Содержание
1. Функциональное описание …………………………………… 2
2. Морфологическое и информационное описание ………… 8
3. Генетико–прогностическое описание ……………………… 23
4. Анализ медико-биологических методов исследования ….. 26
5. Вывод ……………………………………………………….. 37
6. Список используемой литературы ……………………………. 38
1. Функциональное описание
Деятельность организма – это одновременное функционирования разнообразныхсистем и подсистем регулирования, подчиненное главной функции – обеспечению выживания в условиях изменяющейся внешней среды. При этом анализ результатов физиологических исследований позволяет легко обнаружить иерархическую организацию этих подсистем.
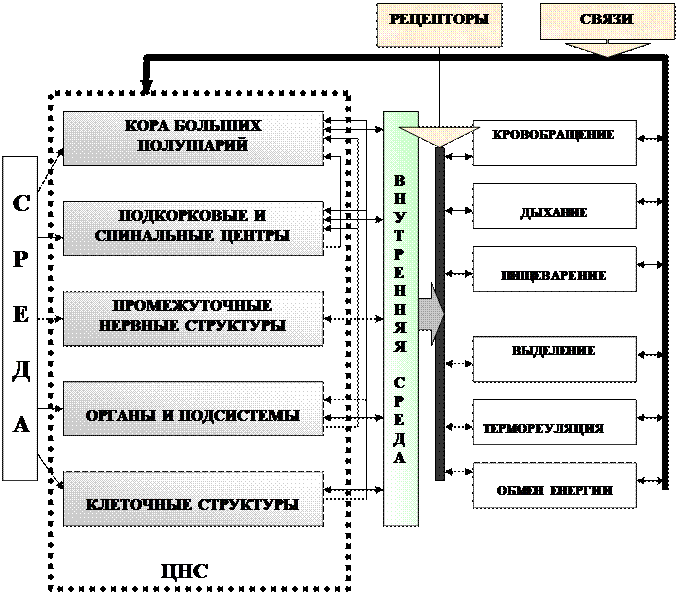
Схема 1.
Многоуровневое управление ЦНС, а также активизация определённых внутренних органов под влиянием нервной системы, что создаёт оптимальные условия во внутренней среде.
Центральная нервная система, состоящая из головного и спинного мозга, играет руководящую роль как в сфере взаимодействия организма с внешним миром, так и в отношении ко всем происходящим в организме сложнейшим процессам.
Основной механизм нервной деятельности – приём сигналов из внутренней и внешней среды, преобразование их в нервные импульсы, передача импульсов в нервные центры головного мозга, где они анализируются, и выработка ответной реакции, которая осуществляется при помощи исполнительных органов.
Нервная система обеспечивает связь организма с внешней средой. Все реакции организма на раздражения, поступающие из внешнего мира, обусловливаются деятельностью нервной системы. Повышение температуры окружающего воздуха приводит к усиленному притоку крови к коже и потоотделение, чем предотвращается перегревание организма. Наоборот, понижение температуры приведёт к сужению сосудов и уменьшению теплоотдачи. Кроме того, высший отдел нервной системы – полушария головного мозга являются органом мышления и психической деятельности.
Нервная система состоит из спинного и головного мозга. Спинной мозг находится в позвоночном канале и представляет собой тяж, который вверху переходит в продолговатый мозг, а внизу заканчивается на уровне второго поясничного позвонка. Спинной мозг состоит из серого вещества, содержащего нервные клетки, и белого, образованного нервными волокнами. Серое вещество расположено внутри спинного мозга и со всех сторон окружено белым веществом. В центре спинного мозга находится узкий канал, содержащий спинномозговую жидкость. Есть передние и задние рога, а также в грудном отделе выделяются боковые рога, в которых заложены тела нейронов, иннервирующих внутренние органы. Спинной мозг имеет два утолщения – шейное и поясничное, от которых отходят нервы к верхним и нижним конечностям. От спинного мозга отходит 31 пара спинномозговых нервов.
Основные функции спинного мозга – рефлекторная и проводниковая. С помощью рефлекторной деятельности осуществляются простые двигательные рефлексы (сгибание, разгибание…). В шейных сегментах расположены центры рефлекторных движений диафрагмы, мышц шеи, плечевого пояса и верхних конечностей, в грудных сегментах – центр межреберных мышц и мышц туловища, в поясничных и крестцовых сегментах – центры мышц тазового пояса и свободных нижних конечностей. В боковых рогах серого вещества грудного и поясничного отделов находятся сосудодвигательные центры и потоотделения, в крестцовом – центры мочеиспускания и др.. Проводниковая функция заключается в том, что по проводящим путям спинного мозга в головной мозг передаются импульсы от рецепторов кожи, мышц и внутренних органов (восходящие пути), а из головного мозга - в спинной, затем на периферию к органам (нисходящие пути). При повреждении спинного мозга указанные функции нарушаются (выпадение рефлексов, параличи и др.). Деятельность спинного мозга находится под контролем головного мозга, регулирующего спинномозговые рефлексы.
В головном мозге различают 5 отделов: продолговатый мозг, собственно задний (мост и мозочок), средний, промежуточный и большие полушария мозга. В мозге находятся 4 сообщающихся между собой полости – мозговые желудочки, заполненные спинномозговою жидкостью: первый и второй желудки расположены в больших полушариях, третий в промежуточном, а четвёртый в продолговатом и заднем мозге.
Продолговатый мозг – продолжение спинного мозга, выполняет две функции: рефлекторную и проводниковую, имеет важное значение в регулировании сердечной деятельности, сосудов и дыхания. Здесь же находятся центры жевания, сосания, глотания, отделения слюны и желудочного сока, а также защитных рефлексов (чихание, рвота, кашель). Проводниковая функция заключается в передаче импульсов из спинного мозга в головной. Продолговатый мозг – жизненно важный отдел центральной нервной системы, его повреждение может быть причиной смерти (прекращение дыхания и деятельности сердца).
Задний мост образован мозочком и мостом. Мост выполняет проводниковую функцию. Мозочок находится позади продолговатого мозга. Он координирует движения, делает их чёткими и плавными, играет важную роль в сохранении равновесия тела в пространстве, а также оказывает влияние на тонус мышц. Мозочок принимает участие в регуляции некоторых вегетативных функций (состав крови, сосудистые рефлексы). Деятельность мозочка также контролируется корой больших полушарий.
В среднем расположены первичные (подкорковые) центры зрения и слуха, которые осуществляют рефлекторные ориентировочные реакции на световые и звуковые раздражения. Эти реакции проявляются в различных движениях туловища, головы и глаз в сторону раздражителей. Средний мозг регулирует и распределяет тонус мышц.
Промежуточный мозг расположен впереди среднего мозга. Здесь находятся чувствительные пути, проводящие импульсы от рецепторов тела к коре больших полушарий. Гипоталамус является высшим центром регуляции вегетативных функций, всех видов обмена веществ, температуры тела, постоянства внутренней среды, через гипофиз регулирует деятельность других желез внутренней секреции – щитовидной, надпочечников, половых.
Большие полушария – наиболее крупный и развитый отдел головного мозга. Серое вещество покрывает полушария снаружи и образует кору головного мозга. Под корой находится белое вещество. Оно состоит из нервных волокон, связывающих кору с расположенными ниже отделами центральной нервной системы и отдельные доли полушарий между собой. Кора имеет извилины, разделённые бороздами на доли: лобную, теменную, височную и затылочною. Возбуждения от разных рецепторов поступает в соответствующие воспринимающие участки коры, называемые зонами, и отсюда передаётся к определённому органу, побуждая его к возбуждению. Таким образом, полушария головного мозга - это высший отдел центральной нервной системы, контролирующей работу всех органов. В коре заключаются условно рефлекторные связи, поэтому она является органом приобретения и накопления жизненного опыта и приспосабливает организм к постепенно меняющимся условиям внешней среды. Мыслительная, речевая деятельность и память связаны с функциями коры. От головного мозга отходит 12 пар черепно-мозговых нервов, по выполняемым функциям одни из них являются чувствительными (обонятельный, зрительный, слуховой), другие – двигательными (блоковый, отводящий, подязичный), третьи – смешанными (тройничный, блуждающий).
