Реферат на тему:
«Птицы. Особенности строения»
2009
Птицы — высокоорганизованные теплокровные
животные, приспособленные к полету. Благодаря большой численности и широкому распространению на Земле они играют исключительно важную и многообразную роль в природе и хозяйственной деятельности человека. Известно свыше 9 тыс. современных видов птиц. В фауне Беларуси насчитывается 305 видов.
Общими чертами организации птиц в связи с их приспособленностью к полету являются следующие:
1. Туловище обтекаемой формы. Передние конечности преобразованы в орган полета — крылья,
задние конечности служат опорой туловищу и для передвижения.
2. Кожа тонкая, сухая,
лишенная желез. Единственная копчиковая железа расположена в хвостовом отделе. Кожа имеет роговые образования в виде перьев,
создающих летательные поверхности и защищающих тело от потерь тепла.
3. Кости скелета тонкие, прочные, в трубчатых костях имеются воздушные полости, облегчающие их массу. Череп образован полностью срощенными, без швов, костями. Все отделы позвоночника (кроме шейного) неподвижны. Грудина у летающих птиц с выступом впереди — килем,
к которому прикрепляются мощные летательные мышцы. В скелете задних конечностей развита длинная цевка,
увеличивающая длину шага птицы.
4. Мышечная система сильно дифференцирована. Самые крупные мышцы — грудные,
опускающие крыло. Хорошо развиты подключичные, межреберные, шейные, подкожные
и мышцы ног.
Движения птиц быстрые и разнообразные: ходьба, бег, прыжки, лазание, плавание. Виды полета —машущий
и парящий.
Птицы многих видов способны совершать дальние перелеты.
5. Особенности строения пищеварительной системы связаны с необходимостью быстрого расщепления больших объемов пищи и облегчения массы пищеварительного тракта. Это достигается благодаря отсутствию зубов, участию клюва и языка в добывании пищи, размягчению ее в расширенной части пищевода —зобе,
смешиванию пищи с пищеварительными соками железистого отдела
желудка и перетиранию ее, как на жерновах, в мышечном отделе
желудка, и укорочению задней кишки, заканчивающейся клоакой. Строение клюва и языка у птиц разнообразное и отражает их пищевую специализацию.
6. Органы дыхания — легкие.
У летящей птицы дыхание двойное:
газообмен в легких осуществляется как при вдохе, так и при выдохе, когда атмосферный воздух из воздушных мешков
поступает в легкие. Благодаря двойному дыханию птица во время полета не задыхается.
7. Сердце четырехкамерное,
все органы и ткани снабжаются чистой артериальной кровью. В результате интенсивного процесса жизнедеятельности вырабатывается много тепла, которое удерживается перьевым покровом. Поэтому все птицы — теплокровные
животные с постоянной температурой тела.
8. Органы выделения и виды конечных продуктов азотистого обмена такие же, как и у пресмыкающихся. Отсутствует лишь мочевой пузырь в связи с необходимостью облегчения массы тела птицы.
9. Как и у всех позвоночных, головной мозг птиц имеет пять отделов. Наиболее развиты большие полушария переднего мозга,
покрытые гладкой корой, и мозжечок,
благодаря которым птицы обладают хорошей координацией движений и сложными формами поведения. Ориентировка птиц в пространстве осуществляется с помощью острого зрения и слуха.
10. Птицы раздельнополы, большинству видов свойственен половой диморфизм. У самок развит только левый яичник.
Оплодотворение внутреннее, развитие прямое. Птицы большинства видов откладывают яйца в гнезда,
обогревают их теплом своего тела (насиживание), вылупившихся птенцов выкармливают. В зависимости от степени развитости вылупившихся из яиц птенцов выделяют гнездовых
и выводковых
птиц.
Особенности строения и жизнедеятельности.
У птиц голова небольшая, шея длинная и чрезвычайно подвижная. Челюсти лишены зубов, вытянуты и образуют клюв, одетый роговым чехлом. Форма клюва сильно варьирует в связи с разнообразием пищевых объектов. По бокам головы расположены крупные глаза, а ниже их имеются наружные слуховые отверстия. Передние конечности превращены в летательный орган — крылья. Задние конечности имеют разнообразное строение, которое зависит от условий обитания и способов добывания пищи. Нижняя часть ног и пальцы покрыты роговыми чешуями. Хвост короткий, снабжен веером рулевых перьев, причем у разных птиц неодинакового строения.
Кожа
птиц тонкая, сухая, лишена желез. Исключение составляет лишь копчиковая железа, расположенная под корнем хвоста. Она выделяет жиросодержащий секрет, которым птица смазывает перья при помощи клюва. Железа сильно развита у водоплавающих птиц. Кожа их покрыта своеобразным роговым покровом, состоящим из перьев. У летающих птиц перья отмечены лишь на определенных участках кожи, а у нелетающих равномерно покрывают все тело.
ОПИСАНИЕ СКЕЛЕТА
В скелете птиц отчетливо выражены черты, связанные с приспособлением к полету. Плоские кости обычно очень тонкие, губчатые. В больших трубчатых костях хорошо развиты полости, заполненные или костным мозгом, или воздухом. Все это обеспечивает повышенную прочность скелета и заметно облегчает его. Нужно, однако, иметь в виду, что общая масса скелета птиц составляет 8—18% от массы тела, т. е. примерно столько же, сколько и у млекопитающих (6—14% веса тела), хотя у последних кости толще, а воздушные полости в них отсутствуют. Это объясняется тем, что у птиц значительно больше относительная длина скелетных элементов конечностей и некоторых других костей.
