 2001. V. 410, № 6827. P. 419–420.
511. Lincoln T. Lucy takes a stroll // Nature. 1998. V. 394, № 6691. P. 325.
512. Linde A. The Universe: inflation out of chaos // New scientist. 1985. V. 105, № 1446. P. 14–18.
513. Lindley D. Cold dark matter makes an exit // Nature. 1991. V. 349, № 6304. P. 14.
514. Lindley D. Not so Great Attractor? // Nature. 1992. V. 356, № 6371. P. 657.
515. Lister A. M. Mammoths in miniature // Nature. 1993. V. 362, № 6418. P. 288–289.
516. Livio M. Cosmic Explosions in an Accelerating Universe // Sciense. 1999. V. 286, № 5445. P. 1689–1690.
517. Lowenstein J., Zihlman A. The invisible ape // New scientist. 1988. V. 120, № 1641. P. 56–59.
518. Lucas S. The rise of the dinosaur dynasty // New scientist. 1990. V. 128, № 1737. P. 44–46.
519. Lumley H. de. Cultural Evolution in France in its Paleoecological Setting During the Middle Pleistocene // After the Australopithecines. Stratigraphy, Ecology, and Culture Change in the Middle Pleistocene. The Hague, P. : Mouton Publishers, 1975. P. 745–808.
520. Lumley H. de. Cadre chronologique absolu, paléomagnétisme, chronologie paléontologique et botanique, esquisse paléoclimatologique, séquences culturelles // La préhistoire française. Les civilisations paléolithiques et mésolithiques de la France. Publié à l’occasion du IXe Congrès de l’U.I.S.P.P., Nice, 1976. Paris: Éditions du Centre national de la recherche scientifique, 1976. T. I, part 1. P. 5–23.
521. Lumley H. de.
Les civilisations du Paléolithique inférieur en Provence // La préhistoire française. Les civilisations paléolithiques et mésolithiques de la France. Publié à l’occasion du IXe Congrès de l’U.I.S.P.P., Nice, 1976. Paris: Éditions du Centre national de la recherche scientifique, 1976. T. I, part 2. P. 819–851.
522. Lynch M., Conery J. S.
The Evolutionary Fate and Consequences of Duplicate Genes // Science. 2000. V. 290, № 5494. P. 1151–1155.
523. Lynch M., Conery J. S.
The Origins of Genome Complexity // Science. 2003. V. 302, № 5649. P. 1401–1404.
524. Lynden-Bell D., Faber S. M., Burstein D., Davies R. L., Dressler A., Terlevich R.
J., Wegner G.
Spectroscopy and photometry of elliptical galaxies. V. Galaxy streaming toward the new supergalactic center // The Astrophisical Journal. 1988. V. 326, № 1, part 1. P. 19–49.
525. Maber K. A., Stevenson D. J.
Impact frustration of the origin of life // Nature. 1988. V. 331, № 6157. P. 612–614.
526. MacLeod K. G., Huber B. T.
Reorganization of deep ocean circulation accompanying a Late Cretaceous extinction event // Nature. 1996. V. 380, № 6573. P. 422–425.
527. Madore B. F., Freedman W. L., Silbermann N., Harding P., Huchra J., Mould J. R., Graham J.A., Ferrarese L., Gibson B.K., Han M., Hoessel J.G, Hughes S.M., Illingworth G D., Phelps R, Sakai S., Stetson P.
A Cepheid distance to the Fornax cluster and the local expansion rate of the Universe // Nature. 1998. V. 395, № 6697. P. 47–50.
528. Maddox J.
Have the heavy neutrinos gone? // Nature. 1986. V. 323, № 6083. P. 11.
529. Maddox J.
Prospects for fifth force fade // Nature. 1987. V. 329, № 6137. P. 283.
530. Maddox J.
The search for dark matter (contd) // Nature. 1992. V. 359, № 6393. P. 267.
531. Maddox J.
Big Bang not yet dead but in decline // Nature. 1995. V. 377, № 6545. P. 99.
532. Malaney R. A., Fowler W. A.
The Transformation of Matter after the Big Bang // American Scientist. 1988. V. 76, № 5. P. 472–477.
533. Mania D.
Auf den Spuren des Urmenchen: Die Funde aus der Steinrinne von Bilzingsleben. Berlin: Deutscher Verlag der Wissenschaften, 1990. 283 S.
534. Margolis S. V.
Paleoglacial history of Antarctica inferred from analysis of Leg 29 sediments by scanning-electron microscopy // Initial Reports of the Deep Sea Drilling Project. Washington: US Government Printing Office, 1975. V. 29. P. 1039–1048.
535. Marshack A.
The roots of civilization. N. Y: McGraw-Hill Book Company, 1972. 413 p.
536. Marshack A.
Implications of the Paleolithic Symbolic Evidence for the Origin of Language // American Scientist. 1976. V. 64, № 2. P. 136–145.
537. Marshack A.
Some Implications of the Paleolithic Symbolic Evidence for the Origin of Language // Current Anthropology. 1976. V. 17, № 2. P. 274–282.
538. Marshack A.
On Paleolithic Ochre and the Early Uses of Color and Symbol // Current Anthropology. 1981. V. 22, № 2. P. 188–191.
539. Marshack A.
Non-utilitarian fragment of bone from the middle palaeolithic layer // Excavation in the Bacho Kiro cave (Bulgaria): Final report. Warszawa: Państwowe wydawnictwo naukowe, 1982. P. 117.
540. Marshall J. C.
The descent of the larynx? // Nature. 1989. V. 338, № 6218. P. 702–703.
541. Martin B., Harvey P. H. Human bodies of evidence // Nature. 1987. V. 330, № 6150. P. 697–698.
542. Martin H. Comment vivait l’Homme de La Quina à l’époque moustérienne // Préhistoire. Paris: Librairie Ernest Leroux, 1936. T. 5. P. 7–23.
543. Martin R. D. New fossils and primate origins // Nature. 1991. V. 349, № 6304. P. 19–20.
544. Martin R. D. Primate origins: plugging the gups // Nature. 1993. V. 363, № 6426. P. 223–234.
545. Martin W., Gierl A., Saedler H. Molecular evidence for pre-Cretaceous angio-sperm origins // Nature. 1989. V. 339, № 6219. P. 46–48.
546. Martinez V. J. Is the Universe Fractal? // Science. 1999. V. 284, № 5413. P. 445–446.
547. Mather J. A look at the primeval explosion // New scientist. 1986. V. 109, № 1491. P. 48–50.
548. Mathewson D. S., Ford V.L., Buchhorn M. No back-side in fall into the Great Attractor // The Astrophisical Journal. 1992. V. 389, № 1, part 2. Letters. P. 5–8.
549. Matsuzawa T. Use of numbers by a chimpanzee // Nature. 1985. V. 315, № 6014. P. 57–59.
550. Matthews J. Good vibrations from the stars // New scientist. 1990. V. 125, № 1701. P. 46–50.
551. Matthews R. Can gravity take a quantum leap? // New scientist. 1994. V. 143, № 1942. P. 26–32.
552. Matthews R. Nothing like a vacuum // New scientist. 1995. V. 145, № 1966. P. 30–33.
553. Matthews R. K. Relative elevation of Late Pleistocene high sea level stands: Barbados uplift rates and their implications // Quaternary research. 1973. V. 3, № 1. P. 147–153.
554. Maurrasse F. J-M. R., Sen G. Impacts, Tsunamis, and the Haitian Cretaceous-Tertiary Boundary Layer // Science. 1991. V. 252, № 5013. P. 1690–1693.
555. May R. M. How Many Species Are There on Earth? // Science. 1988. V. 241, № 4872. P. 1441–1449.
556. McCollum M. A. The Robust Australopithecine Face: A Morphogenetic Perspective // Science. 1999. V. 284, № 5412. P. 301–305.
557. McCrone J. Fired up // New scientist. 2000. V. 166, № 2239. P. 30–34.
558. McDonald M. Iridium: tracking down the extraterrestrial element in sedimentary clays // New scientist. 1988. V. 121, № 1652. P. 58.