Нервная система условно подразделяется на две части – соматическую и вегетативную. Соматическая инвертирует скелетную мускулатуру (двигательные реакции на раздражения), обеспечивает связь организма с внешней средой и быструю реакцию на её изменения. Вегетативная нервная система иннервирует гладкую мускулатуру внутренних органов, сосудов, кожи, мышцы сердца и железы, стимулирует работоспособность скелетных мышц. Вегетативная нервная система управляет деятельностью внутренних органов, участвующих в осуществлении функций питания, дыхания, выделения, циркуляции жидкостей и приспосабливает их работу к потребностям организма и условиям внешней среды.
Вегетативная и соматическая нервные системы состоят из центральных и периферических образований. Вегетативная нервная система подразделяется на два отдела: симпатический и парасимпатический. Как правило, органы и ткани иннервируются из обоих отделов, таким путём осуществляется более надёжная регуляция деятельности органов.
Симпатический отдел усиливает окислительные процессы, потребление питательных веществ, стимулирует дыхательную и сердечную деятельность. Роль парасимпатического отдела охраняющая: сужение зрачка при сильном свете, торможение сердечной деятельности, опорожнение полостных органов.
Таблица 1. Влияние нервной вегетативной системы на деятельность некоторых органов
| Орган
|
Симпатические
|
Парасимпатические
|
| Сердце |
Учащение ритма и усиление сокращений |
Урежение ритма и ослабление сокращений |
Кровеносные
сосуды
|
Сужение |
Расширение |
| Зрачок |
Расширение |
Сужение |
| Железы желудка |
Торможение |
Усиление секреции
|
| Гладкие мышцы желудка и кишечника |
Торможение сокращений |
Усиление сокращений |
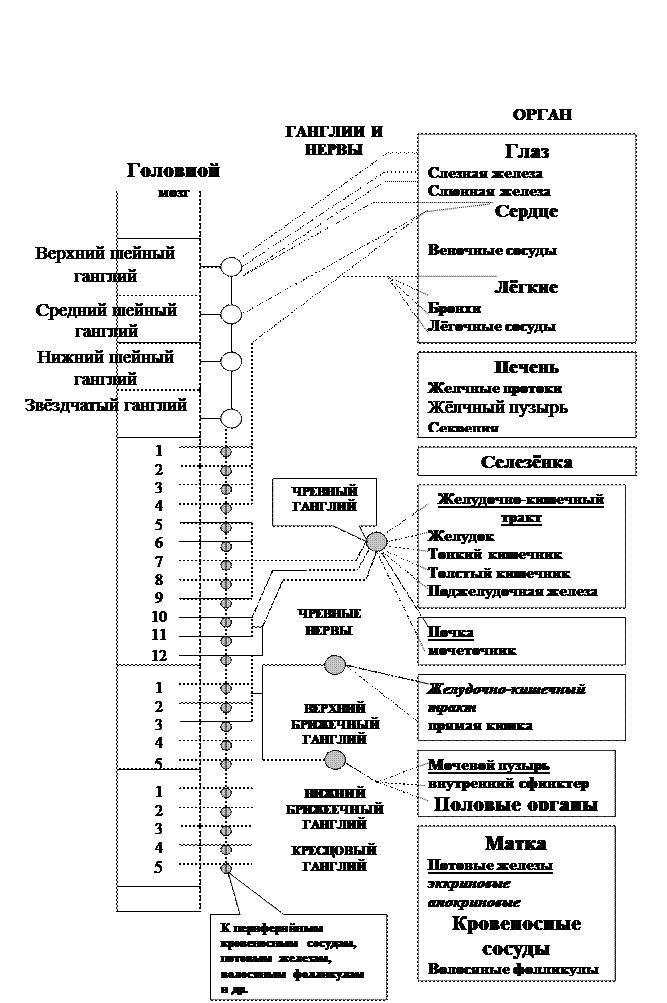
Схема 2.
Пример симпатического отдела вегетативн6ой нервной системы.
Волокна выходят из спинного мозга, входят в периферийные нервные узлы (ганглии) и иннервируют различные органы. Сплошными линиями обозначены холинэргические волокна, пунктирными – постганглионарные адренергические.
Нервная система регулирует деятельность всех органов и систем организма. Работа каждого органа – отделение секрета железами, сокращение скелетных мышц и сердца – происходит под влиянием нервной системы. Она согласовывает их деятельность, обеспечивает единство организма в целом. При изменении деятельности одного органа (или системы органов) происходят изменения в деятельности других органов (или систем).
Наш организм может жить и развиваться лишь в том случае, если между ним и средой обитания происходит постоянный обмен веществ.
Внутренняя же среда создаёт условия для “свободной и независимой” жизни с помощью НС. Нервная система сама формирует для организма свою внутреннюю среду, но формирование это происходит под постоянным и непрекращающимся воздействием со стороны окружающего мира.
2. Морфологическое и информационное описание
Иерархичность структуры организма приводит к тому, что взаимодействие нервной системы с органами и подсистемами строится на принципе последовательности уровней, через которые проходят управляющие сигналы к исполнительным механизмам регуляции. Такое многоуровневое управление более экономично, чем жестко централизованное. Оно сохраняет принцип централизации управления со стороны внешних уровней при относительной независимости (автономности) функционирования низших уровней (подсистем).
В течении многих лет перед исследователями возникал вопрос о том, как проходит взаимодействие во внутренней среде – кровь, лимфа, тканевая жидкость, а главное – биологически активные вещества, которые синтезируют и расщепляют ферменты, связывающие и освобождающие из связанной формы механизмы. Так, например, нелегко ответить, почему в крови удаётся обнаружить свободный ацетилхолин при наличии мощных холинэстераз или обнаруживается гистамин наряду с диминоксидазой, почти мгновенно её расщепляющей.
Медиаторы нервного возбуждения (например, ацетилхолин, норадреналин, серотонин, гамма-аминомасляная кислота и др.), образующиеся нервными окончаниями и передающие нервный импульс с нейрона на клетку-исполнитель (синоптическая передача), избежав ферментативного расщепления или обратного поглощения, поступают в ток крови и разносятся по всему организму. Здесь они начинают свою вторую жизнь, теперь уже в качестве биологически активных веществ. Рецептор, принимающий центробежные нервные импульсы, можно рассматривать как устройство, через которое специфическая информация поступает из нервных окончаний в клетку-исполнительницу.
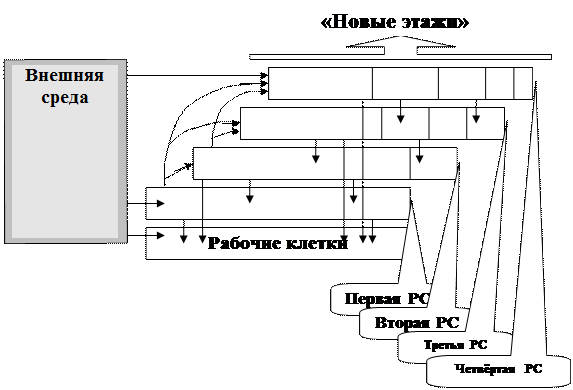
Схема 3
. Развитие взаимоотношений РС. С внешней средой взаимодействуют рабочие клетки первой и четвёртой РС. Все РС связаны между собой и с рабочими клетками. “Новые” этажи развивались главным образом на 3 и 4 РС и не имеют непосредственной связи с рабочими клетками.
В развитии регулирующих систем есть много общих закономерностей:
1) Регулирующая система (РС) возникает при новых условиях существования организма и появлении новых функций;
2) Чем “моложе” система, тем более специализировано её действие, тем уже круг типов клеток, которые она регулирует, тем короче период действия. Первая РС непрерывно воздействует на все клетки; вторая также действует на все клетки, но её эффект весьма изменяется во времени; третья регулирует внутренние органы и сосуды; четвёртая – только поперечно-полосатую мускулатуру;
3) Все РС развиваются в процессе эволюции, но особенно быстро новые, и в первую очередь четвёртая;
4) Клетки новой РС находятся под воздействием старых, но в тоже время оказывает на них и обратное воздействие;
5) Новые РС получают информацию через свои рецепторы или от старых РС. Каждая система имеет свои эффекторы или воздействует через старые системы;
6) В развитии каждой РС можно наметить несколько этажей, объединённых одинаковым принципом действия.
