МИНИСТЕРСТВО СЕЛЬСКОГО ХОЗЯЙСТВА РОССИЙСКОЙ ФЕДЕРАЦИИ
НОВОСИБИРСКИЙ ГОСУДАРСТВЕННЫЙ АГРАРНЫЙ УНИВЕРСИТЕТ
Биолого – технологический институт
Кафедра свиноводства
Курсовой проект
На тему: Генетические маркеры и их использование в селекции.
Выполнила:
Студентка 241 гр.
Габдульбарова О.Н.
Проверил:
Барсукова М.А.
Новосибирск 2010
Содержание:
Введение …………………………………………………………………..3
Обзор литературы
1. ДНК-технологии в свиноводстве……………………………………….4
2.
Использование групп крови в качестве генетических маркеров продуктивности свиней……………………………………………………6
3.Расчет ритмичного производственного потока в свиноводческом потомстве.
Заключение………………………………………………………………..20
Список литературы……………………………………………………….21
Введение
Одним из подходов к прогнозированию продуктивных качеств в раннем возрасте является определение маркирующего действия полиморфных белков и групп крови на различные хозяйственно-полезные признаки. Интерес к использованию иммуногенетических тестов в селекции объясняется доступностью для изучения, относительно простым определением, четким характером их наследования и неизменяемостью в течение постэмбриональной жизни.
Уже первые исследования по установлению связи групп свиней с откормочными и мясными качествами, выполненными в 1964 г., выявили наличие статистически достоверных корреляций, указывающих на существование зависимости между эритроцитарными антигенами и продуктивностью животных.
Эти результаты послужили толчком к широко развернувшимся исследованиями по изучению связей групп крови с репродуктивными, откормочными и мясными качествами у разных пород свиней. Существуют публикации о связи групп крови и полиморфных форм белков с продуктивными признаками животных, устойчивостью к стрессам, резистентностью к заболеваниям, о корреляции этих признаков к гетерозиготным состоянием некоторых систем групп крови с отдельными антигенами факторами. Анализ накопленного материала показал, что взаимосвязь между изучаемыми признаками обнаруживается чаще всего у животных определенных пород, племенных стад, генеалогических групп, т.е является сугубо специфической, не обусловленной плейотропным эффектом.
Одним из объяснений данного факта может быть то, что длительное и интенсивное использование хряков и маток , с высокой наследственной предрасположенностью к какому-то определенному виду продуктивности и имеющих тот или иной антиген или генотип по группам крови, которые были у родителей. То есть, данная связь возникла и закрепилась в конкретной популяции при определенных методах отбора и подбора. Ее нельзя считать характерной для всей породы и всех племенных хозяйств, но в тех стадах, где она выявлена, селекция может проводиться под учетом приоритета этого антигена или генотипа – маркера конкретного признака продуктивности.
Обзор литературы
1.ДНК-технологии в свиноводстве
Эффективность селекционно-племенной работы в животноводстве во многом зависит от точности оценки племенной ценности. Интеграция молекулярной генетики в прикладные науки, начиная с 80-ых годов XX века, сделала возможным оценку генетического потенциала продуктивности животных не только на основании данных фенотипа, но и непосредственно на уровне ДНК [Эрнст Л.К., Зиновьева Н.А., 2008]. И сегодня наряду с математическими методами оценки племенной ценности (селекционные BLUP-индексы), все более широкое применение во всем мире находят ДНК-технологии [Калашникова Л.А. и др., 2000, Захаров И.А., 2006].
Одной из приоритетных задач работы Центра биотехнологии и молекулярной диагностики животных ГНУ ВНИИЖ является исследование молекулярно-генетических основ реализации генотипа племенных животных и разработка ДНК-технологий, направленных на расширение и повышение эффективности использования их генетического потенциала.
В результате проведенных в 2006-2009 гг. исследований предложен проект комплексной системы оценки свиней по ДНК-маркерам. Система включает следующие компоненты:
(I) Маркеры контроля происхождения и породной принадлежности;
(II) Маркеры продуктивности, в т.ч. 5 маркеров многоплодия, 3 маркера качества мяса, 3 маркера мясной и откормочной продуктивности, 2 маркера устойчивости к заболеваниям.
(III) Гены наследственных заболеваний.