559. McDougall I., Brown F. H., Fleagle J.G. Stratigraphic placement and age of modern humans from Kibish, Ethiopia // Nature. 2005. V. 433, № 7027. P. 733–736.
560. McElwain J. C., Beerling D. J., Woodward F. I. Fossil Plants and Global Warming after Triassic-Jurassic Boundary // Science. 1999. V. 285, № 5432. P. 1386–1390.
561. McHone J. F., Nieman R. A., Lewis C. F., Yates F.M. Stishovite at the Cretaceous-Tertiary Boundary, Raton, New Mexico // Science. 1989. V. 243, № 4895. P. 1182–1184.
562. McKay C. P., Borucki W. J. Organic Synthesis in Experimental Impact Shocks // Science. 1997. V. 276, № 5311. P. 390–392.
563. McKinnon W. B. Killer acid at the K/T boundary // Nature. 1992. V. 357, № 6373. P. 15–16.
564. McNamara K.
The great evolutionary handicap // New scientist. 1989. V. 123, № 1682. P. 47–51.
565. Melchor R. N., Valais S. de, Genise J.F.
Bird-like fossil foot-prints from the Late Triassic // Nature. 2002. V. 417, № 6892. P. 936–938.
566. Mellars P.
A new chronology for the French Mousterian period // Nature. 1986.
V 322, № 6078. P. 410–411.
567. Mellars P.
Neanderthals and the modern human colonization of Europe // Nature. 2004. V. 432, № 7016. P. 461–465.
567à. Mellars P.
A new radiocarbon revolution and the dispersal of modern humans in Eurasia // Nature. 2006. V. 439, № 7079. P. 931–935.
568. Mercader J., Panger M., Boesch C.
Excavation of a Chimpanzee Stone Tool Site in the African Rainforest // Science. 2002. V. 296, № 5572. P. 1452–1455.
569. Mercier N., Valladas
H., Joron J.-L., Reyss J.-L., Lévéque F., Vandermeersch B.
Thermoluminescence dating of the late Neanderthal remains from Saint-Césaire // Nature. 1991. V. 351, № 6329. P. 737–739.
570. Miller G H., Magee J. W., Johnson B. J., Fogel M. L., Spooner N. A., McCulloch M. T., Ayliffe L. K.
Pleistocene Extinction of Genyornis newtoni:
Human Impact on Australian Megafauna // Science. 1999. V. 283, № 5399. P. 205–208.
571. Miller S. L., Bada J. L.
Submarine hot springs and the origin of life // Nature. 1988. V. 334, № 6183. P. 609–611.
572. Mirazón Lahr M., Foley R.
Human evolution writ small // Nature. 2004. V. 431, № 7012. P. 1043–1044.
573. Mithen S. J.
To hunt or to paint: animals and art in the Upper Palaeolithic // Man. 1988. V. 23, № 4. P. 671–695.
574. Mojzsis S. J., Arrhenius G., McKeegan K. D., Harrison T. M., Nutman A. P., Friend C. R. L.
Evidence for life on Earth before 3,800 million years ago // Nature. 1996. V 384, № 6601. P. 55–59.
575. Mojzsis S. J., Harrison T. M., Pidgeon R. T.
Oxygen-isotope evidence from ancient zircons for liquid water at the Earth’s surface 4,300 Myr ago // Nature. 2001. V. 409, № 6817. P. 178–181.
576. Montagu M. F. A.
Wolf children // The Origin & Evolution of Man: Readings in physical anthropology. N. Y: T. Y Crowell C°, Inc., 1973. P. 248–253.
577. Montagu M. F. A.
The meaning of neonatal and infant immaturity in man // The Origin & Evolution of Man: Readings in physical anthropology. N. Y: T. Y Crowell C°, Inc., 1973. P. 257–260.
578. Mooers A. 0., Redfield R J.
Digging up the roots of life // Nature. 1996. V. 379, № 6566. P. 587–588.
579. Mook W. G.
Radiocarbon dating // Excavation in the Bacho Kiro cave (Bulgaria): Final report. Warszawa: Panstwowe wydawnìctwo naukowe, 1982. P. 168.
580. Moorbath S., Taylor P. N., Orpen J. L., Treloar P., Wilson J. F.
First direct radiometric dating of Archaean stromatolitic limestone // Nature. 1987. V. 326, № 6116. P. 865–867.
581. Moore B.
Evidence against dissipationless from observations of galaxy halos // Nature. 1994. V. 370, № 6491. P. 629–631.
582. Morey D. F.
The Early Evolution of the Domestic Dog // American Scientist. 1994. V. 82, № 4. P. 336–347.
583. Morgan E. Lucy’s child // New scientist. 1986/1987. V. 112, № 1540/1541. P. 13–15.
584. Morris D. The naked ape. L.: Gorgi books, 1975. 219 p.
584a. Morwood M. J., Brown P., Jatmiko, Sutikna T., Saptomo E. W., Westaway K. E., Due R. A., Roberts R. G., Maeda T., Wasisto S., Djubiantono T. Further evidence for small-bodied hominins from the Late Pleistocene of Flores, Indonesia // Nature/ 2005. V. 437, № 7061. P. 1012–1017.
585. Morwood M. J., Soejono R. P., Roberts R. G., Sutikna T., Turney C. S. M., Westaway K. E., Rink W. J., Zhao J.-x., Bergh G. D. van den, Due R. A., Hobbs D. R., Moore M. W., Bird M. I., Fifield L. K. Archaeology and age of a new hominin from Flores in eastern Indonesia // Nature. 2004. V. 431, № 7012. P. 1087–1091.
586. Moskowitz B. The search for dark matter in the laboratory // New scientist. 1989. V. 122, № 1660. P. 39–42.
587. Movius, Jr. H. L. Early man and pleistocene stratigraphy in Southern and Eastern Asia. Cambridge, Mass., USA: Peabody museum, 1944. IX, 125 p.
588. Mungall J. E., Ames D. E., Hanley J. J. Geochemical evidence from the Sudbury structure for crustal redistribution by large bolide impacts // Nature. 2004. V. 429, № 6991. P. 546–548.
589. Murphy J. B., Nance R. D. Supercontinent model for the contrasting character of Late Proterozoic orogenic belts // Geology. 1991. V. 19, № 5. P. 469–472.
590. Nance R. D., Worsley T. R., Moody J. B. Post-Archean biogeochemical cycles and long-term episodicity in tectonic processes // Geology. 1986. V. 14, № 6. P. 514–518.
591. Narlikar J. What if the big bang didn’t happen? // New scientist. 1991. V. 129, № 1758. P. 48–51.
592. Narlikar J. Challenge for the big bang // New scientist. 1993. V. 138, № 1878. P. 27–30.
593. Navaro-GonzalezR., McKay C. P., Mvondo D. N. A possible nitrogen crisis for Archaean life due to reduced nitrogen fixation by lightning // Nature. 2001. V. 412, № 6842. P. 61–64.
594. Nisbet E. G., Sleep N. H. The habitat and nature of early life // Nature. 2001. V. 409, № 6823. P. 1083–1091.
595. Nishida H., Pigg K. B., Rigby J. F. Swimming sperm in an extinct Gondwanan plant // Nature. 2003. V. 422, № 6930. P. 396–397.
596. Norman D., Weishampel D. Vegetarian dinosaurs chew it differently // New scientist. 1987. V. 114, № 1559. P. 42–45.
597. Normile D. Weighing In on Neutrino Mass // Science. 1998. V. 280, № 5370. P. 1689–1690.
598. Officer C. Victims of volcanoes // New scientist. 1993. V. 137, № 1861. P. 34–38.
599. Ohman J. C., Slanina M., Baker G., Mensforth R.P. Thumbs, Tools, and Early Humans // Science. 1995. V. 268, № 5210. P. 587–588.