В процессе эволюции разные отделы РС развивались неравномерно. Над первоначальными элементами РС, осуществлявшими на периферии самые простые функции, по мере количественного их возрастания появились неустройки – этажи которые уже регулируют эти первоначальные клетки РС. Назначение неустроек – выделение ”высших” пространственных и временных кодов, моделирования, как внешнего мира, так и собственных действий. Вертикальные связи осуществляют субординацию между этажами. Горизонтальные связи обеспечивают циркуляцию информации между соответствующими или близкими этажами разных РС. Кроме того, некоторые этажи самых систем поделились по вертикали на два отдела, действующие в различной степени противоположно- возбуждающий и тормозящий отделы, как, например, симпатический и парасимпатический отделы вегетативной нервной системы.
Первая РС представляет собой среду организма – кровь и лимфу – вместе с рабочими внутренними органами, поддерживающими её состав. Вторая РС эндокринная, состоящая из нескольких этажей: вверху -–гипофиз, ниже специализированные железы (половые), ещё ниже – обще клеточные (например, кора надпочечников, щитовидная железа), ещё ниже – специализированные, выполняющие частные задачи (поджелудочная железа). В самом низу – эндокринные функции некоторых органов, имеющих местное значение. Третья РС – вегетативная нервная система. Она состоит (сверху вниз) из: высших вегетативных центров ВВЦ, с симпатическим С и парасимпатическим ПС отделами, главных центров, ведающих отдельными функциями (например, дыхательный центр), сегментарных углов, местных сплетений в органах. Четвёртая РС – нервно-соматическая система – понятная из рисунка РФ – ретикулярная формация.
Первая РС – система неспецифической химической регуляции. Назвать её системой можно только условно, поскольку в неё входят в разной степени все клетки, в процессе соей жизнедеятельности изменяющие химический состав внутренней среды организма. Первая РС – это кровь и лимфа с их простыми химическими составляющими: вода, соли, газы, простейшие питательные вещества – глюкоза, аминокислоты, белки, продукты распада, различные “шлаки”. Первая РС имеет постоянный состав, РН, осмотическое и онкотическое давление, парциальное давление газов. Постоянство основных компонентов среды регулируется высшими РС, но некоторые из них поддерживаются рабочими клеткамиза счёт рецепторных и эффекторных свойств последних.
Так, можно полагать, что клетки почек сами способны поддерживать определённый уровень “шлаков” в крови помимо влияния РС. Тоже самое касается роли печени в поддерживании уровня сахара. Такое регулирование весьма относительно. Восприятие и переработка информации осуществляется внутри рабочих клеток. Нужно только одно непременное условие – поддерживание циркуляции среды.
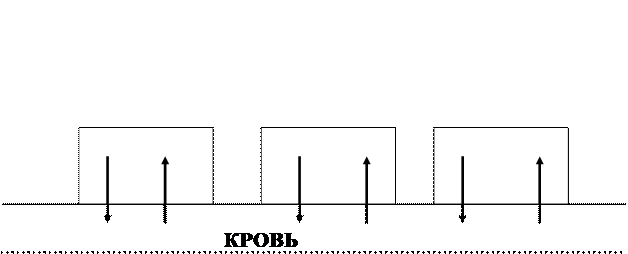
Рисунок 1.
Схема отношений между органами через кровь. Первая регулирующая система. Рабочие органы а, б, в выделяют в кровеносное русло вещества в количествах, зависящих от их содержания в крови.
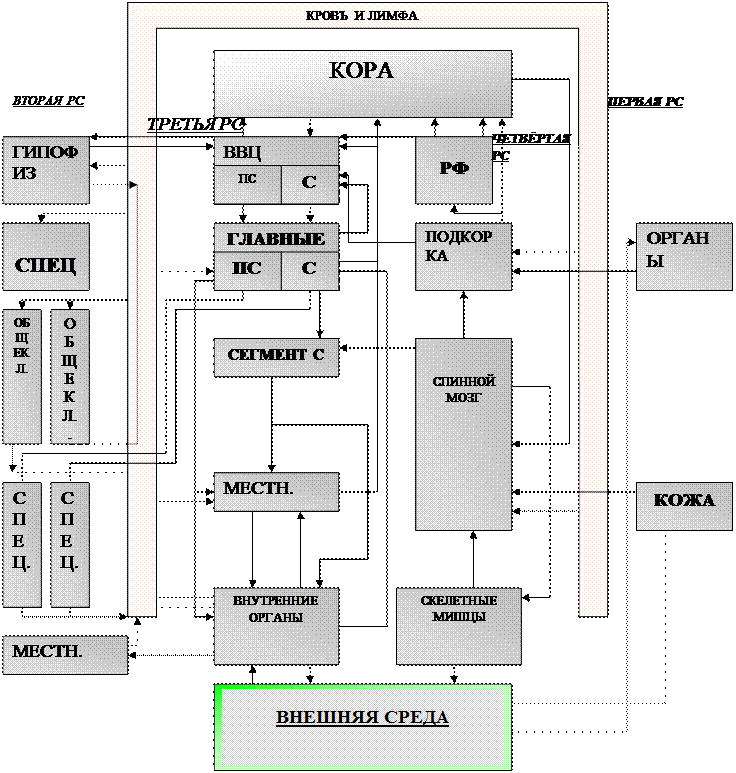
Схема 4.
Упрощённая схема регулирующих систем организма.
Третья РС – нервно-вегетативная. Основной принцип её действия: химия-нерв-химия. Нервные окончания (рецепторы) воспринимают изменения химизма в тканях, преобразовывая их в нервные импульсы, которые на периферии снова превращаются в активное химическое вещество – медиатор вызывающий специфическую реакцию в рабочей клетке. Нервная клетка преобразовывает информацию в нервный импульс – взрыв химических реакций, взрывающихся с положительными обратными связями.

Рисунок 2
. Схема нервной клетки местного сплетения вегетативной нервной системы (третья РС). Она получает раздражитель через рецептор Р из клеток и действует на них же своими окончаниями – эффекторами Э, выделяющими активное химическое вещество – медиатор. Кроме того клетка получает регулирующие импульсы “сверху” через А и посылает сигналы через Б.
Возбуждение - это рабочая деятельность нервной клетки на её первом этаже. Накопление изменений до возбуждения – это кратковременная память. Значение второго этажа очень велико: при частых возбуждениях клетка немного гипертрофируется, причём изменяется не столько всё её тело, сколько избирательно все её части под синапсами, через которые воспринимается возбуждение от других клеток. В этом длительная память клетки.
Она различна на разных этажах нервной системы.
Деятельность третьего клеточного этажа весьма ограничена – клетка способна к регенерации своих частей, но не к размножению. В нервной вегетативной системе можно выделить четыре этапа. Низший этап – местное сплетение в органах (рисунок 3
). Они самые древние, с меньшей специфичностью деятельности. Восприятие информации на этом этаже происходит за счёт отростков или, может быть, самых тел клеток. Кроме химизма окружающей среды они воспринимают информацию “сверху”- со стороны выисших этажей нервной системы. Программа переработки информации – жесткая, простая. Такие нервные сплетения способны осуществлять простую регуляцию некоторых органов. Например, сплетения кишечника – обеспечивают её перистальтику, обеспечиваю сердечную частоту сокращений, клетки в стенке сосудов – изменяют её просвет.

Рисунок 3
. Схема сегментарных и меж органных сплетений (второй этаж третьей РС) НК-1 – клетка местного сплетения (первый этаж), НК-2 и НК-3 – клетки сегментарного сплетения. Они получают воздействия или из первого этажа или непосредственно из тканей и также воздействует на них.
Второй этаж – сегментарные центры и крупные межорганные сплетения (рисунок 4
). Структура их может быть сложной. Информацию получают “снизу” – от местных сплетений или от собственных рецепторов, от ”соседей” таких же клеток и “сверху” - от вышележащих центров. Программа переработки тоже жесткая, но более разнообразная:
a) Роль сервомотора – усиление “приказов” сверху;
b) Пространственное и временное суммирование раздражителей;
c) Передача информации в вышележащие этажи;
d) Передача возбуждения на рабочие клетки через местные сплетения или непосредственно;
Программа осуществляется за счёт структуры и порогов возбудимости. В них заложены “модели” деятельности. Возможна цикличность деятельности благодаря обратным связям. Приспособление - через перестройку (гипертрофии) клеток.
