| 

Уважаемый читатель!
Вы, держите в руках один из учебников нового поколения по биологии для студентов высших учебных заведений, обучающихся по естественнонаучным направлениям и специальностям. Учебник написан известным специалистом в области биологии и прошел сложный и длительный путь конкурсного отбора на Всероссийском конкурсе учебников нового поколения по общим фундаментальным естественнонаучным дисциплинам. Этот конкурс был инициирован. Госкомвузом (в дальнейшем — Минобразованием России) впервые в истории российской высшей школы в связи с реформированием структуры, и содержания программ высшего образования и проведен в 1995-1998 годах на базе Российского университета дружбы, народов.
В конкурсе по одиннадцати номинациям приняли участие свыше трехсот пятидесяти авторских коллективов, чьи разработки более всего соответствовали как новым учебным программам, так и государственным образовательным стандартам по каждой дисциплине.
Конкурсная комиссия выражает надежду, что данный учебник внесет вклад в дело дальнейшего совершенствования российского высшего профессионального образования, и желает всем читателям — студентам и преподавателям — больших творческих успехов.
Первый заместитель Министра
общем и профессиональном образования России,
академик российской академии, образования,
председатель конкурсной комиссии
профессор В. Д. ШАДРИКОВ
ПОСВЯЩАЕТСЯ
ПАМЯТИ МАТЕРИ
ПЕХОВОЙ АННЫ ИЛЬИНИЧНЫ.
ПАРТИЗАНКИ
ВЕЛИКОЙ ОТЕЧЕСТВЕННОЙ ВОЙНЫ.
ОТДАВШЕЙ ЖИЗНЬ ЗА НАШУ СОВЕТСКУЮ РОДИНУ
В 1942 ГОДУ
А. П. ПЕХОВ
БИОЛОГИЯ
С ОСНОВАМИ
ЭКОЛОГИИ
Рекомендовано Министерством образования
Российской Федерации в качестве учебника
для студентов высших учебных заведений,
обучающихся по естественнонаучным
специальностям и направлениям

Санкт-Петербург«
2000
ББК 28
П31
Пехов А. П.
П 31 Биология с основами экологии. Серия «Учебники для вузов. Специальная литература» — СПб.: Издательство «Лань», 2000. — 672 с.
ISBN 5-8114-0219-8
В учебнике освещены основные разделы современной биологии с основами экологии. Он состоит из шести разделов. В разделе I приведены сведения о биоразнообразии, в разделе II — о сущности жизни, свойствах и организации живого, о структуре и свойствах клеток, о росте индивидуальном развитии организмов, в разделе III — о наследственности и изменчивости организмов, о генетическом материале, о действии генов, и о генетике человека, в разделе IV — об эволюции органического мира, включая происхождение человека, в разделе V — об основах экологии, включая экологию человека, в разделе VI — о генетической инженерии, биотехнологии, и об их экологических проблемах, а также о методологических проблемах биологии и экологии. В заключении к учебнику даны краткие обобщения рассмотренных современных данных биологии и экологии.
Учебник предназначен для студентов естественнонаучных, сельскохозяйственных, физкультурных и других направлений бакалавриата. Он также полезен для студентов-медиков, а также студентов, изучающих ветеринарную медицину.
Рисунков — 231. Таблиц — 45.
ББК 28
Рецензенты:
проф. В. А. Голиченков, проф. Ю. Л. Гужов
Оформление обложки С. Л. Шапиро, А. А. Олексенко
Охраняется законом РФ об авторском праве.
Воспроизведение всей книги или любой ее части
запрещается без письменного разрешения издателя.
Любые попытки нарушения закона будут
преследоваться в судебном порядке.
© Издательство «Лань», 2000
© А. П. Пехов, 2000
© Издательство «Лань»,
художественное оформление, 2000
БИОЛОГИЯ — это наука о живом. Долгое время она развивалась как наука описательная, — сейчас ее называют традиционной биологией.
Но с тех пор как в биологию вошли методы физики и химии, ее стали называть новой биологией, или ^'изико-химическои, т. е. молекулярной биологией. В последние десятилетия в этой науке произошли поистине революционные изменения, благодаря чему она выдвинулась на передний план естествознания, начала активно способствовать и, частично, задавать направление научно-техническому прогрессу, идущему вперед семимильными шагами.
Современные данные о живом имеют, прежде всего, гигантское познавательное значение, ибо вносят выдающийся вклад в создание научной картины мира. Однако, непрерывно осуществляя познавательную функцию, биология через генетическую инженерию стремительно вовлеклась в материальное производство, стала одной из производительных сил. С другой стороны, одна из биологических наук, а именно, экология вышла за рамки биологии, стала междисциплинарной наукой. Так произошло благодаря тому, что ученые предпринимали громадные усилия (и продолжают это делать), чтобы общество в целом глубоко осознало тот факт, что деятельность человека в окружающей среде влечет за собой не только положительные, но и отрицательные последствия; последние же могут привести к катастрофе — как к локальной (на отдельно взятой территории), так и к глобальной, мировой. И в наше время невозможно изучать биологию, не уделяя внимания экологии и наоборот. Поэтому данный учебник является учебником по биологии с основами экологии. Можно сказать, что биология и экология — это современный комплекс наук о живом, о происхождении, росте, развитии, наследственности и изменчивости организмов, о взаимоотношениях организмов между собой и со средой, о результатах деятельности человека в окружающей среде и воздействии факторов, порожденных этой деятельностью, на организм человека, животных и растений. Следовательно, задача студентов, которые будут пользоваться этим учебником, заключается в усвоении базовых данных современной биологии и экологии, понимании их фундаментального значения и в использовании приобретенных знаний в практической работе.
Учебник написан на основе примерной программы по дисциплине «Биология с основами экологии» и предназначен для естественнонаучных, сельскохозяйственных, физкультурных и некоторых технических направлений бакалавриата. Однако автору при этом хотелось бы отметить несколько очень важных моментов. Один из них сводится к тому, что, руководствуясь этой программой, мы все же стремились отразить в учебнике свое собственное видение биологии, ее основ и тенденций развития, которое складывалось в результате длительного периода научной и преподавательской деятельности в этой области и развивалось под благотворным влиянием непосредственного общения в те или иные годы с корифеями отечественной науки (такими как Н. П. Дубинин, А. А. Баев, мой учитель Н. Н. Жуков-Вережников, А. А. Им-шенецкий, В. М. Кланов, Ю. А. Овчинников, М. А. Пешков, В. Д. Тимаков и др.). Невозможно не отметить и роль моих зарубежных учителей Г. Понтекорво и У. Хейза (Великобритания), под руководством которых автор изучал генетику и ряд проблем общей биологии, а также благотворность общения с другими выдающимися зарубежными учеными (Р. Картис, Д. Кларк Р. Клаус, США; Ш. Ауэрбах, Н. Датта и М. Ричмонд, Великобритания; Т. Митсухаши, Япония).
Учебник предназначен для студентов разных специальностей, и, учитывая это, автор счел необходимым шире представить не только экологическую проблематику, но и возможности, направления и достижения генетической инженерии, которая позволила получить данные, являющиеся критерием ценности наших представлений о живой материи, и, что не менее важно, подняла на новый уровень биотехнологию.
Наконец, еще один важный момент, которым руководствовался автор, связан с тем, что в свете исключительно быстрого и непрерывного прогресса биологических наук эффективность преподавания курса, изложенного в этом учебнике, полностью зависит не только от новизны и актуальности используемых материалов, но и от уровня методики их преподавания. По этой причине автор стремился подготовить учебник, который был бы насыщен новейшими данными и который оставался бы полезным для студентов как можно дольше.
Чтобы студенты могли более подробно познакомиться с интересующими их проблемами, в конце каждой главы приводится список дополнительной литературы, куда, помимо отечественных источников, вошли и новейшие издания на английском языке. Этот язык играет ведущую роль в общении ученых разных стран, и, не ознакомившись с работами зарубежных специалистов, весьма трудно получить полное представление о современном состоянии науки.
Наконец, в каком бы направлении ни развивалась биология, а вместе с ней и экология, научные открытия всегда используются во имя человека и для пользы человека. По этой основополагающей причине мы стремились в этом учебнике, по возможности, быть «близко» к человеку, к его биологии и здоровью.
Со времен Г. Гегеля (1770-1831) известно, что качество всех вещей и явлений определяется мерой. Непрестанно думая об этом, мы учитывали и то, в какой мере этот учебник будет доступен для тех, кому он может быть полезен. Надеемся, что в общем нам удалось изложить столь сложный материал в приемлемой форме. В этом нас убеждает наш опыт преподавания биологии в Российском университете дружбы народов, а также опыт чтения лекций по биологии в ряде зарубежных университетов, в частности, в университетах штатов Алабамы и Аризоны (США), в Хартумском университете (Судан) и университете г. Дакки (Бангладеш), где мы имели возможность работать некоторое время. Именно материалы прочитанных там лекций в значительной мере составили основу данного учебника.
Как всегда, благодарю своих сотрудников (особенно проф. В. П. Щипкову) за то, что они помогли мне найти время написать этот учебник. Я также очень признателен специалистам, ознакомившимся с учебником в рукописи и высказавшим свои замечания, которые оказались чрезвычайно полезными.
Независимо от специальности студентов, в учебном плане курс «Биология с основами экологии» должен быть представлен лекциями и лабораторными занятиями. При этом общие установочные вопросы должны быть вынесены на лекции, тогда как углубленное изучение этих вопросов и частных закономерностей должно проводиться на лабораторных занятиях (путем постановки соответствующих экспериментов) или на семинарских занятиях в процессе обсуждения. Желательно, чтобы изучение этого курса было согласовано по времени с изучением курсов физики и химии, поскольку многие современные биологические понятия сложились на основе идей, методов та. данных этих наук.
В любом вузе по любой научной дисциплине значительная часть времени в подготовке студентов уделяется их самостоятельной работе, основной формой которой является чтение учебника и дополнительной литературы, ее анализ, а также самоконтроль (поиск ответов на вопросы, возникающие в процессе самостоятельной подготовки, лекционных, лабораторных и семинарских занятий).
* * *
Учебник состоит из 5 разделов, которые отражают основную проблематику биологии и экологии, причем материалы по экологии излагаются с учетом того, что экология является, прежде всего, биологической наукой, которая в наше время стала наукой междисциплинарной. В свою очередь, каждый из разделов состоит из нескольких глав, которые разделены на параграфы. Как правило, в каждой главе излагаются данные по одной проблеме.
Крайне необходимо, чтобы студенты очень' хорошо усвоили определения биологических понятий. В тексте учебника, по мере введения новой терминологии, даются определения тех или иных явлений. Тем не менее, в Приложение включен словарь наиболее важных биологических терминов, которые должны быть усвоены студентами «окончательно и бесповоротно». Очень полезно знать, как развивались биологические понятия во времени. Поэтому в Приложении есть перечень наиболее важных дат в развитии биологии и приведены имена ученых, которые своими открытиями обеспечили это развитие.
Особое внимание уделяется углубленному изучению разделов биологии, пониманию перспектив их дальнейшего развития. По этой причине в конце каждой главы приводится список литературы, рекомендуемой для дополнительного чтения. Как говорилось выше, в перечень книг вошли не только отечественные учебные и научные издания, но и работы ученых (наиболее значительные монографии и фундаментальные обзорные статьи), опубликованные за рубежом на английском языке. Они будут полезны не только студентам, особенно на семинарских занятиях и в научных кружках, но и преподавателям в объяснениях студентам дальнейших перспектив биологического познания.
Вопросы, необходимые для самоконтроля, приведены в конце каждой главы. Эти вопросы — «авторские» и сформулированы так, чтобы они наиболее полно отражали материалы учебника и соответствовали учебной программе (какой она представляется автору). Если студент сумеет ответить на них, то это означает, что он усвоил материал главы. Но студенты, равно как и их преподаватели, могут поставить себе и «свои» вопросы.
Также в конце каждой главы, среди вопросов для самоконтроля, приведены задачи, имеющие аналогичное функциональное значение. Но успешное решение задач особо ценно в том плане, что оно позволяет практически использовать усвоенные фундаментальные данные и приблизиться к моделированию изучаемых процессов.
Чтобы облегчить работу с учебником, в Приложение включены также предметный и именной (авторский) указатели. Предметный указатель содержит все основные термины, используемые в учебнике; авторский указатель содержит фамилии ученых, встречающиеся в тексте.
Наконец, в Приложении приведен перечень единиц измерений, используемых в биологических исследованиях, а также названия наиболее важных отечественных и зарубежных научных журналов и других периодических изданий, в которых публикуются экспериментальные и обзорные статьи по биологии и экологии.
Александр ПЕХОВ
профессор,
доктор биологических наук,
заслуженный деятель науки
Российской Федерации,
Академик
Международной Академии Наук
Высшей Школы Российский университет.
дружбы народов
«Я расскажу вам откровенно,
как было дело,
не прибавив ничего от себя,
что стоит историку немалого труда».
Ф.М.Вольтер. 1757
Биология (от греч. bios — жизнь, logos — наука) — наука о жизни, об общих закономерностях существования и развития живых существ. Предметом ее изучения являются живые организмы, их строение, функции, развитие, взаимоотношения со средой и происхождение. Подобно физике и химии она относится к естественным наукам, предметом изучения которых является природа.
Биология — одна из старейших естественных наук, хотя термин «биология» для ее обозначения впервые был предложен лишь в 1797 г. немецким профессором анатомии Теодором Рузом (1771-1803), после чего этот термин использовали в 1800 г. профессор Дерптского университета (ныне г. Тарту) К. Бурдах (1776-1847), а в 1802 г. Ж.-Б. Ламарк (1744-1829) и Л. Тревиранус (1779-1864).
Биология — естественная наука. Как и другие науки, она возникла и всегда развивалась в связи с желанием человека познать окружающий его мир, а также в связи с материальными условиями жизни общества, развитием общественного производства, медицины, практическими потребностями людей.
Этапы развития биологии. Самые первые сведения о живых существах человек стал собирать, вероятно, с тех пор, когда он осознал свое отличие от окружающего мира. Уже в литературных памятниках египтян, вавилонян, индийцев и др. содержатся сведения о строении многих растений и животных, о применении этих знаний в медицине и сельском хозяйстве. В XIV в. до н. э. многие клинописные таблички, созданные в Месопотамии, содержали сведения о животных и растениях, о систематизации животных путем разделения их на плотоядных и травоядных, а растений на деревья, овощи, лекарственные травы и т. д. В медицинских сочинениях, созданных в VI—I вв. до н. э. в Индии, содержатся представления о наследственности как причине сходства родителей и детей, а в памятниках «Махабхарата» и «Рамаяна» дано довольно подробное описание ряда особенностей жизни многих животных и растений.
В период рабовладельческого строя возникают ионийская, афинская, александрийская и римская школы в изучении животных и растений.
Ионийская школа возникла в Ионии (VII-IV вв. до н. э.). Не веря в сверхъестественное происхождение жизни, философы этой школы признавали причинность явлений, движение жизни по определенному пути, доступность для изучения «естественного закона» , который, по их утверждению, управляет миром. В частности, Алкмеон (конец VI—начало V в. до н. э.) описал зрительный нерв и развитие куриного эмбриона, признавал мозг в качестве центра ощущений и мышления, а Гиппократ (460-377 гг. до н. э.) дал первое относительно подробное описание строения человека и животных, указал на роль среды и наследственности в возникновении болезней.
Афинская школа сложилась в Афинах. Наиболее выдающийся представитель этой школы Аристотель (384—322 гг. до н. э.) создал четыре биологических трактата, в которых содержались разносторонние сведения о животных. Аристотель подразделял окружающий мир на четыре царства (неодушевленный мир земли, воды и воздуха, мир растений, мир животных и мир человека), между которыми устанавливалась последовательность. В дальнейшем эта последовательность превратилась в «лестницу существ» (XVIII в.). Аристотелю принадлежит, вероятно, и самая первая классификация животных, которых он классифицировал на четвероногих, летающих, пернатых и рыб. Китообразных он объединил с сухопутными животными, но не с рыбами, которых он классифицировал на костных и хрящевых.