Скелет птиц подразделяется на осевой скелет, грудную клетку (грудина и ребра), череп, скелет конечностей и их поясов.
Осевой скелет. Осевой скелет, или позвоночный столб (column. vertebralis), как и у пресмыкающихся, разделяется на пять отделов шейный, грудной, поясничный, крестцовый и хвостовой.
Шейный отдел очень подвижен (птицы легко поворачивают голов на 180°, некоторые даже на 270°). Число шейных позвонков варьирует в широких пределах: от 11 (попугаи) до 23—25 (лебеди, утки); у вороны, голубя 14 шейных позвонков.
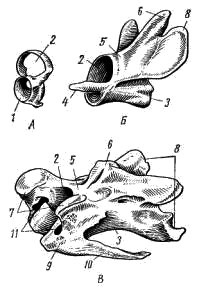
Рис. 1. Шейные позвонки вороны.
А — атлант; Б — эпистрофей; В — средний шейный позвонок:
1 — сочленовная ямка для затылочного мыщелка, 2 —канал для спинного мозга,
3 — тело позвонка, 4 — зубовидный отросток, 5 — верхняя дуга, 6 — остистый отросток,
7 — передние сочленовные отростки, 8 — задние сочленовные отростки,
9 — поперечный отросток, 10 — шейное ребро, 11 — позвоночное отверстие
Первый шейный позвонок — атлас, или атлант (atlas; рис. 105, А
) имеет форму кольца. На нижней части его передней поверхности есть глубокая сочленовная ямка, в которую входит затылочный мыщелок черепа; на нижней задней поверхности — сочленовная плоскость для соединения с телом второго шейного позвонка. Просвет полости кольца поперечной сухожильной, обычно окостеневающей связкой разделен на две части: через верхнюю проходит спинной мозг, а в нижней помещается зубовидный отросток второго позвонка.
Второй шейный позвонок — эпистрофей (epistrophaeus; рис. 1, Б
) имеет хорошо развитое тело с направленным вперед зубовидным отростком (рис. 1, 4
). По происхождению зубовидный отросток, видимо, — тело первого позвонка, обособившееся от него и сросшееся с телом второго позвонка. Сбоку расположены поперечные отростки; к ним крепятся шейные мышцы, соединяющие второй позвонок с затылочным отделом черепа и с другими шейными позвонками. Над телом позвонка расположена хорошо развитая верхняя дуга (рис. 1, 5
), через просвет которой проходит спинной мозг.
Остальные шейные позвонки имеют гетероцельное строение: довольно длинное тело каждого позвонка (corpus vertebrae; рис. 1, 3
) на передней и задней стороне имеет седловидные сочленовные поверхности. Такой характер сочленения обеспечивает значительную подвижность позвонков друг относительно друга, как в вертикальной, Так и в горизонтальной плоскостях. Это обеспечивает наряду со сложной системой дифференцированных шейных мышц большую подвижность шеи, что очень важно для птиц (компенсация неподвижного скелета туловища, возможность менять положение центра тяжести в полете, сгибая и выпрямляя шею, возможность сложных движений головы при чистке оперения, постройке гнезда и т. п.).
Над телом позвонка расположена верхняя дуга (arcus neuralis; рис. 1, 5
), заканчивающаяся остистым отростком (processus spinosus; рис. 1, 6
); в канале дуги проходит спинной мозг. От передней и задней поверхности дуги отходят парные сочленовные отростки (рис. 1, 7, 8
), соединяющиеся с такими же отростками соседних позвонков. Такой тип сочленения обеспечивает прочность соединения позвонков друг с другом при сохранении значительной подвижности. От боковой поверхности тел позвонков отходят короткие поперечные отростки (processus transversus; рис. 1, 5
). Птицы, как и пресмыкающиеся, имеют шейные ребра (рис. 1, 10
); они рудиментарны и только последние одно-два шейных ребра достаточно длинны и подвижны (рис. 2, 5
), но не доходят до грудины. Шейное ребро прирастает своей головкой к телу позвонка, а бугорком — к поперечному отростку. Таким путем образуются позвоночные отверстия (foramen transversarium; рис. 1, 1
), создающие с обеих сторон шеи канал, по которому проходят позвоночная артерия и шейный симпатический нерв. Эта костная защита артерии и нерва особенно важна при длинной, очень подвижной шее.

Рис. 2. Скелет туловища вороны сбоку:
1 — последние шейные позвонки, 2 — сросшиеся грудные позвонки (спинная кость),
3 — шейные ребра, 4 — грудные ребра, 5 — крючковидный отросток, 6 — тело грудины,
7 — киль, грудины, 8 — сложный крестец, 9 — подвижные хвостовые позвонки,
10 — пигостиль, 11 — коракоид, 12 — вилочка, 13 — лопатка,
14 — суставная впадина для головки плеча, 15 — подвздошная кость,
16 — седалищная кость, 17 — лобковая кость,
18 — вертлужная впадина для сочленения с головкой бедра
Грудной отдел у вороны представлен шестью позвонками (у других видов птиц их число варьирует от 3 до 10), которые, как и у большинства птиц, срастаются друг с другом, образуя спинную кость (dorsale; рис. 2, 2
).
