1. популяция. популяционный подход в экологии
(хар-ка, численность, плотность)
популяция
(позднелат. populatio, от лат. populus — народ, население)
в генетике, экологии и эволюционном учении, совокупность особей одного вида, более или менее длительно занимающая определённое пространство и воспроизводящая себя в течение большого числа поколений; особи одной п. имеют большую вероятность скрещиваться друг с другом, чем с особями других п.; это связано с тем, что данная совокупность особей отделена от других таких же совокупностей особей той или иной степенью давления различных форм изоляции (см. изоляция). основной характеристикой п., определяющей её центральное положение как элементарной единицы эволюционного процесса, является её генетическое единство: в пределах п. в той или иной степени осуществляется панмиксия. вместе с тем особям, составляющим п., присуща генетическая гетерогенность (в пределах единого генофонда п.), определяющая приспособленность п. к различным условиям среды обитания и создающая столь важный для эволюции резерв наследственной изменчивости (см. изменчивость). вследствие генетической и морфофизиологической неравноценности особей, неоднородности окружающей среды п. имеет сложную структуру: особи различаются по полу и возрасту, принадлежности к разным, обычно перекрещивающимся поколениям (см. поколение), к разным фазам жизненного цикла, к тем или иным малоустойчивым группировкам внутри п. (стадо, колония, семья и т.п.).
биологический вид обычно представлен множеством взаимодействующих п. число особей, входящих в п., широко варьируя не только у разных видов, но и в пределах одного вида, всегда достаточно велико (обычно не менее нескольких сотен, однако у ряда видов может достигать сотен, тысяч, а может быть, и миллионов особей). границы пространства, занятого п., часто бывает трудно определить, т.к. они всегда динамичны, что обусловлено, например, колебаниями численности и плотности п. (см. волны жизни).
на популяционном уровне постоянно ощущается влияние всех др. основных уровней организации жизни — молекулярно-генетического, организменного, биогеоценотического. генетически детерминированные процессы клеточного метаболизма обусловливают в процессе индивидуального развития разную приспособленность отдельных особей к абиотическим и биотическим условиям среды обитания. характер взаимоотношений между особями, в частности то место в установившейся внутри п. иерархии, какое занимает данная особь, определяет важные свойства п. изменение биогеоценотической среды влечёт за собой изменение генетического состава и структуры п., её численности и величины занимаемого ею пространства. в свою очередь, в процессе жизнедеятельности п. воздействует на все уровни среды и, в частности, влияя на численность и структуру п. др. видов, служит фактором, обусловливающим динамику биогеоценоза.
в пределах п. (и при взаимодействии разных п.) протекают процессы микроэволюции (см. микроэволюция) — основа эволюционного процесса в целом. поэтому динамика генетического состава п., её численности и структуры, обмена веществ и энергии между п. и средой интенсивно изучаются популяционной генетикой (см. популяционная генетика), популяционной экологией (см. популяционная экология), биогеоценологией (см. биогеоценология), а «популяционное мышление» необходимо во многих разделах общей биологии. всестороннее изучение п. (прежде всего, динамики численностей и обусловливающих её факторов) служит основой как рационального использования хозяйственно-полезных животных, так и мер борьбы с вредителями сельского хозяйства, переносчиками инфекционных заболеваний и т.д. изучение генофондов природных п. растений (сородичей культурных растений, древесных пород и т.п.) — непременное условие успешного решения многих проблем селекции, интродукции, лесоведения. наряду с природными п. изучаются лабораторные (экспериментально созданные) п., например дрозофил, мышевидных грызунов, некоторых растений.
в современной англо-американской литературе термин «п.» часто подменяется термином «дем» (в советской научной литературе демом обычно обозначают внутрипопуляционные «семейные» группировки). термин «п.» употребляют также для обозначения отдельных обособленных групп домашних животных (породы, стада) и культурных растений (сорта, клоны, линии). в гистологии, медицине, микробиологии п. называют однородные группы клеток в тканях многоклеточных организмов или в культуре клеток.
любая популяция растений, животных и микроорганизмов – это совершенная живая система, способная к саморегуляции, восстановлению своего динамического равновесия. но она существует не изолированно, а совместно с популяциями других видов, образуя биоценозы. поэтому в природе широко распространены и межпопуляционные механизмы, регулирующие взаимоотношения между популяциями разных видов. в качестве регулятора данных взаимоотношений выступает биогеоценоз, состоящий из множества популяций разных видов. в каждой из этих популяций происходят взаимодействия между особями, и каждая популяция оказывает воздействие на другие популяции и на биогеоценоз в целом.