Аристотелю были известны основные признаки млекопитающих. Он дал описание наружных и внутренних органов человека, половых различий у животных, способов размножения и образа жизни животных, происхождения пола, наследования отдельных признаков, уродств, многоплодия и т. д. Аристотеля считают основоположником зоологии. Другой представитель этой школы Те-офраст (372-287 гг. до н.э.) оставил сведения о строении и размножении многих растений, о различиях между однодольными и двудольными растениями, ввел в употребление термины — плод, околоплодник, сердцевина. Его считают основоположником ботаники.
Александрийская школа вошла в историю биологии благодаря ученым, занимавшимся в основном изучением анатомии. Герофил (расцвет творчества на 300 годы до н. э.) оставил сведения по сравнительной анатомии человека и животных, впервые указал на различия между артериями и венами, а Эразистрат (250 годы до н. э.) описал полушария головного мозга, мозжечок, извилины головного мозга.
Римская школа не дала самостоятельных разработок в изучении живых организмов, ограничившись коллекционированием сведений, добытых греками. Гай Плиний старший (23-79 гг.) создал энциклопедию «Естественная история» из 37 томов, в которой содержались также и сведения о животных и растениях. Диоско-рид (I век н. э.) оставил описание 600 видов растений, обращая внимание на их целебные свойства. Клавдий Гален (130-200 гг.) широко проводил вскрытия млекопитающих (крупный и мелкий рогатый скот, свиньи, собаки, медведи и др.), первым дал сравнительно-анатомическое описание человека и обезьяны. Он был последним великим биологом древности, оказавшим исключительно большое влияние на дальнейшее развитие анатомии и физиологии.
В средние века господствующей идеологией была религия. Однако научные знания как-то все же продолжали развиваться. Можно сказать, что новых знаний почти не получали. Но биологические знания, основанные на описаниях Аристотеля, Плиния, Галена, поддерживались. В частности знания, добытые греками, были отражены в энциклопедии Альберта Великого (1206—1280).
На Руси сведения о животных и растениях были обобщены в том древнем произведении, которое известно под названием «Поучение Владимира Мономаха» (XI в.).
Выдающийся ученый и мыслитель средних веков Абу-Али ибн Сина (980-1037), известный в Европе под именем Авиценны, развивал взгляды о вечности и несотворенности мира, признавал причинные закономерности в природе. В этот период биология еще не выделилась в самостоятельную науку, не отделилась от искаженных религиозно-философских взглядов на окружающий мир-
Как считают историки науки, начала биологии, как и всего естествознания, связаны с эпохой Возрождения (Ренессанса). В эту эпоху происходит крушение феодального общества, разрушается диктатура церкви. Можно сказать, что естествознание более быстро начинает развиваться со второй половины XV в. С того времени успехи естествознания следуют один за другим. Например, выдающийся деятель эпохи Возрождения Леонардо да Винчи (1452-1519) в то время открыл гомологию органов, охарактеризовал многие растения, описал поведение птиц в полете, открыл щитовидную железу, описал способ соединения костей суставами, деятельность сердца и зрительной функции: глаза, отметил сходство костей человека и животных, Андреас Везалий (1514-1564) создал анатомический труд «Семь книг о строении человеческого тела», заложивший основы научной анатомии, В. Гарвей (1578—1657) открыл кровообращение, а Д. Борелли (1608—1679) описал механизмы движения животных, что заложило научные основы физиологии. С того времени анатомия и физиология развивались вместе в течение многих десятков лет, после чего они разделились на самостоятельные науки, в пределах которых возникли более узкие науки (анатомия животных, анатомия человека, физиология животных и т. д.).
Чрезвычайно быстрое накопление научных данных о живых организмах вело к дифференцировке биологических знаний, к разделению биологии на отдельные науки по объектам и задачам изучения. В XVI-XVII вв. стала стремительно развиваться ботаника. С изобретением микроскопа (начало XVII в.) в пределах ботаники возникла микроскопическая анатомия растений, закладываются основы физиологии растений. С XVI в. стала быстро развиваться и зоология.
Большое влияние на развитие зоологии в последующем оказала система классификации животных, созданная К. Линнеем (1707-1778). Введя четырехчленные таксономические подразделения (класс — отряд — род — вид), К. Линней классифицировал животных на шесть классов (млекопитающие, птицы, амфибии, рыбы, насекомые, черви).
Значительное влияние на биологию XVII-XVIII вв. оказал и немецкий ученый Г. Лейбниц (1646-1716) и швейцарский ученый Ш. Бонна, которые разработали учение о «лестнице существ», основные принципы которой были заимствованы из взглядов античного мира.
В XVIII-XIX вв. трудами К. Ф. Вольфа, К. М. Бэра и других закладываются основы эмбриологии. С этого времени эмбриология развивается в качестве самостоятельной науки. В 1839 г. Т. Шванн (1804-1881) и М. Шлейден (1810-1882) формулируют клеточную теорию, явившуюся важнейшим обобщением знаний о клетке, ставших известными к концу первой трети XIX в.
В 1859 г. Ч. Дарвин (1809-1882) публикует «Происхождение видов». В этом труде была сформулирована теория эволюции.
В первой половине XIX в. возникает бактериология, которая благодаря трудам Л. Пастера, Р. Коха, Д. Листера и И. И. Мечникова в последующем перерастает в микробиологию как самостоятельную науку. К концу XIX в. в качестве самостоятельных наук оформляются паразитология и экология.
В 1865 г. была опубликована работа Г. Менделя (1822-1884) «Опыт над растительными гибридами», в которой было обосновано существование генов и сформулированы закономерности, которые в настоящее время называют законами наследственности. После повторного открытия этих законов в XX в. оформляется в качестве самостоятельной науки генетика.
Еще в первой половине XIX в. возникли идеи об использовании физики и химии для изучения явлений жизни (Г. Деви, Ю. Либих). Реализация этой идеи привела к тому, что в середине XIX в. физиология обособилась от анатомии, причем физико-химическое направление заняло в ней ведущее место. На рубеже XIX-XX вв. сформировалась современная биологическая химия. В первой половине XX в. оформляется в качестве самостоятельной науки биологическая физика.
Важнейшим рубежом в развитии биологии в XX в. стали 40-50-е годы, когда в биологию хлынули идеи и методы физики и химии, а в качестве объектов стали использовать микроорганизмы. В 1944 г. была открыта генетическая роль ДНК, в 1953 г. выяснена ее структура, а в 1961 г. был расшифрован генетический код. С открытием генетической роли ДНК и механизмов синтеза белков из генетики и биохимии произошло вычленение молекулярной биологии и молекулярной генетики, которые в совокупности часто называют физико-химической биологией. Основным предметом изучения молекулярной биологии и генетики стали структура и функции нуклеиновых кислот и белков. Возникновение этих наук означало гигантский шаг в изучении явлений жизни на молекулярном уровне живой материи.
12 апреля 1961 г. впервые в истории человек поднялся в космос. Этим первым космонавтом был гражданин СССР Юрий Алексеевич Гагарин. У нас этот день стал Днем космонавтики, а в мире — Всемирным днем авиации и космонавтики. Но можно сказать, что этот день является также и днем космической биологии, родиной которой по праву является наша страна.
В наше время биология характеризуется исключительно широким перечнем разрабатываемых фундаментальных проблем, начиная с исследований элементарных клеточных структур и реакций, протекающих в клетках, и заканчивая познанием процессов, развернутых и развивающихся на глобальном (биосферном) уровне. В относительно короткие исторические сроки были разработаны принципиально новые методы исследований, вскрыты молекулярные основы строения и активности клеток, установлена генетическая роль нуклеиновых кислот, расшифрован генетический код и сформулирована теория генетической информации, определены (секвеннированы) последовательности азотных оснований многих генов, появились новые обоснования теории эволюции, возникли новые биологические науки. Новейший революционный этап в развитии биологии — это создание методологии генетической инженерии, которая открыла принципиально новые возможности для проникновения в глубь биологических процессов с целью дальнейшей характеристики живой материи и создания научной картины мира. Генетическая инженерия подняла также на новый уровень биотехнологию, сделала ее более эффективной и привлекла к ней значительное общественное внимание, заставив людей более внимательно задуматься о своем бытие. Появление генетической инженерии привело к созданию ряда совершенно новых социальных и этических проблем естественных наук.
Классификация биологических наук. Биология — это комплексная наука, ставшая в наше время такой в результате дифференциации и интеграции разных биологических наук. Самыми старыми биологическими науками являются зоология и ботаника, изучающие животных и растения соответственно.
Процесс дифференциации биологических наук возник давно и начался с разделения зоологии, ботаники и микробиологии на ряд самостоятельных наук. В пределах зоологии в XVIII—XIX вв. в разное время возникли зоология позвоночных и беспозвоночных, а также паразитология, протозоология, гельминтология, энтомология, малакология, ихтиология, герпетология, орнитология, тери-ология, предметом изучения которых являются паразиты и паразитизм, простейшие, гельминты (черви), насекомые, моллюски, рыбы, земноводные и рептилии, птицы (соответственно) и другие науки. В ботанике в самостоятельные науки выделились дендрология (наука о деревьях и кустарниках), птеридология (наука о папоротниках), альгология (наука о водорослях), бриология (наука о мхах), биогеоботаника (наука о распространении растений) и другие науки.
Отдельные биологические науки имеют комплексное значение. Например, комплексной наукой стала генетика, предметом изучения которой являются наследственность и изменчивость организмов.
В наше время комплексной наукой стала экология, изучающая взаимоотношения организмов между собой и со средой.
Как в зоологии, так и в ботанике уже давно в самостоятельные науки выделились систематика, анатомия, физиология, цитология, гистология, эмбриология и другие дисциплины. Микробиология разделилась на бактериологию, вирусологию и иммунологию. Одновременно с дифференциацией шел процесс возникновения и оформления новых наук, которые расчленялись на более узкие науки. Например, генетика, возникнув в качестве самостоятельной науки, разделилась на общую и молекулярную, на генетику растений, животных и микроорганизмов. В то же время возникли генетика пола, генетика поведения, популяцион-ная генетика, эволюционная генетика и т. д. В недрах физиологии возникли сравнительная и эволюционная физиология, эндокринология и другие физиологические науки. В последние годы отмечается тенденция оформления узких наук, получающих название по проблеме (объекту) исследования. Такими науками являются энзимология, мембранология, кариология, плазмидология и другие.
В результате интеграции наук возникли биохимия, биофизика, радиобиология, цитогенетика, космическая биология и другие науки.
Ведущее положение в современном комплексе биологических наук занимает физико-химическая биология, новейшие данные которой вносят существенный вклад в представления о научной картине мира, в дальнейшее обоснование материального единства мира. Продолжая отражать живой мир и человека как часть этого мира, глубоко развивая познавательные идеи и совершенствуясь в качестве теоретической основы медицины и сельского хозяйства, биология приобрела исключительно большое значение в научно-техническом прогрессе, стала производительной силой.
Методы исследований. Как известно, новые теоретические представления и продвижение познания вперед в любой науке всегда определялись и определяются созданием и использованием новых методов исследования. Биология не является исключением из этого правила.
Основными методами, используемыми в биологических науках, являются описательный, сравнительный, исторический и экспериментальный.
Описательный метод является самым старым методом и основан на наблюдении организмов. Он заключается в сборе фактического материала и описании его. Возникнув в самом начале биологического познания, этот метод долгое время оставался единственным в изучении строения и свойств клеток, тканей и организмов. Поэтому старая (традиционная) биология была связана с простым отражением живого мира в виде описания растений и животных, т. е. она являлась, по существу, описательной наукой. Использование этого метода позволило заложить основы биологических знаний. Достаточно вспомнить насколько успешным оказался этот метод в систематике и в создании науки о систематике организмов.
Описательный метод широко используется и в наше время, особенно в зоологии, ботанике, цитологии, экологии и других науках. Изучение клеток с помощью светового или электронного микроскопа и описание выявленных при этом микроскопических или субмикроскопических особенностей в их строении представляет собой один из теперешних примеров использования описательного метода.
Сравнительный метод заключается в сравнении изучаемых организмов, их структур и функций между собой с целью выявления сходств и различий. Этот метод утвердился в биологии в XVIII в. и оказался очень плодотворным в решении многих крупнейших проблем. С помощью этого метода и в сочетании с описательным методом были получены сведения, позволившие в XVIII в. Заложить основы систематики растений и животных (К. Линней), а также сформулировать клеточную теорию (М. Шлейден и Т. Шванн) и учение об основных типах развития (К. Бэр). Метод широко был использован в XIX в. в обосновании теории эволюции, а также в перестройке ряда биологических наук на основе этой теории. Однако использование этого метода не сопровождалось выходом биологии за пределы описательной науки.
Сравнительный метод широко используют в разных биологических науках и в наше время.
Сравнение приобретает особую ценность тогда, когда невозможно дать определение понятия. Например, с помощью электронного микроскопа часто получают изображения, истинное содержание которых заранее неизвестно. Только сравнение их со светомикроскопическими изображениями позволяет получить желаемые данные.
Во второй половине XIX в. благодаря Ч. Дарвину в биологию входит исторический метод, который позволил поставить на научные основы исследование закономерностей появления и развития организмов, становления структуры и функций организмов во времени и в пространстве. С введением этого метода в биологии немедленно произошли значительные качественные изменения. Исторический метод превратил биологию из науки чисто описательной в науку, объясняющую, как произошли и как функционируют многообразные живые системы. Благодаря этому методу биология поднялась сразу на несколько ступеней выше. В настоящее время исторический метод вышел, по существу, за рамки метода исследования. Он стал всеобщим подходом к изучению явлений жизни во всех биологических науках.
Экспериментальный метод заключается в активном изучении того или иного явления путем эксперимента. Нельзя не отметить, что вопрос об опытном изучении природы, как новом принципе естественнонаучного познания, т. е. вопрос об эксперименте, как одной из основ в познании природы, был поставлен еще в XVII в. английским философом Ф. Бэконом (1561-1626). Его введение в биологию связано с работами В. Гарвея в XVII в. по изучению кровообращения. Однако экспериментальный метод широко вошел в биологию лишь в начале XIX в., причем через физиологию, в которой стали использовать большое количество инструментальных методик, позволявших регистрировать и количественно характеризовать приуроченность функций к структуре. Благодаря трудам Ф. Мажанди (1783-1855), Г. Гельмгольца (1821-1894), И. М. Сеченова (1829-1905), а также классиков эксперимента К. Вернара (1813-1878) и И. П. Павлова (1849-1936) физиология, вероятно, первой из биологических наук стала экспериментальной наукой.
Другим направлением, по которому в биологию вошел экспериментальный метод, оказалось изучение наследственности и изменчивости организмов. Здесь главнейшая заслуга принадлежит' Г. Менделю, который в отличие от своих предшественников использовал эксперимент не только для получения данных об изучаемых явлениях, но и для проверки гипотезы, формулируемой на основе получаемых данных. Работа Г. Менделя явилась классическим образцом методологии экспериментальной науки.
В обосновании экспериментального метода важное значение имели работы, выполненные в микробиологии Л. Пастером (1822-1895), который впервые ввел эксперимент для изучения брожения и опровержения теории самопроизвольного зарождения микроорганизмов, а затем для разработки вакцинации против инфекционных болезней. Во второй половине XIX в. вслед за Л. Пастером значительный вклад в разработку и обоснование экспериментального метода в микробиологии внесли Р. Кох (1843-1910), Д. Листер (1827-1912), И. И. Мечников (1845-1916), Д. И. Ивановский (1864-1920), С. Н. Виноградский (1856-1953), М. Бейеринк (1851-1931) и другие. В XIX в. биология обогатилась также созданием методических основ моделирования, которое является высшей формой эксперимента. Созданные Л. Пастером, Р. Кохом и другими микробиологами способы заражения лабораторных животных патогенными микроорганизмами и изучение на них патогенеза инфекционных болезней — это классический пример моделирования, перешедшего в XX в. и очень дополненного в наше время моделированием не только разных болезней, но и различных жизненных процессов, включая происхождение жизни.