Третий этаж вегетативной нервной системы условно называется уровнем “главных” центров. Главный центр – это сосредоточение программы управления одной функциональной системой (дыхательной, сердечно-сосудистой).
На этом уровне появляется две системы клеток – симпатическая и парасимпатическая. Общий принцип действия их одинаковый – химический, но действие более или менее противоположно, хотя точки приложения медиаторов на “химическую цепь” не одни и те же (рисунок 4
).
Рисунок 4.
Схема “химической цепочки” в рабочих клетках на разные звенья которой действуют медиаторы симпатического С и парасимпатического ПС отделов вегетативной нервной системы. Их действие противоположно по знаку, но имеет разные точки приложения, поэтому в некоторой степени они дополняют друг друга.
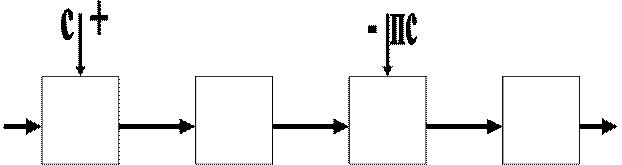 Возможно, что парасимпатический отдел более поздний и связан с развитием соматической нервной системы. Возможно, что парасимпатический отдел более поздний и связан с развитием соматической нервной системы.
Центр – это комплекс клеток, имеющих между собой прочные связи, хотя и необязательно расположенных рядом, но обеспечивающих одну программу. В некоторых главных центрах имеются два связанных между собой ”подцентра” – симпатический и парасимпатический. Возбуждение одного тормозит другой. Оба имеют связи с высшими и низшими этажами.
Главный центр выполняет большую программу переработки инофырмации. В результате в нём вырабатываются группы импульсов, обеспечивающих циклическую деятельность рабочих органов. Программа состоит из:
a) Самостоятельной деятельности центров – в них возникают импульсы и без внешних воздействий;
b) Восприятия возбуждающих и тормозящих воздействий из органов
c) Восприятие аналогичных воздействий из вышестоящих этажей нервной системы;
d) Возбуждение и торможение гормонами.
Программа воздействия на рабочие органы осуществляется через симпатический и парасимпатический отделы, возможно с разной частотой импульсов. Непосредственное претворение регулирующего действия идёт через сервомотор первой и второй ступени и через специализированные эндокринные железы.
Четвёртый этаж – высшие вегетативные центры (ВВЦ). Горизонтальными связями они тесно связаны с высшими этажами четвёртой РС – подкоркой, ретикулярной формацией, с “эндокринным мозгом” – гипофизом. Через вертикальные связи они соединены с корой и уровнем главных центров. ВВЦ имеет отношение к психике мышления. Их основные назначения: выделение высших кодов регулирования – пространственное и временное интегрирование информации, поступающей со стороны тела и со стороны четвёртой РС, для управления внутренними органами.
Цель – обеспечение программы внешней деятельности организма, осуществляемой скелетной мускулатурой под управлением четвёртой РС. В тоже время программы самой четвёртой РС находятся под воздействием третьей РС, которая не только обеспечивает обратную связь, но и часто играет роль пусковых организмов, включающих программу поведения, т.е. в них заложена значительная часть “программы жизни” – инстинктов.
Схема 5.
Схема высших вегетативных центров (высших этажей вегетативной нервной системы – третья РС). ВВЦ и главные центры ГЦ состоят из симпатических С и парасимпатических ПС отделов. Некоторые органы (А,Б) получают импульсы от обеих отделов, другие (В,Г) только от симпатического.
На уровне ВВЦ чувствительная и двигательная сферы разделены. Каждая имеет свою структуру и программу. Чувствительную сферу можно представить себе как комплекс центров чувств и ощущений, опять-таки понимая под центрами сочетания клеток, объединенных одной функцией, а не соседством расположения. Чувствительная вегетативная сфера тесно связанная с подкорковыми соматическими чувствительными центрами из четвёртой РС.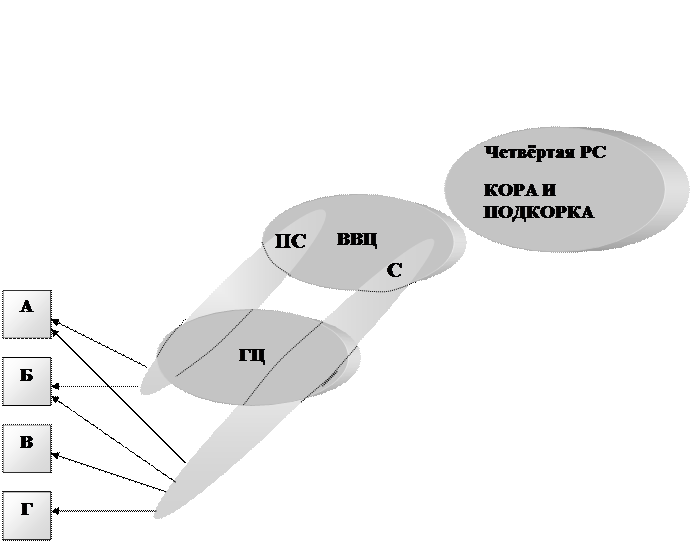 Можно говорить о целой гамме чувств и ощущений: голод – сытость, жажда. Утомление, удушье, холод – тепло, боль и т.д. Все они представлены в вегетативном (ВВЦ) и соматическом отделах подкорки и является структурами модели, связанными с различными центрами на периферии, с нижележащими этажами (ступенями) третьей и четвёртой РС. Кроме специализированных центров есть ещё два общих – центры “приятного” (ПР) и “неприятного” (НПР). Они противоположны друг другу и связаны обратными связями – возбуждение одного тормозит другой. С ними связано несколько моделей-структур, представляющих собой центры эмоций – страха, гнева, горя, радости. Большинство из этих центров имеют представительство в коре, посылая в неё и получая обратно сигналы. Есть центры которые и не представлены в коре. Всякое возбуждение определено какой-то степенью возбуждения одного или нескольких центров и центра ПР или НПР. Можно говорить о целой гамме чувств и ощущений: голод – сытость, жажда. Утомление, удушье, холод – тепло, боль и т.д. Все они представлены в вегетативном (ВВЦ) и соматическом отделах подкорки и является структурами модели, связанными с различными центрами на периферии, с нижележащими этажами (ступенями) третьей и четвёртой РС. Кроме специализированных центров есть ещё два общих – центры “приятного” (ПР) и “неприятного” (НПР). Они противоположны друг другу и связаны обратными связями – возбуждение одного тормозит другой. С ними связано несколько моделей-структур, представляющих собой центры эмоций – страха, гнева, горя, радости. Большинство из этих центров имеют представительство в коре, посылая в неё и получая обратно сигналы. Есть центры которые и не представлены в коре. Всякое возбуждение определено какой-то степенью возбуждения одного или нескольких центров и центра ПР или НПР.
Двигательная сфера или точнее “сфера действий” ВВЦ представлена структурами – моделями сложных безусловных рефлексов, часть которых относится к вегетативной нервной системы, часть – к соматической. Они осуществляют много программ действия. Одни универсальные включающиеся от центров ПР, НПР и эмоций, другие – специализированные. Работающие в паре со специальными центрами ощущений и чувств. Большинство соматических и двигательных центров подкорки подчиняются коре, вегетативные, наоборот, чаще всего не имеют коркового контроля. Понятие модели-структуры, осуществляющей определённую программу действий, более соответствует действительности, чем старый термин “двигательный центр”, то есть, с одной стороны, модель может объединять клетки различных участков нервной системы, а с другой стороны, одни и те же клетки могут входить в несколько моделей, участвуя у нескольких программах действий.
Кроме всех перечисленных структур чувствительной и двигательной сфер есть ещё одно образование – ретикулярная формация (РФ), не относящаяся ни к одной из них. Назначение – осуществлять механизм выбора и усиления одной программы при одновременном торможении всех других. Этот механизм чрезвычайно важен, т.е. за счёт его организм выбирает из массы программ одну, имеющую наибольшее значение в данный момент времени. РФ регулирует все отдели центральной нервной системы. В коре она осуществляет внимание, в спинном мозгу, а также в ВВЦ усиливает одни и тормозит другие рефлексы.