основные характеристики популяций
к основным характеристикам популяции относят численность и плотность.
численность
– общее количество особей на данной территории или в данном объеме.
плотность
– количество особей или их биомасса на единице площади или объема. в природе происходит постоянные колебания численности и плотности.
динамика численности и плотности определяется в основном рождаемостью, смертностью и процессами миграции.
рождаемость
различают абсолютную и удельную. абсолютная рождаемость – это количество новых особей, появившихся за единицу времени, а удельная – количество новых особей, отнесенных к определенному числу особей. согласно правилу максимальной рождаемости в идеальных условиях в популяциях появляется максимально возможное количество новых особей, однако рождаемость ограничивается физиологическими особенностями вида. например, одуванчик за 10 лет способен заполонить весь земной шар, при условии, что все его семена прорастут.
смертность
различают абсолютную и удельную. она характеризует скорость снижения численности популяции от гибели из-за болезней, старости, хищников, недостатка корма, и играет главную роль в динамике численности популяции.
различают три типа смертности
: а) одинаковая на всех стадиях развития; б) повышенная смертность в раннем возрасте (характерна для большинства видов растений и животных, так у деревьев к возрасту зрелости доживает менее 1% всходов, у рыб – 1-2% мальков, у насекомых – менее 0,5% личинок); в) высокая смерть в старости (обычно наблюдается у животных, чьи личиночные стадии проходят в благоприятных мало изменяющихся условиях, например в почве, древесине, живых организмах).
популяция приспосабливается к изменению условий среды путем обновления и замещения особей, т.е. процессами рождения (возобновления) и убывания (отмирания), дополняемыми процессами миграции. в стабильной популяции темпы рождаемости и смертности близки, сбалансированы. в растущей популяции рождаемость превышает смертность. для растущих популяций характерны вспышки массового размножения, особенно у мелких животных (саранча, амброзия). нередко растущими становятся популяции крупных животных в условиях заповедного режима. в сокращающейся популяции смертность превышает рождаемость.
структура и динамика популяций
динамика, состояние и воспроизводство популяций согласуются с их возрастной и половой структурой.
в жизненном цикле особей обычно выделяют три возрастных периода: предрепродуктивный, репродуктивный и пострепродуктивный. для растений характерен еще период первичного покоя, который они проходят в стадии поящихся семян.
классификация животных по возрасту г.а. новикова: новорожденные – до момента прозревания; молодые – подрастающие особи, “подростки”; полувзрослые – близкие к половозрелым особям; взрослые – половозрелые животные; старые – особи, переставшие размножаться.
в геоботанике используют классификацию растений по возрасту н.м. черновой и а.м. быловой: покоящиеся семена; проростки (всходы) – растения первого года жизни, многие из них живут за счет питательных веществ в семядолях; ювенильные – переходят к самостоятельному питанию, но морфологически различаются от отличаются от взрослых растений; имматурные – обладают переходными признаками от ювенильных к взрослым растениям; виргинильные – “взрослые подростки”, могут достигать размеров взрослых особей, но регенеративные органы отсутствуют; молодые генеративные – характерно наличие генеративных органов, завершается формирование облика, типичного для взрослого растения; средневозрастные генеративные – отличаются максимальным годичным приростом и максимальной репродуктивностью; старые генеративные – растения продолжают плодоносить, но у них полностью прекращаются рост побегов и образование корней; субсенильные – плодоносят очень слабо, идет отмирание вегетативных органов, новообразование побегов идет за счет спящих почек; сенильные – старые особи, появляются черты ювенильных растений: крупные одиночные листья, порослевые побеги.
ценопопуляция, в которой представлены все перечисленные стадии, называется нормальной полночленной.
пространственное распределение особей в популяциях бывает случайным, групповым и равномерным.
случайное — неравномерное распределение особей в пределах популяции, взаимосвязи между особями выражены слабо. случайное распределение свойственно популяциям в начальный период расселения; популяциям растений, испытывающим сильное угнетение со стороны эдификаторов сообществ; популяциям животных, у которых социальная связь выражена слабо.
групповое распределение встречается наиболее часто, отражает неоднородность условий обитания или разные онтогенетические (возрастные) закономерности популяции, при групповом распределении обеспечивается наибольшая устойчивость популяции. например, формирование стадо у животных, распределение деревьев в лесу.
равномерное размещение в природе встречается редко. им характеризуются вторичные одновозрастные древостои после смыкания крон, большинство животных-хищников, ведущих активный образ жизни.