Начиная примерно с 40-х годов XX в. экспериментальный метод в биологии подвергся значительному усовершенствованию за счет повышения разрешающей способности многих биологических методик и разработки новых экспериментальных приемов. Например, была очень повышена разрешающая способность генетического анализа, ряда иммунологических методик. В практику исследований были введены культивированные соматические клетки, выделение биохимических мутантов микроорганизмов и соматических клеток и т. д. Экспериментальный метод стал широко обогащаться методами физики и химии, которые оказались исключительно ценными не только в качестве самостоятельных методов, но и в сочетаниях с биологическими методами. Например, структура и генетическая роль ДНК были выяснены в результате сочетанного использования химических методов выделения ДНК, химических и физических методов определения ее первичной и вторичной структуры и биологических методов (трансформации и генетического анализа бактерий), доказательства ее роли как генетического материала.
В настоящее время экспериментальный метод характеризуется исключительными возможностями в изучении явлений жизни. Эти возможности определяются использованием микроскопии разных видов, включая электронную с техникой ультратонких срезов, биохимических методов, высокоразрешающего генетического анализа, иммунологических методов, разнообразных методов культивирования и прижизненного наблюдения в культурах клеток, тканей и органов, маркировки эмбрионов, техники оплодотворения в пробирке, метода меченых атомов, рентгене структурного анализа, ультрацентрифугирования, спектрофотометрии, хроматографии, электрофореза, секвенирования, конструкции биологически активных рекомбинантных молекул ДНК и т. д. Новое качество, заложенное в экспериментальном методе, вызвало качественные изменения и в моделировании. Наряду с моделированием на уровне организмов в настоящее время очень развивается моделирование на молекулярном и клеточном уровнях, а также математическое моделирование различных биологических процессов.
Успехи, достигнутые в результате использования экспериментального метода, сопровождались изменениями в подходах к изучению явлений жизни. Новое, заложенное в экспериментальном методе и его техническом оснащении, определило и важные подходы к изучению явлений жизни. Продвижение вперед биологических наук в XX в. во многом определилось также возникновением и развитием системно-структурного подхода к изучению организации и функций живых организмов, анализом и синтезом получаемых данных о структуре и функциях исследуемых объектов. Экспериментальный метод в современном оснащении и в сочетании с системно-структурным подходом в корне преобразил биологию, углубил ее познавательные возможности, расширил представления о научной картине мира, еще больше связал ее с производством, с медициной.
Применение биологических знаний. Прежде всего биологические знания имеют познавательное значение. Однако чрезвычайно велико и их практическое значение. Впервые практика стала формулировать свои заказы биологии с введением в эту науку экспериментального метода. Но тогда биология оказывала влияние на практику опосредованно, в частности, через медицину и сельское хозяйство.
Прямое влияние биологии на материальное производство началось с создания основ биотехнологии в тех областях промышленности, которые основываются на биосинтезирующей деятельности микроорганизмов. На основе биологических знаний уже давно в промышленных условиях осуществляется микробиологический синтез многих органических кислот, которые широко используются в народном хозяйстве и медицине.
В 40—50-е годы было создано промышленное производство антибиотиков, а в начале 60-х годов — производство аминокислот. Важное место в микробиологической промышленности сейчас занимает производство ферментов. Микробиологическая промышленность производит сейчас в больших количествах витамины и другие вещества. Как аминокислоты и антибиотики, так и витамины крайне необходимы в народном хозяйстве и медицине. На основе трансформирующей способности микроорганизмов основано промышленное производство веществ с фармакологическими свойствами из стероидного сырья растительного происхождения.
Наибольшие успехи в производстве различных веществ, в том числе лекарственных (инсулин, соматостатин, интерферон и др.), связаны с генетической инженерией, составляющей сейчас основу биотехнологии.
Исключительно важное значение биология имеет для сельскохозяйственного производства. Например, теоретической основой селекции растений и животных является генетика. В последние годы в сельскохозяйственное производство также вошла генетическая инженерия. Она открыла новые перспективы в увеличении производства пищи.
Генетическая инженерия оказывает существенное влияние на поиск новых источников энергии, новых путей сохранения окружающей среды, очистки ее от различных загрязнений.
Развитие биотехнологии, теоретическую основу которой составляет биология, а методическую — генетическая инженерия, является новым этапом в развитии материального производства. Появление этой технологии есть один из моментов новейшей революции в производительных силах.
Биологическое познание прямым образом связано с медициной, причем эти связи уходят в далекое прошлое и датируются тем же временем, что и возникновение самой биологии. Больше того, многие выдающиеся медики далекого прошлого были одновременно и выдающимися биологами (Гиппократ, Герофил, Эразистрат, Гален, Авиценна, Мальпиги и другие). Тогда и позднее биология стала обслуживать медицину путем «поставки» ей сведений о строении организмов. Однако роль биологии, как теоретической основы медицины в современном понимании, стала формироваться лишь в прошлом веке.
Создание в XIX в. клеточной теории заложило подлинно научные основы связи биологии с медициной. В 1858 году немецкий ученый Р. Вирхов (1821-1902) опубликовал книгу <Щеллю-лярная патология», в которой было сформулировано положение о связи цитологического процесса с клетками, с изменениями в строении клеток. Соединив клеточную теорию с патологией, Р. Вирхов прямым образом «подвел» биологию под медицину в качестве теоретической основы.
Значительные заслуги в укреплении связей биологии и медицины в XIX в. и начале XX в. принадлежат К. Бернару и И. П. Павлову, которые раскрыли и общебиологические основы физиологии и патологии, Л. Пастеру, Р. Коху, Д. И. Ивановскому и их последователям, создавшим учение об инфекционной патологии. Исследуя процессы пищеварения у низших многоклеточных животных, И. И. Мечников заложил биологические основы учения об иммунитете, имеющего большое значение в медицине. На основе микробиологии, иммунологии и паразитологии разрабатываются вопросы диагностики и профилактики инфекционных и паразитарных болезней, развивается эпидемиология.
В укреплении связей биологии с производством и медициной существенный вклад принадлежит генетике, данные которой имеют важнейшее значение в разработке основ диагностики, лечения и профилактики наследственных болезней.

«На протяжении многих тысячелетий
человек стремился
познать окружающий его мир и то место,
которое он занимает в этом мире,
на маленькой Земле или в большой Вселенной».
1977
Совокупность растений называют флорой (новолат. flora, от лат. Flora — богиня цветов и весны), тогда как совокупность животных — фауной (новолат. fauna, от лат. Fauna — богиня лесов и полей, покровительница стад животных).
На Земле нет места, где бы ни обитали растения и животные. Можно сказать, что жизнь на Земле вездесуща.
Между растениями и животными существует ряд сходств (одинаковое клеточное строения, одинаковый генетический материал и т. д.). В то же время для них характерны существенные различия (наличие целлюлозы в клеточных стенках растений, которой нет в мембранной системе клеток животных, присущий многим растениям неограниченный рост, который не характерен для животных и т. д.). Между растениями и животными существуют различия и по численному составу.
Растения и животные создают живую оболочку Земли, и в этом заключается их планетарная роль. Для человека они являются прежде всего источником пищи. Они обеспечивают многие отрасли промышленности и хозяйственную деятельность человека сырьем и материалами (древесина, хлопок, шерсть и др.), являются лекарственным сырьем для медицинской промышленности, а также обладают многими другими ценными для человека свойствами.
Как полагают многие ученые, растения и животные начали раздельное развитие от общего предка свыше 1 млрд лет назад.
А. И. ОПАРИН,
На Земле идентифицировано около 2 000 000 видов животных и растений, включая виды, которые жили в далеком прошлом, но затем вымерли. На долю растений приходится 500 000 видов, а на долю животных около 1 500 000 видов. Непрерывно открываются также новые виды растений и животных. Чтобы изучать это огромное множество животных и растений, их необходимо вначале классифицировать на сходные группы или категории. Задачу классификации организмов решает наука, которую называют систематикой и предметом которой является изучение и описание особенностей этого бесконечного множества форм.
Кроме того, в задачу систематики входит нахождение названий единиц классификации (таксонов), а также изучение эволюционных взаимоотношений между всеми единицами классификации. Разделами систематики являются таксономия, название которой еще в 1813 г. предложил швейцарский ученый О. Декандолль (1778— 1841) и которая представляет собой теорию и практику классификации, номенклатура (совокупность названий таксонов) и филоге-нетика (установление родства между организмами в историческом плане).
Самые первые попытки классификации организмов принадлежат Аристотелю (384-322 гг. до н. э.), который считал, что общее количество видов растений и животных составляет всего лишь несколько сотен. Аристотель и его ученик Теофраст (370-285 гг. до н. э.) подразделяли растения на травы, кустарники и деревья, а животных на ряд групп в зависимости от того, где они живут — водные, земные, воздушные. Последователи этих выдающихся греков использовали далее для классификации организмов такие признаки, как их полезность, вредность или безвредность.
Названные системы классификации были эмпирическими или, как их еще называют, искусственными системами, ибо они не основывались на признании единства естественного происхождения всех живых существ и не отражали естественных связей между разными организмами. Тем не менее даже в средние века в Европе продолжали называть животных и растения именами, данными им еще древними греками и римлянами. Однако это не приводило к успеху, т. к. разные европейские растения и животные характеризовались различиями на разных территориях европейского континента.
Искусственные системы в классификации организмов условно используют и сейчас, когда характеризуют организмы, учитывая их хозяйственные признаки. Например, растения подразделяют на культурные и дикорастущие, съедобные и ядовитые, лекарственные и кормовые и т. д. Животных подразделяют на домашних и диких, на вредителей полей, садов и огородов, на паразитов человека и животных, на переносчиков возбудителей болезней человека и животных и т. д.
Такие подразделения не лишены значения в хозяйственной и другой деятельности человека, однако для научной классификации организмов они не имеют ценности.
Уже давно выявилась тенденция выделять с целью классификации естественные системы, которые бы отражали естественные связи между организмами. Чрезвычайно важным шагом на пути к научной классификации организмов оказались создание в 1663 г. английским естествоиспытателем Д. Реем (1627-1705) концепции вида. Он считал, что видом является группа сходных организмов, имеющих сходных предков, и что «...один вид никогда не зарождается из семян другого вида». Принимая вид в качестве реальной, но неизменной категории, Д. Рей классифицировал животные организмы на несколько групп по некоторым анатомическим особенностям, например, по строению копыт, рогов. Конечно, эта классификация была примитивной, но она все же дала начало естественным системам классификации.
Основы современной классификации растений и животных были заложены в XVIII в. шведским ученым К. Линнеем (1707-1778). Считая, что нахождение определенного порядка в природе является главнейшей целью науки, К. Линней в качестве основной (начальной) систематической (таксономической) единицы (таксона) определил вид, под которым он понимал совокупность организмов, сходных между собой, как сходны дети от одних родителей, и способных давать плодовитое потомство. Однако К. Линней считал, что со времени создания виды постоянны и неизменны. Он полагал также, что каждый классифицируемый организм должен быть сравним с идеальным типом и что все подобные организмы должны группироваться вокруг идеального типа. На основе изучения сходства организмов он подразделил животных на млекопитающих, птиц, пресмыкающихся, рыб, насекомых, червей, а также ввел в оборот такие таксономические единицы, как вид, род, отряд, класс, разместив их в виде иерархической системы и определив их сопод-чиненность, заключающуюся в том, что каждая категория более высокого уровня включает в себя одну или несколько категорий низшего порядка. Например, класс включает в себя несколько отрядов, отряд — несколько родов, а род — несколько видов.
Кроме того, для научного наименования организмов он ввел так называемую бинарную номенклатуру, в соответствии с которой наименование организмов одного вида, принадлежащих к одному роду, состоит из родового и видового латинских названий, причем первым словом является обозначение рода, вторым — вида. Например, научное название (бинарное обозначение) ландыша майского есть Convallaria majalis, садового гороха — Pissum sativum, домашней кошки — Fells doinast.ica, тигра — Felis tigris. По К. Линнею вслед за бинарным обозначением вида обычно указывают первооткрывателя вида и год открытия вида. Например, научное наименование аскариды человеческой будет Ascaris lumbricoides L., 1758. Это означает, что род аскариды человеческой есть Ascaris, вид есть lumbricoides, и что этот организм впервые был классифицирован К. Линнеем в 1758 г. Линнеевская система является естественной системой классификации.
Значительный вклад в систематику принадлежит французскому ученому Ж. Ламарку (1744-1829), который разделил животных на беспозвоночных и позвоночных, а также определил основные группы (типы) червей (плоские, круглые и кольчатые).
В XIX в. французский ученый Ж. Кювье (1769-1832) ввел в оборот понятие о типе животных и описал несколько типов.
Позднее, когда было разработано понятие о семействе, в соответствии с принципами естественной классификации виды животных и растений (лат. Species) стали объединяться в роды (лат. genus), роды — в семейства (лат. familia), семейства — в отряды (лат. ordo), отряды — в классы (лат. classis), классы — в типы (лат. typos), типы — в царства (лат. regnurn).
В XIX в. немецкий ученый Э. Геккель (1834-1919) разделил живой мир на три царства, а именно: протисты, животные и растения. Он ввел в обиход также понятие о генеалогическом древе, в котором главными категориями стали так называемые стволы. Из одного ствола происходят классы, отряды, семейства, роды. Позднее были предложены и другие подразделения царств.
В связи с совершенствованием классификации сейчас выделяют еще более дифференцированные систематические единицы в пределах основных систематических групп (таксонов), добавляя к ним приставку над или под (надцарство, подцарство, надсемейство, подсемейство, надтип, подтип). Наконец, часто выделяют такие таксо-ны, как раздел, надраздел, триба.
С введением в биологию теории эволюции Ч. Дарвина систематика организмов стала развиваться с учетом их естественного родства и происхождения (филогенетического родства). Поскольку сходство строения и функциональной активности обусловлено эволюционными связями, то учет этих особенностей определил эволюционное направление в систематике. Как считал Ч. Дарвин «Всякая истинная классификация есть генеалогическая». Таким образом, учет сходств строения и эволюционных связей между организмами стал классическим направлением в систематике.
С целью классификации организмов используют ряд методов. В частности, используют сравнительно-морфологический, сравнительно-эмбриологический, кариологический, эколого-генетический, географический, палеонтологический, молекулярно-генетический и другие методы. Что касается свойств организмов, важных для классификации, то ими являются одноклеточность или многокле-точность, дифференциация клеток, развитие зародышевых листков, процесс и степень развития определенных систем (кровеносной, пищеварительной и других), наличие или отсутствие целома, тип симметрии (радиальная или билатеральная), наличие или отсутствие сегментации тела, генетическое сходство, количество и морфология хромосом, строение пыльцевых зерен у растений, биохимические и иммунологические свойства. В наше время чрезвычайное значение приобрело установление последовательностей азотистых оснований в ДНК или секвенирование ДНК (генетическая дактилоскопия), а также установление последовательностей аминокислот в белках. Молекулярно-генетическая филогения основывается на представлениях о том, что последовательность азотистых оснований в ДНК и аминокислот в белках одного организма отличается от этих последовательностей другого организма. Следовательно, различия в этих последовательностях у разных организмов являются мерой эволюционных «расстояний» между организмами. Образцы различий могут быть выстроены в генеалогический ряд.
В обработке полученных результатов широко используют компьютерную технику.
Современная концепция в систематике является динамической. Она основана не только на использовании названных выше свойств, но и на учете географического распространения, экологических потребностей, генетических механизмов и степени, репродуктивной изоляции классифицируемых организмов.
В современной классификации растений и животных имеется много спорных вопросов, т. к. одни биологи склонны укрупнять систематические единицы, тогда как другие стремятся их детализировать. Поэтому существует несколько классификаций как растений, так и животных. В приводимом ниже описании разнообразия организмов используется классификация, исходным моментом которой является разделение живого мира на царства растений и животных.
1. Дайте определение систематике и назовите ее основные разделы.