I. Маркеры контроля происхождения и породной принадлежности.
Наиболее распространенным типом маркеров для генетического контроля происхождения свиней в России является анализ систем групп крови (ГК), представляющих собой определенные сочетания эритроцитарных антигенов [Безенко С. П., 1982, Попов Н.А., 2002, Марзанов Н.С., 1994]. Существующие на сегодня системы тестирования животных, по группам крови характеризуются большой трудоемкостью, низкой производительностью, высокими материальными и временными затратами, а так же зависимостью от субъективных факторов (так как идентификация результатов анализа осуществляется визуально), что существенно сказывается на их точности.
Развитие ДНК-технологий обусловило открытие микросателлитов (МС) [Tautz D., 1989]. Согласно директиве Европейского общества генетиков с 2004 года исследование МС является основным методом подтверждения индивидуального происхождения племенных животных в Европе.
В рамках сравнительной оценки ГК и МС в качестве критериев в генетической экспертизе происхождения показано, что вероятность совпадения генотипов свиней при тестировании по 20-и ЭА 10-и систем ГК составляет 0,0005. Это означает, что у одного животного из каждых 2000 протестированных особей возможно появление идентичного генотипа по всем протестированным ЭА, в то время как при тестировании по 12-и локусам МС вероятность совпадения генотипов составляет 10-13, то есть практически исключена. Вероятность совпадения генотипов сибсов по ГК составляет 0,02358, по МС – 0,00001. Учитывая, что число сибсов одного возраста, как правило, не превышает 15 голов, обе системы являются информативными для генетической идентификации сибсов.
С другой стороны, расчет вероятности исключения отцовства на основании анализа по ГК и МС, показал, что если известны оба родителя, точность определения несоответствия происхождения с использованием ГК составляет 76,5-87,3% (в зависимости от породы), в то время с использованием МС – 99,78-99,99%. В случае если известен один родитель, точность составляет 41,1-64,3 и 96,5-99,5%, соответственно. Точность исключения обоих родителей по ГК и МС составляет, соответственно, 91,7-96,6 и 99,9999%.
Другим аспектом прикладного использования МС является контроль породной принадлежности. Комбинирование результатов анализа МС с современными программными пакетами обработки данных молекулярно-генетического анализа (GenAlEx, версия 6.0, 2007 г., Structure, версия 3.2.1, 2009 г.) делает возможным определение породной принадлежности и подтверждение чистопородности животных. Последнее является наиболее актуальным для племенных свиней, завозимых из-за рубежа.
II. Маркеры продуктивности.
Проведенный анализ мировых информационных ресурсов позволил выявить ряд потенциальных ДНК-маркеров продуктивных признаков свиней, для определения полиморфизма которых нами были разработаны аналитические тест-системы. Предлагаемые для внедрения в сельхозпредприятиях России маркеры свиней охватывают широкий спектр экономически значимых показателей: многоплодие, сохранность поросят, качество мяса, откормочная и мясная продуктивность. Результаты влияния генотипов ДНК-маркеров по результатам собственных исследований обобщены в таблице.
В качестве задач на ближайшую перспективу нами ставится дальнейшее расширение спектра маркеров и разработка систем диагностики, позволяющих выполнять одновременный анализ полиморфизма нескольких генов, и направленных на снижение себестоимости и повышение производительности ДНК-технологий.
III. Маркеры (гены) наследственных заболеваний.
Широкое распространение искусственного осеменения приводит к многократному тиражированию генотипов лучших хряков-производителей, вместе с которым происходит распространение генетических аномалий. Анализ мировых баз данных показывает, что в настоящее время у свиней описано более 200 генетических дефектов, многие из которых с достаточно высокой частотой распространены в популяциях свиней. Так, 50% хряков-производителей породы йоркшир, используемых в системе искусственного осеменения Финляндии, в 1999 году являлись носителями скрытого наследственного дефекта, характеризующегося укорочением хвоста сперматозоидов. Определенную проблему в свиноводстве представляет наследственно обусловленный стресс-синдром у свиней мясных и беконных пород (одним из его проявлений является PSE-порок мяса), а так же синдром «кислое мясо» у гемпширов.
О перспективности исследований в области ДНК-диагностики в Российской Федерации свидетельствует факт хозяйственного внедрения разработок в области ДНК-диагностики, объем которых за последние 5 лет вырос более чем в 15 раз, а так же возросшее число предприятий, востребующих данные разработки.
По предварительным оценкам внедрение ДНК-диагностики позволит повысить точность оценки племенной ценности животных и ускорить принятие селекционных решений, что уже в ближайшие 3-5 лет сделает возможным повышение на 5-10% эффективности селекционно-племенной работы в свиноводстве.
2.Использование групп крови в качестве генетических маркеров продуктивности свиней.
