Московская Государственная академия
тонкой химической технологии им. M.В. Ломоносова
Реферат
по курсу: Регуляция клеточной активности
Москва -2008
Введение
Парагормон и кальцитонин
Кальцитриол
Другие гормоны, влияющие на обмен кальция в организме
Заключение
Литература
Введение
В организме взрослого человека в костной ткани содержится более 1 кг кальция. Наиболее важной минеральной составляющей костной ткани является нерастворимый фосфат кальция в виде гидроксилапатита или карбонатапатита (Ca10
(PO4
)6
(OH)2
и Ca10
(PO4
)6
СО3
соответственно). В костях присутствуют также карбонаты других щелочноземельных элементов. За счет активности костеобразующих клеток, остеобластов, и клеток, разрушающих костную ткань, остеокластов, кальций постоянно откладывается и вновь вымывается из кости. Кальциевый обмен контролируется гормонами: кальцитонин повышает отложение кальция в костном матриксе, парагормон стимулирует мобилизацию кальция, а кальцитриол улучшает процесс минерализации.
Недостаток кальцитриола у детей приводит к заболеванию рахитом, а у взрослых может вызвать нарушение обмена веществ в костной ткани. Отрицательный баланс между процессами отложения и вымывания кальция, особенно в пожилом возрасте, вызывает заболевание остеопорозом (рис.1).
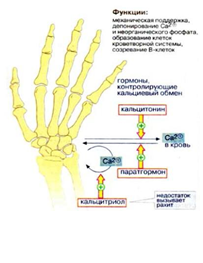
Рисунок 1. Гормоны, контролирующие кальциевый обмен
Парагормон и кальцитонин
Гормоны секретируются клетками паращитовидной железы. Местом синтеза кальцитонина является также щитовидная железа.
Парагормон был получен в частично очищенном состоянии в 1925 году из паращитовидной железы быка. Он представляет собой простую полипептидную цепь, состоящую из 84 аминокислотных остатков с молекулярной массой 9500 Да; период полураспада около 2 мин. Ген, кодирующий биосинтез парагормона, локализуется на 11-й хромосоме (11р15).
Парагормон образуется на рибосомах в виде препропарагормона – полипептида, состоящего из 115 аминокислотных остатков. В результате локального протеолиза отщепляется 31 аминокислотный остаток с N-конца и образуется активный гормон, который запасается в секреторных гранулах.
Регуляция секреции парагормона осуществляется несколькими механизмами. В течение короткого времени биосинтез парагормона регулируется ионизированным кальцием, а в течение длительного времени – 1,25(ОН)2
D3
- совместно с кальцием. Скорость секреции обратно пропорциональна концентрации в плазме крови.
Метаболизм и деградация парагормона осуществляется в основном в печени (около 62-70%), а также в почках (30-38%).
Парагормон оказывает многообразное действие в зависимости от ткани-мишени. Все это позволило L. Mallette (1991) высказать мнение, что парагормон является прогормоном, а его фрагменты обладают биологическим действием. Считается, что его аминотерминальный домен (аминокислотные остатки 1-34) ответствен за регуляцию минерального обмена посредством взаимодействия с соответствующими рецепторами в костях и почках; карбокситерминальный домен (аминокислотные остатки 53-84) – за регуляцию функции остеокластов, а средний домен (аминокислотные остатки 28-48), возможно, за транспорт кальция и фосфора
Парагормон взаимодействует с плазматическими рецепторами, которые являются гликопротеинами с молекулярной массой около 800 кДа и состоят из 585-594 аминокислотных остатков. Рецептор парагормона, как и все другие рецепторы, относящиеся к семейству рецепторов, оперирующих через G-белок, имеет 3 цепи внеклеточного фрагмента, 7 трансмембранных фрагментов и внутриклеточную часть рецептора, также представленную 3 петлями полипептидной цепочки.
Такое взаимодействие приводит к активации аденилатциклазы и повышению синтеза цАМФ, который активирует протеинкиназу, фосфолипазу С, диацилглицерин, инозитолтрифосфат и участвует в регуляции транспорта ионов кальция, натрия и калия через клеточные мембраны.