колебания численности
при благоприятных условиях в популяциях наблюдается рост численности и может быть столь стремительным, что приводит к популяционному взрыву. совокупность всех факторов способствующих росту численности называется биотическим потенциалом. он достаточно высок для разных видов, но вероятность достижения популяцией предела численности в естественных условиях низка, т.к. этому противостоят лимитирующие (ограничивающие) факторы. совокупность факторов, лимитирующих рост численности популяции, называют сопротивлением среды. состояние равновесия между биотическим потенциалом вида и сопротивлением среды поддерживающее постоянство численности популяции получило название гомеостаза или динамического равновесия. при нарушении его происходят колебания численности популяции.
различают периодические и непериодические колебания численности популяций. первые совершаются в течение сезона или нескольких лет, вторые – это вспышки массового размножения, например некоторых вредителей. колебания численности популяций под влиянием биотических и абиотических факторов среды, свойственные всем популяциям, и назывыются популяционными волнами.
для популяций характерно два принципа: минимальные и максимальные размеры. популяция не может состоять из меньшего числа особей, чем необходимо для стабильного развития и устойчивости популяции к факторам внешней среды. однако наряду с принципом минимального размера существует правило популяционного максимума, оно заключается в том, что популяция не может увеличиваться бесконечно, в силу истощения пищевых ресурсов и условий размножения (теория андреварты – бирча ) и ограничения воздействия комплекса абиотических и биотических факторов среды (теория фредерикса).
кроме экологических факторов, включаются внутренние (генетические и физиологические) механизмы регулирования численности популяций: при сокращении жизненного пространства и запасов корма сокращается плодовитость особей (многие насекомые, мышевидные грызуны), повышение смертности на ранних стадиях жизни (паразиты, многие насекомые), задерживается наступление половой зрелости (полевые мыши) и стадии плодоношения (виды деревьев 2 и 3 ярусов в густом лесу), имеет место каннибализм (грызуны, насекомые, рыбы), и др.; снижается выход личинок из яиц (майский хрущак), уменьшаются размеры взрослых особей. при чрезмерном росте численности популяции у млекопитающих, общественных насекомых, птиц начинается эмиграция на новые места.
репродуктивный потенциал
biotic (reproductive) potencial - биотический (репродуктивный) потенциал.
видоспецифичный показатель скорости увеличения численности особей (как правило, в логарифмической шкале) при отсутствии лимитирующих факторов, характеризующий способность данного вида к размножению.
актуальность темы. в последние десятилетия демографические проблемы привлекают внимание как специалистов, так и широкой общественности, приобретая при этом все большее значение.
особенно актуальны исследования уровня рождаемости, который определяет темпы воспроизводства населения и демографическую ситуацию в стране.
в первой половине 90-х годов xx в. наша страна вступила в демографическую стадию, которую принято называть депопуляцией, что означает превышение смертности над рождаемостью в сопровождении таких негативных явлений, как высокий уровень развода и сокращение продолжительности жизни, особенно среди мужской и сельской части населения. с 1992 г. население россии, (а также и северной осетии - алании) не растет, а сокращается достаточно быстрыми темпами: за прошедшие годы оно сократилось в российской федерации почти на 2 млн. человек, или на 1,3 %. надо отметить, что сокращение населения компенсировалось за счет миграционного притока из ближнего зарубежья. естественная убыль населения, т.е. превышение числа умерших над числом родившихся за указанный период составляет 4,2 млн. человек.
социальный опыт последних лет свидетельствует, что для более полной ясности в понимании этой проблемы необходимо включить анализ ряда социальных факторов, влияющих на показатели рождаемости. так, экономические и социально-психологические факторы оказывают непосредственное влияние на процесс деторождения.
кроме того, существенное значение имеет анализ культурно-бытовых традиций, этнических особенностей в изучении социально-психологических и социально-исторических аспектов рождаемости.
к важнейшим свойствам популяции относятся:
-скорость прироста популяции
-средняя продолжительность жизни особей
-количество и качество потомства
максимальная мгновенная скорость прироста популяции обратно пропорциональна продолжительности жизни организмов и отражает их способность увеличивать свою численность.