Чаще других в качестве маркеров различных продуктивных качеств у свиней используют группы крови систем A, E, F,G, Kзначительно реже H, L, M, J, остальные группы крови - лишь эпизодически. Объяснение состоит, по–видимому, в том, что первые пять систем крови достоверно связаны с такими важнейшими хозяйственными признаками, как продуктивность маток, жизнеспособность приплода, скороспелость, использование корма и мясные качества свиней.
Целым рядом исследований, выполненных на разных породах свиней, подтверждается достоверный маркирующий эффект высокого многоплодия маток эритроцитными антигенами Acp, Ea, Ga, Fa, генотипами Ga/b, Eedg/edf, Eaeg/edf, Eaeglh/bdgи трансферинами типа АА. У свиноматок с указанными маркерами среднее количество поросят в помете при рождении выше на 0, 26-1,4 гол., чем у маток не имеющих антигенов и генотипов.
Отмечена достоверная связь антигенов Gb, Eb, Ee, Eg, Ko, генотипов Ga/b, Ebdf/ edf, Eaegjh/ begiи трансферина АВ с более высокой сохранностью поросят в подсосный период, а генотипов Ebdf/edfи Lbegi\ begi с высокой массой при отъеме. Признаком низкого многоплодия и молочности маток являются генотип Е edf/ edf.
В тех породах и племенных стадах, где выявлены отмеченные закономерности, целесообразно использовать их для осуществления индивидуального и группового подбора с целью повышения репродуктивных свойств маток путем создания сочетающихся отцовских и материнских форм по группам крови и полиморфным белкам.
Довольно хорошо изучена взаимосвязь полиморфных систем групп крови с откормочными и мясными качествами свиней. Большинство исследований сходятся во мнении, что представляют животные, имеющие генотипы A- /-, Ga/b, Ebdf /bdf, Ebdf/edf, Ebdg/edf, Eaeg/adf, Eedghkmnp/edghkmnp, Ebdgkmp/ bdgkmpи некоторые другие. У них, по сравнению с животными, не имеющих генотипов, преимущество по скороспелости достигает 12-15 дней, по среднесуточным приростам более 150 г. Маркерами низкой скороспелости являются генотипы Eedg/edf и Eedf/edf.
К специфическим маркерам высокой оплаты корма чаще других относят аллели A-,Gb, Ka, Ebdg.Исходным материалом для исследований по данному вопросу послужило поголовье свиней племенных заводов «Кубань» и «Краснодарское» , в которых около 40 лет применяется внутрилинейный подбор свиней в заводских линиях без привлечения животных ихз других линий, а также животные трех племенных ферм, в которых разведение свиней осуществлялось путем завоза и спаривания хряков и маток из разных хозяйств. Наследственная основа таких животных значительно шире, чем в племзаводах, и менее консолидирована, что предлагает наличие у более низкого генетического сходства по группам крови.
Для решения поставленной задачи провели иммуногенетический анализ пяти племенных стад свиней по 4 системам групп крови (A, E, G, K) на поголовье 733 гол.
Установлено, что в генетических разнородных стадах племенных ферм маркерами высокой скорости роста являются генотипы Acp/- в системе A, Ga/bв системе G и K -/- в системе К. Животные с генотипом А ср/- по величине среднесуточного прироста достоверно превосходили на 33,4 г среднесуточный показатель по всей выборке животных и на 47,5 г сверстников с генотипом А-/-.
Лучшим использованием корма отличались животные с генотипом Ga/bв системе G(3,97 корм.ед.), с генотипами К ас/- К -/-(3,88ти 3,91 корм.ед на 1кг прироста). Больше других расходовали кормов на прирост свиньи с генотипом Ga/a(4,25 корм.ед.)
Более существенные различия по откормочным качествам установлены среди животных по группам крови в системе Е: по среднесуточному приросту 128,5 г, по расходу кормов на 1 кг прироста 0,3 корм.ед.
Гомозиготные животные с генотипами Eaeg/aeg, Ebdg/bdg и Eedg/edg по скорости роста и затратам корма практически мало чем отличались и были близки к средним показателям по всей группе животных. Гомозиготные животные с генотипом Eedf/edf уступали средним показателям по среднесуточному приросту на 20,4 г, по оплате корма на 0,15 корм.ед. Однако самыми лучшими по скороспелости и использованию кормов оказались именно животные, имеющие аллель edf: гетерозиготные свиньи с генотипом Eaeg/edfпри откорме от 30 до 100 кг давали в сутки по 808,3 г прироста при затратах корма на 1 кг прироста 3,85 корм.ед. Высокой скороспелостью характеризуются гетерозиготные животные с генотипами Eaef/edf и bdg/edf , превышающие средние значения признака по группе на 26,0-54,6 г.