К грудным позвонкам подвижно причленяются грудные ребра (costa; рис. 2, 4
); число их пар равно числу грудных позвонков. Это плоские, слегка изогнутые костные пластинки, состоящие из двух подвижно соединенных друг с другом отделов — спинного и брюшного. Верхний конец спинного отдела ребра имеет два сочленовных отростка: бугорок, причленяющийся к поперечному отростку, и головку, сочленяющуюся с телом позвонка. Нижний конец брюшного отдела ребра подвижно сочленяется с телом грудины (sternum рис. 2, 6
). Подвижное соединение ребер с грудиной и позвоночным столбом вместе с подвижным сочленением спинного и брюшного отделов ребра обеспечивает возможность значительных изменений объема полости тела путем опускания грудины вниз и поднимания ее кверху. Эта подвижность возможна благодаря сильно развитой реберной мускулатуре. Жесткость грудной клетки усиливается костными образованиями — крючковидными отростками (processus uncinatus; рис. 2, 5
) ребер, укрепляющимися на спинном отделе и налегающими на последующее ребро.
Грудина, или грудная кость (sternum; рис. 2, 6
) — широкая и длинная, вогнутая изнутри костная пластинка, несущая по средней линии высокий костный гребень — киль грудины (crista sterni; рис. 2, 7
). Резкое увеличение поверхности грудины, обусловленное как ее разрастанием, так и образованием киля, — прямое приспособление к полету; оно обеспечивает возможность прикрепления мощных мышц, приводящих в движение крыло. На переднем конце грудины по бокам киля расположены большие сочленовные площадки, обеспечивающие прочное прикрепление к телу грудины коракоидов (рис. 2, 11
). На боковых краях грудины размещаются мелкие углубления, которые служат для сочленения с нижними концами брюшных отделов ребер.
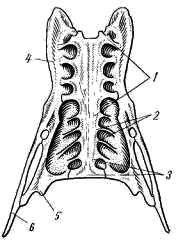
Рис. 3. Сложный крестец вороны (вид снизу):
1 — поясничные позвонки, 2 — крестцовые позвонки, 3 — хвостовые позвонки,
4 — подвздошная кость, 5 — седалищная кость, 6 — лобковая кость
За грудным отделом позвоночного столба расположен сложный крестец (synsacrum; рис. 2, 8
; рис. 3), представляющий собой слившиеся в общую монолитную кость все поясничные (у ворон и голубей 6), все крестцовые (почти у всех видов птиц 2) и часть хвостовых (3—8) позвонков. Сложный крестец неподвижно срастается с последним грудным позвонком. Границы между отделами в сложном крестце провести трудно; крестцовые позвонки (рис. 3, 2
) бывают заметны лишь по отверстиям между их поперечными отростками и подвздошными костями. Кости тазового пояса также неподвижно срастаются с позвонками сложного крестца.
Благодаря срастаниям позвонков туловищный участок осевого скелета птиц представляет собой прочное, монолитное образование, обеспечивающее неподвижность туловища, что крайне важно при полете. Образование сложного крестца, слившегося с костями тазового пояса, создает прочную опору для задних конечностей, на которые давит вся тяжесть тела при движении птицы по земле (рис. 3).
Число свободных, подвижно соединенных друг с другом хвостовых позвонков (рис. 2, 9
) у птиц невелико (от 5 до 9; у вороны и голубя — обычно 7). Последние хвостовые позвонки (4—8) сливаются в вертикально уплощенную пластинку — копчиковую кость (pygostyle; рис. 2, 10
), к боковым поверхностям которой веером прикрепляются основания рулевых перьев. Укорочение хвостового скелета увеличивает общую компактность тела (важно аэродинамически). Характер прикрепления рулевых перьев позволяет использовать хвост в полете (добавочная несущая плоскость) и при посадке (торможение).
Череп. Череп птиц тропибазального типа (мозг не лежит между глазницами). По сравнению с пресмыкающимися резко бросается в глаза значительное увеличение объема мозговой полости, большие глазницы, появление клюва, полное исчезновение зубов, у большинства птиц — заметное смещение вниз большого затылочного отверстия. Кости черепа настолько тонки, что соединение их друг с другом при помощи швов становится невозможным. Поэтому у птиц большинство костей черепа сливается друг с другом; границы между отдельными костями хорошо видны только на черепе птенцов. Тонкие кости и редукция зубов сильно облегчают скелет головы. В то же время благодаря сложной губчатой внутренней структуре кости черепа сохраняют большую прочность.
Череп (cranium) делится на мозговой череп (neurocranium), включающий собственно мозговую (черепную) коробку, носовой отдел и область глазниц, и висцеральный череп (splanchnocranium), к которому относятся челюсти (клюв), кости нёба и подъязычный аппарат.

Рис. 4. Череп вороны.