600. Ohmoto H., Kakegawa T., Lowe D. R. 3.4-Billion-Year-Old Biogenic Pyrites from Barberton, South Africa: Sulfur Isotope Evidence // Science. 1993. V. 262, № 5133. P. 555–557.
601. Ohmoto H., Watanabe Y., Kumazawa K. Evidence from massive siderite beds for a CO2-rich atmosphere before ~ 1,8 billion years ago // Nature. 2004. V. 429, № 6990. P. 395–399.
602. Olsen P. E. A 40-Million-Year Lake Record of Early Mesozoic Orbital Climatic Forcing // Science. 1986. V. 234, № 4778. P. 842–848.
603. Olsen P. E., Kent D. V., Sues H.-D., Koeberl C., Huber H., Montanari A., Rainforth E. C., Fowell S. J., Szajna M. J., Hartline B. W. Ascent of Dinosaurs Linked to an Iridium Anomaly at the Triassic-Jurassic Boundary // Science. 2002. V. 296, № 5571. P. 1305–1307.
604. Oppenheimer B. R., Hambly N. C., Digby A. P., Hodgkin S. T., Saumon D. Direct Detection of Galactic Halo Dark Matter // Science. 2001. V. 292, № 5517 // P. 698–702.
605. Overbye D. Weighing the Universe // Science. 1996. V. 272. V. 5267. P. 1426–1428.
606. Padmanabhan T., Vasanthi M. M. Possible cosmological scenario with an unstable 17-keV neutrino // Nature. 1985. V. 317, № 6035. P. 335–336.
607. Partridge T. C., Granger D. E., Caffee M. W., Clarke R.J. Lower Pliocene Hominid Remains from Sterkfontein. // Science. 2003. V. 300, № 5619. P. 607–612.
608. Patel T. Stone age Picassos // New scientist. 1996. V. 151, № 2038. P. 32–35.
609. Peacock J. A., Cole S., Norberg P., Baugh C. M., Bland-Hawthorn J., Bridges T., Cannon R. D., Colless M., Collins C., Couch W., Dalton G., Decley K., De Propris R., Driver S. P., Efstathiou G., Ellis R. S., Frenk C. S., Glazebrook K., Jackson C., Lahave O., Lewis I., Lumsden S., Maddox S., Percival W. J., Peterson B. A., Price I., Sutherland W., Taylor K. A measurement of the cosmological mass density from clustering in the 2dF Galaxy Redshift Survey // Nature. 2001. V. 410, № 6825. P. 169–173.
610. Peebles P. J. E. Evolution of the cosmological constant // Nature. 1999. V. 398, № 6722. P. 25–26.
611. Peebles P. J. E., Schramm D. N., Turner E. L., Kron R. G. The case for the rela-tivistic hot Big Bang cosmology // Nature. 1991. V. 352, № 6338. P. 769–776.
612. Pennisi E. Genetic Study Shakes Up Out of Africa Theory // Science. 1999. V. 283, № 5409. P. 1828.
613. Pennisi E. Genome Duplications: The Stuff of Evolution? // Science. 2001. V. 294, № 5551. P. 2458–2460.
614. Penny D. What was the first living cell? // Nature. 1988. V. 331, № 6152. P. 111–112.
614a. Penny D. Reativity for molecular clocks // Nature. 2005. V. 436, № 7048. P. 183–184.
615. Perimutter S., Aldering G., Valle M. D., Deustua S., Ellis R. S., Fabbro S., Fruchter A., Goldhaber G., Groom D.E., Hook I. M., Kim A. G., Kim M. Y., Knop R. A., Lidman C., McMahon R. G., Nugent P., Pain R., Panagia N., Pennypacker C.R., Ruiz-Lapuente P., Schaefer B., Walton N. Discovery of a supernova explosion at half the age of the Universe // Nature. 1998. V. 391, № 6662. P. 51–54.
616. Perrett D. I., Lee K. J., Penton-Voak I., Rowland D., Yoshikawa S., Burt D. M., Henzi S. P., Castles D. L., Akamatsu S. Effects of sexual dimorphism on facial attractiveness // Nature. 1998. V. 394, № 6696. P. 884–887.
617. Peterson J. Universe in the balance // New scientist. 2000. V. 168, № 2269. P. 26–29.
618. Petley B. W., Kibble B. P., Hartland A. A measurement of the Planck constant // Nature. 1987. V. 327, № 6123. P. 605–606.
619. Pettigrew J. Q. Flying Primates? Megabats Have the Advanced Pathway from Eye to Midbrain // Science. 1986. V. 231, № 4743. P. 1304–1306.
620. Peyrony D. Le Moustier, ses gisements, ses industries, ses couches géologiques // Revue anthropologique. 1930. T. 40, № 1–3. P. 48–76.
621. Peyrony D. Le Moustier. Ses gisements, ses industries, ses couches géologiques // Revue anthropologique. 1930. T. 40, № 4–6. P. 155–176.
622. Peyrony D. La Ferrasie: Moustérien, Périgordien, Aurignacien // Préhistoire. Paris: Librairie Ernest Leroux, 1934. T. 3. P. 1–92.
623. Pickford M., Senut B. The geological and faunal context of Late Miocene hominid remains from Lukeino, Kenya // Comptes rendus de l’Académie des sciences. Série II. F. a. 2001. T. 332, № 2. P. 145–152.
624. Pierce M. J., Welch D. L., McClure R. D., Bergh S. van den, Racine R., Stetson P. B. The Hubble constant and Virgo cluster distance from observations of Cepheid variables // Nature. 1994. V. 371, № 6496. P. 385–389.
625. Pigeaud R. La grotte ornée Mayenne-Sciences (Thorigné-en-Charnie, Mayenne): grotte-limite aux marges du monde anté-magdalénien. – L’anthropologie. 2002. V. 106, № 4. P. 445–489.
626. Plazza A. Towards a genetic history of China // Nature. 1998. V. 395, № 6703. P. 636–638.
627. Ponce de Leô
n M.S., Zollikofer C.P.E. Neanderthal cranial ontogeny and its implications for late hominid diversity // Nature. 2001. V. 412, № 6846. P. 534–538.
628. Pond C. The great ape debate // New scientist. 1987. V. 116, № 1586. P. 39–42.
629. Ponman T. J., Bertram D. Hot gas and dark matter in a compact galaxy group // Nature. 1993. V. 363, № 6424. P. 51–54.
630. Pope G. G. Bamboo and Human Evolution // Natural history. 1989, № 10. P. 48–57.
631. Pradel L. et J. H. Le Moustérien évolué de l’Ermitage // L’Anthropologie. 1955. T. 58, № 5–6. P. 433–443.
632. Rampino M. Dinosaurs, comets and volcanoes // New scientist. 1989. V. 121, № 1652. P. 54–58.
633. Rampino M. R. Late Permian Extinctions // Science. 1996. V. 274, № 5292. P. 1551.
634. Rampino M. R., Haggerty B. M. Mass Extinction and Periodicity // Science. 1995. V. 269, № 5224. P. 617–618.
635. Raymo M. E., Ruddiman W. F. Tectonic forcing of late Cenozoic climate // Nature. 1992. V. 359, № 6391. P. 117–192.
636. Redecker D., Kodner R., Graham L. E. Glomalean Fungi from the Ordovician // Science. 2000. V. 289, № 5486. P. 1920–1921.
637. Reichow M. K., Saunders A. D., White R. V., Pringle M. S., Al’Mukhamedov A. I., Medvedev A. I., Kirda N. P. 40Ar/39Ar Dates from the West Siberian Basin: Siberian Flood Basalt Province Doubled // Science. 2002. V. 296, № 5574. P. 1846–1849.
638. Ren D. Flower-Associated Brachycera Flies as Fossil Evidence for Jurassic Angiosperm Origin // Science. 1998. V. 280, № 5360. P. 85–88.
639. Renne P. R., Zichao Z., Richards M. A., Black M. T., Basu A. R. Synchrone and Causal Relations Between Permian-Triassic Boundary Crises and Siberian Flood Volca-nism // Science. 1995. V. 269, № 5229. P. 1413–1416.