Самый низкий среднесуточный прирост при высоких затратах корма получен у свиней с генотипом Е aeg/edg: прирост ниже средних данных по группе на 40,65 г, оплата корма - на 0,11 корм.ед. Невыскую скорость роста показали животные с генотипом Ebdg/edg. Из полученных результатов следуют, что в данной популяции аллель aegв гетерозиготном состоянии является маркером низкой, а аллель edf– высокой интенсивности роста. У всех животных, носителей аллеля edg, среднесуточный прирост находился в пределах 600-707 г, а у животных с аллелем edfон равнялся 746-808 г.
Не установлено четкой зависимости между группами крови свиней в системе Е и по затратам корма на прирост. Имеющиеся различия носят бессистемный характер и большей частью недостоверны.
В исследованиях на свиньях консолидированных линий из племзаводов «Кубань» и «Краснодарское» установлено, что в генетической системе А животные с антигеном А ср отличаются наименьшей скоростью роста и меньшей величиной среднесуточного прироста по сравнению с животными, не имеющими его. По средним данным двух племзаводов свиньи с генотипом А-/- при откорме до 100 кг живой массы превышали сверстников с генотипом А ср/ - по среднесуточному приросту на 15,4 г.
В генетической системе G не установлено существенных различий по интенсивности роста между животными с разными антигенными факторами и разными генотипами. В обоих племзаводах наибольшей скоростью роста отличались свиньи с генотипом К -/-, а самой низкой с генотипом К b/b. Разница между животными этих генотипов по среднесуточному приросту в двух племзаводах составила 58,6-33,8 г.
Высокой скоростью роста характеризуются также свиньи с генотипом К ас/-, которые превосходят животных с генотипом К b/b по величине среднесуточного прироста на 42,8-34,7 г.
Сравнение животных девяти различных генотипов генетических систем групп крови A, G, Kпо среднесуточному приросту со средними данными по стаду показало, что достоверно превосходят по скорости роста своих сверстников свиньи с генотипом К-/-(среднесуточный прирост выше на 21,5 г) и уступают сверстникам животные с генотипом К b/b( среднесуточный прирост ниже средних данных по стаду на 22,4 г).
В генетической системе Е в целом по двум племенным хозяйствам гомозиготные животные с генотипами aeg/aegи bdg/bdg дали по 693 г прироста в сутки, cгенотипом edg/edg – по 677 г и с генотипом edf/edf – по 655 г; гетерозиготные животные с аллелем edf (138 гол.) имели среднесуточный прирост 640-660 г, с аллелем aeg(142 гол.) – 647-669 г, с аллелем edg(254 гол.) – 653-679 г и с аллелем bdg (236 гол.) 665-679 г. Как видно, специфическим маркером высокой скороспелости свиней является аллель bdg, носители которого проявляют более высокую скороспелость среди гетерозиготных генотипов и максимальную среди гомозиготных.
Животные с генотипом bdg/bdg превосходили по среднесуточному приросту средний показатель по стаду на 20,8 г, средний уровень продуктивности животных с генотипом edf/edf– на 36,4 г.
Почти анологично проявляется маркирующее действие на скороспелость свиней и аллеля aeg. Гомозиготы aeg/aegимели наивысшей среднесуточный прирост, как и гомозиготы bdg/bdg(693 г), а гетерозиготы – носители аллеля aeg– занимали промежуточное положение между гомозиготами по величине среднесуточного прироста. При ютом самый низкий прирост имели гетерозиготы aeg/edf: разница между животными с генотипами aeg/aegи aeg/edf составляет 45,7 г. Генотипы aeg/bdgи aeg/edg занимали по среднесуточному приросту живой массы промежуточное положение между вышеуказанными генотипами.
Эти данные свидетельствуют, по–видимому, о тесном сцеплении маркирующих генов aeg, bdg и edg с группой генов, определяющих высокую скороспелость свиней. Противоположный по направленности эффект по скорости роста оказывает аллель edf. Носители этого гена занимают последнее место по скорспелости как среди гомозиготных генотипов. Разница статистически достоверна.
В обоих племзаводах выявлена тенденция более экономического использования кормов на прирост свиньями с генотипом А -/- по сравнению с животными, имеющими генотип А ср/-. По обобщенным данных двух хозяйств, разница между этими генотипами по затратам кормов на 1 кг прироста составила 0,09 корм. ед.