А — сбоку; Б — снизу; В — сверху:
1 — большое затылочное отверстие, 2 — основная затылочная кость,
3 — боковая затылочная кость, 4 — верхняя затылочная кость, 5 — затылочный мыщелок,
6 — теменная кость, 7 —лобная кость, 8 — носовая кость, 9 — чешуйчатая кость,
10 — наружный слуховой проход, 11 — боковая клиновидная кость,
12 — средняя обонятельная кость, 13 — предлобная кость, 14 — предчелюстная кость,
15 — верхнечелюстная кость, 16 - скуловая кость, 17 — квадратно-скуловая кость,
18 — квадратная кость, 19 — основная височная кость, 20 — клювовидный отросток парасфеноида, 21 — сошник, 22 — нёбная кость, 23 — крыловидная кость, 24 — зубная кость,
25 — сочленовная кость, 26 — угловая кость
Начнем рассмотрение черепа с затылочной области. На задне-нижней стенке черепной коробки находится большое затылочное отверстие (foramen occipitale magnum; рис. 4, 1
), окруженное четырьмя сросшимися затылочными костями. Вентрально расположена основная затылочная кость (basioccipitale; рис. 4, 2
), по бокам — парные боковые затылочные кости (occipitale laterale или exooccipitale; рис. 4, 3
). Основная и боковые затылочные кости образуют единственный, как и у пресмыкающихся, затылочный мыщелок (condylus occipitalis; рис. 4,5), сочленяющий череп с первым шейным позвонком. Над затылочным отверстием располагается верхняя затылочная кость (supraoccipitale; рис. 4, 4
).
Спереди от верхней и боковых затылочных костей лежат парные теменные кости (parietale; рис. 4, 6
). Крышу черепа над глазницами, верхние и задние стенки глазниц и всю переднюю часть мозговой коробки образуют длинные и широкие лобные кости (frontale; рис. 4, 7
). Впереди лобных костей у основания клюва лежат парные носовые кости (nasale; рис. 4, 8
) с двумя отростками: верхний идет вперед сбоку от гребня клюва и ограничивает носовое отверстие, сверху, челюстной — вперед и вниз к верхнечелюстной кости и ограничивает носовое отверстие сзади. Боковую часть мозговой коробки образуют парные крупные чешуйчатые кости (squamosum; рис. 4, 5
), лежащие книзу от лобных и спереди от теменных и боковых затылочных костей. Верхняя височная дуга у птиц отсутствует. Чешуйчатая кость прикрывает ушные кости, которые срастаются вместе и образуют костные стенки среднего и внутреннего уха; снаружи они обычно не видны. При хорошей препарировке черепа в центре ушного прохода (рис. 4, 10
) бывает виден конец тонкой столбчатой косточки (или стремечка, stapes), образовавшийся в результате окостенения подвеска (hyomandibulare) — верхнего элемента подъязычной дуги.
Ниже лобных костей переднюю часть мозговой коробки (являющуюся также задней стенкой глазницы) образуют небольшие парные боковые клиновидные кости (laterosphenoidem; рис. 4, 11
). Практически почти всю тонкую межглазничную перегородку образует непарная средняя обонятельная кость (mesethmoideum; рис. 4, 12
). Передние стенки глазницы образуют парные (по одной с каждой стороны) предлобные кости (praefrontale; рис. 4, 13
); у вороны и других воробьиных птиц они очень малы, и основную часть передней стенки глазницы заполняет отросток средней обонятельной кости.
Клюв птицы состоит из двух частей: надклювья, образованного предчелюстными, верхнечелюстными и носовыми костями, и подклювья. Большую часть надклювья составляют слившиеся парные предчелюстные кости (praemaxillare; рис. 4, 14
). Спереди они образуют выпуклую сверху л слегка вогнутую снизу вершину клюва, а назад продолжаются в виде трех парных отростков:
1) лобные отростки соединяются с верхними отростками носовых костей и образуют верхний гребень клюва (ограничивают носовое отверстие сверху и спереди);
2) челюстные отростки отграничивают носовое отверстие сбоку;
3) нёбные отростки образуют костное дно надклювья (они входят в состав твердого нёба).
Небольшие верхнечелюстные кости (maxillare; рис. 4, 15
) образуют лишь заднюю нижнюю часть надклювья, соединяясь спереди с челюстными отростками предчелюстных костей; снизу к ним примыкают (сливаясь) челюстные отростки нёбных костей.
От заднего края верхнечелюстной кости отходит назад тонкая костная перекладина, состоящая из двух слившихся костей — скуловой (jugale; рис. 4, 16
) и квадратно-скуловой (quadrato-jugale; рис. 4, 17
). По своему положению это типичная нижняя дуга, поэтому череп птиц относят к диапсидному типу с редуцированной верхней дугой. Квадратно-скуловая кость причленяется к квадратной кости (quadratum; рис. 4, 18
). Нижний конец квадратной кости образует суставную поверхность для причленения нижней челюсти, а другой удлиненный ее конец подвижно сочленяется с чешуйчатой и переднеушной костями.
Посмотрим теперь на череп снизу. В основании черепа, впереди основной затылочной кости (рис. 4, 2
) лежит небольшая основная клиновидная кость (basisphenoideum). Она полностью прикрыта широкой основной височной костью (basitemporale; рис. 4, 19
) — производной парасфеноида. Передняя часть парасфеноида сохраняется в виде направленного вперед узкого клювовидного отростка (rostrum parasphenoidei; рис. 4, 20
). У его переднего конца лежит сошник (vomer; рис. 4, 21
). По бокам сошника располагаются хоаны — внутренние отверстия ноздрей.
Нёбные отростки предчелюстных и верхнечелюстных костей сливаются с удлиненными парными нёбными костями (palatinum; рис. 4, 22
) и образуют костное дно надклювья. Задние, имеющие сложный профиль, концы нёбных костей налегают на клювовидный отросток парасфеноида. В этом месте к нёбным костям причленяются (суставом) парные крыловидные кости (pterygoideum; рис. 4, 23
), задние концы которых также суставом соединяются с квадратными костями.