2. Что вы понимаете под искусственными системами, когда их стали использовать и какова их роль сейчас в классификации организмов?
3. Что вы понимаете под естественными системами и какова их роль в классификации организмов?
4. Перечислите основные методы, используемые в систематике. Какие из них являются главными?
б. Назовите основные таксономические единицы и правила использования бинарной номенклатуры.
6. Почему в классификации организмов много спорных вопросов?
Грин Н„ Стаупг У., Тейлор Д. Биология. М.: Мир. 1998. 368 стр.
Нидон К„ Петерман И„ Шеффель П., Шайба Б. Растения и животные. М.: Мир. 1991. 260 стр.
Пехов А. П. Биология и общая генетика. М.: РУДН. 1993. 439 стр.
Полянский Ю.А. (ред.). Жизнь животных, т. 1. М.: Просвещение. 1987. 445 стр.
Хадоры Э„ Венер Р. Общая зоология. М.: Мир. 1989. 523 стр.
Яковлев Г. П., Челомбитъко В. А. Ботаника. М.: Высшая школа. 1990. 367 стр.
Rosenzweig M. L. Species Diversity in Space and Time. Cambridge University Press, 1995. 436 pp.
Для описания разнообразия растении мы используем систему классификации, основанную на макросистеме, предложенной А. Л. Тах-таджяном (1971). В обобщенном виде под влиянием этой макросистемы классификации мира растительных организмов начинают с разделения его на надцарство доядерные организмы (Procaryota) и надцарство настоящие ядерные организмы (Eucaryota).
(
PROCARYOTA)
К этому надцарству относят микроскопические организмы, тело которых представлено слоевищем, или талломом, не расчлененным ни на корень, ни на стебель, ни на листья. У них нет ядерной мембраны и организованного ядра. У них нет также стадии эмбрионального развития. Прокариотами являются в основном одноклеточные организмы. К ним относят и некоторые колониальные формы. Одноклеточные организмы растительной природы вместе с одноклеточными животными составляют значительную часть биомассы Земли.
Генетический материал у прокариотов представлен молекулой ДНК, находящейся в ядерной (центральной) зоне. Размножаются прокариоты путем простого деления, хотя у некоторых из них отмечается наличие аналога полового процесса в виде конъюгации.
Прокариоты не обладают ни хлоропластами, ни митохондри-ями. У них нет комплекса Гольджи и центриолей, но есть рибо-сомы. Характерной особенностью прокариот является наличие у них клеточной стенки. Известны подвижные формы этих организмов.
Некоторые прокариоты способны к фиксации азота атмосферы, но не способны к жизни в отсутствие кислорода (анаэробы).
В пределах этого надцарства Доядерные организмы выделяют лишь одно царство — царство дробянок (Mychota), которое классифицируют далее на подцарства Архебактерии (Archaeobacteria, или Archaeobacteriobionta), Настоящие бактерии (Bacteria, или Bacteriobionta) и Оксифотобактерии (Oxyphotobacteria, или Oxyphotobacteriobionta).
Подцарство Архебактерии (
Archaeobacteria
).
Организмы этого подцарства представлены метаногенными, галофильными и серо-зависимыми бактериями. Известно около 50 видов архебактерий. Выделяя здесь эту группу организмов в самостоятельное подцар-ство нельзя не отметить, что многие биологи классифицируют архебактерий в качестве самостоятельного царства (кроме царств растений и животных).
Метаногенные бактерии образуют метан путем восстановления диоксида углерода молекулярным водородом. Метан является основным продуктом их метаболизма. Считают, что весь метан биогенного происхождения на Земле образован деятельностью метано-генных бактерий. Их ежегодная производительность составляет около 1,0 х 109
т метана.
Эти бактерии обитают в строго анаэробных условиях в иле водоемов, в болотах и других местах, а также в желудочно-ки-шечном тракте человека и животных. Особенно много их в рубце жвачных.
Галобактерии — это обитатели горячих соленых водоемов. Благоприятной для них является среда, которая содержит Nad в количестве 20—30%, т. е. является насыщенным раствором.
Серозависимые бактерии являются обитателями горячих кислых водоемов и почв, вулканических расщелин. Образование месторождений серы обязано серозависимым бактериям.
В отличие от большинства настоящих бактерий для архебактерий характерен ряд особенностей. Например, плазматическая мембрана архебактерий имеет однослойную структуру, а в пептидогли-кане их клеточных стенок в отличие от клеточных стенок настоящих бактерий отсутствует муреин.
Уникальностью характеризуются также мембранные липиды архебактерий, ибо они не содержат эфиров глицерина и жирных кислот, но содержат изопреноидные углеводороды, которые обычно встречаются в нефти.
В ДНК некоторых архебактерий отмечается наличие повторяющихся последовательностей азотистых оснований, чего нет у настоящих бактерий. У галофилов обнаружен родопсиноподобный белок, обычно содержащийся в зрительном пурпуре многих позвоночных.
Схема синтеза белков, осуществляемого архебактериями, является такой же, как и у настоящих бактерий, однако в тРНК этих организмов нет ни тимина, ни урацила. Последний представлен псевдоуридином. Есть также отличия и в структуре рРНК.
Перечисленные особенности делают архебактерии по существу промежуточной формой между растениями и животными, что и явилось основанием для некоторых ученых к выделению их в самостоятельное царство.
Однако следует заметить, что многие ученые не разделяют эту точку зрения. Большинство специалистов классифицируют архебактерии в рамках подцарства.
Среди архебактерии встречаются как аэробы, так и хемоавтот-рофы и хемогетеротрофы. Архебактерии играют важную роль в природе.
Классификация архебактерии еще плохо разработана.
Архебактерии являются древнейшими прокариотами. Возможно, что они были самыми первыми организмами на Земле.
Подцарство Настоящие бактерии (
Bacteria
).
Бактерии являются одноклеточными микроскопическими организмами, размеры которых измеряются микронами. На основе метода окраски по Граму различают грамположительные и грамотрицательные бактерии. В зависимости от формы среди бактерий различают бациллы, стафилококки, диплококки, стрептококки, вибрионы, спириллы (рис. 1). Бактерии многих видов подвижны, обладая жгутиками или ресничками.
Бактерии содержат почти все структурные элементы, которые характерны для клеток растений и животных (см. гл. VI). Однако в отличие от клеток растений и животных бактерии не обладают хлоропластами (митохондриями), центриолями, ядерной мембраной, ядрышком.
Бактерии являются обитателями практически всех экологических ниш.
Размножение бактерий происходит путем простого деления или путем спорообразования. У бактерий многих видов открыта конъюгация, являющаяся аналогом полового процесса и зависимая от содержания в них плазмид. Бактерии, обладающие плазмидами, служат в качестве доноров генетического материала. Бактерии, не обладающие плазмидами, являются реципиентами этого материала (см. гл. X).
Большинство бактерий является гетеротрофами, а часть их является автотрофами или хемосинтезирующими. Бактерии многих видов являются аэробами, но встречаются и анаэробы.
Значение бактерий в природе очень велико. Они вызывают брожение, гниение, а также минерализацию органических веществ, результатом деятельности бактерий является накопление карбонатов, сульфидов, фосфатов, бокситов, образование железных руд и т- д. Клубеньковые бактерии, колонизируя корневые клубеньки бобовых растений, усваивают азот атмосферы благодаря наличию в них специальных органелл — симбиосом. Из симбиосом азот в форме NHg или NH44
' под действием бактериального мембранного белка транспортируется затем в клетки растений.

Рис. 1. Морфологические формы бактерий:
1 – стафилококки; 2 – стрептококки; 3 – диплококки; 4 – бациллы;
5 – спириллы; 6 – капсульные бациллы и кокки; 7 – жгутиковые бациллы
Бактерии широко используют в практике. Например, бактерии молочного брожения используют в производстве молочнокислых продуктов, а также в консервировании овощей, силосовании кормов для животных.
Известны также виды бактерий, являющиеся продуцентами антибиотиков. Бактерии используют в генной инженерии для клони-рования и поддержания в них векторных молекул ДНК (плазмид) и гибридных молекул ДНК (см. раздел VI).
Многие виды бактерий являются возбудителями бактериальных болезней человека, животных и растений. Наиболее известными бактериальными болезнями человека являются дизентерия, чума, холера, дифтерия и другие. У человека и животных бактерии вызывают туберкулез, бруцеллез, сибирскую язву и другие болезни.
Бактерии являются древнейшими обитателями Земли. Однако их происхождение не совсем ясно. Предполагают, что они произошли от самых примитивных организмов, называемых проге-нотами и оцениваемых в качестве предшественников прокариот. Но сведения о строении и функциях прогенот отсутствуют и это мешает установлению действительных связей между прогенота-ми и бактериями.
Подцарство Оксифотобактерии (
Oxyphotobacteria
, или
Oxyphotobacteriobionta
)
. Это подцарство представлено отделами ци-анобактерий и хлороксибактерий.
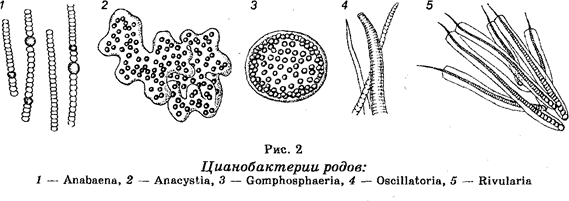
Отдел Цианобактерии (Cyanobacteria). Строение цианобакте-рий (по старой ботанической терминологии — сине-зеленых водорослей) до некоторой степени сходно со строением бактерий (рис. 2). Известно около 2500 видов. В основном они являются одноклеточными организмами разной формы (округлой, цилиндрической), но могут образовывать длинные многоклеточные нити или даже объединяться в колонии. Однако они отличаются от настоящих бактерий тем, что их клеточные стенки содержат некоторое количество целлюлозы и что они способны к фотосинтезу, т. к. в цитоплазме содержат хлорофилл (в гранулах, но не в хлоропластах) и другие пигменты (каротин, ксантофил и фикобилины), создающие их окраску. Некоторые клетки в многоклеточных цианобактериях обладают способностью фиксировать азот атмосферы.
Обитают в пресной и соленой воде, входя в состав фитопланктона, а также являются обитателями почвы. Отдельные виды встречаются в морях. При неблагоприятных условиях способны образовывать споры. Они могут также находиться в симбиотических отношениях с грибами (см. § 33).
Размножение цианобактерий происходит путем простого деления. При интенсивном размножении вызывают «цветение» воды.
Осуществляя фотосинтез, цианобактерий ответственны за появление значительного количества кислорода в атмосфере. Хозяйственного значения не имеют, если не считать, что их несгнившие остатки участвуют в образовании лечебных грязей.
Считают, что цианобактерий тоже являются древнейшими обитателями Земли. Они существовали еще до появления папоротников (щитовников), мхов и семенных растений. Их возраст составляет несколько миллиардов лет. По мнению многих ботаников они составляют тупиковую ветвь в эволюции.
Отдел Хлороксибактерии (род РгосЫогоп). Организмы этого отдела немногочисленны (в видовом составе). Они также способны к фотосинтезу, т. к. содержат хлорофилл. Они содержат и другие пигменты, обнаруживаемые в клетках зеленых растений.
В пределах этого надцарства у растений выделяют царство грибов и царство растений.
Царство Грибы —
Mycota
(
Fungi
).
Для организмов этого царства, которое представляет собой очень гетерогенную группу организмов, характерны значительная выраженность клеточной оболочки, неподвижность в вегетативном состоянии, гетеротрофный тип питания путем всасывания (адсорбции) и неограниченный рост. Насчитывают около 100 000 видов грибов, характеризующихся разнообразием как по строению, размерам (рис. 3), местам обитания, так и по физиологическим функциям. Оптимальные температуры для роста грибов равны 20—26°С. Обитают практически во всех географических зонах, встречаясь в воде (пресной и морской) и почве, на мертвом органическом материале, многие виды паразитируют в тканях растений и животных, включая человека, причем степень паразитизма весьма различна.
Грибы могут вступать в симбиотические отношения с другими организмами, например, с водорослями или цианобактериями, образуя лишайники. Они могут также вступать в симбиоз с высшими растениями, обволакивая и проникая в корни растений своими гифами и формируя структуры (корень + гриб), получивший название микориз. Такой симбиоз с растениями обеспечивает потребность последних в фосфатах. Например, 80% наземных растений, включая и многие сельскохозяйственные растения, формируют симбиоз с грибом Glornus versiforme, который обитает на их корнях и облегчает им восприятие фосфатов и минеральных питательных веществ из почвы.
Среди организмов этого царства встречаются как одноклеточные (микроскопические), или низшие, так и многоклеточные (высшие) грибы.
Грибы классифицируют на отделы: Настоящие грибы, Оомице-ты и Лишайники.
Среди Настоящих грибов различают классы Хитридиевые грибы, Зигомицеты, Аскомицеты (Сумчатые грибы), Базидиомицеты и Несовершенные грибы (Дейтеромицеты).
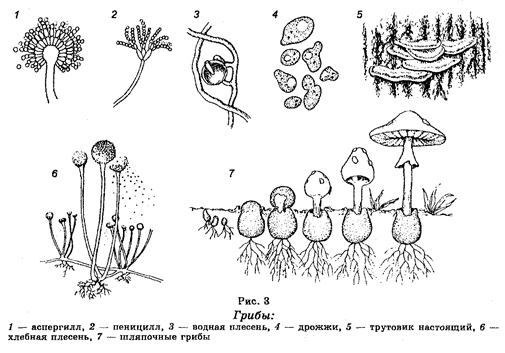
Хитридиевые грибы представлены микроскопическими организмами, тело которых имеет вид цитоплазматической массы. Они ведут паразитический образ жизни на водных растениях и животных. Известен также вид этих грибов, вызывающий рак картофеля.
Зигомицеты являются наземными грибами. Тело их представлено неклеточным мицелием. Эти грибы ведут как сапрофитичес-кий, так и паразитический образ жизни, паразитируя на разных животных. Среди сапрофитов наиболее известными являются так называемые мукоровые грибы (плесени хлеба, овощей и других продуктов).
Отдельные виды зигомицетов являются паразитами, вызывая микозы человека и животных.
Аскомицеты представляют собой наиболее многочисленную группу грибов (более 30000 видов), различающихся между собой прежде всегоразмерами. Встречаются как одноклеточные, так и многоклеточные формы. Тело их представлено гаплоидным мицелием. Образуют аски (сумки), содержащие аскоспоры, что является характерным признаком этих грибов. Среди грибов данной группы наиболее известными являются дрожжи (пивные, винные, кефирные и другие). Например, дрожжи Saccharomices cerevisiae влияют на ферментацию глюкозы (CgH^Og). Одна молекула глюкозы дает в ходе этого ферментативного процесса две молекулы этилового спирта.
Базидиомицеты являются высшими грибами. Они характеризуются большими размерами, которые могут доходить даже до полуметра. Их тело также состоит из мицелия (грибницы), но многоклеточного, формирующего грибы. Протопласт грибных клеток содержит не только ядра, но и митохондрии, рибосомы, аппарат Гольджи и даже гликоген в качестве запасного вещества. Гифы переплетаются, образуя плодовые тела, которые в обыденной жизни называют грибами, состоящими из ножки и шляпки.
Эти грибы размножаются как вегетативным и бесполым, так и половым путем. Наиболее известными базидиомицетами являются шляпочные грибы, среди которых имеются как съедобные, так и ядовитые.
Несовершенные грибы тоже весьма многочисленны (около 30 000 видов). Очень распространены в разных географических зонах. Среди них есть как сапрофиты, которые широко участвуют в разложении остатков растений и в почвообразовательных процессах, так и паразитические формы. Наиболее известными видами этих организмов являются грибы из рода Fusarium, которые вызывают у многих культурных растений (хлопчатник, лен и другие) болезнь, называемую вилтом. В числе несовершенных грибов широко известны также грибы из родов Penicillium и Aspergillus. Они широко используются в промышленном производстве лимонной, фумаровой и других органических кислот, а также ряда ферментов. Отдельные виды этих родов являются продуцентами антибиотиков, широко используемых в медицине и ветеринарной медицине.