Наибольшие различия по оплате корма наблюдаются среди животных группами крови системы К. больше всего затрачивают кормов на прирост свиньи с генотипом К ас/-, а лучшую оплату корма имели животные с генотипами К-/- и Kac/b. В целом по двум хозяйствам разница между этими генотипами составила 0,26 корм. ед. в пользу животных с генотипом К-/-. Достоверные различия по оплате корма установлены также между генотипами Kac/bи К ас/- в пользу животных –носителей аллеля «b». Таким образом, наибольшую селекционную ценность по оплате корма в генетической системе А представляет генотип А-/-, в генетической системе К животные с генотипом К-/- и Kac/b, являющиеся маркерами лучшего использования кормов.
Максимальные различия по затратам кормов на 1 кг прироста между животными разных генотипов по группам крови в генетической системе Е составили 18,5 %. Следовательно, эффективность селекции по оплате корма с использованием групп крови свиней может быть довольно высокой. При такой направленности селекции в число наиболее желательных следует занести генотипы bdg/bdg( 3,74-3,84 корм. ед.), bdg/edf (3,89-3,95 корм. ед.), edg/edg(3,73-3,84 корм. ед.) и edg/edf( 3,96-3,97 корм. ед.). как видно, преимущественно это гомозиготные животные по аллелям bdgи edg и гетерозиготы с указанной парой аллелей. Гомозиготные животные по аллелям aeg и edf по оплате корма уступает гомозиготам – носителям аллелей bdg и edg– затрачивают на 1 кг прироста живой массы в обоих племзаводах более 4,0 корм. ед. Все гетерозиготные животные с аллелем aegтакже расходуют на 1 кг прироста свыше 4,0 корм. ед., причем самым высоким расходом кормов на единицу прироста по данным двух хозяйств (4,48 корм. ед.) характеризуются животные, объединяющие в своем генотипе оба аллеля – маркера низкой оплаты корма – aegи edf.
Изучение ценными, с точки зрения селекции свиней по оплате корма, являются животные с генотипом bdg/bdg, которые на каждом килограмме прироста лают экономию 0,19 корм. ед. по сравнению с другим генотипом aeg/aegи 0,32 корм. ед. по сравнению с худшим из гомозигот – генотипом edf/edf/. Гетерозиготы с аллелем bdg показывают лучшую оплату корма среди всех гетерозиготных животных(3,19-4,08 корм. ед.), за ними следуют гетерозиготы с аллелем aeg, маркирующим самую низкую оплату корма (4,08-4,48 корм. ед.).
При изучении сочетаемости генотипов нескольких генетических систем групп крови, наиболее тесно связанных с откормочными качествами свиней установлено, что наиболее удачным как по скороспелости, так и по оплате корма является сочетание у животного генотипов Kac/b и Ebdg/bdg. Таких животных среди 570 гол. выявлено 9.
Среднесуточный прирост у них был почти самый высокий(733 г), оплата корма наивысшей (3,62 корм. ед). в отдельности, без учета сочетания, у животных с генотипом Kac/bи Ebdg/bdg среднесуточный прирост равнялся 679-693 г, оплата корма – 3,78 -3,96 корм. ед.
Здесь мы имеем налицо как бы суммирующий эффект генотипов – маркеров высокой скороспелости и оплаты корма. Остальные генотипы генетической системы Е наиболее удачно сочетаются с генотипом К-/-. У животных, имеющих такое сочетание генотипов по группам крови, величина среднесуточного прироста при откорме до 100 кг достигает 697,7 – 750 г при затратах корма на 1 кг прироста 3,62-3,90 корм. ед. Таким образом, сочетание двух генотипов – маркеров высокой продуктивности – усиливают их маркирующий эффект. Выявление животных, сочетающих в себе эти генотипы, и интенсивное использование их в селекции позволит повысить ее эффективность и сократить сроки получение конечного селекционного достижения.
Сравнительная оценка маркирующего эффекта 13 антигенов и 20 генотипов по группам крови оплаты корма и скорости роста свиней консолидированных заводских линий, разводимых с использованием отдаленного инбридинга, и аутбредных генеалогических линий, показала, что специфическими маркерами высокой интенсивности роста только для свиней заводских линий являются гомозиготные генотипы Eaeg/aeg и Ebdg/bdg, а для свиней генеалогических линий - Eaeg/bdg, Eaeg/edf, Ebdg/edf; специфическими маркерами низкой скорости роста в заводских линиях был аллель edf, в генеалогических – аллель edg.