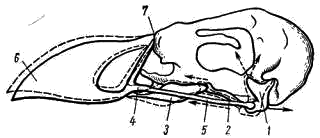
Рис. 5. Схема кинетизма черепа птицы.
Сплошной линией и пунктиром показано изменение положения надклювья при движении костного нёба: 1 — квадратная кость, 2 — крыловидная кость, 3 — нёбная кость, 4 — скуловая кость, 5 — квадратно-скуловая кость, 6 — надклювье, 7 — место перегиба костей
Эти элементы твердого нёба имеют очень важное значение для движения клюва (рис. 5). Если сокращаются мышцы, соединяющие направленный вперед длинный глазничный отросток квадратной кости со стенками глазницы, то направленный вниз конец квадратной кости несколько подвигается вперед и толкает вперед как крыловидные и нёбные кости (место сочленения этих костей может скользить по клювовидному отростку парасфеноида, на который они налегают), так и квадратно- скуловую и скуловую. Давление по этим двум костным мостикам передается на основание надклювья, благодаря чему вершина надклювья приподнимается кверху. При этом прогибаются кости у основания вершины надклювья, в области «переносицы». Перегиб облегчает очень сильное истончение костей в этом месте; у ряда видов здесь сохраняется хрящевая перемычка или даже образуется настоящий сустав. При сокращении мышц, соединяющих череп с нижней челюстью, нижний конец квадратной кости сдвигается назад, оттягивая на себя эти костные связи, и вершина клюва сдвигается книзу.
Описанный сложный костный механизм твердого нёба (основа его — подвижно соединенная с черепом сложной формы квадратная кость) дополняется системой дифференцированных жевательных мышц. Все это обеспечивает возможность довольно разнообразных движений клюва, облегчающих захват добычи, чистку оперения, постройку сложного гнезда и т. п. Возможность дифференцированных движений клюва выработалась, видимо, в связи с преобразованием передних конечностей в крылья, выполняющие только функцию полета.
Подклювье — нижняя челюсть — представляет собой ряд слившихся костей, из которых наиболее крупные зубная (dentale; рис. 4, 24
), сочленовная (articulare; рис. 4, 25
) и угловая (angulare; рис. 4, 26
). Челюстной сустав образуют, соединяясь друг с другом, квадратная и сочленовная кости.
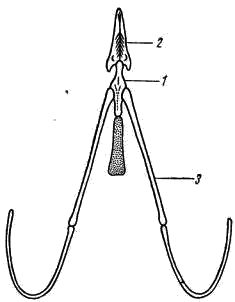
Рис. 6. Подъязычный аппарат вороны:
1 — тело, 2 — его передний отросток, служащий основанием языка, 3 — рожки
Остатки гиоида и жаберных дуг превращаются в подъязычный аппарат (рис. 6). У некоторых птиц (например, дятлов) благодаря большой длине рожков и общей подвижности подъязычного аппарата язык может выдвигаться вперед почти на длину клюва, что облегчает захват добычи.
Конечности и их пояса. Пояс передних конечностей (плечевой пояс) птиц образован парными коракоидами, лопатками и ключицами.
Мощные удлиненные коракоиды (coracoideum; см. рис. 6, 11
) своими расширенными нижними концами прочно соединяются малоподвижными суставами с передним краем грудины. Между передними концами коракоидов расположена прикрепляющаяся к ним вилочка (furcula, см. рис. 6, 12
), возникшая путем слияния обеих ключиц. Длинные и узкие лопатки (scapula; см. рис. 6, 13
) лежат над ребрами; их передние концы прочно соединены со свободными концами коракоидов. В месте соединения лопатки и коракоида расположена довольно глубокая суставная впадина (см. рис. 6, 14
), в которую входит головка плечевой кости.
Мощность костей плечевого пояса и их прочное соединение с грудиной обеспечивают опору для крыльев в полете. Удлинение коракоидов увеличивает площадь прикрепления мышц крыла, а также позволяет вынести вперед плечевой сустав, что обеспечивает и удобное складывание крыла в покое, и аэродинамически выгодное его положение в полете, когда центры площадей крыльев лежат на линии, проходящей через центр тяжести птицы. Расположенная между свободными концами коракоидов вилочка обычно играет роль пружины, амортизирующей резкие толчки при взмахах крыла в полете.
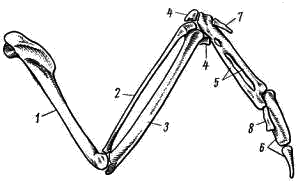
Рис. 7. Скелет крыла вороны:
1 — плечевая кость, 2 — лучевая кость, 3 — локтевая кость, 4 — самостоятельные косточки запястья, 5 — пряжка (слившиеся косточки запястья и пясти), 6 — фаланги II пальца,
7 — единственная фаланга I пальца, 8 — единственная фаланга III пальца
Скелет передней конечности, превратившейся в крыло, подвергся значительным изменениям. Плечо (humerus; рис. 7, 1
) — мощная трубчатая кость, проксимальный конец которой образует уплощенную головку плеча, входящую в суставную впадину плечевого сустава. Характер суставных поверхностей заметно ограничивает возможность вращательных движений в плечевом суставе, что обеспечивает устойчивость крыла в полете. Мощные эпифизы дистального конца плеча образуют суставные поверхности для сочленения с двумя костями предплечья: более прямой и тонкой лучевой (radius; рис. 7, 2
) и более мощной, слегка изогнутой локтевой костью (ulna; рис. 7, 3
). На поверхности локтевой кости видны бугорки — места прикрепления очинов второстепенных маховых перьев. Характер суставных поверхностей локтевого сустава обеспечивает прочное соединение костей плеча и предплечья и ограничивает вращательные движения и в этом суставе. При этом сохраняется большая подвижность в одной плоскости — плоскости крыла, что дает возможность птице складывать крыло в покое и изменять его площадь при изменении режима полета.