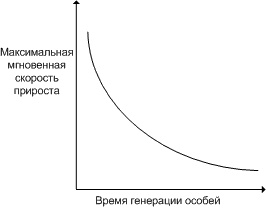
с увеличением размера особей время генерации увеличивается, период размножения отодвигается, поэтому максимальная мгновенная скорость прироста популяции уменьшается. крупные организмы, несмотря на меньшие значения максимальной мгновенной скорости прироста популяции имеют больше преимуществ перед мелкими организмами: 1. меньшее количество потенциальных хищников. 2. крупные организмы легче переносят изменения состояния окружающей среды. таким образом, крупные размеры тела позволяют особям этих популяций существовать и развиваться, несмотря на их меньшую численность.
кривые выживания - графическая зависимость числа выживших особей от их возраста при условии, что первоначальное число особей составляет 100 или 1000.
различают пять основных типов кривых выживания.
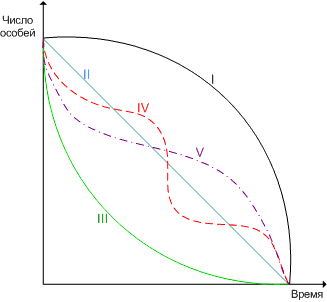
кривая i - характерна для популяций, в которых большинство особей имеет продолжительность жизни, близкую к максимально возможной для данного вида, и умирают в течение короткого отрезка времени. эту форму кривой называют выпуклой. она свойственна насекомым, многим крупным млекопитающим, человеку.
кривая ii - теоретическая, отражает равную вероятность гибели особей в любом возрасте, то есть коэффициент смертности остается постоянным в течение всей жизни особей. например, у многих природных популяций птиц и насекомых, постоянно находящихся в оптимальных условиях.
кривая iii соответствует очень высокой смертности в раннем возрасте, а для особей, переживших этот период, вероятность смерти низка. эта форма кривой - вогнутая. к таким популяциям относятся многие растения, беспозвоночные и рыбы.
кривая iv имеет ступенчатый характер и показывает, что при переходе от одной стадии развития к другой происходит резкое изменение выживания особей.
кривая v - имеет s-образную форму, приближающуюся к теоретической кривой.
свойства популяций
от чего же зависят значения коэффициентов рождаемости и смертности? от
очень многих факторов, действующих на популяцию извне, а также от
собственных ее свойств. объективный показатель способности организмов
увеличивать свою численность —максимальная мгновенная скорость прироста
популяции. этот параметр обратно пропорционален продолжительности жизни
организмов. в этом легко убедиться, обратившись к гиперболической
зависимости между врожденной скоростью увеличения численности популяции и
средним временем генерации, выраженным в днях (рис. 1). мелкие организмы
имеют более высокие значения rтах, чем крупные, что объясняется меньшим
временем генерации. причина подобной корреляции понятна, поскольку
организму для достижения крупных размеров требуется много времени. отсрочка
периода размножения также неизбежно ведет к уменьшению rтах.
тем не менее преимущества, даваемые большими размерами тела, должны
превышать недостатки, связанные с уменьшением rтах, так как в противном
случае крупные организмы никогда не появились бы в эволюции. тенденция к
возрастанию размеров тела с течением геологического времени, прослеженная
по ископаемым остаткам, послужила основанием для введения понятия
увеличение филетического размера.
крупные размеры тела дают совершенно очевидные преимущества: более
крупный организм должен привлекать меньше потенциальных хищников и,
следовательно, он имеет больше шансов не стать жертвой и должен отличаться
лучшей выживаемостью; мелкие организмы находятся в тесной зависимости от
физической среды, даже очень слабые изменения которой могут оказаться для
них губительными. более крупные организмы легче переносят воздействие
подобных изменений и соответственно лучше от них защищаются. однако более
крупным организмам требуется больше вещества и энергии в расчете на одну
особь в единицу времени, чем мелким. кроме того, для них существует гораздо
меньше укрытий и безопасных мест.
в жизни всех организмов в популяции можно выделить три основных
периода: предрепродуционный, репродукционный и пострепродукционный.
относительная продолжительность каждого из них у разных видов весьма
различна. у многих животных первый период самый длинный. ярким примером
могут служить поденки, у которых предрепродуционный период достигает 3 лет,
а репродукционный занимает всего от 2-3 ч до 1 суток. у американской цикады
предрепродуционный период составляет 17 лет. но есть виды, особи которых,
едва появившись на свет, начинают интенсивно размножаться (большинство
бактерий).
[pic]
репродукционные возможности популяции зависят от ее возрастного
состава. продолжительность жизни особей популяции можно оценить, используя
кривые выживания. существует три типа кривых выживания (рис. 2).
[pic]
первый тип (кривые 1) соответствует ситуации, когда большее число особей
имеет одинаковую продолжительность жизни и умирает в течение очень
короткого отрезка времени. кривые характеризуются сильно выпуклой формой.