640. Renton P. Has the Higgs boson been discovered? // Nature. 2004. V. 428, № 6979. P. 141–144.
641. Repenning C. A., Fejfar O. Evidence for earlier date of ’Ubeidiya, Israel, hominid site // Nature. 1982. V. 299, № 5881. P. 344–347.
642. Retallack G. Cretaceous – Tertiary Dinosaur Extinction // Science. 1986. V. 234, № 4781. P. 1170–1171.
643. Retallack G. J. Feakes C. R. Trace Fossil Evidence for Late Ordovician Animals on Land // Science. 1987. V. 235, № 4784. P. 61–63.
644. Rich P. V., Rich T. H., Wagstaff B. E., McEven Mason J., Douthitt C. B., Gregory R. T., Felton E. A. Evidence for Low Temperatures and Biologic Diversity in Cretaceous High Latitudes of Australia // Science. 1988. V. 242, № 4884. P. 1403–1406.
645. Riess A. G. et al. Observational evidence from supernovae for an accelerating universe and a cosmological constant // The Astronomical Journal. 1998. V. 116, ¹ 3. P. 1009–1038.
646. Risch N., Kidd K. K., Tishkoff S. A. Genetic Data and the African Origin of Humans // Science. 1996. V. 274, № 5292. P. 1548–1549.
647. Roberts R. G., Flannery T. F., Ayliffe L. K., Yoshida H., Olley J. M., Prideaux G. J., Laslett G. M., Baynes A., Smith M. A., Jones R., Smith B. L. New Ages for the Last Australian Megafauna: Continent-Wide Extinction About 46,000 Years Ago // Science. 2001. V. 292, № 5523. P. 1888–1892.
648. Roberts R. G., Jones R., Smith M. A. Thermoluminescence dating of a 50,000-year-old human occupation site in northern Australia // Nature. 1990. V. 345, № 6271. P. 153–156.
649. Robin E., Froget L., Jehanno C., Rocchia R. Evidence for a K/T impact event in the Pacific Ocean // Nature. 1993. V. 363, № 6430. P. 615–617.
650. Robinson A. L. Values of Fundamental Conctants Adjusted // Science. 1987. V. 235, № 4789. P. 633–634.
651. Roca A. L., Bar-Gal G. K., Elzirik E., Helgen K. M., Maria R., Springer M. S., O’Brien S. J., Murphy W. J. Mesozoic origin for West Indian insectivores // Nature. 2004. V. 429, № 6992. P. 649–651.
652. Roche H., Delagnes A., Brugal J.-P., Feibel C., Kibunjia M., Mourre V., Texier P.-J. Early hominid stone tool production and technical skill 2.34 Myr ago in West Turkana, Kenya // Nature. 1999. V. 399, № 6731. P. 57–60.
653. Rogers R. R., Swisher III C. C., Sereno P. C., Monetta A. M., Forster C. A., Martinez R. N. The Ischigualasto Tetrapod Assemblage (Late Triassic, Argentina) and 40Ar/39Ar Dating of Dinosaur Origins // Science. 1993. V. 260, № 5109. P. 794–797.
654. Rohde D. L. T., Olson S., Chang J. T. Modelling the recent common ancestry of all living humans // Nature. 2004. V. 431, № 7008. P. 562–566.
655. Romer A. S. Vertebrate paleontology. 3d ed. Chicago, L.: The University of Chicago press, 1971. IX, 468 p.
656. Ron N., Porat N., Ronen A., Tchernov E., Horwitz L. K. Magnetostratigraphy of the Evron Member – implications for the age of the Middle Acheulian site of Evron Quarry // Journal of Human Evolution 2003. V. 44, № 5. P. 633–639.
657. Rowan-Robinson M. Dark doubts for cosmology // New scientist. 1991. V. 129, № 1759. P. 30–44.
658. Rozzi F. V. R., Bermudez de Castro J. M. Surprisingly rapid growth in Neanderthals // Nature. 2004. V. 428, № 6986. P. 936–939.
659. Ruben J. A., Hillenius W. J., Geist N. R., Leitch A., Jones T. D., Currie P. J., Horner J. R., Espe III G. The Metabolic Status of Some Late Cretaceous Dinosaurs // Science. 1996. V. 273, № 5279. P. 1204–1207.
660. Ruben J. A., Jones T. D., Geist N. R., Hillenius W. J. Lung structure and ventilation in Theropod Dinosaurs and Early Birds // Science. 1997. V. 278, № 5341. P. 1267–1270.
661. Rubner M. Die Regelung des Stoff– und Energieverbrauchs beim Wachstum der Wirbeltiere // Die Naturwissenschaften. 1924. Bd. 12. H. 25. S. 493–495.
662. Rubner M. Die Beziehung zwischen Nahrungsaufwand und körperlichen Leistungen des Menschen // Die Naturwissenschaften. 1927. Bd. 15. H. 9. S. 203–212.
663. Rú
jula A. De. Superstrings and supersymmetry // Nature. 1986. V. 320, № 6064. P. 678.
664. Sackett P. D., Morrison H. L., Harding P., Boroson T.A. A faint luminous halo that may trace the dark matter around spiral galaxy NGC 5907 // Nature. 1994. V. 370, № 6489. P. 441–443.
665. Sahu K. C. Stars within the Large Magellanic Cloud as potential lenses for observed microlensing events // Nature. 1994. V. 370, № 6487. P. 275–276.
666. Sahu K. C., Casertano S., Livio M., Gilliand R. L., Panagia N., Andrew M. D., Potter M. Gravitational microlensing by low-mass objects in the globular cluster M 22 // Nature. 2001. V. 411, № 6841. P. 1022–1024.
667. Saitou N. Reconstruction of Molecular Phylogeny of Extant Hominoids From DNA Sequence Data // American Journal of Physical Anthropology. 1991. V. 84, № 1. P. 75–85.
668. Sancetta C., Imbrie J., Kipp N. G. Climatic record of the past 130000 years in North Atlantic deep-sea core V 23–82: Correlation with the terrestrial record // Quaternary research. 1973. V. 3, № 1. P. 110–116.
669. Sansom I. J., Smith M. P., Armstrong H. A., Smith M. M. Presence of the Earliest Vertebrate Hard Tissues in Conodonts // Science. 1992. V. 256, № 5061. P. 1308–1311.
670. Sarich V. M. The Origin of the Hominids: An Immunological Approach // Perspectives on Human Evolution. V. 1, N. Y., Chicago, San Francisco, Atlanta, Dallas, Montreal, Toronto, L.: Holt, Rinehart and Winston, Inc., 1968. P. 94–121.
671. Savolainen P., Zhang Y.-p., Luo J., Lundeberg J., Leitner T. Genetic Evidence for an East Asian Origin of Domestic Dogs // Science. 2002. V. 298, № 5598. P. 1610–1613.
672. Scarre C. Painting by resonance // Nature. 1989. V. 338, № 6214. P. 382.
673. Scheuer P. Weighing the Universe // Nature. 1993. V. 361, № 6408. P. 112–113.
674. Schindewolf O. H. Neokatastrophismus? // Zeitschrift der Deutschen Geologischen Gesellschaft. 1962. Bd. 114. T. 2. 1963. S. 430–445.
675. Schmandt-Besserat D. The Earliest Precursor of Writing // Scientific American. 1978. V. 238, № 6. P. 38–47.
676. Schmandt-Besserat D. From Tokens to Tablets: A Re-evaluation of the So-called “Numerical Tablets” // Visible language. 1981. V. 15, № 4. P. 321–344.
677. Schopf J. W. Microfossils of the Early Archean Apex Chert: New Evidence of the Antiquity of Life // Science. 1993. V. 260, № 5108. P. 640–646.
678. Schopf J. W., Packer B.M. Early Archean (3.3-Billion to 3.5 billion-Year-Old) Microfossils from Warrawoona Group, Australia // Science. 1987. V. 237, № 4810. P. 70–73.