Особенно резкие изменения в связи с приспособлением к полету произошли в строении кисти. В проксимальном отделе запястья (carpus) сохраняются только две самостоятельные косточки (рис. 111, 4
). Они связками почти неподвижно соединяются с костями предплечья. Подвижный сустав расположен между ними и остальными костями запястья, поэтому его называют интеркарпальным. Остальные кости запястья и все кости пясти (metacarpus) сливаются в единое образование— пряжку (carpometacarpus; рис. 7, 5
). Резко редуцируется скелет пальцев. Сохраняются две фаланги II пальца (рис. 7, 6
), продолжающие ось пряжки. От I пальца сохраняется одна фаланга (рис. 7, 7
), к ней прикрепляется пучок перьев, образующий крылышко. III палец представлен также одной фалангой (рис. 7, 8
), прикрепленной к основанию первой фаланги II пальца.
Эти преобразования в скелете кисти (образование carpometacarpus, редукция пальцев) обеспечивают прочную опору для первостепенных маховых перьев — части крыла, несущей в полете наибольшую нагрузку.
Тазовый пояс состоит из трех пар срастающихся вместе костей: Широкая и длинная подвздошная кость (ilium; см. рис. 6, 15
; рис. 7, 4
) на большом протяжении срастается со ложным крестцом. К ее наружному краю прирастает седалищная кость (ischium; см. рис. 6, 16
; рис. 7, 5
), а ниже расположена направленная назад палочкообразная лобковая кость (pubis; см. рис. 6, 17
; рис. 7, 6
). Все три кости таза образуют вертлужную впадину (acetabulum; см. рис. 6, 18
), в которую входит головка бедра.
Большая поверхность тазовых костей и их прочное (неподвижное) соединение с осевым скелетом обеспечивают прочную опору для прикрепления мощных мышц. Эти особенности тазового пояса определяются тем, что передние конечности выполняют только функцию полета, а при движении по земле вся тяжесть тела поддерживается лишь задними конечностями.
Лобковые и седалищные кости правой и левой сторон не соединяются друг с другом на брюшной поверхности. Поэтому таз птиц называют открытым; он дает возможность откладывать крупные яйца. Можно предполагать, что открытый таз, увеличивающий подвижность брюшной стенки в тазовой области, способствует интенсификации дыхания в полете.
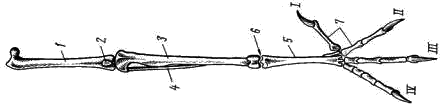
Рис. 8. Скелет задней конечности вороны:
1 — бедренная кость, 2 — коленная чашечка, 3 — голено-предплюсна (tibio-tarsus),
4 — малая берцовая кость, 5 — цевка (слившиеся кости предплюсны и плюсны),
6 – интертарзальный сустав, 7 — фаланги пальцев; I — IV — пальцы
Скелет задней конечности представлен мощными трубчатыми костями. Бедро (femur; рис. 8, 1
) на проксимальном конце имеет хорошо развитую округлую головку для сочленения с тазовым поясом. На дистальном конце образуются рельефные суставные поверхности для сочленения с костями голени. В области коленного сустава в мышечном сухожилии лежит коленная чашечка (patella; рис. 8, 2
).
Основной элемент голени — массивная большая берцовая кость, с нижним дистальным концом которой сливаются две проксимальные кости предплюсны; образуется костный комплекс, представляющий собой голено-предплюсну, или тибио-тарзус (tibio-tarsus; рис. 8, 3
). Малая берцовая кость (fibula; рис. 8, 4
) сильно редуцирована и в виде маленькой тоненькой косточки прирастает к верхней части наружной поверхности большой берцовой кости.
Дистальные элементы предплюсны (tarsus) и все элементы плюсны (metatarsus) сливаются у птиц в единую кость — цевку, или плюсну-предплюсну (tarso-metatarsus; рис. 8, 5
). Благодаря этому в конечности появляется добавочный рычаг. Подвижное сочленение располагается между двумя рядами костей предплюсны (между костями приросшими к голени, и костями, слившимися с плюсной), поэтому, как и у пресмыкающихся, этот сустав называется интертарзальным (рис. 8, 6
).
На дистальном конце цевки образуются хорошо выраженные суставные поверхности для причленения фаланг пальцев (phalanges digitorum; рис. 8, 7
). У подавляющего большинства птиц в задней конечности развиваются четыре пальца, из которых I направлен назад, а II, III, IV — вперед.
Мощные длинные кости скелета задних конечностей, появление добавочного рычага (образование tarso-metatarsus), резкий рельеф суставных поверхностей — все это обеспечивает прочность и подвижность задних конечностей.