Оомицеты — это в основном водные и почвенные грибы. Среди этих грибов очень известны виды из рода Phytophtora, которые вызывают болезни картофеля, томатов и других пасленовых.
Грибы играют значительную роль в природе. В частности, они являются организмами-разрушителями. Входя в состав многих экологических систем, ответственны за разрушение органического материала растительного происхождения, т. к. продуцируют ферменты, действующие на целлюлозу, лигнин и другие вещества растительных клеток. Их широко используют в сыроваренной промышленности для производства многих популярных сортов сыра. Нельзя не отметить, что Neurospora crassa принадлежит выдающаяся роль в качестве экспериментального объекта в познании многих метаболических путей.
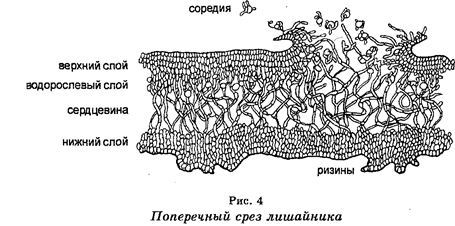
Лишайники
представляют собой сложные организмы, образованные в результате симбиоза между грибами, водорослями зелеными, или цианобактериями, и азотобактером (рис. 4). Следовательно, лишайник — это комбинированный организм, т. е. гриб + водоросль + азотобактер, существование которого обеспечивается тем, что гифы гриба ответственны за поглощение воды и минеральных веществ, водоросль — за фотосинтез, а азотобактер — за фиксацию азота атмосферы. Лишайники являются обитателями всех ботанико-географических зон. Размножаются вегетативным, бесполым и половым путем.
Значение лишайников в природе велико. Из-за высокой чувствительности к загрязнителям среды лишайники используют в качестве индикаторов чистоты атмосферы. На севере они являются главным кормом для оленей. Их используют также в аптечном деле и в парфюмерии.
Грибы имеют древнее происхождение. Их ископаемые остатки отмечены в силуре и девоне. Отдельные ботаники предполагают, что они произошли от зеленых водорослей, потерявших хлорофилл. Более распространенный взгляд заключается в том, что грибы произошли от жгутиковых (простейших).
Ископаемые остатки лишайников также найдены в девоне, что определяет их возраст примерно в 400 млн лет. Предполагают, что образование лишайников явилось первым случаем установления симбиотических отношений между организмами. Это обеспечило возможность их широкого распространения в разных экологических нишах.
Царство Растения (
Plantae
или
Vegetabilia
)
. Это царство представлено организмами, клетки которых обладают плотными клеточными стенками и которые способны к фотосинтезу. Растения этого царства классифицируют на три подцарства, а именно: багрянки (Phycobionta), настоящие водоросли (Phycobionta) и высшие растения (Embryophyta).
Тело багрянок и настоящих водорослей не расчленено на ткани и органы. По этой причине их часто называют низшими, или слоевцовыми растениями. Напротив, остальные растения известны в качестве высших растений, т. к. характеризуются наличием разных тканей и расчленением тела на органы. Эти растения приспособлены к жизни в наземных условиях.
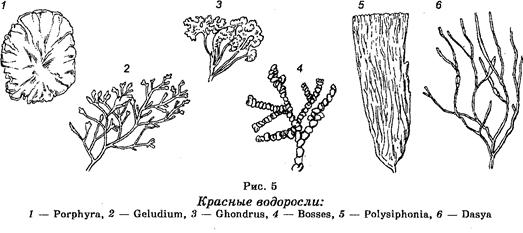
Подцарство Багрянки (
Rhodophyta
).
Растения этого подцарства являются многоклеточными организмами (рис. 5). Тело багрянок представлено слоевищем. Насчитывают около 4000 видов багрянок, среди которых наиболее известными являются порфира, не-малион, кораллины и другие. Их багряная окраска зависит от содержания в них хлорофилла, каротиноидов, красных фикоэритри-нов, синих фикоцианинов и других пигментов. Являются обитателями больших глубин морей и океанов. Часто их называют красными водорослями. Особенно ими богато Красное море.
Размножаются как бесполым, так и половым путем с чередованием полового и бесполого поколений.
Имеют хозяйственное значение. Отдельные виды служат сырьем, из которого добывают агар-агар. В ряде стран их используют на корм скоту
Багрянки являются древними организмами, но происхождение их и филогенетические связи между отдельными видами остаются невыясненными.
Подцарство Настоящие водоросли (
Phycobionta
).
Настоящие водоросли являются растениями, тело которых представлено слоевищем. Известно около 30 000 видов этих организмов. Встречаются как одноклеточные, так и многоклеточные водоросли. Они являются обитателями в основном пресноводных водоемов и морей, но встречаются почвенные водоросли и даже водоросли снега и льда. Размножение одноклеточных водорослей происходит путем деления, многоклеточные формы размножаются как бесполым, так и половым путем. Когда-то Вергилий писал — «nigilvilor algo» (ничего нет хуже водорослей). В наше время водоросли приобрели Другие оценки.
Альгологи классифицируют водоросли на несколько отделов.
Отдел Зеленые водоросли (
Chlorophyta
).
Этот отдел представлен подвижными и неподвижными одноклеточными и многоклеточными организмами, обладающими довольно толстой клеточной стенкой и имеющими форму нитей, трубочек (рис. 6). Некоторые виды формируют подвижные и неподвижные колонии. Насчитывают свыше 13 000 видов этих водорослей, большинство которых является обитателями пресных водоемов. Но известны и морские формы.
Одноклеточные и многоклеточные зеленые водоросли способны к фотосинтезу, т. к. содержат хлоропласты, в которых концентрируется хлорофилл и от наличия которого они имеют зеленую окраску. Они обладают также ксантофилом и каротином.
Типичными представителями одноклеточных зеленых водорослей являются хламидомонады (из рода Chlamidomonas), обитающие в лужах и других небольших пресных водоемах, и хлорелла из одноименного рода (Chlorella), которая обитает в пресных и соленых водах, на поверхности сырой земли, на коре деревьев. Хлорелла обладает исключительной фотосинтезирующей активностью, будучи способной улавливать и использовать 10-12% световой энергии. Содержит ряд ценных белков, витамины В, С и К.
Примером многоклеточных зеленых водорослей является обитатель прудов вольвокс (Volvox). Формируя колонию, этот организм состоит из 500—60 000 клеток, каждая из которых снабжена двумя жгутиками, а также содержит глазок, дифференцированное ядро и хлоропласт. Толстая студенистая оболочка окружает каждую клетку и отделяет ее от соседних клеток. Если в колонии одна клетка погибает, остальные продолжают жить. Расположение клеток в колонии обеспечивает движение этого организма.
Размножаются путем деления или образования подвижных зооспор, которые отделяются от материнского организма, прикрепляются к какому-либо субстрату, а затем развиваются в новый организм. У спирогир имеет место половой процесс в виде конъюгации.
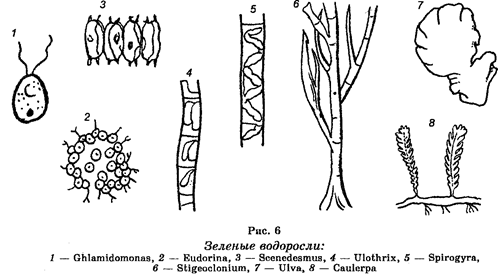
Хозяйственное значение этих водорослей невелико, если не считать, что из-за богатого содержания белков и витаминов хлореллу используют в корм для животных. Будучи компонентом фитопланктона, служит кормом для рыб.
Предполагают, что зеленые водоросли возникли в результате ароморфозов, которыми оказались образование ядра, появление мно-гоклеточности и полового процесса. Предполагают также, что они дали начало примитивным наземным растениям, ставшими пред-ковыми формами мохообразных.
Отдел Диатомовые
водоросли, или диатомеи (
Chrysophyta
) представлен в основном многоклеточными организмами, а иногда даже и колониальными формами (рис. 7). Встречаются и одноклеточные формы. Известно 5700 видов. Характеризуются четкой дифференциацией тела на цитоплазму и ядро. Клеточная стенка «пропитана» кремнеземом, в результате чего ее называют панцирем. Являются обитателями пресных водоемов, морей и океанов и входят в состав фитопланктона.
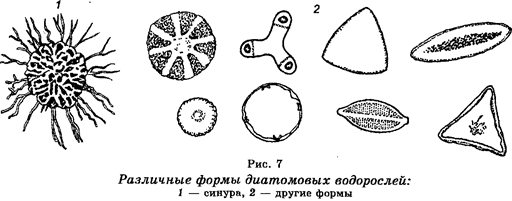
В клетках этих водорослей имеются хлоропласты в виде зерен или пластинок, которые окрашены в разные цвета из-за содержания разных пигментов (каротин, ксантофилл и его вариант диато-мин). По этой причине диатомовые водоросли часто называют золотисто-коричневыми.
Размножение происходит путем деления клеток пополам. У отдельных видов существует половое размножение. Диатомеи — это диплоидные организмы.
Напластования отмерших диатомовых водорослей дали начало диатомиту, который на 50-80% состоит из их панцирей и который используют в качестве поглотителей в химии и пищевой промышленности.
Значение диатомовых водорослей в природе очень большое. Они занимают исключительно важное место в круговороте веществ, являясь главным кормом для рыб. Их пищевая ценность является очень высокой.
Эволюционно диатомовые водоросли стоят ближе всего к зеленым водорослям, но происхождение их неясно.
Отдел Бурые водоросли (
Phaeophyta
).
Эти водоросли являются многоклеточными организмами. Каждая клетка содержит лишь одно ядро. По размерам они самые большие (длинные) водоросли, достигая в длину нескольких десятков метров (рис. 8). Известно около 900 видов. Являются обитателями морей и океанов, включая северные. Их пигментация определяется тем, что они содержат хлоропласты, окрашенные в бурый цвет из-за содержания хлорофилла, а также бурых пигментов (каротина, ксантофилла и фукок-сантина).
Наиболее известными являются водоросли из родов Laminaria и Fucus.
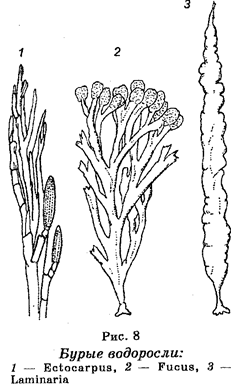 Размножаются вегетативным, бесполым и половым путем. Вегетативное размножение происходит частями сло-евища, бесполое (споровое) — с помощью гаплоидных спор, развивающихся в гаметофит, половое — путем изогамии, гетерогамии или оогаши. Характерно чередование гаплоидного и диплоидного поколений. Половые клетки снабжены жгутиками. Размножаются вегетативным, бесполым и половым путем. Вегетативное размножение происходит частями сло-евища, бесполое (споровое) — с помощью гаплоидных спор, развивающихся в гаметофит, половое — путем изогамии, гетерогамии или оогаши. Характерно чередование гаплоидного и диплоидного поколений. Половые клетки снабжены жгутиками.
Хозяйственное значение этих водорослей, особенно ламинарий, очень велико. Из них добывают йод, калийные соли, агароподоб-ные вещества, используемые в пищевой промышленности. Ламинарии, известные под названием «морской капусты», используются в пищу человеком. Некоторые водоросли используют в качестве удобрения.
Бурые водоросли — древнейшие водные растения. Предполагают, что они дали начало папоротниковидным растениям.
Заканчивая краткое изложение данных о водорослях, следует отметить, что в целом водоросли имеют важное значение во многих экологических системах. Фактически, они являются главным источником органических веществ в водоемах. Подсчитано, что водоросли ответственны за ежегодный синтез в Мировом океане органического вещества в количестве 550 млрд тонн, что составляет значительную часть продуктивности всей биосферы. Далее им принадлежит очень значительная роль в обогащении кислородом атмосферы. Наконец, водоросли участвуют в самоочищении водоемов, в почвообразовании.
Подцарство Высшие растения (
Embryophyta
или
Embryobionta
). Растения, входящие в состав этого подцарства, часто называют ли-стостебельными, поскольку их тело расчленено на стебель, лист и корень. Кроме того, их называют еще зародышевыми, т. к. они содержат зародыш. Наконец, их называют сосудистыми растениями (кроме моховидных), поскольку в органах их спорофитов имеются сосуды и трахеиды.
Высшие растения в ходе исторического развития приспособились к жизни в наземных условиях. У этих растений отмечается чередование полового (гаметофит) и бесполого (спорофит) поколений. Гаметофит продуцирует гаметы и защищает зародыш, тогда как спорофит продуцирует споры, которые обеспечивают следующую генерацию гаметофита. У высших растений доминирует диплоидный спорофит, который и определяет внешний вид растения.
В подцарстве Высшие растения различают высшие споровые и высшие семенные растения. Для высших споровых характерно разделение полового и бесполого размножения. В первом случае размножение происходит одноклеточными спорами, образующимися в спорангиях спорофитов, во втором — гаметами, образующимися в половых органах гаметофитов. Для высших семенных растений характерно наличие многоклеточного образования — семени, образующегося в процессе размножения и придающего семенным растениям важнейшее эволюционное преимущество перед споровыми.
Подцарство Высшие растения классифицируют на несколько отделов. В частности, высшие споровые растения классифицируют на отделы Риниофиты (Rhyniophyta) и Зостерофиллофиты (Zostrophyllophyta), организмы которых полностью вымерли, а также на ныне существующие отделы Моховидные (Bryophyta), Плау-новидные (Lycopodiophyta), Псилотовидные (Psilotophyta), Хвощевидные (Eguisetophyta), Папоротниковидные (Polypodiophyta). Высшие семенные растения классифицируют на отделы Голосеменные (Gymnospermae) и Покрытосеменные, или Цветковые (Angiospermae, или Magnoliophyta). Голосеменные и Покрытосеменные — это семенные растения, тогда как все остальные — это высшие споровые растения. У части высших споровых все споры одинаковы (равноспоровые растения), а у некоторых споры имеют разную величину (разноспоровые растения).
Из растений современных отделов ниже будут рассмотрены лишь отдельные из них.
 Отдел Моховидные
(Bryophyta). Этот отдел представлен низкорослыми, многолетними растениями. У некоторых из них тело представлено слоевищем, но у большинства расчленено на стебель и листья (рис. 9). Насчитывают около 25 000 видов моховидных. Являются обитателями сырых мест во всех географических зонах. К почве прикрепляются с помощью волосовидных выростов, называемых ризоидами. Через эти структуры они осуществляют почвенное питание. Наиболее известными представителями этого типа являются кукушкин лен, маршанция многообразная, мхи рода сфагнум (300 видов). Отдел Моховидные
(Bryophyta). Этот отдел представлен низкорослыми, многолетними растениями. У некоторых из них тело представлено слоевищем, но у большинства расчленено на стебель и листья (рис. 9). Насчитывают около 25 000 видов моховидных. Являются обитателями сырых мест во всех географических зонах. К почве прикрепляются с помощью волосовидных выростов, называемых ризоидами. Через эти структуры они осуществляют почвенное питание. Наиболее известными представителями этого типа являются кукушкин лен, маршанция многообразная, мхи рода сфагнум (300 видов).