Механизм действия РФ не совсем ясен, поскольку неясна природа торможения: есть ли специальная тормозная природа клеток, или тормозные сигналы. Либо же тормозной эффект получается на обычных синапсах за счёт особого ритма. Так или иначе, усиливающий эффект РФ осуществляется по принципу положительной обратной связи. Переключение на тормозной эффект осуществляется вследствие “утомления” клеток РФ или наиболее чувствительного центра.
Схема связей ВВЦ. Они состоят из центров чувств и действий, получающих от уровня главных центров их чувствительного отдела, от коры и от двигательных подкорковых центров четвертой РС. Кроме того они связаны с ретикулярной формацией (РФ), состоящей из двух частей - облегчающей (ОБЛ) и тормозящей (ТОР), которые действуют противоположно и взаимно тормозят друг друга (Г). РФ усиливает одну из текущих программ и ослабляет другие. Кроме того, чувствительные центры связаны с универсальными центрами чувств “приятного” (ПР) и “неприятного” (НПР), которые тоже взаимно тормозят друг друга. Они связаны с центрами эмоций, включающими свою программу действий. Имеются связи со второй РС.
Четвёртая регулирующая система – соматическая, или анимальная, нервная система.
Её основное назначение – управление скелетной мускулатурой и восприятие и переработка информации из внешнего мира.
Высший этап переработки, в котором суммируется информация из вне и изнутри, и представляет собой процесс мышления, осуществляемый в коре.
Разберём отношение четвёртой РС к регулированию внутренних органов.
Очень грубо соматическую нервную систему можно разделить на три этажа:
- Спинной и продолговатый мозг;
- Подкорковые образования;
- Кора.
На первом уровне осуществляются простые безусловные рефлексы с врождёнными программами переработки информации, поступающей с различных участков тела.
Схема 6.
Схема связи ВВЦ с другими отделами подкорки.
Сегменты спинного мозга связаны с соответствующими сегментами третьей РС и её местными сплетениями. Здесь осуществляется первый уровень связи.
Подкорка интегрирует простые рефлексы в модели сложных двигательных актов: ходьба, равновесие, даже целые комплексы в системе движения, связанных с эмоциями (мимика, защита, нападение). У человека двигательная сфера находится в коре. В подкорке происходит первичная обработка информации, поступающей из вне через действующие на расстоянии рецепторы – слух, обаяние, зрение. Видимо она и обрабатывает, но врождённой, очень сложной программой, давая сигналы на включение соответствующих двигательных моделей. Чувствительная и двигательная сферы подкорковой четвёртой РС тесно связаны с центром приятного (ЦП) и неприятного (ЦНП), центрами эмоций и ВВЦ третьей РС.
Важнейшей частью нервной системы является кора головного мозга. Хотя она относится к четвёртой РС, но оказывает приличное влияние и на третью РС. Кора способна сама управлять инстинктами и полностью управлять поведениями человека, хотя эта способность реализовывается далеко не у всех и не всегда.
Функции коры:
1. В коре собирается информация, собирающаяся извне и изнутри, вернее, от промежуточных станций в подкорке. В ней имеются представительства центров чувств, эмоций, приятного (ЦП) и (ЦНП).
2. Кора осуществляет функцию памяти:
-кратковременной, когда в течении некоторого времени возбуждения “циркулирует” по клеткам, составляя модель внешнего или внутреннего раздражителя;
-длительной – когда в результате повторного возбуждения какой-либо части временной модели в клетках, составляющих её, образуются структурные изменения, обеспечивающие распространение возбуждения.
3. Между моделями расположенными в разных участках коры, возбуждающимися одновременно, или близко по времени, возникли условные связи по типу той же кратковременной и длительной памяти. В результате образуются сложные комплексные модели, объединенные только общностью времени возникновения возбуждения.
4. В коре осуществляется большая программа обработки информации с выделением и запоминанием целой гаммы высших кодов и моделей. Таким образом кора является моделирующим устройством для внешнего и внутреннего мира и устанавливает связи между ними.
5. В двигательной области коры сосредоточены модели двигательных актов (программ), расположенных в несколько этажей. Реализация движений осуществляется под контролем обратных связей.
6. Возбуждение корковых моделей внутреннего мира извне, от условных раздражителей даёт такой же эффект в подкорке, как и действие внутренних раздражителей (условный рефлекс). В обычных условиях он слабее “настоящего”, но при тренировке может оказаться таким же и даже более сильным. Возбуждение корковых центров чувств со стороны коры вызывает такие же сдвиги во внутренних органах, как и возбуждение этих центров “снизу”.
7. У человека особенно развит механизм внимания, представляющий собой усиление возбуждения одной модели за счёт торможения всех других. Этот механизм осуществляет РФ.
8. Клетки, составляющие корковую модель, при частом возбуждении и усилении могут гипертрофироваться и стать источником самопроизвольного возбуждения. Таким образом, корковая модель становиться сильнее реальных раздражителей и вызывает те же изменения в регулировании внутренних органов, что и истинный внешний или внутренний раздражитель.
Рассматривая четвёртую РС, нельзя обойти вопрос о сознании. Сознание – это выделение среди большого количества параллельно идущих в коре процессов переработки информации, одного, который доминирует и усиливается над всеми другими в каждый данный момент. Это осуществляется за счёт механизма внимания, выполняемого через верхние отделы РФ. Иначе: сознание – это способность выделить и усилить одну мысль среди множества других, одновременно идущих в коре, а мышление – это содержание самых процессов переработки информации. Механизмы усиления построены таким образом, что одни и те же группы клеток не могут усиливать статически, длительное время. Усиливающие импульсы всё время переключаются с одних моделей – образов, на другие.
Однако, кроме основного, усиление РФ, пути информации перерабатываются ещё в огромном количестве других клеток. Это и есть “подсознание”. Когда человек внимательно слушает, то, кажется, что он в это время не видит. Действительно, зрительный канал получения информации заторможен РФ, усиливающий в это время слуховой. Но торможение неполное, информация всё равно поступает и перерабатывается. И так по всем другим каналам. Разумеется, модели в подсознании возбуждаются в гораздо меньшей степени и поэтому импульсы циркулируют только по хорошо проходимым путям. Примером может служить выполнение сложных, но хорошо заученных двигательных актов без привлечения внимания.
В подсознательной сфере коры регулируется огромное количество разнообразных рабочих процессов, в первую очередь происходящих во внутренних органах. В подсознании всегда могут быть скрытые очаги повышенной возбудимости, если в них постепенно поступает поток импульсов с периферии. Даже не привлекая внимания они могут существенно влиять на сознание, менее общую настройку коры. Это особенно важно для патологии.
3. Генетико–прогностическое описание
В процессе эволюции возникли многоклеточные существа (второй уровень систем), в которых клетки организовались в определённые группы по однородности выполняемых функций. Создались структуры из клеток. Несравненно большие изменения потерпели регулирующие элементы. Первые многоклеточные организмы представляли собой структуру с замкнутой полостью по средине (но при сохранении выхода всех клеток наружу), дополненной жидкость, которая и била первой внутренней средой организма. Клетки, таким образом, получили возможность взаимодействовать друг с другом не только через непосредственное соприкосновение, но и через эту среду, выделяя в неё продукты своего обмена. Так возникла первая регулирующая система – химиконеспецифическая. Действующими началами в системе являлись продукты обмена, общие для всех клеток: ионы, газы, простые химические вещества.
На следующем этапе эволюции часть клеток оказалась внутри, и полностью утеряли связь с внешней средой. Их жизнь стала зависеть от наружных клеток, обеспечивающих определённый состав внутренней жидкости. Если он отклонялся от обычного, внутренние клетки отвечали резким нарушением обмена и выделением в жидкую среду различных промежуточных продуктов. Эти продукты и стали в дальнейшем гормонами, активирующими деятельность внешних клеток, т. е. Регулирующими их. Так возникла вторая регулирующая система - эндокринная (рисунок 6
).

Рисунок 5
. Первая регулирующая система. В процессе эволюции возникла замкнутая внутренняя среда, через которую клетки общаются друг с другом.