Заключение
Морфологически класс птиц во многих чертах сохраняет большое сходство со своими предками — пресмыкающимися. На это указывает целый ряд признаков: почти полное отсутствие кожных желез, то, что дистальные отделы задних конечностей покрыты роговыми чешуями, роговой покров клюва, диапсидного типа череп (с редуцированной, однако, верхней дугой), имеющий только один затылочный мыщелок, состав костей черепа, то, что имеются шейные ребра и крючковидные отростки грудных ребер, открытый таз (как у некоторых ископаемых динозавров), большое сходство строения мочеполовой системы и периферических отделов кровеносной системы, присутствие клоаки, размножение откладыванием яиц, сходство эмбрионального развития и т. п.
В процессе эволюции птицы обособились от рептилиеобразных предков путем многих, но часто относительно небольших преобразований морфологических структур, обеспечиваюших существенную интенсификацию физиологических функций и тем самым резко повысивших общий уровень организации птиц по сравнению с пресмыкающимися. Полет и высокая общая подвижность птиц связаны с большим расходом энергии, требующим быстрой компенсации. Поэтому в эволюции птиц большое значение имело становление особенностей организации, связанных не только с обеспечением физической возможности полета, но и с общим повышением интенсивности обмена веществ.
Возможность полета птиц обеспечивается целым рядом прямых приспособлений. К ним относятся: компактность тела, жесткость и неподвижность туловищного отдела осевого скелета и редукция его хвостового отдела, резкое увеличение размеров грудины и образование на ней киля, прочность пояса передних конечностей (в том числе и образование специального амортизирующего устройства — вилочки), строение скелета крыла (включая и особенности суставных поверхностей и особенно резкие изменения в строении кисти), мощность мускулатуры, обеспечивающая движения крыла, развитие перьев — легких и прочных роговых образований, создающих несущие плоскости крыльев и хвоста и обеспечивающих обтекаемость тела в по лете, редукция зубов, облегчающая скелет черепа, общее увеличение объема головного мозга и особенно сильное развитие мозжечка — центра равновесия и координации движений, увеличение массы нервных клеток спинного мозга в области крыльев и т. д.
Клюв с его подвижным скелетным механизмом и дифференцированной мускулатурой стал очень тонко действующим инструментом, способным к сложным движениям. Образование клюва и одновременно увеличение длины и подвижности шеи освободило передние конечности от многих добавочных функций (захват и удерживание добычи, чистка покровов и т. п.) и тем самым способствовало их превращению только в орган полета — крылья. Удлинение шеи и возрастание ее подвижности компенсировало неподвижность туловища и дало возможность, изменяя положение головы, в каких-то пределах менять положение центра тяжести тела, что важно при изменении режима полета.
Преобразование передних конечностей в орган полета — крылья сопровождалось укреплением тазового пояса и скелета задних конечностей, а также возрастанием массы их мышц, обеспечивающих возможность «двуногого» движения на земле, по ветвям деревьев и т. п.
Полет требует затраты большого количества энергии. Например у голубя, летящего со скоростью 70 км/ч, расход энергии в 27 раз больше, чем в покое. Резкое возрастание энергетических возможностей обеспечивается изменениями многих систем органов.
Легкие птиц относительно малы по объему, но велика их губчатость (и тем самым большая поверхность окисления). Сложная система воздушных мешков, связанная с легкими, занимает до 20% объема тела — она резко увеличивает объем вдыхаемого воздуха и дает возможность насыщать кровь кислородом и во время вдоха, и во время выдоха. Чем интенсивнее полет, тем чаще дыхание, что обеспечивает более интенсивное насыщение крови кислородом и удаление из нее углекислоты при ускорении движения. Одновременно воздушные мешки осуществляют интенсивную теплоотдачу во время полета, предохраняя организм летящей птицы от перегрева.
Полное разделение большого и малого кругов кровообращения резко увеличивает возможность насыщения тканей кислородом. Сердце у птиц относительно очень велико — составляет 0,8—2,5% от веса тела (у рептилий 0,2—0,3%; у млекопитающих 0,2—1,5%). Относительно очень велика у птиц и частота сердцебиений: сердце крупных птиц сокращается 140—200 раз в 1 мин, мелких — до 500—600. В полете частота сердцебиений резко возрастает: так, у голубя в покое около 170 сердцебиений в 1 мин, а в полете 350—600; у мелких птиц частота пульса в полете до 1000 и более. Большой объем сердца и высокая частота сердцебиений создают быструю циркуляцию крови по организму, обеспечивая интенсивное насыщение тканей и органов питательными веществами и кислородом и освобождение их от углекислоты и продуктов распада.
Дифференцировка желудка на два отдела — железистый и мускульный — обеспечивает механическую переработку пищи (в мускульном желудке может развиваться давление до 20—30 кг на 1 см2), что значительно увеличивает интенсивность ее переваривания. Этому же помогает свойственная ряду видов птиц способность отбрасывания погадок, т. е. удаления непереваренных остатков пищи (хитин, шерсть, кости и т. п.) из желудка через пищевод. Обеспечивает интенсивное пищеварение и высокая активность пищеварительных ферментов. Поэтому скорость переваривания пищи у птиц заметно выше, чем у пресмыкающихся и млекопитающих. Так, воробей переваривает гусеницу за 15 мин, жука — за 1 ч, зерно — за 3—4 ч (считая от времени принятия пищи до вывода ее непереваренных остатков). Интенсивное пищеварение позволяет перерабатывать большое количество пищи, обеспечивая высокие энергетические потребности организма. Особенности пищеварительной системы и органов захвата пищи наряду с большой подвижностью, высоким уровнем высшей нервной деятельности и совершенством органов чувств способствовали образованию широких пищевых связей и создали возможность использования очень большого числа кормовых объектов.