такие кривые выживания свойственны человеку (рис. 2, 1), причем кривая
выживания для мужчин по сравнению с аналогичной кривой для женщин менее
выпуклая, поэтому страховой полис для мужчин в большинстве стран запада в
1,5 раза дороже, чем для женщин. для большинства копытных кривые выживания
также выпуклые (рис. 3), хотя и в различной степени для разных видов, а
также в зависимости от пола. второй тип (рис. 2, 2) свойствен видам,
коэффициент смертности которых остается постоянным на протяжении всей их
жизни. поэтому кривая выживания трансформируется в прямую линию. такая
форма кривой выживания свойственна пресноводной гидре. третий тип (рис. 2,
3) — сильно вогнутые кривые, отражающие высокую смертность особей в раннем
возрасте. так характеризуется продолжительность жизни некоторых птиц, рыб,
а также многих беспозвоночных.
знание типа кривой выживания дает возможность построить пирамиду
возрастов (рис. 4). следует различать три типа таких пирамид. пирамида с
широким основанием, что соответствует высокому проценту молодняка,
характерна для популяции с большим значением коэффициента рождаемости.
средний тип пирамиды соответствует равномерному распределению особей по
возрастам в популяции со сбалансированными коэффициентами рождаемости и
смертности — пирамида выровненная. пирамида с узким основанием
(обращенная), отвечающая популяции с численным преобладанием старых особей
над молодняком, характерна для сокращающихся популяций. в таких популяциях
коэффициент смертности превышает коэффициент рождаемости.
[pic]
[pic]
большое значение для увеличения численности популяции имеют затраты на
потомство, выражающиеся в определенной тактике размножения. не все потомки
равноценны: те из них, которые произведены на свет в конце вегетационного
сезона, обычно имеют меньшую вероятность дожить до взрослого состояния по
сравнению с потомками, появившимися на свет раньше. сколько усилий должны
родители затрачивать на каждого потомка? при постоянной величине
репродуктивного усилия средняя приспособленность отдельного потомка связана
обратным соотношением с их числом. один крайний вариант тактики размножения
— вложить все в единственного очень крупного и хорошо приспособленного
потомка, другой — максимально увеличить общее число произведенных на свет
потомков, вложив в каждую особь как можно меньше. однако наилучшая тактика
размножения —это компромисс между производством максимально большого числа
потомков и образованием потомства максимально высокой приспособленности.
указанное соотношение количества и качества потомства иллюстрирует
простая графическая модель (рис. 5).
[pic]
в маловероятном случае, т. е. в случае линейной зависимости
приспособленности потомков от затрат на них родителей, приспособленность
каждого отдельного потомка уменьшается с увеличением размера помета или
кладки. так как приспособленность родителей или, что то же самое, общая
приспособленность всех потомков - постоянная величина, с точки зрения
родителей не существует оптимального размера кладки. однако поскольку
первоначальные расходы на потомство вносят больший вклад в
приспособленность потомков, чем последующие (имеет место 5-образный
характер зависимости приспособленности потомков при увеличении вклада
родителей; см. рис. 7.6), то очевидно, что существует некоторый оптимальный
размер кладки. в данном гипотетическом случае родители, расходующие только
20% своего репродуктивного усилия на каждого из пяти своих потомков,
получат большую отдачу от своего вклада, чем при любом другом размере
кладки. подобная тактика, будучи оптимальной для родителей, не является
наилучшей для каждого отдельно взятого потомка, максимальная
приспособленность которого достигается в том случае, если он единственный
отпрыск, получивший полный вклад усилий от своих родителей. следовательно,
в данном случае налицо «конфликт родителей и детей».
особенно большое влияние на форму s-образной кривой оказывает
конкурентная обстановка. в сильно разреженной среде (конкурентный вакуум)
наилучшей репродуктивной стратегией следует считать максимальный вклад
вещества и энергии в размножение для продуцирования как можно большего
количества потомков в самые короткие сроки. поскольку конкуренция невелика,
потомки могут выжить, даже если они имеют очень малые размеры и низкую
приспособленность. однако в насыщенной среде обитания, где заметно
проявляются эффекты массы, а конкуренция остра, оптимальной стратегией
будет расходование большого количества энергии на преодоление конкуренции,
повышение собственной выживаемости и на продуцирование более
конкурентоспособных потомков. при подобной стратегии лучше иметь крупных
потомков, а поскольку энергетически они дороже, их может быть произведено
на свет меньше.