679. Schrenk F., Bromage T. G., Betzler C. G., Ring U., Juwayeyi Y.M. Oldest Homo and Pliocene biogeography of the Malawi Rift // Nature. 1993. V. 365, № 6449 P. 833–836.
680. Schultz P. H., Zarate M., Hames W., Camiliô
n C., King J. A 3.3-Ma Impact in Argentina and Possible Consequences // Science. 1998. V. 282, № 5396 P. 2061–2063.
681. Scott A. Update on Genesis // New scientist. 1985. V. 106, № 1454. P. 30–33.
682. Seilacher A., Bose P. K., Pflü
ger F. Triploblastic Animals More Than 1 Billion Years Ago: Trace Fossil Evidence from India // Science. 1998. V. 282, № 5386. P. 80–83.
683. Sellers A., Meadows A. J. Longterm variations in the albedo and surface temperature of the Earth // Nature. 1975. V. 254, № 5495. P. 44.
684. Semaw S., Renne P., Harris J. W. K., Feibel C.S., Bernor R. L., Fesseha N., Mow-bray K. 2.5-million-year-old stone tools from Gona, Ethiopia // Nature. 1997. V. 385, № 6614. P. 333–336.
685. Semaw S., Rogers M. J., Quade J., Renne P. R., Bulter R. F., Dominguez-Rodrigo M., Stout D., Hart W. S., Pickering T., Simpson S. W. 2.6-Million-year-old stone tools and associated bones from OGS-6 and OGS-7, Gona, Afar, Ethiopia // Journal of Human Evolution. 2003. V. 45, № 2. P. 169–177.
686. Semaw S., Simpson S. W., Quade J., Renne P. R., Bulter R. F., McIntosh W. C., Levin N., Dominguez-Rodrigo M., Rogers M. J. Early Pliocene hominids from Gona, Ethiopia // Nature. 2005. V. 433, № 7023. P. 301–305.
687. Senut B., Pickford M., Gommery D., Mein P., Cheboi K., Coppens Y. First hominid from the Miocene (Lukeino Formation, Kenya) // Comptes rendus de l’Académie des sciences. Série II. F. a. 2001. T. 332, № 2. P. 137–144.
688. Sereno P. C. The Evolution of Dinosaurs // Science. 1999. 284, № 5423. P. 2137–2147.
689. Sereno P. C., Forster C. A., Rogers R. R., Monetta A. M. Primitive dinosaur sceleton from Argentina and the early evolution of Dinosauria // Nature. 1993. V. 361, № 6407. P. 64–66.
690. Sereno P. C., Novas F. E The Complete Skull and Skeleton of an Early Dinosaur // Science. 1992. V. 258, № 5085. P. 1137–1140.
691. Service R. F. A Biomolecule Building Block From Vents // Science. 1998. V. 281, № 5385. P. 1936–1937.
692. Shackleton N. J. The Stratigraphic Record of Deep-Sea Cores and Its Implications for the Assessment of Glacials, Interglacials, Stadials, and Interstadials in the Mid-Pleistocene // After the Australopithecines. Stratigraphy, Ecology, and Culture Change in the Middle Pleistocene. The Hague, P. : Mouton Publishers, 1975. P. 1–24.
693. Shackleton N. J., Kennett J. P. Late cenozoic oxygen and carbon isotopic changes at DSDP site 284: Implications for glacial history of the Northern Hemisphere and Antarctica // Initial Reports of the Deep Sea Drilling Project. Washington: US Government Printing Office, 1975. P. 801–807.
694. Shackleton N. J., Opdyke N. D. Oxygen Isotope and Palaeomagnetic Stratigraphy of Equatorial Pacific Core V 28–238: Oxygen Isotope Temperatures and Ice Volumes on a 105 Year and 106 Year Scale // Quaternary research. 1973. V. 3, № 1. P. 39–55.
695. Shectman S. A., Landy S. D., Oemler A., Tucker D. L., Lin H., Kirshner R. P., Schechter P. L. The Las Campanas Redshift Survey // The Astrophysical Journal. 1996. V. 470, № 1, part 1. P. 172–188.
696. Sheehan P. M., Morse C. L. Cretaceous – Tertiary Dinosaur Extinction // Science. 1986. V. 234, № 4781. P. 1171–1172.
697. Shipman P. Birds do it… did dinosaurs? // New scientist. 1997. V. 53, № 2067. P. 26–31.
698. Shixing Z., Huineng C. Megascopic Multicellular Organisms from the 1700-Million-Year-Old Tuanshanzi Formation in the Jixian Area, North China // Science. 1995. V. 270, № 5236. P. 620–622.
699. Shu D.-G., Chen L., Han J., Zhang X.-L. An Early Cambrian tunicate from China // Nature. 2001. V. 411, № 6836. P. 472–473.
700. Shu D.-G., Conway Morris S., Han J., Zhang Z.-F., Liu J.-N. Ancestral echinoderms from the Chengjiang deposits of China // Nature. 2004. V. 430, № 6998. P. 422–428.
701. Shu D., Conway Morris S., Zhang Z. F., Liu J. N., Han J., Chen L., Zhang X. L., Yasui K., Li Y. A New Species of Yunnanozoan with Implications for Deuterostome Evolution // Science. 2003. V. 299, № 5611. P. 1380–1384.
702. Shu D-G., Luo H-L., Conway Morris S., Zhang X-L., Hu S-X., Chen L., Han J., Zhu M., Li Y., Chen L-Z. Lower Cambrian vertebrates from south China // Natura. 1999. V. 402, № 6757. P. 42–46.
703. Shu D., Zhang X., Chen L. Reinterpretation of Yunnanozoon as the earliest known hemichordate // Nature. 1996. V. 380. 6573. P. 428–430.
704. Silk J. What makes nearby galactic clusters all move as one? // Nature. 1986. V. 322, № 6076. P. 207–208.
705. Silk J. Dark matter comes in from the cold // Nature. 1993. V. 361, № 6408. P. 111.
706. Silk J., Boemen H. A gamma-ray constraint on the nature of dark matter // The Astrophysical Journal. 1987. V. 313, ¹ 2, part 2. Letters. P. 47–51.
707. Simon H. A. A Mechanism for Social Selection and Successful Altruism // Science. 1990. V. 250, № 4988. P. 1665–1668.
708. Simons E. L. Human Origins // Science. 1989. V. 245, № 4924. P. 1343–1350.
709. Simons E. L. Discovery of the Oldest Known Anthropoidean Skull from the Paleogene of Egypt // Science. 1990. V. 247, № 4950. P. 1567–1569.
710. Sinclair A. Art of the ancients // Nature. 2003. V. 426, № 6968. P. 774–775.
711. Sinclair A. R. E., Leakey M.D., Norton-Griffiths M. Migration and hominid bipedalism // Nature. 1986. V. 324, № 6095. P. 307–308.
712. Siveter D. J., Williams M., Waloszek D. A Phosphatocopid Crustacean with Appendages from the Lower Cambrian // Science. 2001. V. 293, № 5529. P. 479–481.
713. Sloan R. E., Rigby, Jr., J. K. Cretaceous – Tertiary Dinosaur Extinction // Science. 1986. V. 234, № 4781. P. 1173–1175.
714. Sloan R. E., Rigby, Jr., J. K., Van Valen L. M., Gabriel D. Gradual Dinosaur Extinction and Simultaneous Ungulate Radiation in the Hell Creek Formation // Science. 1986. V. 232, № 4750. P. 629–633.
715. Small M. F. Ay up, a chimp wi’ an accent // New scientist. 1994. V. 142, № 1928. P. 33–37.
716. Smitson T. R. The earliest known reptile // Nature. 1989. V. 342, № 6250. P. 676–678.
717. Sneden C., Cowan J. J. Genesis of the Heaviest Elements in the Milky Way Galaxy // Science. 2003. V. 299, № 5603. P. 70–75.
718. Sogin M. L., Gunderson J. H., Elwood H. J., Alonso R. A., Peattie D. A. Phylogenetic Meaning of the Kingdom Concept: An Unusual Ribosomal RNA from Giardia lamblia // Science. 1989. V. 243, № 4887. P. 75–77.