Рисунок 6.
Эндокринная регулирующая система. Клетка а выделяет в неё продукты своей жизнедеятельности - гормоны.
Действия первой и второй систем - диффузное. Оно оказалось достаточным лишь до тех пор, пока все наружные клетки были одинаковыми. По мере же специализации клеток возникли и новые регулирующие элементы, обеспечивающие целенаправленное воздействие со стороны внутренних регулирующих клеток на избранные наружные. При этом сохранялся принцип химического воздействия, только оно стало более ограниченным. Так возникла третья регулирующая система – вегетативная нервная система.

Рисунок 7.
Вегетативная нервная система.
У некоторых внутренних клеток вытянулись отростки, один из которых приобрёл особую чувствительность к изменению внутренней среды – стал рецептором, а другой проник в толщу специализированных рабочих клеток, чтобы целенаправленно влиять на их деятельность- превратился в эффектор.
Четвёртая система возникла вместе со специальными органами движения – мышцами. Часть напруженных клеток приобрела особую чувствительность, превратилась в рецепторы, и, соединившись с мышцами, стала управлять последними. Может быть, эту функцию взяли на себя некоторые изменившиеся клетки вегетативной нервной системы. Так или иначе, возникла четвёртая регулирующая система – анимальная, или соматическая, нервная система.

Рисунок 8.
Соматическая нервная система.
4. Анализ медико-биологических методов исследования
На сегодняшний день неврология располагает широким спектром высокоинформативных инструментальных и аппаратных методов исследования нервной системы (КТ, МРТ, УЗДГ, ЭЭГ, ЭНМГ и др.).
Электроэнцефалография (ЭЭГ)
—
1) метод электрофизиологического объективного исследования функцио-нального состояния головного мозга, основанный на графической регистрации его биопотенциалов;
2) раздел электрофизиологии центральной нервной системы, изучающий биоэлектрические процессы, связанные с возникновением активности в структурах мозга, с переходом их от относительного покоя к деятельному, активированному или тормозному состоянию.
Данные ЭЭГ наиболее широко используют для решения следующих основных исследовательских и диагностических задач: установления локализации патологического очага в головном мозге; дифференциального диагноза органических и функциональных заболеваний ц.н.с; изучения механизмов эпилепсии и выявления на ее ранних стадиях эпилептогенного фокуса при отсутствии типичных клин. симптомов заболевания; характеристики реакции активации коры головного мозга, а также ориентировочных реакций с оценкой латентных периодов и длительности периодов последействия раздражителей как с диагностической целью, так и при различных тестах (напр., при отборе контингентов лиц, устойчивых к экстремальным условиям их будущей производственной деятельности); при определении эффективности проводимой терапии (нейротропными, противосудорожными и психотропными средствами); для обоснования физиол. безвредности действия новых анестетиков на ц.н.с. и определения глубины наркоза в процессе оперативного вмешательства; оценки обратимых и необратимых изменений мозга по нарушению биопотенциалов мозга в процессе затухания его функции и с целью констатации смерти мозга
.
В 1913 г. В.В. Правдич-Неминскийвпервые с помощью струнного гальванометра зарегистрировал различные типы колебаний потенциалов обнаженного головного мозга собаки, а также представил их описание и классификацию. В 1928 г. немецкий психиатр Бергер впервые записал биотоки головного мозга человека, используя в качестве отводящих электродов иглы, которые вводил под сухожильный шлем головы в лобной и затылочной областях. Такой способ отведения биотоков мозга с поверхности неповрежденного черепа был вскоре заменен прикладыванием к коже головы пластинок из неполяризующегося материала. Эта модификация отведения биопотенциалов мозга вошла в клин. практику, получив название электроэнцефалографии; регистрируемая при этом кривая колебаний биопотенциалов мозга была названа электроэнцефалограммой (ЭЭГ). Бергер установил регулярность ритмических колебаний биопотенциалов коры головного мозга, дал первую классификацию спонтанных ритмов ЭЭГ человека, описал их особенности в покое и изменения при функциональной пробе (открывании глаз испытуемого во время записи ЭЭГ). Дальнейшее развитие Э. как метода электрофизиологического объективного исследования и внедрение ее в неврологическую, нейрохирургическую и психиатрическую практику связано с прогрессом электроники и разработкой многоканальных высокочувствительных электроэнцефалографов — приборов, предназначенных для регистрации биоэлектрических процессов в структурах мозга (см. ниже).
Отечественная электроэнцефалография как раздел электрофизиологии ц.н.с. развивалась в тесной связи с общей и экспериментальной нейрофизиологией и уже в начале 50-х гг. не ограничивалась лишь описанием формы колебаний биопотенциалов мозга, их частотных и амплитудных характеристик в норме и при патологии. Начальный этап развития клинической электроэнцефалографии характеризовался наряду с накоплением фактического материала об особенностях общих (диффузных) и локальных изменений ЭЭГ при различных органических заболеваниях и функциональных состояниях мозга переходом к изучению условий возникновения тех или иных форм электрической активности, механизмов генерации биотоков мозга, связи их с динамическими процессами возбуждения или торможения в коре головного мозга.
В 60-е гг. широкое развитие получила так наз. функциональная электроэнцефалография, изучающая реактивные изменения ЭЭГ в ответ на сплошную и ритмическую афферентную стимуляцию (световое и звуковое раздражения) с целью выявления очага патол. активности, особенно при опухолевых и сосудистых заболеваниях головного мозга. В это же время были начаты исследования локальных вызванных реакций мозга при кратковременных раздражениях разной модальности и так наз. неспецифических ответов коры. На основании изучения последних было установлено, что характер регистрируемых на ЭЭГ ответных реакций в значительной степени определяется соотношением возбуждений, поступающих в кору полушарий головного мозга по специфической (лемнисковой) и неспецифической (ретикулярная формация мозгового ствола) афферентным системам. Одновременно в ЭЭГ выделилось направление электрофизиологического изучения высшей нервной деятельности человека с описанием сложной и мозаичной нейродинамики, сопутствующей процессу выработки условных реакций человека в норме и при патологии.
В 60—70-е гг. заметно повысился интерес к прямой регистрации электрической активности подкорковых структур головного мозга человека с помощью имплантированных в мозг электродов — электросубкортикографии (ЭСубКоГ), способствующей изучению роли глубинных образований головного мозга в механизмах эмоциональных реакций и др. проявлений психической деятельности человека.
Современный этап развития клинической Э. характеризуется разработкой машинных методов количественной оценки частотно-амплитудных и фазных изменений биопотенциалов мозга в норме и при патологии. Математический анализ ЭЭГ позволяет выявлять ее особенности, скрытые при обычной визуальной оценке. В наст. время путем корреляционного анализа и вычисления спектров когерентности исследуют взаимосвязанность отдельных ритмов в разных областях коры головного мозга в норме и при патологии.
Запись биоэлектрических процессов в структурах мозга производят с помощью электроэнцефалографа. Он состоит из коммутатора отведений, усилителя отводимых биопотенциалов, регистрирующего устройства, устройства калибровки, конструктивно объединенных и заключенных в общий корпус. Неотъемлемыми частями электроэнцефалографа являются также электроды
, световой и звуковой стимуляторы. Существуют разные типы электродов: накладные, приклеивающиеся, игольчатые, кортикографические, имплантируемые (долгосрочные), многоконтактные зонды и др. Для исследования биопотенциалов базальной поверхности головного мозга применяют специальные электроды, подводимые через носовые ходы к задней стенке носоглотки. Наиболее удобными в клин. практике являются накладные электроды-мостики, которые укрепляют на голове с помощью резиновых шлемов-сеток. С целью экспресс-диагностики, напр. при тяжелой черепно-мозговой травме, используют игольчатые электроды. Электроды, применяемые для отведения биопотенциалов мозга с определенных точек головы человека, имеют малое переходное сопротивление (между электродом и кожей головы), а также малое напряжение поляризации. Их, как правило, изготавливают из токопроводящих металлов, обладающих антикоррозионными свойства ми; обычно используют чистое серебро или его смесь с хлоридом серебра. Необходимый контакт при накладывании электродов на кожу головы создает специальная электродная паста. Коммутатор отведений представляет собой многопози-ционный переключатель. В наст. время вместо механических коммутаторов отведений применяют электронные (программные), которые позволяют автоматизировать процесс регистрации биопотенциалов мозга и сократить время записи ЭЭГ. Чувствительность электроэнцефалографов достигает 1—0,2 мкв/мм.