итак, свойства популяции можно оценить по таким показателям, как
рождаемость, смертность, возрастная структура, соотношение полов, частота
генов, генетическое разнообразие, скорость и форма кривой роста и т. д.
плотность популяции определяется ее внутренними свойствами, а также
зависит от факторов, действующих на популяцию извне.
факторы динамики численности популяций
известно три типа зависимости численности популяции от ее плотности
(рис. 6). при первом типе (кривая 1) скорость роста популяции уменьшается
по мере увеличения плотности. это широко распространенное явление позволяет
понять, почему популяции некоторых животных относительно устойчивы. во-
первых, при увеличении плотности популяции наблюдается снижение
рождаемости. так, в популяции большой синицы при плотности меньше одной
пары на 1 га на одно гнездо приходится 14 птенцов; когда же плотность
достигает 18 пар на 1 га, выводок составляет менее 8 птенцов. во-вторых,
при увеличении плотности популяции меняется возраст наступления половой
зрелости. например, африканский слон в зависимости от плотности популяции
может достигать половой зрелости в возрасте от 12 до 18 лет. кроме того,
этот вид при низкой плотности дает приплод 1 слоненок за 4 года, тогда как
при высокой — рождаемость составляет 1 слоненок за 7 лет.
при втором типе зависимости (кривая 2) темп роста популяции максимален
при средних, а не при низких значениях плотности. так, у некоторых видов
птиц (например, чаек) число птенцов в выводке увеличивается с повышением
плотности популяции, а затем, достигнув наибольшей величины, начинает
уменьшаться. этот тип влияния плотности популяции на скорость размножения
особей характерен для видов, у которых отмечен групповой эффект. при
третьем типе (кривая 3) темп роста популяции не изменяется до тех пор, пока
она не достигнет высокой плотности, затем резко падает.
[pic]
подобная картина наблюдается, например, у леммингов. при пике
численности плотность леммингов становится избыточной, и они начинают
мигрировать. элтон так описал миграции леммингов в норвегии: животные
проходили через деревни в таком количестве, что собаки и кошки, которые
вначале нападали на них, просто перестали их замечать. достигнув моря,
обессиленные лемминги тонули.
регуляция численности равновесных популяций определяется
преимущественно биотическими факторами. среди них главным фактором часто
оказывается внутривидовая конкуренция. примером может служить борьба птиц
за мести для гнездования.
внутривидовая конкуренция может быть причиной физиологического
эффекта, известного под названием шоковой болезни. его отмечают, в
частности, у грызунов. когда плотность популяции становится слишком
большой, шоковая болезнь приводит к снижению плодовитости и увеличению
смертности, что возвращает плотность популяции к нормальному уровню.
у некоторых видов животных взрослые особи питаются собственным
приплодом. это явление, известное как каннибализм, снижает численность
популяции. каннибализм свойствен, например, окуням: в озерах западной
сибири 80% пищи крупных особей составляет молодь того же вида. молодь, в
свою очередь, питается планктоном. таким образом, когда нет других видов
рыб, взрослые особи живут за счет планктона.
межвидовые взаимодействия также играют существенную роль в контроле
плотности популяции. взаимодействия паразит-хозяин и хищник-жертва часто за
висят от плотности. болезни также являются фактором, принимающим участие в
регуляции плотности популяции. когда кролики болеют миксоматозом,
вызываемым вирусом, распространение инфекции идет гораздо быстрее в
популяциях с повышенной плотностью.
хищничество как ограничивающий фактор само по себе имеет большое
значение. причем если влияние жертвы на численность популяции хищника не
вызывает сомнений, то обратное воздействие, т. е. на популяцию жертвы,
бывает не всегда. во-первых, хищник уничтожает больных животных, тем самым
он улучшает средний качественный состав популяции жертвы. во-вторых, роль
хищника ощутима только тогда, когда оба вида обладают приблизительно
одинаковым биотическим потенциалом. в противном случае из-за низкого темпа
размножения хищник не в состоянии ограничить численность своей жертвы.
например, только одни насекомоядные птицы не могут остановить массовое
размножение насекомых. иными словами, если биотический потенциал хищника
намного ниже биотического потенциала жертвы, действие хищника приобретает
постоянный характер, не зависящий от плотности его популяции.
численность насекомых-фитофагов нередко определяется комбинацией
видоспецифичных реакций насекомых и растений на воздействие загрязняющих
веществ. загрязнение понижает резистентность растений, в результате чего
численность насекомых возрастает. однако при слишком большом загрязнении
численность насекомых падает, несмотря на уменьшение сопротивляемости
растений.