Парагормон оказывает множественное действие на костную ткань. Он опосредованно активирует ферменты коллагенозу и глюкуронидазу, что вызывает деструкцию органических компонентов кости, в частности коллагена и гликозамингликанов. В минеральных компонентах костной ткани под действием парагормона происходит солюбилизация гидроксиапатита и высвобождение в кровь кальция и фосфора.
Было установлено, что парагормон активирует процессы транскрипции в остеокластах – клетках, резорбирующих кости. Влияние парагормона на резорбцию костной ткани на пострецепторном уровне осуществляется в основном через инозитолтрифосфат и диацилглицерин, но не через цАМФ, которые образуются посредством активации фосфолипазы С и ионов Са.
Наряду с этим парагормон оказывает влияние на обмен фосфора и магния. Обмен кальция, фосфора и магния в организме показан на рис.2.
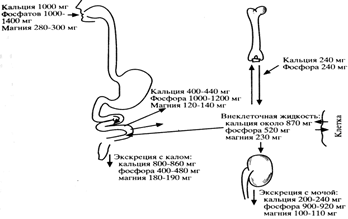
Рисунок 2. Суточный баланс и обмен кальция, фосфора и магния в организме.
Кальций всасывается в верхнем отделе тонкого кишечника. Это активный процесс, осуществляемый транспортным кальцийсвязывающим белком, который активизируется 1,25-дигидроксивитамином D. Всасывание кальция в кишечнике усиливается при увеличении поступления кислот с пищей, диете с высоким содержанием белка, саркоидозе, беременности, тогда как щелочи, глюкокортикоиды, избыток фосфатов и оксалатов снижают его всасывание в кишечнике.
Свое влияние парагормон на натрий-фосфатный котранспорт оказывает путем повышения образования цАМФ и путем активизации фосфолипазы С и образования диацилглицерина и инозитолтрифосфата.
Кальцитонин был впервые получен С. Н. Коопом и соавторами в 1962 году. Кальцитонин человека представляет собой полипептид, состоящий из 32 аминокислот с мол. м. 3000 Да с периодом полураспада около 5 мин. Гормон может образовывать путем ковалентной связи димерные и, не исключено, полимерные формы, однако биологически активной является только мономерная форма гормона. Было показано, что в процессе трансляции образуется препрокальцитонин и прокальцитонин с мол. м. около 13 кДа.
Специфическим стимулятором секреции кальцитонина является повышение концентрации кальция в крови более 2,25 ммоль/л (9 мг/100 мл). Кроме того, стимуляторами секреции кальцитонина являются катехоламины, осуществляющие свое действие через β-адренергические рецепторы, холецистокинин, глюкагон, гастрин. Глюкагон и катехоламины, взаимодействуя с рецепторами, увеличивают содержание цАМФ, который стимулирует секрецию кальцитонина, так же как и парагормона, т.е. цАМФ является внутриклеточным медиатором секреции кальцитонина. Кальцитонин метаболизируется в почках, печени и, возможно, в костной ткани.
Биологический эффект кальцитонина проявляется снижением уровня кальция и фосфора в крови, что является следствием влияния кальцитонина на костную ткань и почки. В кости кальцитонин угнетает процессы резорбции кальция. Это проявляется снижением экскреции гидроксипролина и содержания кальция в крови. Одновременное уменьшение фосфора в сыворотке крови является результатом снижения мобилизации фосфора из кости и непосредственной стимуляции поглощения фосфора костной тканью. Кальцитонин ингибирует активность и количество остеокластов. Уже через 1 ч после введения кальцитонина уменьшается образование остеокластов из клеток-предшественников. Механизм действия кальцитонина опосредуется цАМФ и активацией протеинкиназ, что сопровождается изменением активности щелочной фосфатазы, пирофосфатазной активности и активности ферментов.