Для обеспечения такой чувствительности в диапазоне частот 0,5—100 гц
существуют различные устройства, подавляющие внешние помехи полезного сигнала. Для количественной оценки амплитудных характеристик ЭЭГ в электроэнцефалографы встраивают устройство калибровки чувствительности, амплитуда напряжения которого находится в диапазоне от 20 до 5000 мкв.
Регистрирующее устройство электроэнцефалографа имеет несколько скоростей движения бумаги, чаще используют скорости 15, 30 и 60 мм/сек.
Эффективная ширина записи не должна превышать 20 мм
.
В электроэнцефалографах применяют перьевой и струйный чернильные виды записи, а также термическую запись на специальной бумажной ленте. В устройство электроэнцефалографа входят частотные фильтры, предназначенные для ограничения полосы ЭЭГ в пределах 15, 30, 75 гц.
По числу каналов записи ЭЭГ электроэнцефалографы бывают 8-, 16- и 32-канальными. Наибольшее применение в медицинской практике получили 8- и 16-канальные приборы, установленные для удобства эксплуатации на передвижных тележках. С помощью дополнительных датчиков, приставок, встраиваемых блоков электроэнцефалографы позволяют регистрировать и другие электрофизиологические параметры — электрокардиограмму, электромиограмму, кривые дыхания и др.
Совершенствование электроэнцефалографов идет по пути автоматизации их управления и внедрения обработки ЭЭГ с помощью микро-ЭВМ и микропроцессоров. Результаты обследования больного при этом получают в цифровом или буквенном виде с заключением по диагнозу.
Для исследования биопотенциалов мозга, кроме электроэнцефалографов, применяют электроэнцефало-скопы — приборы, позволяющие одновременно исследовать биопотенциалы многих областей ц.н.с. На экране электронно-лучевой трубки электроэнцефалоскопа пространственное распределение биопотенциалов можно наблюдать по изменению яркости луча в соответствии со знаком и интенсивностью биоэлектрических потенциалов в каждой точке отведения.
Запись ЭЭГ производят в свето- и звукоизолированном помещении для нивелирования ориентировочных реакций испытуемого на влияние внешних факторов. Современные электроэнцефалографы снабжены специальными фильтрами для подавления сетевых наводок тока, поэтому ранее использовавшиеся для записи ЭЭГ экранированные от электрических помех камеры необязательны.
Обследуемый в зависимости от тяжести состояния во время записи ЭЭГ находится в положении полулежа в специальном удобном кресле или лежит на кушетке с несколько приподнятым подголовником. Перед электроэнцефалографией обследуемого предупреждают о том, что процедура записи ЭЭГ безвредна, безболезненна, продолжается не более 20—25 мин., что надо обязательно закрыть глаза, расслабить мышцы туловища, конечностей, шеи и мышцы лица.
Фиксацию отводящих электродов на голове производят по международной схеме (рис. 9
), в которой представлены основные точки наложения электродов, имеющие определенные обозначения и соответствующие конкретным областям поверхности мозга: лобные — F
3
,
F
4
,
нижнелобные — F
7
,
F8
, лобно-полюсные — Fp
1
F
p
2
, центральные — С3
, С4
,
теменные — Р3
, Р4
, затылочные — О1
, О2
,
передние и задние височные — Т3
, Т4
и T
5
,
T
6
, сагиттальные — Pz
, Cz
, Fz
.
Электроды устанавливают симметрично по отношению к средней линии головы так, чтобы расстояния между соседними электродами с обеих сторон были одинаковыми. Индифферентный электрод, чаще накладываемый на мочку уха, обозначают буквой А. Электроды, расположенные на правой половине головы, обозначаютчетными номерами, на левой — нечетными. В клинической Э. используют схемы отведений с уменьшенным количеством электродов (10—12). Различают два способа отведений: биполярный, при котором запись ЭЭГ осуществляется от двух активных электродов, и монополярный, когда один электрод активный, а второй — индифферентный. Перед установкой электродов кожу на голове обезжиривают (протирают смесью спирта, эфира и ацетона), что обеспечивает нормальную величину переходного сопротивления.
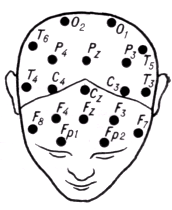 |
Рис. 9. Схема расположения отводящих электродов на коже головы при электроэнцефало
графии:
буквами обозначены точки наложения электродов, соответствующих конкретным областям поверхности мозга, с которых ведется запись биопотенциалов (О — затылочные; Т
— височные;Р
— теменные; С
— центральные; F
—
лобные; Fp
— лобно-полюсные); четными цифрами обозначены точки наложения электродов, расположенные на правой половине головы; нечетными — на левой; точки, расположенные по средней линии (сагиттально), имеют индекс z.
|
В настоящее время признано, что регистрация только ЭЭГ покоя (фоновой, спонтанной ЭЭГ) недостаточно информативна для оценки нечетко выраженных локальных нарушений и определения активности патол. очага, так или иначе влияющего на биопотенциалы соседних и отдаленных областей полушарий головного мозга. С целью выявления патол. активности на ЭЭГ при клинической Э. используют ряд методических приемов. К ним относятся пробы с открыванием и закрыванием глаз во время записи ЭЭГ, а также экстероцептивные раздражения ритмическим или сплошным светом и звуком, проприоцептивные и вестибулярные раздражения. Другими функциональными пробами являются гипервентиляция, темновая адаптация, сон и депривация (лишение) сна. Выбор той или иной функциональной пробы с целью уточнения соотношения очаговых и общемозговых нарушений у больных с локальными поражениями мозга определяется состоянием больного, а также конкретными задачами исследования.
По современным представлениям, ЭЭГ, регистрируемая через неповрежденные покровы головы, является суммарной, усредненной биоэлектрической активностью мозга, обусловленной синхронной активностью множества нейронов. При рассмотрении теоретических вопросов Э. следует различать происхождение электрической активности головного мозга и происхождение ритмики биопотенциалов коры и подкорковых образований.
Наличие постоянной регулярной ритмики на ЭЭГ свидетельствует против предположения о том, что на ЭЭГ регистрируется независимая деятельность нейронных популяций, находящихся вблизи отводящего электрода. Суммарная ЭЭГ представляет собой организованный колебательный процесс, в котором визуально и особенно четко при автоматическом машинном анализе можно выделить регулярные частотные составляющие.
ЭЭГ характеризуется такими основными показателями, как частота, амплитуда и фаза колебаний. Для трактовки физиол. значения тех или иных изменений ЭЭГ в норме и при патологии важно оценивать форму биопотенциалов, их устойчивость, пространственно-временные параметры.
В ЭЭГ выделяют следующие основные физиол. ритмы с диапазонами частот: 1—3 гц
— дельта (Δ); 4— 7гц —
тета (θ); 8—13 гц —
альфа (α); 14—20 гц —
низкий бета (βН
); 20—30 гц
— высокий бета (βв
); свыше 30 гц
— гамма (γ)(рис. 10
). Кроме того, известны регистрируемые в определенных условиях роландический, или мю (μ)-, и каппа (χ)-ритмы, частота которых находится в пределах α-диапазона.

Рис. 10. Различные физиологические ритмы электроэнцефалограмм:
1 — дельта (Δ)-ритм; 2 — тета (θ)-ритм; 3 — альфа (α)-ритм; 4 — бета (β)-ритм; 5 — гамма (γ)-ритм.