 В развитии мхов характерно чередование полового (гаметофи-та) и бесполого (спорофита) поколений. На растениях полового поколения образуются споры разных размеров. После оплодотворения женских половых клеток мужскими развивается спорофит (спорангий со спорами), клетки которого имеют диплоидный набор хромосом. Образующиеся в результате мейоза в спорангии споры имеют гаплоидный набор хромосом. Высыпаясь на почву, споры прорастают, давая начало растению, гаметофиту, имеющему в размножающихся митозом клетках гаплоидный набор хромосом. Гаплоидный гаметофит доминирует в цикле развития. На гаметофите вновь образуются половые клетки, и процесс повторяется. Специфической особенностью этих растений является не только доминирование гаплоидного гаметофита, но также и то, что гаметофит (половое поколение) и спорофит (бесполое поколение) представляют собой одно растение. В развитии мхов характерно чередование полового (гаметофи-та) и бесполого (спорофита) поколений. На растениях полового поколения образуются споры разных размеров. После оплодотворения женских половых клеток мужскими развивается спорофит (спорангий со спорами), клетки которого имеют диплоидный набор хромосом. Образующиеся в результате мейоза в спорангии споры имеют гаплоидный набор хромосом. Высыпаясь на почву, споры прорастают, давая начало растению, гаметофиту, имеющему в размножающихся митозом клетках гаплоидный набор хромосом. Гаплоидный гаметофит доминирует в цикле развития. На гаметофите вновь образуются половые клетки, и процесс повторяется. Специфической особенностью этих растений является не только доминирование гаплоидного гаметофита, но также и то, что гаметофит (половое поколение) и спорофит (бесполое поколение) представляют собой одно растение.
Значение моховидных в природе заключается в том, что, находясь в экосистемах, они воздействуют на среду обитания многих видов других растений, равно как и животных. Интенсивное размножение мхов способствует ухудшению почвы. Отмирая, сфагновые мхи «оторфовываются» и образуют залежи торфа. Некоторые виды используются в медицинской промышленности.
Полагают, что растения этой группы были одними из первых наземных растений и широко произрастали еще 450-500 млн лет назад и что эволюция их заключалась в регрессивном развитии спорофита. Считают, что моховидные являются слепой эволюционной ветвью.
Отдел Папоротниковидные
(Palypodiophyta). В пределах этого отдела классифицируют травянистые растения, также обитающие в сырых местах (рис. 10). Некоторые Папоротниковидные, обитающие в тропиках, представлены древесными формами, отдельные из которых достигают 25 метров в высоту. Насчитывают более 10 000 видов этих растений. Типичными представителями папоротниковидных являются папоротники.
Для папоротниковидных также характерно чередование полового и бесполого поколений, однако, в отличие от моховидных, у организмов, принадлежащих к этому отделу, преобладающим является спорофит, для которого характерна диплоидность. У спорофита имеются основные органы — стебель, листья, корень. Напротив, гаметофит характеризуется очень малыми размерами, представляя небольшую пластинку, прикрепленную к почве с помощью ризоидов.
Для папоротниковидных характерен сложный цикл развития. Цикл начинается с развития изоспор гаметофита (заростка), на котором образуются половые органы в виде антеридиев и архегониев. В последних развиваются половые клетки. После их оплодотворения из зиготы образуется спорофит, на котором образуются споры, дающие начало гаметофиту. Большинство папоротниковидных представлено разноспоровыми растениями.
Значение папоротниковидных в природе большое, т. к. они входят в состав многих экосистем. Хозяйственное значение современных папоротниковидных небольшое, если не считать, что растения отдельных видов служат лекарственным сырьем.
Папоротниковидные классифицируют на 7 отделов, большинство из которых представлено вымершими видами.
Папоротниковидные являются наиболее древними споровыми растениями. Они уже были в девоне, а в карбоне составляли леса из растений, высота которых достигала до 30 м. Остатки этих растений принимали участие в образовании каменного угля.
Отдел Голосеменные
(Gymnospermae). Растения этого отдела дают семена, которые представляют собой, по существу, готовые зародыши будущих растений. Основными органами семени являются зародышевый корешок, зародышевый стебелек, зародышевые листки. Однако у голосеменных семя не покрыто плодолистиками. По этой причине их называют голосеменными.
Голосеменные представлены деревьями, кустарниками и лианами. Количество видов составляет около 700. Распространены по всему земному шару. В северном полушарии занимают огромные площади, образуя хвойные леса.
Наиболее известными представителями голосеменных являются сосна, пихта, ель, лиственница. У растения этого отдела тканями ствола и корня являются камбий, ксилема, флоэма. У многих листья представлены «иголками».
Для голосеменных характерно чередование поколений, связанное со сменой гаплоидного и диплоидного состояний, однако у них налицо уменьшение гаметофита. Можжевельник, саговник, туя, ель, сосна, лиственница — это спорофиты. Как и все семенные растения, голосеменные являются разноспоровыми. Органами размножения являются женские и мужские шишки, которые формируются на одном и том же дереве и в которых находится гаметофит.
Образование семени является первым этапом в развитии спорофита. Женские шишки построены из крупных чешуек, называемых мегаспорофиллами, каждая из которых несет по два мегаспорангия на внутренней поверхности, а каждый мегаспорангий в свою очередь содержит мегаспору, которая развивается в многоклеточный гаметофит, содержащий две или три архегонии. Каждая архегония состоит из одиночной большой яйцеклетки и нескольких малых вытянутых клеток. Мегаспорангий покрыт так называемым интегу-ментом. Мегаспорангий с интегументом называют семязачатком.
Мужские шишки несут на внутренней поверхности их чешуи (на микроспорофиллах) по два микроспорангия, содержащих микроспоры, каждая из которых развивается в гаплоидную пыльцу. Пыльцевые гранулы (зерна) составляют мужской гаметофит.
Мегаспорофиллы и микроспорофиллы собраны в мега- и мик-ростробиллы (соответственно) на укороченном спороносном побеге, представляющем собой стебель со спороносными листьями.
 Когда пыльца попадает на женские шишки, она проходит в семязачаток, причем каждая пыльцевая гранула развивается в тычиночную трубочку и два спермоядра, а когда тычиночная трубочка проникает в яйцеклетку, происходит слияние спермоядра с ядром яйцеклетки. Это и есть оплодотворение. Диплоидная зигота становится диплоидным зародышем. Со временем внешний инте-гумент семязачатка превращается в оболочку семени, а из остатков мегаспорангия образуется эндосперм. Следовательно, семязачаток превращается в семя. После созревания семена из шишек выпадают наружу. Когда пыльца попадает на женские шишки, она проходит в семязачаток, причем каждая пыльцевая гранула развивается в тычиночную трубочку и два спермоядра, а когда тычиночная трубочка проникает в яйцеклетку, происходит слияние спермоядра с ядром яйцеклетки. Это и есть оплодотворение. Диплоидная зигота становится диплоидным зародышем. Со временем внешний инте-гумент семязачатка превращается в оболочку семени, а из остатков мегаспорангия образуется эндосперм. Следовательно, семязачаток превращается в семя. После созревания семена из шишек выпадают наружу.
Хозяйственное значение голосеменных очень большое. Они дают древесину, сырье для медицинской промышленности. Многие виды являются декоративными.
Голосеменные представляют собой очень древнюю группу высших растений. Появившись в девоне (около 350 млн лет назад), голосеменные в конце палеозоя — начале мезозоя заняли место папо-ротниковидных, поскольку оказались более приспособленными к жизни в наземных условиях. Одна их гипотез заключается в том, что голосеменные произошли от древнейших папоротниковидных.
Отдел Покрытосеменные
, или Цветковые
(Angiospermae, или Magnoliophyta). Растения этого отдела встречаются почти повсеместно. На их долю приходится 250 000-300 000 видов, т. е. почти две трети видов царства растений. В настоящее время они являются самой процветающей группой растений.
В пределах этого отдела различают однодольные и двудольные растения, которые бывают как травянистыми и кустарниковыми видами, так и деревьями. Типичными представителями этого отдела являются рожь, пшеница, роза, береза, осина и другие. Различают однодольные и двудольные покрытосеменные растения.
Для этих растений также характерно чередование поколений, но у них произошло значительное уменьшение гаметофита.
Замечательной особенностью этих растений является наличие у них цветка, который представляет собой видоизмененный побег и является производным спорофита (рис. 11). Именно по этой причине растения, образующие цветки, называют цветковыми. Как правило, цветки обоеполы, но иногда и раздельнополы. В цветке различают пестик и тычинки, которые являются его главными частями. В нижней части пестика (завязи) развиваются семена. По этой причине эти растения получили название покрытосеменных. Нижняя часть пестика представлена завязью, узким столбиком и рыльцем. Что касается тычинок, то каждая из них состоит из тычиночной нити и пыльника.
У обоеполых растений, которые среди покрытосеменных составляют большинство, цветки имеют как пестики, так и тычинки, т. е. эти растения имеют пестичные (женские) и тычиночные (мужские) цветки. Но у многих видов одни цветки имеют только пестики, на другом — только тычинки. Такие растения называют двудомными. Опыление является результатом переноса пыльцы с тычинок на рыльце пестика.
Общая схема репродукции покрытосеменных на рис. 12.
Женский гаметофит цветковых растений состоит из 8 клеток зародышевого мешка, одна из которых является яйцеклеткой. Эта микроскопическая структура развивается из одиночной мегаспоры. Мужской гаметофит развивается из микроспоры, или пыльцевой гранулы, располагающейся в микроспорангии пыльника. Попав на рыльце пестика, пыльцевая гранула в результате деления дает начало генеративной клетке и клетке, развивающейся в пыльцевую трубку. Далее пыльцевая трубка врастает в полость завязи. Ядро трубки генеративной клетки мигрирует к низу пыльцевой трубки, где генеративная клетка делится, давая два спермин. Один из этих спермиев сливается с яйцеклеткой, образуя диплоидную зиготу, тогда как второй спермий сливается с ядром (в центре зародышевого мешка, в семязачатке), давая триплоидное ядро, развивающееся затем в эндосперм. В конечном итоге обе структуры оказываются в семени, а семя оказывается в завязи, которая развивается в плод. Последний может содержать от одного до нескольких семян. Такое оплодотворение называют двойным (рис. 13). Оно было открыто в 1898 г. С. Г. Навашиным (1857—1950). Биологический смысл двойного оплодотворения заключается в том, что развитие триплоидного эндосперма в сочетании с огромным числом поколений обеспечивает экономию пластических и энергетических ресурсов растений.
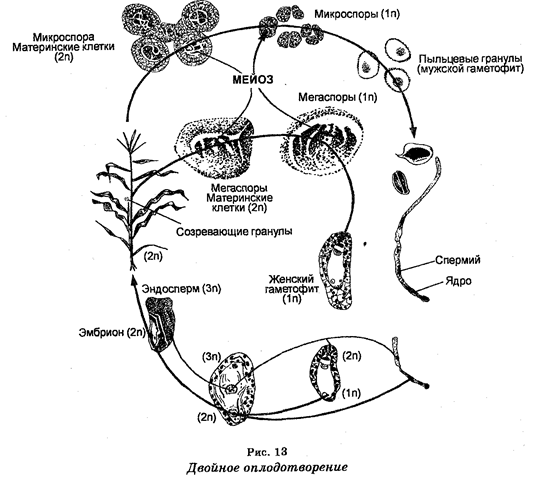
 Оно было открыто в 1898 г. С. Г. Навашиным (1857—1950). Биологический смысл двойного оплодотворения заключается в том, что развитие триплоидного эндосперма в сочетании с огромным числом поколений обеспечивает экономию пластических и энергетических ресурсов растений. Оно было открыто в 1898 г. С. Г. Навашиным (1857—1950). Биологический смысл двойного оплодотворения заключается в том, что развитие триплоидного эндосперма в сочетании с огромным числом поколений обеспечивает экономию пластических и энергетических ресурсов растений.
 Стебель является органом растений, к которому прикрепляются листья, корни, цветки. (Строение стебля древесного растения показано на рис. 14.) Стебель является органом растений, к которому прикрепляются листья, корни, цветки. (Строение стебля древесного растения показано на рис. 14.)
Листья являются важнейшим органом растений. Они характеризуются разной формой и построены из нескольких слоев клеток, содержащих большое количество хлоропластов. Служат органом газообмена между растениями и средой. Из-за наличия хлорофилла в листьях происходит фотосинтез, основу которого составляют две реакции — фотолиз воды и фиксация COg.
Корень является органом растения, который адсорбирует воду и минеральные вещества из почвы и проводит их к стеблю. У покрытосеменных, как и голосеменных, вода и питательные вещества из почвы адсорбируются корневыми волосками и проводятся в ксилему в результате осмотического давления в корневой системе, действия капилляров, отрицательного давления в ксилеме, доходящего иногда у некоторых древесных форм до 100 бар, и транспирации, т. е. испарения воды из листьев (рис. 15).
Хозяйственное значение покрытосеменных переоценить очень трудно, т. к. они исключительно широко используются в жизни человека (источник продовольствия, сырье для промышленности, корм для животных и т. д.).
Покрытосеменные растения являются господствующими растениями нашей планеты. Поэтому объяснение их происхождения уже давно оказалось одной из самых важных задач в учении об эволюции. Начиная с Ч. Дарвина, для объяснения покрытосеменных растений было выдвинуто несколько гипотез. По одной из них предполагают, что покрытосеменные произошли от каких-то голосеменных, а однодольные происходят от каких-то древних двудольных. Однако эта и другие гипотезы не являются исчерпывающими. Существуют разногласия и в определении времени появления покрытосеменных. По новейшим представлениям главная диверсификация цветковых растений, в том числе разделение на однодольные и двудольные, произошла 130—90 млн лет назад, и это дало тогда начало изменениям земных экосистем.
1. Как вы понимаете различия между доядерными и ядерными организмами?
2. Назовите подцарства доядерных организмов.
3. Что вы знаете об архебактериях и об их свойствах, которых нет у других доядерных организмов?
4. Какова роль бактерий в природе и в жизни человека? Какие морфологические формы бактерий Вы знаете?
5. Перечислите основные свойства грибов. Чем отличаются грибы от лишайников?
6. Каковы сходства и различия между клетками растений и клетками животных?
7. Чем отличаются зеленые водоросли от цианобактерий?
8. Обладают ли водоросли какими-либо признаками, имеющими хозяйственное значение?
9. Какие свойства характерны для высших растений?
10. Что означает у растений чередование поколений и какова его биологическая роль?
11. Существуют ли различия между моховидными и папоротникооб-разными растениями? Существует ли общность в их происхождении?
12. Что вы знаете о происхождении голосеменных растений? Каково их значение в природе и в жизни человека?
13. Почему покрытосеменные имеют такое название?
14. Каково значение цветка?
16. В чем заключается двойное оплодотворение у покрытосеменных?
16. Какое значение в жизни человека имеют покрытосеменные?
17. Что вы знаете о происхождении покрытосеменных растений?
Грин Н., Стаут У.. Тейлор Д. Биология. М.: Мир. 1996. 368 стр.
Нидон К., Петерман И., Шеффель П., Шайба Б. Растения и животные. М.: Мир. 1991. 260 стр.
Старостин Б. А. Ботаника. В кн. «История биологии». М.: Наука. 1975. 52-77.
Яковлев Г. П., Челомбитько В. А. Ботаника. М.: Высшая школа. 1990. 367 стр.
Rosemweig М. L. Species Diversity in Space and Time. Cambridge University Press. 1995. 436 pp.
В соответствии с наиболее распространенной классификацией царство животных подразделяют на подцарства Одноклеточные (Monocytozoa), или Простейшие (Protozoa), и Многоклеточные (Metazoa). Многоклеточные подразделяют на животных, не имеющих настоящих органов и тканей (Parazoa), и на настоящих многоклеточных животных (Eumetazoa). Примером первых являются губки, примером вторых — все остальные многоклеточные животные (остальные типы животных). Наконец, животных подразделяют на беспозвоночных и позвоночных. К беспозвоночным относят животных, не имеющих типичного скелета, к позвоночным — животных, имеющих скелет. Важно заметить, что эти подразделения не основаны на естественной системе классификации, но они имеют практическое значение, отражая общие черты организации живого мира.
Все животные, за исключением простейших, являются многоклеточными гетеротрофными организмами, способными к движению. Для них (кроме губок) характерна координация частей тела с помощью нервной системы.
Поскольку зоологи по-разному понимают систематический ранг различных таксонов, то в настоящее время трудно назвать подлинное количество типов в каждом подцарстве. В большинстве случаев все же считают, что количество типов животных составляет не менее 35, тогда как количество классов приближается к 100.