Интенсивность пищеварения, кровообращения и дыхания обеспечивает высокий уровень обмена веществ, что наряду с образованием высоко эффективного теплоизолирующего покрова из пуха и перьев приводит к теплокровности птиц (гомойотермия), т. е. к их способности поддерживать температуру тела на постоянном уровне независимо от изменения температуры окружающей среды. Косвенным показателем высокого уровня обмена веществ птиц может служить температура их тела, которая у крупных видов равна 38—40°С, а у мелких видов достигает 41—43°С.
Значительное увеличение объема головного мозга и возрастание числа нервных клеток в нем, по сравнению с рептилиями, определяют - более высокий уровень высшей нервной деятельности птиц. Это проявляется и в очень сложных системах врожденных рефлексов, определяющих поведение каждой особи, и в значительном возрастании роли индивидуального опыта, т. е. в выработке условных рефлексов.
Плодовитость птиц меньше, чем у пресмыкающихся. Увеличение размеров яиц (откладка которых обеспечивается открытым тазом) увеличивает размеры вылупившегося детеныша. Сопутствующие размножению сложные биологические явления (постройка гнезда, насиживание, выкармливание, обогрев и охрана птенцов и т. п.) обеспечивают ускорение эмбрионального и постэмбрионального развития и повышенное выживание потомства.
Совершенство органов движения (большая подвижность), гомойотермия, сложность высшей нервной деятельности и совершенство органов чувств (особенно зрения и слуха) позволили птицам широко расселиться по всему земному шару. Практически нет на земле ни одного района, включая высокие широты Арктики и Антарктики, где не встречались бы те или иные виды птиц.
Более высокий уровень организации птиц, по сравнению с пресмыкающимися, находит свое отчетливое выражение не только в значительно более широком их распространении по всему земному шару, но и в более сложных и разнообразных взаимоотношениях со средой, с условиями жизни. Так, при сезонных неблагоприятных изменениях условий жизни (наступление зимы) пресмыкающиеся и земноводные прекращают свою активность, впадают в состояние оцепенения. У птиц в целом реакции при наступлении неблагоприятных сезонов года многообразнее и шире. Часть птиц совершает миграции (протяженностью до 5—10 тыс. км и более), улетая на зимовки в благоприятные по погодным и кормовым условиям районы. Однако значительное число видов остается в умеренных и северных широтах в течение всего года. Переживать суровые условия зимы им помогают довольно разнообразные приспособления.
Для многих видов свойственна резко выраженная сезонная смена кормов: переход зимой на питание менее калорийными, но массовыми кормами (например, тетерев летом питается насекомыми, ягодами, зелеными частями растений, зимой — преимущественно почками и сережками некоторых пород деревьев). Неперелетные (оседлые) птицы совершают небольшие перемещения, концентрируясь в участках с большим количеством пищи (глухарь в сосновых лесах, где питается хвоей сосны; синицы, вороны — в населенных пунктах и т. п.). Некоторые виды осенью запасают пищу, пряча ее в укромных местах (кедровки, сойки, поползни — орехи лещины, «орешки» кедра, желуди; синицы — семена деревьев и кустарников). Оседлые птицы во время осенней линьки получают перьевой покров с лучшими теплоизолирующими свойствами, чем летний (большее развитие пуховой части пера, большее число перьев и пуха на единицу поверхности тела и т. п.); у некоторых видов зимний наряд резко отличается по окраске, помогая маскировке (белая куропатка).
У части видов отчетливо выражены и приспособительные особенности поведения: ночевки в укрытиях (тетеревинные и некоторые другие птицы — в снегу; мелкие воробьиные — в дуплах, щелях за корой и т. п.). При этом птицы некоторых видов в сильные морозы ночуют группами, взаимно обогревая в убежище друг друга (синицы, воробьи и др.).
Несмотря на резкое повышение уровня жизнедеятельности и широкие экологические возможности, открываемые гомойотермией, отчетливая специализация птиц к полету в известной мере ограничивает круг экологических приспособлений этих животных. Отсутствие столь узкой специализации у млекопитающих создало предпосылки к возникновению в процессе эволюции этого класса значительно большего разнообразия жизненных форм и более многообразных взаимоотношений со средой.
Литература
1. Гладков Н.А., Дементьев Г.П., Птушенко Е.С., Судиловская А.М. Определитель птиц СССР. М., 1964.
2. Гуртовой Н.Н., Матвеев Б.С, Дзержинский Ф.Я. Практическая зоотомия позвоночных. Птицы, млекопитающие. М., 1980.
3. Дементьев Г.П. Птицы. Руководство по зоологии, т. 6. М., 1940.
4. Дементьев Г.П. Птицы нашей страны. М., 1962.
5. Иванов А.И., Штегман Б.К. Краткий определитель птиц СССР. Л., 1964.
6. Карташев Н.Н. Систематика птиц. М., 1974.
7. Михеев А.В. Биология птиц. М., 1960.
8. Шмальгаузен И.И. Основы сравнительной анатомии. М., 1947.
9. Шульпин Л.М. Орнитология. Л., 1940.
|