приведенная дифференциация факторов динамики численности популяций
позволяет понять их реальное значение в жизни и воспроизводстве популяций.
современная концепция автоматического регулирования численности популяций
базируется на сочетании двух принципиально различных явлений: модификаций,
или случайных колебаний численности, и регуляций, действующих по принципу
кибернетической обратной связи и нивелирующих колебания. в соответствии с
этим выделяют модифицирующие (независящие от плотности популяции) и
регулирующие (зависящие от плотности популяции) экологические факторы,
причем первые из них воздействуют на организмы либо непосредственно, либо
через изменения других компонентов биоценоза. по существу, модифицирующие
факторы представляют собой различные абиотические факторы. регулирующие
факторы связаны с существованием и активностью живых организмов
(биотические факторы), поскольку лишь живые существа способны реагировать
на плотность своей популяции и популяций других видов по принципу
отрицательной обратной связи (рис. 7).
[pic]
если воздействия модифицирующих факторов приводят лишь к
преобразованиям (модификациям) колебаний численности, не устраняя их, то
регулирующие факторы, выравнивая случайные отклонения, стабилизируют
(регулируют) численность на определенном уровне. однако на разных уровнях
численности популяции регулирующие факторы принципиально различны (рис. 8).
например, хищники-полифаги, способные при изменении численности жертв
ослабить или усилить свою активность—функциональная реакция,— оказывают
действие при сравнительно низких значениях численности популяции жертвы.
хищники - олигофаги, характеризующиеся в отличие от полифагов
численной реакцией на состояние популяции жертвы, оказывают на нее
регулирующее действие в более широком диапазоне, чем полифаги. при
достижении популяцией жертвы еще более высокой численности создаются
условия для распространения болезней и, наконец, предельный фактор
регуляции — внутривидовая конкуренция, ведущая к исчерпыванию доступных
ресурсов и развитию стрессовых реакций в популяции жертвы. на рис. 8
представлена многозвенная буферная система регулирования численности
популяции под воздействием биотических факторов, степень влияния которых
зависит от плотности популяции. в реальной ситуации данный параметр зависит
от большого числа факторов, в частности и тех, которые не оказывают
регулирующего воздействия на плотность популяции по принципу обратной
связи. взаимодействие между модифицирующими, регулирующими, а также такими
специфическими факторами, как размеры тела, группы и индивидуального
участка, при их влиянии на плотность популяции млекопитающих показано на
рис. 9.
таким образом, чтобы получить исчерпывающую информацию о том, какие
факторы вызывают колебания численности, теоретически требуются данные о
физико-химических условиях, обеспеченности ресурсами, жизненном цикле этих
организмов и влиянии конкурентов, хищников, паразитов и т. д., причем нужно
знать, как все эти факторы влияют на рождаемость, смертность и миграцию.
все популяции непрерывно изменяются: новые организмы рождаются или
прибывают как иммигранты, а прежние гибнут или эмигрируют. несмотря на это,
флуктуации размера популяции не безграничны. с одной стороны, она не может
расти беспредельно, а с другой — вымирают виды достаточно редко.
следовательно, один из основных признаков популяционной динамики —сочетание
изменений с относительной стабильностью. при этом колебания размеров
популяций сильно различаются у разных видов.
[pic]
[pic]
особи в популяции взаимодействуют между собой, обеспечивая свою
жизнедеятельность и устойчивое воспроизводство популяции.
у животных, ведущих одиночный образ жизни или создающих семьи,
регулирующим фактором является территориальность, которая влияет на
обладание определенными пищевыми ресурсами и имеет большое значение для
размножения. особь защищает пространство от вторжения и открывает ее для
другой особи только при размножении. самое рациональное использование
пространства достигается в том случае, если образуется настоящая территория
— участок, из которого другие особи изгоняются. так как владелец участка
психологически господствует на нем, для изгнания чаще всего достаточно лишь
демонстрации угроз, преследования, самое большее — притворных атак, которые
прекращаются еще на границах участка. у данных животных огромное значение
имеют индивидуальные различия между особями — наиболее приспособленные
обладают большим индивидуальным пищевым ареалом.
у животных, ведущих групповой образ жизни и образующих стаи, стада,
колонии, групповая защита от врагов и совместная за- бота о потомстве
повышает выживаемость особей, что влияет на численность популяции и ее
выживаемость. данные животные организованы иерархически. иерархические
отношения соподчинения построены на том, что ранг каждого известен каждому.