Общим для всех хозяйств была высокая интенсивность использования корма у животных с генотипом K -/- и Ebdg/edg, а низкая степень использования корма – у животных с генотипом Eaeg/edf. В остальных случаях маркирующий эффект групп крови имел специфическое действие для заводских и генеалогических линий. Так, например, свиньи с генотипом Eaeg/edfв заводских линиях характеризовались самыми высокими затратами корма на 1 кг прироста(4,48 корм. ед.), а в генеалогических линиях, наоборот, самыми низкими (3,85 корм. ед.).
Вычисление бисериальных коэффициентов связи с генотипами по группам крови и оплатой корма свиней из племзаводов «Кубань» и «Краснодарское» позволило установить, что на протяжении четырех лет в обоих племзаводах сохранялась устойчивая отрицательная связь оплаты корма только с генотипами К -/- и Ebdg/bdg. Это дает основание полагать, что маркирующее действие генотипов К -/- и Ebdg/bdg и на интенсивность использование кормов явление не случайное, а устойчиво проявляющееся длительное время во всех стадах и заслуживающее внимания для использования в селекции специализированных линий свиней.
Принимая во внимание тот факт, что в двух племенных заводах, занимающихся продолжительное время совершенствование свиней в одном направлении и одинаковыми методами, маркирующий эффект групп крови по скорости роста и оплате корма совпал практически полностью, склонны считать, что при одинаковом направлении отбора в одной и той же породе в косвенный отбор вовлекаются одинаковые генотипы по группам крови, маркирующий эффект которых можно использовать и в других хозяйствах, разводящих ту же породу и применяющих те же методы разведения.
При селекции крупной белой породы свиней в условиях России по скороспелости и оплате корма предпочтение следуют отдавать животным с сочетанием генотипов по группам крови К -/- и Ebdg/bdg.
При селекции свиней на повышение мясности предпочтительнее отбирать животных с аллелями Ga, Kb, Ebdgkmp, Eedfnkmpили имеющим генотип Ebdf/edg , Eedfhkmnp/edfhkmnp, которые являются маркерами низкой толщины шпика, большей длины туши, площади «мышечного глазка» и массы задней трети полутуши.
Имеется достаточно данных, свидетельствующих о том, что применение в оценке генотипа свиней определённого набора генов может повысить эффективность селекции в стаде до 20%, что превышает затраты на тестирование животных по группам крови в 10-15 раз. Владея методами определенного генотипа свиней в раннем возрасте, селекционеры способны целенаправленно ускорять селекционный прогресс в желательном направлении.
Иммуногенетическая характеристика племенных стад, заводских типов и линий свиней служит для селекционера богатейшим материалом для осуществления целенаправленного подбора с целью закрепления у животных хозяйственно-полезных признаков, свойственных родителям, а также повышение сохранности и продуктивности получаемого потомства.
В одном из опытов, цель которого заключается в получении у свиней гетерозиготного потомства по максимально возможному числу иммуногенетических факторов, в основу подбора пар был положен уровень гомозиготности хряков и маток по группам крови, трансферинам, церулоплазмину, амилазе. По каждому фактору желательным считался подбор при уровне гомозиготности 0,4-0,5; допустимы – при 0,6-0,7. Для большинства маток удалось подобрать по 1-5 желательных и допустимых вариантов спариваний с неродственными хряками.
От 95 маток получило 346 опоросов, из которых 123 желательные и допустимые и 233 нежелательные – с повышенной гомозиготностью у полученного потомства. У маток опытной группы в первом опросе многоплодие было выше на 0,69 гол., выход поросят к отъему на 1,67 гол., масса одного поросенка при отъеме- 2,7 кг; у маток с 2 и более опоросами, соответственно, на 0,88, 1, 23 гол. и 1,6 кг.
Таким образом, применяя сочетаемость родительских пар по группам крови, направленную на повышение гетерозиготности потомства, можно добиться реального и довольно существенного повышение репродуктивных качеств свиноматок.
При подборе пар учетом групп крови, по сравнению с другими формами подбора, у маток отмечается самая высокая оплодотворяемость и жизнеспособность приплода. Наибольшее число опоросов получают обычно у маток с низким уровнем гомозиготности, преимущественно которых над матками с высокой гомозиготностью по группам крови заключается еще и более высоком многоплодии(на 1,1-1,2 гол.). эти данные свидетельствуют о целесообразности получения таких маток и интенсивном их использовании, что способствовало бы снижению потребности в ремонтном молодняке почти на 20 %. Таким образом, иммуногенетический анализ стада позволяет определить у животных степень генетического сходства и использовать полученные данные при составлении подбора пар.