719. Sponheimer M., Lee-Thorp J. A Isotopic Evidence for the Diet of an Early Hominid, Australopithecus africanus // Science. 1999. V. 283, № 5400. P. 368–370.
720. Spoor F., Wood B., Zonneveld F. Implications of early hominid labyrinthine morphology for evolution of human bipedal locomotion // Nature. 1994. V. 369, № 6482. P. 645–648.
721. Spray J. G., Kelley S. P., Rowley D. B. Evidence for a late Triassic multiple impact event on Earth // Nature. 1998. V. 392, № 6672. P. 171–173.
722. Stanford C. B. Chimpanzee Hunting Behavior and Human Evolution // American Scientist. 1995. V. 83, № 3. P. 256–261.
723. Stanley S. M., Yang X. A Double Mass Extinction at the End of the Paleozoic Era // Science. 1994. V. 266, № 5189. P. 1340–1344.
724. Steinhardt P. J., Turok N. A Cyclic Model of the Universe // Science. 2002. V. 296, № 5572. P. 1436–1439.
725. Stenger V. J. Photinos from cosmic sources // Nature. 1985. V. 317, № 6036. P. 411–413.
726. Stewart C.-B., Disotell T. R. Primate evolution – in and out of Africa // Current Biology. 1998. V. 8, № 16. P. 582–588.
727. Stokes M. D., Holland N. D. The Lancelet // American Scientist. 1998. V. 86, № 6. P. 552–560.
728. Straus L. G. Age of the modern Europeans // Nature. 1989. V. 342, № 6249. P. 476–477.
729. Stringer C. The dates of Eden // Nature. 1988. V. 331, № 6157. P. 565–566.
730. Stringer C. The Asian connection // New scientist. 1990. V. 128, № 1743. P. 33–37.
731. Stringer C. Secrets of the Pit of the Bones // Nature. 1993. V. 362, № 6420. P. 501–502.
732. Stringer C. Out of Ethiopia // Nature. 2003. V. 423, № 6941. P. 692–695.
733. Stringer C. B., Grü
n R. Time for the last Neanderthals // Nature. 1991. V. 351, № 6329. – P. 701–702.
734. Stringer C. B., Grü
n R., Schwarcz H.P., Goldberg P. ESP dates for the hominid burial site of Es Skhul in Israel // Nature. 1989. V. 338, № 6218. P. 756–758.
735. Stuart A. Who (or what) killed the giant armadillo? // New scientist. 1986. V. 111, № 1517. P. 29–32.
736. Stuart A. J., Kosintsev P. A., Higham T. F. G., Lister A. M. Pleistocene to Holo-cene extinction dynamics in giant deer and woolly mammoth // Nature. 2004. V. 431, № 7009. P. 684–689.
737. Susman R. L. Hand of Paranthropus robustus from Member 1, Swartkrans: Fossil Evidence for Tool Behavior // Science. 1988. V. 240, № 4853. P. 781–784.
738. Susman R. L. Fossil Evidence for Early Hominid Tool Use // Science. 1994. V. 265, № 5178. P. 1570–1573.
739. Sutton C. Universe in the balance // New scientist. 1986. V. 111, № 1518. P. 45–48.
740. Sutton C. The secret life of the neutrino // New scientist. 1988. V. 117, № 1595. P. 53–57.
741. Suwa G., Asfaw B., Beyene Y., White T. D., Katoh S., Nagaoka S., Nakaya H., Uzawa K., Renne P., WoldeGabriel G. The first skull of Australopithecus boisei // Nature. 1997. V. 389, № 6650. P. 489–492.
742. Svensen H., Planke S., Malthe-Serenssen A., Jamtveit B., Myklebust R., Eldem T. R., Rey S. S. Release of methane from a volcanic basin as a mechanism for initial Eocene global warming // Nature. 2004. V. 429, № 6991. P. 542–545.
743. Swinburne N. It came from outer space // New scientist. 1993. V. 137, № 1861. P. 28–32.
744. Swisher III C. C., Curtis G. H., Jacob T., Getty A. G., Suprijo A., Widiasmoro. Age of the Earliest Known Hominids in Java, Indonesia // Science. 1994. V. 263, № 5150. P. 1118–1121.
745. Swisher III C. C., Grajales-Nishimura J. M., Montanary A., Margolis S. V., Claeys P., Alvarez W., Renne P., Cedillo-Pardo E., Maurrasse F. J-M. R., Curtis G. H., Smity J., McWilliams M. O. Coeval 40Ar/39Ar Ages of 65.0 Million Years Ago from Chicxulub Crater Melt Rock and Cretaceous-Tertiary Boundary Tektites // Science. 1992. V. 257, № 5072. P. 954–958.
746. Swisher III C. C., Rink W. J., Antó
n S. C., Schwarcz H. P., Curtis G. H., Suprijo A., Widiasmoro. Latest Homo erectus of Java: Potential Contemporaneity with Homo sapiens in Southeast Asia // Science. 1996. V. 274, № 5294. P. 1870–1874.
747. Swisher III C. C., Wang Y-q., Wang X., Xu X., Wang Y. Cretaceous age for the feathered dinosaurs of Liaoning, China // Nature. 1999. V. 400, № 6739. P. 58–61.
748. Tammann G. A. The Hubble Constant: A Discourse // Publications of the Astronomical Society of the Pacific. 1996. V. 108, № 730. P. 1083–1090.
749. Tapleton A. R. Out of Africa again and again // Nature. 2002. V. 416, № 6876. P. 45–51.
750. Tarduno J. A., Brinkman D. B., Renne P.R., Cottrell R. D., Scher H., Castillo P. Evidence for Extreme Climatic Warmth from Late Cretaceous Arctic Vertebrates // Science. 1998. V. 282, № 5397. P. 2241–2244.
751. Tata: Eine mittelpaläolithische Travertin-Siedlung in Ungarn. (Archaeologia hungarica, series nova, t. 43). Budapest: Akadémiai kiadó, 1964. 284 S.
752. Thewissen J. G. M., Hussain S. T., Arif M. Fossil Evidence for the Origin of Aquatic Locomotion in Archaeocete Whales // Science. 1994. V. 263, № 5144. P. 210–212.
753. Tice M. M., Lowe D. R. Photosynthetic microbial mats in the 3,416-Myr-old ocean // Nature. 2004. V. 431, № 7008. P. 549–552.
754. Tiedemann H. “Killer” Impacts and Life’s Origins // Science. 1997. V. 277, № 5332. P. 1687–1688.
755. Tiemei C., Quan Y., En W. Antiquity of Homo sapiens in China // Nature. 1994. V. 368, № 6466. P. 55–56.
756. Tishkoff S. A., Dietzsch E., Speed W., Pakstis A. J., Kidd J. R., Cheung K., Bonné
-Tamir B., Santachiara-Benerecetti A. S., Moral P., Krings M., Pää
bo S., Watson E., Risch N., Jenkins T., Kidd K.K. Global Patterns of Linkage Disequilibrium at the CD4 Locus and Modern Human Origins // Science. 1996. V. 271, № 5254. P. 1380–1387.
757. Tozzi C. L’industria musteriana della Grotta di Gosto sulla Montagna di Cetona (Siena) // Rivista di scienze preistoriche. Firenze. 1974. V. 29, № 2. P. 271–304.
758. Trinkaus E., Long J. C. Species Attribution of the Swartkrans Member 1 First Metacarpals: SK 84 and SKX 5020 // American Journal of Physical Anthropology. 1990. V. 83, № 4. P. 419–424.
759. Tully R. B. Alignment of clusters and galaxies on scales up to 0,1 c // The Astro-physical Journal. 1986. V. 303, № 1, part 1. P. 25–38.