Простейшие — это одноклеточные организмы животной природы, хотя часто к ним относят и некоторые одноклеточные формы, подобные по своим свойствам растениям и называемые фитофлагеллятами. Примером таких организмов является эвглена зеленая (Euglena viridis). В пределах этого подцарства известно свыше 25 000 видов, большинство из которых обитает в пресных и соленых водах, в почве. Около 3500 видов являются паразитами растений, животных и человека, заселяя их клетки, ткани и полости тела.
Строение простейших в принципе сходно со строением клеток многоклеточных организмов. Однако для них характерны специфические отличия, определяемые тем, что любой представитель простейших не только является клеткой, но и представляет собой самостоятельный организм.
Форма тела простейших является овальной или вытянутой, а размеры в большинстве случаев микроскопические (3-150 мкм), хотя встречаются и более крупные формы.
Типичные простейшие покрыты трехслойной мембраной, толщина которой варьирует у организмов разных видов. Каждый слой мембраны построен в основном из белков. Отдельные простейшие (раковинные корненожки) имеют наружный цитоскелет в виде раковины.
Цитоплазма дифференцирована на экто- и эндоплазму. Эктоплазма является уплотненным образованием, из которого образуется периферическая пленка, называемая пелликулой. Эндоплазма имеет более рыхлую структуру.
Для простейших характерно наличие органелл (органоидов) двух типов — общего назначения и специальных.
Органеллами общего назначения являются ядро, митохондрии, рибосомы, центриоли, комплекс Гольджи, лизосомы и др. Наиболее крупные простейшие — многоядерны, мелкие — одноядерны. Ядро окружено двойной мембраной. Количество хромосом различно у организмов разных видов и колеблется в пределах от двух (вероятно, гаплоидное число) до более чем 160. Длина хромосом составляет 1—50 мкм.
Органеллами специального назначения являются органеллы движения, а также пищеварительные и сократительные вакуоли. Органеллы движения (локомоторные структуры) представлены псевдоподиями (ложноножками), жгутиками, или ресничками. Например, движение амебоидных организмов основано на движении цитоплазмы. Эктоплазма давит на эндоплазму, в результате чего цитоплазма перетекает в другой участок тела, где образуются псевдоподии, с помощью которых организмы передвигаются в разных направлениях. Жгутики построены из скрученных фибрилл (нитей), основания которых формируют особую гранулу (базальное тельце), называемую блефаропластом (кинетосомой). У свободноживущих простейших жгутики действуют в качестве «винтов», позволяющих вращение тела вокруг оси. Реснички имеют фибриллярную структуру.
Питание простейших характеризуется значительным разнообразием в способах поглощения пищи. Одни из них воспринимают пищу из растворов всем телом посредством пиноцитоза, другие поглощают твердую пищу через цитостом (клеточный рот), третьи захватывают пищу псевдоподиями. Пища, поступающая в эндоплазму, переваривается в специализированных вакуолях, содержащих пищеварительные ферменты. Частицы пищи, остающиеся непереваренными, выбрасываются в среду вместе с пищеварительной вакуолью.
Многие водные простейшие обладают одной или более сократительной вакуолью, которая у них обеспечивает постоянство осмотического давления, а также снабжение кислородом с помощью поступающей и затем выбрасываемой воды.
Как показало культивирование простейших, они нуждаются в минеральном питании, а также в факторах роста (витамин В^д, ти-амин, биотин, рибофлавин, никотиновая кислота, пиридоксин, фо-лиевая и пантотеновая кислоты и др.). Некоторые простейшие нуждаются в стероидах.
Часть простейших, сходных с растениями, имеет хроматофоры, в которых происходит фотосинтез. Из-за содержания хлорофилла и запасных пигментов хроматофоры по цвету могут быть зелеными, желтыми, красными, коричневыми и даже голубыми. Пищевые потребности простейших с хроматофорами и без хроматофоров упрощены по сравнению с простейшими животной природы.
Энергию большинство простейших получают в результате окисления органических соединений (углеводов, жирных кислот). В отличие от бактерий они не способны к использованию неорганических веществ в качестве главного источника энергии.
Размножение простейших происходит как бесполым, так и половым путем. Бесполое размножение заключается в делении организма на две половины, причем ему предшествует деление ядра. Иногда деление тела и деление ядра происходят одновременно. Размножение половым путем осуществляется с помощью синга-мии (слияния двух гамет), конъюгации (обмена гаметическими ядрами) и аутогамии, заключающейся в формировании гаплоид-ных ядер и слияния их в синкарионы. Некоторые простейшие размножаются одновременно бесполым и половым путем. Например, в размножении малярийных плазмодиев бесполая фаза встречается, когда они находятся в организме позвоночных, половая, когда находятся в организме комаров. Установлено, что Euplotes raikovi продуцирует ферромоны полипептидной природы, которые индуцируют как половое, так и вегетативное размножение простейших этого вида.
Принципиальной особенностью простейших является прохождение ими циклов развития. Различают простые и сложные циклы развития. Простым циклом развития является такой цикл, в котором имеется лишь одна (вегетативная) стадия. Напротив, сложные циклы развития связаны с развитием простейших в разных тканях и органах, причем, принадлежащих разным организмам-хозяевам.
Простейшие способны к раздражимости, т. е. к ответным реакциям на воздействие различных факторов. В частности, одной из важнейших форм раздражимости простейших является способность к превращению их вегетативных форм при неблагоприятных условиях в цисты, что называют инцистированием. Благодаря инцистированию простейшие способны выживать в самых неблагоприятных условиях (высыхание среды, появление в ней вредных веществ, изменение температуры и др.) в течение времени, измеряемого годами. При попадании в благоприятные условия цисты развиваются в активные вегетативные формы (трофозоиты). Таким образом, инцистирование способствует распространению простейших, попаданию их в новые экологические ниши.
Большинство простейших являются свободноживущими. Известны также простейшие, которые могут обитать в организме другого вида, не принося ему ущерба, а даже помогая ему. Например, некоторые жгутиконосцы, обитающие в кишечнике термитов, могут переваривать там целлюлозу и этим обеспечивают пищевые потребности термитов, поскольку последние самостоятельно не утилизируют это соединение. Однако значительная часть простейших ведет паразитический образ жизни, причем отдельные паразиты очень опасны для человека и животных. Человек является потенциальным хозяином около 25 видов простейших, из которых 2 вида могут обитать в полости рта, 12 видов — в кишечнике, 1 — в мочеполовом тракте, около 10 — в крови и других тканях.
Простейших классифицируют на основе способов их движения (строения локомоторных органов) и особенностей размножения на пять типов, а именно: Саркомастигофоры (Sarcomastigophora), Споровики (Sporozoa), Книдоспоридии (Cnidosporidia), Микроспоридии (Microspora) и Ресничные, или Инфузории (Infusoria).
 Тип Саркомастигофоры (
Sarcomastigophora
).
Этот тип представлен наиболее примитивными организмами среди всех простейших. Общими свойствами организмов этого типа являются наличие у них одного ядра и способность образовывать в результате переливания цитоплазмы псевдоподии (ложноножки), которые служат для передвижения и захвата пищи, или жгутики. Насчитывают около 18 000 видов. Тип Саркомастигофоры (
Sarcomastigophora
).
Этот тип представлен наиболее примитивными организмами среди всех простейших. Общими свойствами организмов этого типа являются наличие у них одного ядра и способность образовывать в результате переливания цитоплазмы псевдоподии (ложноножки), которые служат для передвижения и захвата пищи, или жгутики. Насчитывают около 18 000 видов.
Саркомастигофоры являются обитателями в основном соленых (морских) вод, однако живут также и в пресной воде, во влажной почве. Многие паразитируют в организме животных и человека.
 В составе типа Саркомастигофоры классифицируют два подтипа — Саркодовые (Sarcodina) и Жгутиконосцы (Mastigophora). В составе типа Саркомастигофоры классифицируют два подтипа — Саркодовые (Sarcodina) и Жгутиконосцы (Mastigophora).
Подтип Саркодовые
(Sarcodina). Типичными представителями Саркодовых являются пресноводная амеба (Amoeba proteus, рис. 16) и дизентерийная амеба (Entamoeba histolytica). Пресноводная амеба обитает в лужах, прудах. В ее цитоплазме различают два слоя — эктоплазму и эндоплазму. Содержит одно ядро. Передвигается с помощью псевдоподий. Размножается бесполым путем (делением). Способна к инцистированию в неблагоприятных условиях. Другими известными свободноживущими Саркодовыми являются пресноводные раковинные корненожки и фораминиферы, обитающие в морях.
Дизентерийная амеба является паразитом человека. Для нее характерен сложный цикл развития. Человек заражается в результате заглатывания с загрязненными продуктами цист этого организма. Болезнь, вызываемая этим паразитом, носит название амебиаза.
Подтип Жгутиконосцы
(Mastigophora). Эти организмы обитают в морских и пресных водах или ведут паразитический образ жизни в организме растений и животных, принадлежащих к разным систематическим группам, а также в организме человека. Насчитывают около 8000 видов.
Жгутиконосцы имеют овальную, шаровидную или вытянутую форму (рис. 17). Размеры микроскопические. Тело покрыто двойной мембраной, на внешней поверхности которой имеется тонкая пелликула. В цитоплазме располагается одно или несколько сходных ядер.
Для организмов, классифицируемых в пределах этого подтипа, характерно наличие одного или более жгутиков. Каждый жгутик состоит из фибрилл и прикрепляется к базальному тельцу (блафа-ропласту или кинетосоме), располагающемуся в эктоплазме.
 Большинство органелл общего назначения (митохондрии, микросомы, комплекс Гольджи и др.) сравнимы по морфологии и функциям с такими же органеллами клеток высших растений и животных, включая сходство на субмикроскопическом уровне. Центриоли у многих жгутиковых играют важную роль в организации не только митотического аппарата, но и являются локусом, вокруг которого формируются набор органелл, образованных в основном фибриллярными белками (жгутик, аксостиль и др.). Среди организмов этого подтипа различают свободноживущих (Е. viridis) и паразитов, для которых характерны сложные циклы развития. Наиболее известными паразитами человека являются три-паносомы (Trypanosoma brucei gambiense и Т. brucei rhodesiense), которые вызывают трипаносомоз (африканскую сонную болезнь), лейшмании (Leishmania tropica major, L. tropica minor и L. donovani), которые вызывают лейшманиоз, трихомонады (Trichomonas vaginalis), вызывающие трихомоноз, и лямблии (Lamblia intestinalis), вызывающие лямблиоз. Большинство органелл общего назначения (митохондрии, микросомы, комплекс Гольджи и др.) сравнимы по морфологии и функциям с такими же органеллами клеток высших растений и животных, включая сходство на субмикроскопическом уровне. Центриоли у многих жгутиковых играют важную роль в организации не только митотического аппарата, но и являются локусом, вокруг которого формируются набор органелл, образованных в основном фибриллярными белками (жгутик, аксостиль и др.). Среди организмов этого подтипа различают свободноживущих (Е. viridis) и паразитов, для которых характерны сложные циклы развития. Наиболее известными паразитами человека являются три-паносомы (Trypanosoma brucei gambiense и Т. brucei rhodesiense), которые вызывают трипаносомоз (африканскую сонную болезнь), лейшмании (Leishmania tropica major, L. tropica minor и L. donovani), которые вызывают лейшманиоз, трихомонады (Trichomonas vaginalis), вызывающие трихомоноз, и лямблии (Lamblia intestinalis), вызывающие лямблиоз.
Саркомастигофоры представляют собой группу древнейших орга-низмов-эукариотов.
Тип Споровики (
Sporozoa
).
Организмы этого типа характеризуются простотой организации. Они не имеют органоидов пищеварения и выделения. Питание, дыхание и выделение обеспечиваются всей поверхностью их тела. Все споровики являются внутриклеточными паразитами. Насчитывают около 20 000 видов. Все они являются паразитами животных многих систематических групп. Среди них наиболее известными паразитами являются возбудители малярии (Plasmodium falciparum, P. vivax, P. ovale, P. malariae) и токсоплазмоза (Toxoplasma gondii).
Особенностью споровиков является то, что для них характерны сложные циклы развития, происхождение которых связано с образованием спор и сменой хозяев. Например, у малярийных плазмодиев жизненный цикл связан с чередованием бесполого и полового размножения (рис. 18). В организме человека плазмодии размножаются бесполым путем с образованием промежуточных форм в виде мерозоитов, шизонтов и гамонтов. Лихорадка и повышение температуры связаны с тем, что, размножаясь в эритроцитах, шизонты разрушают их. Время генерации паразита (шизогонии) для Р. falciparum, P. vivax, Р. ovale составляет 48 часов, а для Р. malariae — 72 часа, в результате чего в первом случае у больных имеет место 2-дневная, а во втором — 3-дневная лихорадка.
Половое размножение малярийных плазмодиев происходит в организме комара и связано с развитием там гамонтов и превращением последних в гаметы, которые сливаются и дают начало диплоидным зиготам. Когда вокруг зигот появляется мембрана, то это сопровождается их инцистированием, т. е. образованием ооцист (ооки-нет). Ооцисты подвергаются затем мейотическому делению, т. е. спорогонии, в результате которой внутри ооцист формируются гаплоид-ные спорозоиты. Являясь инвазионной формой, последние при питании комара на коже человека попадают в его кровь и цикл развития начинается сначала.
Возбудитель токсоплазмоза (Т. gondii), которым болеют человек и животные, также характеризуется сложным циклом развития. Часть цикла токсоплазм проходит в организме кошки, часть — в организме другого млекопитающего, а — часть в организме человека. Человек заражается ооцистами возбудителя при разделке туш животных, а цистами — при употреблении в пищу недостаточно прожаренного или проваренного мяса.
Эволюция споровиков не совсем понятна. Тем не менее предполагают, что они берут начало от жгутиконосцев.
Тип Книдоспоридии (
Cnidosporidia
).
Этот тип представлен более 1000 видами, представители которых распространены в пресных и соленых водоемах.
Все книдоспоридии ведут паразитический образ жизни, поражая исключительно пресноводных и морских рыб и вызывая у них миксоспоридиозы. Миксоспоридиоз лососевых (форель, лосось) известен под названием «вертежа».
Предполагают, что книдоспоридии произошли от саркодовых.
Тип Микроспоридии (
Microsporidia
).
Организмы этого типа являются паразитами как позвоночных, так и беспозвоночных. Насчитывают свыше 300 видов.
Микроспоридии ведут внутриклеточный паразитический образ жизни. Наиболее известными микроспоридиями являются Nosema apis и Nosema bombycis, которые вызывают белый понос у медоносных пчел и пебрину у тутового шелкопряда (соответственно). Микроспоридии вызывают также болезни рыб (корюшки, снетка).
Предполагают, что микроспоридии также произошли от сарко-довых.
Тип Инфузории (
Infusoria
).
В составе этого типа насчитывают около 6000 видов. Являются обитателями очень многих экологических ниш.
Различают свободноживущих и паразитирующих инфузорий. Наиболее известным свободноживущим организмом является парамеция, или туфелька (Paramecium caudatum). Форма тела этого организма вытянута, но постоянна (рис. 19), т. к. снаружи окружена плотной пелликулой, а внутри (в экто- и эндоплазме) имеются к тому же и выполняющие роль опорного скелета нити. Имеются также два ядра (макронуклеус и микронуклеус). Реснички, покрывающие тело, имеют базальные гранулы. Для парамеций характерна цитологическая специализация (наличие предротово-го углубления — перистома клеточного рта, глотки, ануса и трихоциста). Размножаются как бесполым (деление), так и половым путем в виде конъюгации (рис. 20).
 Патогенным представителем этого типа является балантидий (Balantidium coli), вызывающий у человека балантидиоз. Человек инвазируется цистами баланти-дия в результате приема зараженной им пищи и воды, а также пользования загрязненными предметами. Патогенным представителем этого типа является балантидий (Balantidium coli), вызывающий у человека балантидиоз. Человек инвазируется цистами баланти-дия в результате приема зараженной им пищи и воды, а также пользования загрязненными предметами.
 Известны инфузории, часть цикла которых является паразитической, часть — свободно-живущей. Такой инфузорией является ихтиафтириус, паразитирующий в организме рыб. Известны инфузории, часть цикла которых является паразитической, часть — свободно-живущей. Такой инфузорией является ихтиафтириус, паразитирующий в организме рыб.