Наиболее характерным, основным в ЭЭГ здорового человека является α-ритм, имеющий амплитуду в диапазоне 45—70 мкв,
двухфазный характер и синусоидальную форму. Наибольшую выраженность α-ритм имеет в затылочных, затылочно-височных и затылочно-теменных областях головного мозга в состоянии относительного покоя обследуемого при закрытых глазах и расслаблении мышц. У 75—90% здоровых лиц α-ритм является доминирующим, у остальных здоровых лиц он может иметь уменьшенную амплитуду и не превышать выраженности других частотных компонентов. Открывание глаз во время регистрации ЭЭГ, кратковременное световое раздражение вызывают четкую реакцию десинхронизации α-ритма, которую ранее называли блокадой или депрессией α-ритма. В настоящее время эти изменения α-ритма называют реакцией активации, возникающей в результате нарушения синхронности работы нервных элементов мозга в ответ на воздействие какого-либо внешнего или внутреннего фактора (рис. 11
). Наличие или отсутствие реакции активации, степень ее выраженности во время раздражения и скорость восстановления исходного α-ритма после прекращения действия раздражителя (или при закрывании глаз) используются в диагностических целях, а также в экспертной практике как объективный критерий реактивности ц.н.с.
В норме на ЭЭГ, кроме α-ритма, выявляются и другие частотные компоненты, более частые и более медленные, чем α-ритм.
Бета-ритм имеет частоту 14—30 гци амплитуду в диапазоне 15—20 мкв.
Наиболее выражен этот ритм в центральных и лобных областях полушарий головного мозга. Он отражает активированное состояние подкорковых образований и коры головного мозга. Во время регистрации ЭЭГ при произвольном движении правых или левых конечностей испытуемого или мысленном представлении им этого произвольного движения, особенно при записи с открытой поверхности головного мозга, т.е. при записи электрокортикограммы (ЭКоГ), проявляются изменения амплитуды β-ритма и α-ритма в передних отделах полушарий головного мозга. Причем изменения β-ритма более выражены, чем наступающая при этом десинхронизация α-ритма. При перераздражении афферентных нервов амплитуда β-ритма увеличивается в 2—3 раза, α-ритм дезорганизуется, развивается состояние патологически устойчивой гиперактивации подкорково-корковых связей (ирритация). Такой тип диффузных изменений ЭЭГ наблюдается при гиперкинезах, болевых синдромах, в период обостренияцеребральной гипертензии и др.
 |
Рис. 11. Электроэнцефалограмма в норме при одновременной многоканальной регистрации биопотенциалов:
α-ритм (указан стрелками) преобладает в затылочно-теменных монополярных (О1
, О2
, Р3
, Р4
) и биполярных (О2
РА
; Ot
P3
)отведениях; при функциональной пробе — открывании глаза видна депрессия α-ритма (указана квадратными скобками); после закрывания глаз α-ритм восстанавливается. |
Тета-ритм имеет частоту 4—7 гц,
амплитуда его не превышает амплитуды α-ритма. У здоровых лиц β-ритм проявляется с двух сторон в виде кратковременных эпизодов гл. обр. в передних областях полушарий. Он отражает активность срединно-стволовых образований головного мозга и изменяется по мере возрастного созревания корково-стволовых связей в сторону снижения амплитуды и длительности проявления на ЭЭГ. При поражении глубинных структур мозга амплитуда 0-ритма увеличивается до 400— 500 мкв,
что отражает состояние чрезмерного, стойкого возбуждения образований мозгового ствола и снижения собственно корковой активности.
Дельта-ритм имеет частоту 1—3 гци амплитуду, не превышающую у взрослых амплитуду 6-ритма. Наличие в ЭЭГ волн А-ритма указывает на снижение уровня функциональной активности коры, напр. при дремотном состоянии, утомлении. В норме Л-волны регистрируются кратковременно, диффузно, во всех отведениях; при включении раздражений любой модальности Δ-ритм сменяется α-ритмом. При органических поражениях, напр. при опухолях мозга, А-волны регистрируются локально в зоне коры, окружающей патол. очаг, при раздражении они не исчезают, а замедляются, амплитуда колебаний при этом нарастает. Изменения А-ритма при раздражениях еще более подчеркивают зону локальных патол. изменений на ЭЭГ или ЭКоГ.
В норме в суммарной ЭЭГ у 12 — 15% испытуемых можно выделить так наз. роландический ритм, или γ-ритм. Он имеет частоту 8—12 колебаний в 1 сек. и амплитуду 40—60 мкв.
От α-ритма роландический ритм отличается топографией и формой. Он регистрируется преимущественно в области центральной (роландовой) борозды полушарий головного мозга, имеет форму аркообразных, однофазных колебаний потенциала, которые не изменяются при световых раздражениях и десинхронизируются при мышечной нагрузке (сжатие кисти в кулак и др.).
Каппа-ритм по частотному диапазону (8—12 гц)
совпадает с α-ритмом. Он регистрируется в височных областях головного мозга при умственном напряжении, когда α-ритм в других областях полушарий подавляется. Полагают, что α-, χ- и γ-ритмы относятся к одной категории ритмической деятельности коры, они, по-видимому, сходны по происхождению и связаны с активностью таламических структур.
Гамма-ритмом называют колебания потенциалов коры мозга с частотой более 30 гци амплитудой колебаний8—12 мкв.
Частота γ-ритма варьирует
в диапазоне от 30 до 100 гц.
Закономерные изменения γ-ритмаотмечаются при различных вадахумственной деятельности. Приобычной чернильной записи ЭЭГ выявить не удается, для его обнаружения необходима специальная обработка ЭЭГ.
На ЭЭГ иногда проявляются таксверхмедленные потенциалы,длительность которых измеряется секундами и даже десятками секунд.Сверх медленные потенциалы, по-видимому, играют роль в корреляции активности разных отделов мозга, в их «сонастройке» при выработкеновых временных связей. На обычных электроэнцефалографах, не снабженных усилителями постоянного тока, записать сверхмедленные потенциалы через покровы черепа практически невозможно.
При разных формах функциональной активности мозга в норме, а также при патологических и пограничных состояниях необходима комплексная оценка ЭЭГ. При этом важно учитывать не только выраженность отдельных ритмов, но и соотношения их частотно-амплитудных показателей и пространственного распределения. В настоящее время широко внедряется математический анализ ЭЭГ с использованием ЭВМ, который позволяет проводить частотную оценку суммарной ЭЭГ и выражать мощность каждого ритма в количественных показателях (рис. 12
).

Рис. 12. Спектрограмма нормальной электроэнцефалограммы затылочной (О2
)
и вентральной (С4
) областей:
по оси ординат отложены количественные показатели мощности каждого частотного спектра вдецибелах; по оси абсцисс отложены частоты колебаний, составляющие суммарную электроэнцефалограмму (Д — дельта-ритм;
Т — тета-ритм; А - альфа-ритм; Б — бета-ритм; автоматическая отметка частотного диапазона обозначена восклицательным знаком).
5. Вывод
Невозможно назвать хотя бы одну отрасль экспериментальной, лечебной или профилактической медицины, которая могла бы рассчитывать на даже малый успех без применения электронной медицинской аппаратуры. Инструментальные методы оценки параметров жизнедеятельности организмов человека, животных и других представителей биосферы прочно вошли в повседневную практику специалистов связанных с исследованием биологических объектов – физиологов, биологов, ботаников, медицинских работников и др. В современной медицине и биологии широкое распространение получила измерительная и регистрирующая техника для решения самих различных задач по исследованию организма. При этом используется большой арсенал методов и средств, предназначенных для измерения разных медико-биологических показателей, а также для регистрации и анализа разных физиологических процессов, протекающих в организме. Результаты исследований представляются в виде набора чисел и графиков, отражающих состояние биологического объективно время проведения исследований. Необходимым условием эффективности таких исследований является оснащение лечебных и исследовательских учреждений современными методами лечения и исследования, а также современным оборудованием. Выбор методов в значительной степени зависит от решаемых медицинских и биологических задач. Количество методик, которые можно реализовать только с применением соответствующих технических средств постоянно растёт. Получили дальнейшее развитие методы исследования центральной нервной системы.
6.
Список использованной литературы
1. М.А. Лемези.
Пособие для поступающих в вузы. – Минск: Университетское, 1995.
2. Г.Н. Кассиль.
Внутренняя среда организма. – М.: Наука, 1983.
3. В.М. Ахутин
. Биотехнические системы. - Издательство ленинградского университета, 1981.
4. Анатомия и физиология человека. – К.: Высшая школа.
5. Ю.Г. Антомонов.
Моделирование биологических систем. – К., 1994.
6. А.С. Батуев, А.П. Таиров.
Мозг и организация движений. – Львов, 1979.
|