Г.Н. Сердюк, подбирая к маткам хряков с противоположными генотипами по группам крови, добился в условиях промышленных комплексов увеличения рождаемости гетерозиготного молодняка на 25-30% и это дополнительно сохранил в каждом до 3-4 тыс. поросят в год. В пометах, полученных маток генетически менее сходных с хряками(степень сходства 0,25-0,30), сохранность молодняка была на 11.6 -13,5% выше, чем при более высоком генетическом сходстве родителей, равным 0,4.
Своевременное выявление и выбраковка хряков, иммуносовместимых с большинства свиноматок, позволяет значительно повысить сохранность полученного приплода и сократить численность маточных стад.
Нормативы выполнения курсового проекта по технологическим условиям и зоотехническим показателям животных.
Таблица 1. Варианты показателей для курсового проекта
| Показатель |
Вариант 2 |
| 1. Годовой объем выращивания и откорма свиней, тыс.голов |
54 |
| 2.Производство свинины в живой массе в год, тыс. ц |
63 |
| 3. Ритм производства, дней |
2 |
| 4. Количество подсосных маток в группе, голов |
60 |
| 5. Оплодотворяемость маток, % |
80 |
Таблица 2. Варианты продолжительности подсосного периода доращивания, живой массы и отхода молодняка
| Показатели |
Вариант 2 |
| 1. Продолжительность подсосного периода (возраст при отъеме), дней |
35 |
| 2. За период до отъема до 4-х месячного возраста (доращивание), дней |
85 |
| 3. Живая масса при отъеме, кг |
8 |
| 4. Отход молодняка в группе сосунов, % |
5 |
| 5. Отход молодняка на доращивание (старше 4-х мечяцев), % |
2 |
| 6. Отход молодняка на откорме (старше 4-х месяцев), % |
1 |
| Итого отход молодняка для убоя, % |
8 |
Таблица 3. Средние показатели продуктивности животных
| Показатели |
Значение |
| 1. Многоплодие свиноматки за 1 опорос, голов |
8 |
| 2. Живая масса 1 головы в 4-х месячном возрасте при постановке на откорм, кг |
43 |
| 3. Среднесуточный прирост на откорме, г |
550 |
| 4. Живая масса при убое, кг |
110 |
Расчет ритмичного производственного потока в свиноводческом потомстве.
1.
Годовая потребность в поросятах:
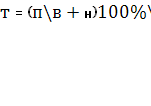 , ,
где Т – годовая потребность в поросятах, голов;
П – годовой план продажи свинины, ц;
В – средняя сдаточная масса 1 головы, ц;
Н – расход свинины внутри хозяйства и продажи своим рабочим и служащим, в случае наличия такой потребности, до 6% от годового производственного задания;
К – сохранность поросят до убоя, %
Т =63000/1,10*100%/ 92 = 62253 голов
2. Ц = Пс+Пн+П
Ц - цикл воспроизводства, дней;
Пс - продолжительность периода супоросности, дней;
Пн - продолжительность подсосного периода, дней;
П – период от отъема поросят до оплодотворяемости, дней (22 дня)
П = 35+114+22=171дней
3. С = 365/ Ц
С – количество опросов от матки в год;
365 – количество дней в году;
Ц – полный цикл воспроизводства матки;
С = 365/171 = 2
4. М =Т*Ц/ 365*А*Кс
М – среднее число маток, голов;
А – количество поросят при рождении на 1 опорос;
Кс – коэффициент сохранности
М = 62253*171/365*8*0,92 = 10645263/2686 = 3963 голов
5. Ко= Т/А
Ко – количество опросов за год по предприятию;
Ко = 62253/8=7781
6. Ц1= 365/Р
Ц1 - количество циклов в год;
Р – ритм производства:
Ц1 = 365/ 2 = 183
7. О1= М/Ц*Р
О1= количество опоросов в течении 1 ритма;
М = среднегодовое число маток на предприятии, голов
О1 = 3963/171*2 = 46
8. Мо = О1*100/ % опл.