760. Turner M. S. Why Is the Temperature of the Universe 2,726 Kelvin? // Science. 1993. V. 262, № 5135. P. 861–867.
761. Ucko P. J., Rosenfeld A. L’Art paléolithique. Texte français de M.Dumartheray. Paris: Hachette, 1966. 256 p.
762. Valla F. R. Le natoufien: une culture préhistorique en Palestine. P. : J. Gabalda et Cie, Éditeurs, 1975. 133 p.
763. Valladas H., Cachier H., Maurice P., Bernaldo de Quiros F., Clottes J., Cabrera Valdé
s V., Uzquiano P., Arnold M. Direct radiocarbon dates for prehistoric paintings at the Altamira, El Castillo and Niaux caves // Nature. 1992. V. 357, № 6373. P. 68–70.
764. Valladas H., Geneste J. M., Joron J. L., Chadelle J. P. Thermoluminescence dating of Le Moustier (Dordogne, France) // Nature. 1986. V. 322, № 6078. P. 452–454.
765. Valladas H., Joron J. L., Valladas G., Arensburg B., Bar-Yosef O., Belfer-Cohen A., Goldberg P., Laville H., Meignen L., Rak Y., Tchernov E., Tillier A. M., Vandermeersch B. Thermoluminescence dates for the Neanderthal burial site at Kebara in Israel // Nature. 1987. V. 330, № 6144. P. 159–160.
766. Valladas H., Reyss J. L., Valladas G., Bar-Yosef O., Vandermeersch B. Thermolu-miniscence dating of Mousterian “Proto-Cro-Magnon” remains from Israel and the origin of modern man // Nature. 1988. V. 331, № 6157. P. 614–616.
767. Vallois H. V. The Fontéchevade fossil men // American Journal of Physical Anthropology. 1949. V. 7, № 3. P. 339–362.
768. Vartanyan S. L., Garutt V. E., Sher A. V. Holocene dwarf mammoths from Wrangel Island in the Siberian Arctic // Nature. 1993. V. 362, № 6418. P. 337–340.
769. Vaufrey R. Le Paléolithique italien (Archives de l’Institut de paléontologie humaine. Mémoire 3). Paris: Masson et Cie, éditeurs, 1928. 196 p.
770. Vekua A., Lordkipanidze D., Rightmire G. P., Agusti J., Ferring R., Maisuradze G., Mouskhelishvili A., Nioradze M., Ponce de Leô
n M., Tappen M., Tvalchrelidze M., Zollikofer C. A New Skull of Early Homo from Dmanisi, Georgia // Science. 2002. V. 297, № 5578. P. 85–89.
771. Venkatesan M. I., Dahl J. Organic geochemical evidence for global fires at the Cretaceous/Tertiary boundary // Nature. 1989. V. 338, № 6210. P. 57–60.
772. Verhaegen M. Origin of hominid bipedalism // Nature. 1987. V. 325. № 6102. P. 305–306.
773. Vé
zian J. Gisement moustérien de la grotte du Portel, à Loubens (Ariège) // Bulletin de la Société d’Histoire naturelle de Toulouse. Toulouse. 1955. T. 90, № 1–2. P. 211–217.
774. Vigilant L., Stoneking M., Harpending H., Hawkes K., Wilson A.C. African Populations and the Evolution of Human Mitochondrial DNA // Science. 1991. V. 253, № 5027. P. 1503–1507.
775. Vignaud P., Duringer P., Mackaye H. T., Likius A., Blondel C., Boisserie J.-R., de Bonis L., Eisenmann V., Etienne M.-E., Geraads D., Guy F., Lehmann T., Lihoreau F., Lopez-Martinez N., Mourer-Chauvivé
C., Otero O., Rage J.-C., Schuster M., Viriot L., Zazzo A., Brunet M. Geology and palaeontology of the Upper Miocene Toros-Menalla hominid locality, Chad // Nature. 2002. V. 418, № 6894. P. 152–155.
776. Vilà
C., Leonard J. A., Gö
therströ
m A., Marklund S., Sandberg K., Lidé
n K., Wayne R.K., Ellegren H. Widespread Origins of Domestic Horse Lineages // Science. 2001. V. 291, № 5503. P. 474–477.
777. Vilà
C., Savolainen P., Maldonado J.E., Amorim I. R., Rice J. E., Honeyoutt R. L., Crandall K. A., Lundeberg J., Wayne R. K. Multiple and Ancient Origins of the Domestic Dog // Science. 1997. V. 276, № 5319. P. 1687–1689.
778. Vines G. Are chimps chumps? // New scientist. 2000. V. 168, № 2260. P. 38–42.
779. Vogel G. Chimps in the Wild Show Stirrings of Culture // Science. 1999. V. 284, № 5423. P. 2070–2073.
780. Wainright P. O., Hinkle G., Sogin M. L., Stickel S. K. Monophyletic Origins of the Metazoa: An Evolutionary Link with Fungi // Science. 1993. V. 260, № 5106. P. 340–342.
781. Wainscoat J. Out of the garden of Eden // Nature. 1987. V. 325, № 6099. P. 13.
782. Wainscoat J. S., Hill A. V. S., Boyce A.L., Flint J., Hernandez M., Thein S. L., Old J. M., Lynch J. R., Falusi A. G., Weatherall D. J., Clegg J. B. Evolutionary relationships of human populations from an analysis of nuclear DNA polymorphism // Nature. 1986. V. 319, № 6053. P. 491–493.
783. Waldrop M. M. In Search of Dark Matter // Science. 1986. V. 234, № 4773. P. 152–154.
784. Waldrop M. M. The Large-Scale Structure of the Universe Gets Larger – Maybe // Science. 1987. V. 238, № 4829. P. 894.
785. Waldrop M. M. Cosmologists Begin to Fill in the Blanks // Science. 1991. V. 251, № 4989. P. 30–31.
786. Walkden G., Parker J., Kelley S. A Late Triassic Impact Ejecta Layer in Southwestern Britain // Science. 2002. V. 298, № 5601. P. 2185–2188.
787. Walker A., Leakey R. E., Harris J. M., Brown F. H. 2.5-Myr Australopithecus boisei from west of Lake Turkana, Kenya // Nature. 1986. V. 322, № 6079. P. 517–522.
788. Walsh M. M., Lowe D. R. Filomentous microfossils from the 3,500-Myrold Onverwacht Group, Barberton Mountain Land, South Africa // Nature. 1985. V. 314, № 6011. P. 530–532.
789. Wang Y., Mukherjee P. Model-independent constraints on dark energy density from flux-averaging analysis of type Ia supernova data // The Astrophysical Journal. 2004. V. 606, № 2. Part 1. P. 654–663.
790. Ward S., Brown B., Hill A., Kelley J., Downs W. Equatorius: A New Hominoid Genus from the Middle Miocene of Kenya // Science. 1999. V. 285, № 5432. P. 1382–1386.
791. Watson A. Mathematics of a fake world // New scientist. 1988. V. 118, № 1615. P. 41–45.
792. Watson A. The Universe Shows Its Age // Science. 1998. V. 279, № 5353. P. 981–983.
793. Weiss G., Haeseler A. von. Estimating the Age of the Common Ancestor of Men from the ZFY Intron // Science. 1996. V. 272, № 5266. P. 1359–1360.
794. Wellman C. H., Osterloff P. L., Mohiuddin U. Fragments of the earliest land plants // Nature. 2003. V. 425, № 6955. P. 282–285.
795. White S. D. M., Davis M., Efstathiou G., Frenk C. S.
Galaxy distribution in a cold dark matter universe // Nature. 1987. V. 330, № 6147. P. 451–53.
796. White T. D., Asfaw B., DeGusta D., Gilbert H., Richards G. D., Suwa G., Howell C.
Pleistocene Homo sapiens
from Middle Awash, Ethiopia // Nature. 2003. V. 423, № 6941. P. 742–747.
797. White T. D., Suwa G., Asfaw B. Australopithecus ramidus,
a new species of early hominid from Aramis, Ethiopia // Nature. 1994. V. 371, № 6495. P. 306–312.