как правило, высший ранг принадлежит самому старшему самцу. иерархия
контролирует все взаимодействия внутри популяции: брачные, особей разных
возрастов, родителей и потомства.
у животных особую роль играют отношения «мать—дитя». родители передают
потомкам генетическую информацию и информацию о внешней среде.
Мутуали́зм
— взаимовыгодные отношения между организмами. Более общим понятием является симбиоз, который представляет собой сосуществование различных биологических видов. Но в отличие от мутуализма, симбиоз может быть и не выгоден одному из партнеров, например, в случае паразитизма.Преимущества, которые получает организм, вступающий в мутуалистические отношения, могут быть различны. Часто по крайней мере один из партнеров использует другого в качестве пищи, тогда как второй получает защиту от врагов или благоприятные для роста и размножения условия. В других случаях вид, выигрывающий в пище, освобождает партнера от паразитов, опыляет растения или распространяет семена. Каждый из участников мутуалистической пары действует эгоистично, и выгодные отношения возникают лишь потому, что получаемая польза перевешивает затраты, требуемые на поддержание взаимоотношений.Взаимовыгодные связи могут формироваться на основе поведенческих реакций, например, как у птиц, совмещающих собственное питание с распространением семян. Иногда виды-мутуалисты вступают в тесное физическое взаимодействие, как при образовании микоризы (грибокорня) между грибами и растениями.Тесный контакт видов при мутуализме вызывает их совместную эволюцию. Характерным примером служат взаимные приспособления, которые сформировались у цветковых растений и их опылителей. Часто виды-мутуалисты совместно расселяются.Самая тесная форма мутуализма — когда один организм живет внутри другого. Поразительным примером этого служит система органов пищеварения коров и других жвачных животных. Коровы, как и человек, не способны переварить целлюлозу — вещество, которое в большом количестве содержится в растениях. Но у жвачных животных есть особый орган — рубец. Он представляет собой полость, в которой живет множество микробов. Растительная пища, после того как животное ее прожевало, попадает в рубец, и там эти микробы разрушают целлюлозу. (Животное может отрыгнуть и вновь прожевать частично расщепленную пищу — именно этим и занимаются коровы, когда пережевывают свою жвачку.) Рубец коровы — это замкнутая микроэкосистема, образованная множеством различных микроорганизмов, задача которых состоит в переваривании целлюлозы для своего хозяина. Аналогично корневая система высших растений образована переплетением корневой ткани и грибных нитей, так что грибы снабжают растение минеральными веществами.
Комменсализм
Комменсализм, сотрапезничество, нахлебничество, сожительство животных разных видов, характеризующееся тем, что один из них (комменсал) постоянно или временно живёт за счёт другого, не причиняя ему вреда (ср. Аменсализм). При К. один из организмов может использовать другой для защиты, как средство передвижения или питаться за его счёт. В зависимости от характера взаимоотношений животных, которым свойствен К., их делят на три группы. 1) Комменсал ограничивается использованием пищи организма др. вида; например, в извивах раковины, занятой раком-отшельником, обитает кольчатый червь из рода Nereis, поедающий остатки пищи рака. 2) Комменсал прикрепляется (временно или постоянно) к организму другого вида, который в этом случае называется хозяином; например, рыба-прилипало своим спинным плавником, превращенным в присоску, прикрепляется к коже акул и др. крупных рыб, пользуясь ими для передвижения; морские гидроиды, поселяющиеся на коже рыб и питающиеся их экскрементами; некоторые ракообразные (например, морские жёлуди), живущие на коже китов и на раковинах моллюсков. 3) Комменсал поселяется во внутренних органах хозяина; например, инфузории опалины, живущие в задней кишке лягушек; некоторые жгутиконосцы, обитающие в кишечнике млекопитающих.
Аменсализм
Аменсализм (от греч. а — отрицательная частица и лат. mensa — стол, трапеза), форма взаимоотношений между организмами, полезная для одного вида, но вредная для другого. Например, обитающие в норах сусликов и кротов "квартиранты" (клещи, блохи и др.) могут служить источником распространения среди грызунов инфекционных заболеваний; "нахлебники" хищника могут навлечь на него врага или затруднить охоту (например, вороны, подбирающие остатки трапезы волка, своим криком привлекают внимание охотника). Четко отграничить А. от др. форм взаимоотношений между организмами не всегда возможно (см. Межвидовые взаимоотношения). Чтобы определить пользу или вред от сосуществования организмов, необходимо учитывать сложный комплекс условий среды.
|