Мо = 46*100/80= 58 голов
9. Мn = О1 – (О1*Км)
Мn = 46 – (46 *0,1) = 41 голов
10. М1= Мо*100/ 5*Р
М1 = 58*100/5 *2 = 580
11. ПС = Мn*А*Кс
ПС – число поросят – сосунов в технологической группе, голов
ПС = 41*8*0,95 = 312 голов
12. Гхол = (Д1+Д)/Р
Д – продолжительность пребывания на участке, дней
Д1 – количество дней дезенфекции
1.Гхол = (22+2)/2=12
2.Гсуп =(112+2)/2= 57
3. Гподсос = (35+5)2=20
4. Гдоращ=(85+5)/2=45
5. Г мол на откорме = (122+5)/2=63
Д = (110-43)/0,55=122
13. Пд = пс*кс = 312*0,98=306
14. По = пд*кс = 306*0,99=303
15.См=Ф*Г
См суп=58*57=3306
См хол = 580*12=6960
См подсос мат= 41*20 = 820
См пор-отъемышей=306*45=13770
См мол на откорме=303*63,5 =19240
Таблица 5. Одновременное поголовье по группам животных на комплексе (ритм 2 дня)
/Эээ\\эээ
| группа |
Продолжительность фазы, дней |
Количество дней дезинфекции |
Количество групп |
Количество животных в группе |
Общее количество скотомест |
| Матки холостые |
24 |
2 |
12 |
6960 |
| супоростные |
114 |
2 |
57 |
3306 |
| подсосные |
42 |
5 |
20 |
820 |
| Поросята-сосуны |
| Молодняк на доращивании |
90 |
5 |
45 |
13770 |
| Молодняк на откорме |
63 |
5 |
63 |
19240 |
Таблица 6. Технологические показатели комплекса
| показатель |
Производственный срок |
| ритм |
месяц |
год |
| Осеменение маток |
58 |
870 |
10440 |
| Получение опоросов |
46 |
690 |
8280 |
| Выбраковка маток |
- |
| Получение поросят |
312 |
4680 |
56160 |
Выращивание поросят
до……..дней
|
312 |
4680 |
56160 |
| До…….дней |
306 |
4590 |
55080 |
| Снятие молодняка с откорма |
303 |
4545 |
54540 |
| Снятие с откорма взрослых животных |
- |
| Сдача на мясо голов |
303 |
4545 |
54540 |
| ц |
454,5 |
6817,5 |
81810 |
Заключение
Селекция свиней, направленная на повышение генетического потенциала продуктивности, является основой интенсивного развития свиноводства, при котором достигается наивысшая эффективность производства, способствующая поддержанию высокой конкурентоспособности продукции на рынке.
Теоретические разработки и исследования показывают, что в проблеме интенсификации свиноводства наряду с выведением новых генотипов становится необходимым создание условий среды, обеспечивающих наиболее полное проявление генетического потенциала продуктивности. При этом центральное место занимают регулирование механизмов генетической и физиологической адаптации, контролирование взаимосвязи генотипа с окружающей средой.
Для дальнейшей интенсификации селекционного процесса становится необходимым изыскание новых приемов генетического потенциала, использование современных научных методов, таких как генетические маркеры, в качестве которых могут служить полиморфные системы групп крови, сывороточные белки, ферменты и др. Установлена связь между определенными генотипами по нескольким системам группы крови, эритроцитарными ферментами и продуктивностью. В геноме свиньи в последнее время известно 239 генетических маркеров, использование которых позволит значительно интенсифицировать селекцию.
Принимая во внимание, что пригодность к гибридизации существующих и создаваемых типов линий свиней определятся не только уровнем их продуктивности, но и консолидированностью генотипов, важное практическое и научное значение приобретает изучение маркирующего действия групп крови на откормочные качества свиней в генетических систем разводимых по принципу закрытых линий, с использованием инбридинга в отдаленных и умеренных степенях, а также при разведение свиней путем постоянно меняющихся кроссов генеалогических линий.
Список литературы:
1. Бажов Г.М. /Племенное свиноводство: уч.пособие/ .- СПб.: издательство «Лань», 2006.- 384 с.
2. Бекенев В.А./ Селекция свиней/. - Новосибирск, 1997.- 184 с.
3. Волкопялов Б.П. /Свиноводство/.- Издательство «Колос», Ленинград, 1968.- 431 с.
4. Ладан П.Е /Свиноводство/.- Издательство «Колос», Москва, 1978.- 304 с.
5. Уитеса А. И. /Приусадебное хозяйство. Свиноводство/.- Издательство «Лик пресс», Москва, 2001.- 208 с.
6. Ухвертов М.П., Тихонов И.Т., Мичурин В.П./Селекционные достижения в племенном свиноводстве/.- Госагропромиздат, Москва, 2000.-100 с.
7. Лесли Д.Ф. /Генные основы селекции сельского хозяйства/. – М, Колос, 1982. – 394 с.
|