798. White T. D., Suwa G., Asfaw B. Australopithecus ramidus,
a new species of early hominid from Aramis, Ethiopia (Corrigendum) // Nature. 1995. V. 375, № 6526. P. 88.
799. White T. D., Suwa G, Hart W. K., Walter R. C, WoldeGabriel G, Heinzelin J. de, Clark J. D., Asfaw B., Vrba E.
New discoveries of Australopithecus
at Maka in Ethiopia // Nature. 1993. V. 366, № 6452. P. 261–265.
800. Whiten A., Goodall J., McGrew W. C, Nishida T., Reynolds V., Sugiyama Y, Tutin C. E. G, Wrangham R.W., Boesch C.
Cultures in chimpanzees // Nature. 1999. V. 399, № 6737. P. 682–685.
801. Whitfield L. S., Sulston J. E., Goodfellow P. N.
Sequence variation of the humane Y chromosome // Nature. 1995. V. 378, № 6555. P. 379–380.
802. Wignall P. B., Twitchett R. J.
Oceanic Anoxia and the End Permian Mass Extinction // Science. 1996. V. 272, № 5265. P. 1155–1158.
803. Wilczek F.
Masses and molasses // New scientist. 1999. V. 162, № 2181. P. 32–37. 803 a. Wild E. M., Teschler-Nicola M., Kutschera W., Steier P., Trinkaus E., Wanek W.
Direct dating of Early Upper Palaeolithic human remains from Mladec // Nature. 2005.
V 435, № 7040. P. 332–335.
804. Wilde P., Berry W. B. N., Quinby-Hunt M. S., Orth C. J., Quintana L. R, Gilmore J. S.
Iridium Abundances Across the Ordovician-Silurian Stratotype // Science. 1986. V. 233, № 4761. P. 339–341.
805. Wilde S. A., Valley J. W., Peck W.
H., Graham C. M.
Evidence from detrital zircons for the existence of continental crust and oceans on the Earth 4.4 Gyr ago // Nature. 2001. V. 409, № 6817. P. 175–178.
806. Williams G. E.
The Acraman Impact Structure: Source of Ejecta in Late Precambrian Shales, South Australia // Science. 1986. V. 233, № 4760. P. 200–203.
807. Williams G. E., Sonett C. P.
Solar signature in sedimentary cycles from the late Precambrian Elatina Formation, Australia // Nature. 1985. V. 318, № 6046. P. 523–527.
808. Willick J. A.
The pecular velocity field in the Perseus-Pisces supercluster // The Astrophisical Journal. 1990. V. 351, № 1. Part 2. Letters. P. 5–8.
809. Wilson A. C, Sarich V. M.
A molecular time scale for human evolution // Proceedings of the National Academy of Sciences of the USA. 1969. V. 63, № 4. P. 1088–1093.
810. Winterhalder B.
Work, resources and population in foraging societies // Man. 1993. V. 28, № 2. P. 321–340.
811. WoldeGabriel G., Haile-Selassie Y, Renne P. R, Hart W. K., Ambrose S. N., Asfaw B., Heiken G., White T.
Geology and palaeontology of the Late Miocene Middle Awash valley, Afar rift, Ethiopia // Nature. 2001. V. 412, № 6843. P. 175–178.
812. WoldeGabriel G., Renne P., White T. D., Suwa G., Heinzelin J. de, Hart W. K., Heiken G.
Age of early hominids // Nature. 1995. V. 376, № 6541. P. 559.
813. WoldeGabriel G., White T. D., Suwa G., Renne P., Heinzelin J. de, Hart W. K., Heiken G. Ecological and temporal placement of early Pliocene hominids at Aramis, Ethiopia // Nature. 1994. V. 371, № 6495. P. 330–333.
814. Wolpoff M. H., Senut B., Pickford M., Hawks J. Sahelanthropus or «Sahelpithecus»? // Nature. 2002. V. 419, № 6907. P. 581–582.
815. Wood B. Who is the «real» Homo habilis? // Nature. 1987. V. 327, № 6119. P. 187–188.
816. Wood B. Origin and evolution of the genus Homo // Nature. 1992. V. 355, № 6363. P. 783–790.
817. Wood B. Four legs good, two legs better // Nature. 1993. V. 363, № 6430. P. 587–588.
818. Wood B. Rift on the record // Nature. 1993. V. 365, № 6449. P. 789–790.
819. Wood B. The oldest whodunnit in the world // Nature. 1997. V. 385, № 6614. P. 292–293.
820. Wood B. Ecce Homo – behold mankind // Nature. 1997. V. 390, № 6656. P. 120–121.
821. Wood B. Hominid revelations from Chad // Nature. 2002. V. 418, № 6894. P. 133–135.
822. Wood B., Brooks A. We are what we ate // Nature. 1999. V. 400, № 6741. P. 219–220.
823. Wood B., Turner A. Out of Africa and into Asia // Nature. 1995. V. 378, № 6554. P. 239–240.
824. Wray G. A., Levinton J.S., Shapiro L.H. Molecular Evidence for Deep Precam-brian Divergences Among Metazoan // Science. 1996. V. 274, № 5287. P. 568–573.
825. Wynn T., McGrew W.C. An ape’s view of the Oldowan // Man. 1989. V. 24, № 3. P. 383–398.
825a. Xie S., Pancost R. D., Yin H., Wang H., Evershed R. P. Two episodes of micro-bial change coupled with Permo/Triassic faunal mass extinction // Nature. 2005. V. 434, № 7032. P. 494–497.
826. Xu X., Norell M. A. A new troodontid dinosaur from China with avian-like sleeping posture // Nature. 2004. V. 431, № 7010. P. 838–841.
827. Xu X., Norell M.A., Kuang X., Wang X., Zhao Q., Jia C. Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids // Nature. 2004. V. 431, № 7009. P. 680–684.
828. Xu X., Zhou Z.-h., Prum R. O. Branched integumental structures in Sinornitho-saurus and the origin of feathers // Nature. 2001. V. 410, № 6825. P. 200–204.
829. Xu X., Zhou Z., Wang X., Kuang X., Zhang F., Du X. Fourwinged dinosaurs from China // Nature. 2003. V. 421, № 6921. P. 335–340.
830. Yahil A. Meet your cosmic neighbour // Nature. 1996. V. 379, № 6565. P. 489.
831. Yellen J. E. The longest human record // Nature. 1986. V. 322, № 6082. P. 774.
832. Zehavi I., Dekel A. Evidence for a positive cosmological constant from flows of galaxies and distant supernovae // Nature. 1999. V. 401, № 6750. P. 252–254.
833. Zhang F., Zhou Z. Leg feathers in an Early Cretaceous bird // Nature. 2004. V. 431, № 7011. P. 925.
834. Zhu M., Ahlberg P.E., Zhao W., Jia L. First Devonian tetrapod from Asia // Nature. 2002. V. 420, № 6917. P. 760–761.
835. Zhu M., Yu X. A primitive fish close to the common ancestor of tetrapods and lungfish // Nature. 2002. V. 418, № 6899. P. 767–770.
836. Zhu M., Yu X., Janvier P. A primitive fossil fish sheds light on the origin of bony fishes // Nature. 1999. V. 397, № 6720. P. 607–610.
837. Zhu Z.-H., Fujimoto M.-K. Constraints on the Cardassian scenario from the expansion turnaround redshift and the Sunyaev – Zeldovich/X-ray data // The Astro-physical Journal. 2004. V. 602, № 1, part 1. P. 12–17.
838. Zimmer C. Fossils Give Glimpse of Old Mother Lamprey // Science. 1999. V. 286, № 5442. P. 1064–1065.
839. Zollikofer C. P. E., Ponce de Leô
n M. S., Lieberman D. E., Guy F., Pilbeam D., Likius A., Mackaye H. T., Vignaud P., Brunet M. Virtual cranial reconstruction of Sahelanthropus tchadensis // Nature. 2005. V. 434, № 7034. P. 755–759.
840. Zuckerman S. The social life of monkeys and apes. L.: Kegan, Trench, Trubner & Co., 1932. XII, 357, 20 p.
|