Кальцитриол
Наряду с парагормоном и кальцитонином в поддержании фосфорно-кальциевого гомеостаза большое участие принимает витамин в (D-гормон, холекальциферол или витамин D3), который при гидроксилировании в печени и почках образует гормон кальцитриол (1α,25-дигидроксихолекальциферол)
Витамин D3
образуется в коже из 7-дегидрохолестерина под влиянием ультрафиолетового облучения (рис.3). Однако витамин D3
является неактивным. Некоторые авторы рассматривают его как прогормон, который путем гидроксилирования (25-гидроксилаза) сначала в печени превращается в 25-гидроксихолекальциферол (25-ОНD2
), а затем в почках, также путем гидроксилирования (1-гидроксилаза), в 1,25-дигидроксихолекальциферол (1,25-(ОН)2
D3
. Гидроксилирование в почках может осуществляться и другим путем (24-гидроксилаза) с образованием 24-25-(ОН)2
D3
. Гидроксилирование витамина в происходит в митохондриях. Необходимо отметить, что витамин D2 (эргокальциферол), содержащийся в растениях, в организме гидроксилируется тем же путем с образованием 1,25-(ОН)2
D2
, который по биологической активности эквивалентен 1,25-(ОН)2
D3
.

Рисунок 3. Образование кальцитриола из 7-дегидрохолестерина.
Помимо парагормона, кальцитонина и витамина D, на гомеостаз кальция в организме влияют и другие гормоны:
1. Глюкокортикоиды увеличивают глюконеогенез, используя матрикс костной ткани и повышая резорбцию кости, снижают активность остеобластов и скорость образования новой костной ткани, повышают экскрецию кальция и снижают абсорбцию кальция в желудочно-кишечном тракте.
2. Гормон роста повышает экскрецию кальция почками, активность остеобластов и процессы минерализации во вновь образовавшейся костной ткани и увеличивает активность остеокластов и деминерализацию в ранее образовавшейся кости.
3. Тифоидные гормоны (избыток Т3 и Т4), повышают экскрецию кальция с мочой, увеличивают процессы резорбции кости. Недостаток тифоидных гормонов задерживает образование и созревание костной ткани;
4. Глюкагон способствует развитию гипокальциемии посредством прямого влияния на кости (снижая процессы резорбции) и опосредованно – через стимуляцию высвобождения кальцитонина.
5. Половые гормоны снижают экскрецию кальция с мочой и калом, стимулируют активность остеобластов.
Для нормального функционирования клеток помимо обмена веществ и энергии необходима интеграция функций, осуществляемая, в частности, гормонами – веществами, способными контролировать различные стороны клеточного метаболизма. Недостаток или избыток гормона необратимо ведет к нарушению функционирования клеток и возникновению различных заболеваний.
В случае нарушения фосфорно-кальциевого обмена в организме могут возникать такие заболевания как:
- гипопаратироз – cиндром, развитие которого связано с недостаточностью секреции парагормона околощитовидными железами, снижением реабсорбции кальция в канальцах почек, уменьшением абсорбция кальция в кишечнике, что приводит к гипокальциемии;
- гиперпаратироз – заболевание, развитие которого связано с избыточной секрецией парагормона и повышением вследствие этого содержания кальция в сыворотке крови. Гиперкальциемия, которая является одним из ведущих симптомов гиперпаратироза, встречается и при других заболеваниях.
Для лечения подобных заболеваний в медицине применяют синтетический кальцитонин, или же полученный из лосося (кальцитрин). При дефиците парагормона и, как следствие, содержания кальция и фосфора, для лечения применяют параткогмон, лекарственная форма которого называется паратиреоидин.
1. Комов В.П., Шведова В.Н. Биохимия, М.: Дрофа, 2004, 638 стр.
2. Кольман Я., Рем К.-Г. Наглядная биохимия, М.: Мир, 2000, 469 стр.
3. Балаболкин М.И. Эндокринология, М.: Универсум Паблишинг, 1998.
4. Сингер М., Берг П. Гены и геномы. В 2 т., М.: Мир, 1998.
|