Инфузории — наиболее организованные существа среди простейших. Они являются также процветающей группой среди животных этого подцарства.
Простейшие являются древнейшими обитателями Земли. Предполагая родство между Capкодовыми и Жгутиковыми, многие протистологи считают, что архетипами современных простейших являются Саркодовые.
Однако в объяснениях эволюции простейших много противоречий, поскольку существует мнение и о том, что саркодовые и жгутиконосцы произошли от разных предков, каких-то самых древних живых существ. Большое значение придается ароморфозам, примером которых является развитие ресничного и ядерного аппаратов у инфузорий, и идиоадаптациям в виде приспособлений к планктонному образу жизни или инцистированию у ряда простейших. В любом случае считают, что самой прогрессивной формой в мире простейших являются инфузории.
Тип Губки (
Spongia
).
Этот тип представлен наиболее примитивными многоклеточными организмами, клетки которых, однако, дифференцированы (рис. 21). Известно около 3000 видов губок. Будучи обитателями морей, ведут неподвижный образ жизни на дне или на различных подводных предметах.
Губки — это колониальные организмы. Для них характерны разнообразная форма, мягкое и эластичное тело, пронизанное порами, в которые поступает вода, различная окраска. Имеют внутренний скелет (известковый, кремневый или роговой). Имеют также центральную (гастральную) полость.
Для дыхания используют растворенный в воде кислород. Питаются органическими остатками растений и животных, а также простейшими и бактериями. Новые исследования показали, что морские губки способны поглощать и растворять мелкие кварцевые частицы, что связывают с потребностью губок в аскорбиновой кислоте. Эта способность является уникальной в мире животных.
Для губок характерны бесполое размножение (почкование), а также половая дифференциация и половое размножение.
Отдельные виды губок ядовиты для человека. Известны губки, используемые в хозяйственной деятельности человека (туалетные губки, бодяги, стеклянные губки).
Происхождение губок не совсем ясно. Предполагают, что они являются слепой ветвью эволюции.
Тип Кишечнополостные (
Coelenterata
).
Организмы этого типа являются обитателями в основном морей, но они проникли и в пресные воды. Известно около 9000 видов. Кишечнополостным присуща довольно простая организация. Для них характерна радиально-осевая симметрия. Их тело состоит из экто- и энтодермы, между которыми находится мезоглея, представляющая собой слой неклеточного вещества.
Типичными представителями, обитающими в морских водах, являются коралловые полипы (рис. 22), ведущие неподвижный образ жизни, образуя часто колонии, и медузы, одиночные организмы, ведущие подвижный образ жизни (плавающие). Наиболее известным пресноводным представителем кишечнополостных является гидра.
Размножаются бесполым и половым путем.
Кишечнополостные склонны к образованию симбиотических отношений в форме комменсализма, мутуализма, паразитизма с другими морскими организмами.
Кишечнополостные имеют важное значение в природе. Они являются участниками многих морских экологических систем. Образуя колонии, коралловые полипы формируют рифы, коралловые острова.
Кишечнополостные — самые древние многоклеточные животные, процветающие в современную эпоху. Они встречались еще в докембрии. Входят в основной ствол эволюции. Предполагают, что их предками были когда-то примитивные многоклеточные животные, для которых характерен был свободноплавающий образ жизни.
Тип Плоские черви (
Plathelminthes
).
К этому типу относят животных, характеризующихся вытянутой уплощенной билатерально-симметричной формой и обитающих в воде, почве, организме растений, животных и человека. Они составляют один из наиболее больших по численности типов животных (около 9000 видов).
Размеры плоских червей в длину чрезвычайно варьируются — от 0,5 мм до 30 м. У одних плоских червей поверхностный (покровный) эпителий снабжен ресничками или даже жгутиками, у других он характеризуется тем, что ядра и цитоплазма смещаются в глубь тела, формируя так называемый погружной эпителий (тегумент). Покровный эпителий и располагающаяся под ним кожная мускулатура формируют кожно-мускульный мешок. Пространство между мешком и внутренними органами заполнено паренхимой, являющейся производной мезодермы. Пищеварительная система у некоторых плоских червей представлена ротовым отверстием, глоткой, пищеводом и кишкой, а у некоторых (у ленточных) ее совсем нет. Все плоские черви гермафродиты.
Плоские черви являются первичноротыми животными. Они характеризуются также отсутствием метамерии, скелетной, сосудистой и дыхательной систем.
Среди гельминтов, принадлежащих к этому типу, широко распространен паразитизм. В пределах типа прослеживаются группы (таксоны), представители которых обладают чертами постепенного перехода от свободноживущих к паразитическим формам. Это проявляется в том, что органы, которые наиболее активно функционируют у свободноживущих, подвергаются прогрессивной редукции вплоть до полного исчезновения у паразитов. В частности, у паразитов отмечается инактивация и атрофия сенсорных и локомоторных органов. С развитием в условиях паразитизма способности к адсорбции пищи поверхностью тела подвергается регрессии али-ментарный тракт. В связи с этим у ленточных гельминтов уже нет следов пищеварительной системы.
Общий уровень метаболизма у паразитических видов существенно не изменен. Однако, живя в кишечнике животных или человека, где количество кислорода ограничено, многие плоские черви развили анаэробный тип дыхания, при котором кислород они получают путем восстановления углеводов (по типу брожения).
Нервная система характеризуется по сравнению с кишечнопо-лостными значительным усложнением. У ленточных гельминтов в головной части тела имеется парный мозговой ганглий или окологлоточное нервное кольцо и отходящие от них нервные стволы, сформированные нервными клетками.
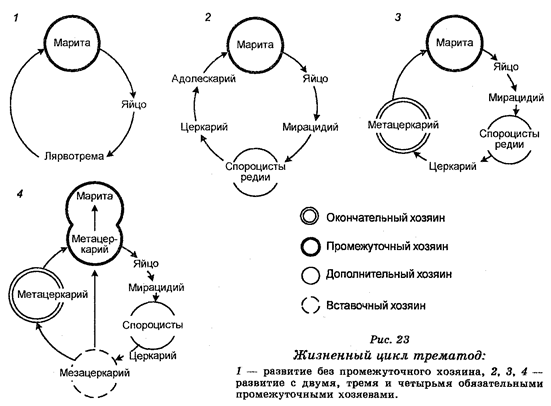
У паразитических форм неподвижный образ жизни и обилие пищи нашли отражение в их гигантской репродуктивной способности, которая компенсирует потери при переходе паразита от одного хозяина к другому.
Для плоских червей характерно развитие с неполным метаморфозом. Их личиночные стадии являются мелкими организмами, не способными к восприятию пищи, а также с ограниченной локомоторной способностью и временем жизни. У некоторых видов лишь один экземпляр на 1 млн особей является удачливым в переживании. Поэтому переживание паразитических видов обеспечивается, возможно, лишь их гигантской репродуктивной способностью.
Тип делят на три класса: Ресничные, или Турбеллярии (Turbellaria), Сосальщики, или Трематоды (Trematoda) и Ленточные (Cestoda).
Ресничные
, или Турбеллярии
(Turbellaria) являются наиболее примитивными плоскими гельминтами. Их размеры в длину составляют около 5 мм. Известно примерно 3000 видов турбеллярий. Они являются обитателями в основном пресных и солоноватых вод. Ведут свободный образ жизни. Наиболее известным представителем животных этого класса является планария. Однако многие турбеллярий паразитируют в организме морских ракообразных, рыб и других животных.
Турбеллярии не имеют ни хозяйственного, ни какого-либо другого выраженного значения, если не принимать во внимание их использования в качестве экспериментальной модели в некоторых медико-биологических исследованиях.
Трематоды
(Trematoda) представлены примерно 6000 видов, все из которых являются паразитами животных и человека. Для трематод характерно непрямое развитие, причем их жизненные циклы довольно сложны и связаны со сменой хозяев. По этой причине их называют биогельминтами. Циклы развития связаны с циркуляцией в организме постоянных хозяев (дефинитивных, окончательных) мариты (взрослой формы) и в организме промежуточных хозяев личиночных форм (рис. 23). Дефинитивными хозяевами трематод являются человек и другие животные.
 В качестве патогенных видов этого класса являются печеночный, кошачий и ланцетовидный сосальщики, клонорхис (Fasciola hepatica, Opisthorchorchis felineus, Dicrocoelium lanceatum, Clonorchis sinensis соответственно) и другие. В качестве патогенных видов этого класса являются печеночный, кошачий и ланцетовидный сосальщики, клонорхис (Fasciola hepatica, Opisthorchorchis felineus, Dicrocoelium lanceatum, Clonorchis sinensis соответственно) и другие.
Ленточные (Cestoda) черви являются паразитами млекопитающих и человека. В этом классе насчитывают более 3000 видов гельминтов.
 Наиболее известными видами являются цепень вооруженный (Taenia solium рис. 24), цепень невооруженный (Taeniarhynchus saginatis), эхинококк (Echinococcus granulosus), лентец широкий (Diphyllobothrium latum) и др. Наиболее известными видами являются цепень вооруженный (Taenia solium рис. 24), цепень невооруженный (Taeniarhynchus saginatis), эхинококк (Echinococcus granulosus), лентец широкий (Diphyllobothrium latum) и др.
Ленточные
гельминты также являются биогельминтами, т. к. их жизненные циклы проходят со сменой разных хозяев(рис. 25).
Предполагают, что плоские черви произошли либо от древних кишечно-полостных, либо от предков, общих с древними кишечнополостными. Что касается происхождения червей, принадлежащих к разным классам, то предполагают, что они возникли в результате дивергенции предковых форм и адаптации образовавшихся форм к разным экологическим нишам. В частности, ресничные черви произошли либо от гребневиков, либо от бескишечных предковых форм, перешедших к ползанию. Сосальщики произошли от ресничных червей, перешедших к паразитическому образу жизни. В соответствии с одной из гипотез ленточные черви произошли тоже от ресничных, но в процессе паразитизма у них развились органы фиксации, подверглась интенсивному развитию половая система. По другой гипотезе ленточные черви произошли от древних трематод.
Тип Круглые черви (
Nema
-
thelminthes
).
Круглые черви характеризуются удлиненным цилиндрическим телом, не имеющим сегментации и ресничек на поверхности. Насчитывают более 10 000 видов этих организмов. Они адаптированы к жизни почти во всех экологических нишах, многие из них являются паразитами растений, животных и человека.
Круглые черви характеризуются трехслойностью, билатеральной симметрией, наличием первичной полости тела и кожно-мус-кульного мешка. Помимо круглой формы, типичным для гельминтов этого типа является наличие мышечной, пищеварительной, выделительной, нервной и половой систем, для которых характерны дальнейшие прогрессивные изменения. Например, у этих червей отмечается раздельнополость, а для свободноживущих форм характерно очень большое разнообразие в половом размножении. Для пищеварительной системы характерно наличие анального отверстия. Кровеносная и дыхательная системы отсутствуют.
Круглые черви доступны для культивирования в лабораторных условиях.
Среди круглых гельминтов встречаются как свободноживущие, так и паразитические формы. Многие паразитические формы являются геогельминтами, т. к. развиваются обычно в одном хозяине. Известны среди них также и биогельминты.
Тип Круглые черви классифицируют на несколько классов, из которых наибольшее распространение и значение имеет класс Собственно круглые черви (Nematoda).
Среди паразитических нематод наиболее известными являются аскарида человеческая (Ascaris lumbricoides, рис. 26), угрица кишечная (Strongyloides stercoralis), кривоголовка (Ancylostoma duodenale), трихинелла (Trichinella spiralis) и другие, которые вызывают у человека аскаридоз, стронгилоидоз, анкилостоматоз, трихинеллез (соответственно).
Происхождение круглых червей до конца не выяснено. Тем не менее многие гельминтологи считают, что они произошли от тур-беллярий и развились в обособленную эволюционную ветвь филогенетического древа, животного мира.
В эволюции круглых червей важную роль играли ароморфо-зы, в частности, большое значение имело появление первичной полости тела, образованной по принципу «трубка в трубке» с отверстиями на обоих концах. Ароморфозами являются также прогрессивное развитие пищеварительной системы, появление раздельнополости и живорождения (у некоторых видов). Предполагают, что в эволюции этих организмов существенную роль играли гены кластера Нох, которые аналогичны генам Нох членистоногих.
Тип Кольчатые черви (
Annelides
).
Кольчатые черви, или кольчецы — это высокоорганизованные гельминты, являющиеся обитателями пресных и морских водоемов, почвы и других сред. Насчитывают около 10 000 видов этих животных. Их длина составляет от долей миллиметра до 2,5 м. Характеризуются прежде всего метамерией, заключающейся в разделении их тела внутренними перегородками на сегменты, которые функционально практически самостоятельны. Тело дифференцировано на головную часть, среднюю часть и анальный район.
Для кольчецов характерны билатеральная симметрия, трехслой-ность, вторичная (целомическая) полость тела, кожно-мускульный мешок, наличие пищеварительной, дыхательной, кровеносной, нервной и половой систем, а также органов движения (параподий) в каждом сегменте. Кольчатые черви построены таким образом, что внешняя трубка, или стенка их тела, отделена от внутренней трубки пищеварительного канала целомическим пространством.
Кольчатые черви имеют значение в круговороте веществ в природе, служат пищей рыб, в том числе и промысловых. Земляные черви способствуют рыхлению почвы и обогащению ее органическими веществами. Они улучшают также водный и газовый баланс почвы.
В медицинской практике значение имеет обитающая в неглубоких пресных водоемах медицинская пиявка (Hirudo medicinalis), которую давно используют в лечении гипертонии, повышенной свертываемости крови, тромбозов и других болезней.
Представления о происхождении кольчатых червей противоречивы. Одни гельминтологи предполагают, что они произошли от турбеллярий, другие — от форм, родственных с гребневиками. В любом случае можно считать, что происхождение кольчецов связано с крупными ароморфозами в виде появления метамерии, пе-лома, кровеносной и дыхательной систем, усложнения выделительной и нервной систем. Предполагают, что пиявки произошли от малощетинковых червей или от их предков.
 Кольчатые черви связаны филогенетически с членистоногими и мягкотелыми, поэтому они являются узлом в родословном древе животного мира. Кольчатые черви связаны филогенетически с членистоногими и мягкотелыми, поэтому они являются узлом в родословном древе животного мира.
Тип Членистоногие (
Arthropoda
).
Членистоногие — это высший тип беспозвоночных животных. В его состав входит около 650 000 видов, основная часть которых приходится на насекомых. По количеству видов он является самым процветающим типом в мире животных. Членистоногие распространены во всех средах обитания, являясь существенным компонентом всех экологических систем.
Для членистоногих характерна билатеральная симметрия. Как и кольчатые гельминты, они имеют вторичную полость тела. У большинства членистоногих тело сегментировано на три отдела — голову, грудь, брюшко. В голове сконцентрированы сенсорные органы и внешние нервные центры. К голове часто примыкают грудные сегменты, в результате чего образуется головогрудь. Имеются членистые конечности, выполняющие роль органов передвижения, чувств, нападения и т. д.
Для членистоногих характерны также трехслойность, наличие внешнего скелета в виде хитинизированной кутикулы, а также пищеварительной, дыхательной, выделительной, кровеносной, нервной, эндокринной и половой систем, характеризующихся дальнейшим усовершенствованием.
Пищеварительная система разделена на передний, средний и задний отделы. В среднем отделе расположены пищеварительные железы (зачатки). Ротовой аппарат составлен передними конечностями (тремя парами).
Дыхательная система у водных членистоногих представлена жабрами, с помощью которых используется кислород, растворенный в воде, а у наземных организмов — легочными мешками или трахеями (паукообразных и трахейнодышащих), позволяющими использовать кислород атмосферы.
Кровеносная система состоит из дорсального сосуда, представляющего собой сердце и аорту. Она незамкнута. Сердце построено из мышечных клеток. Аналогом крови является гемолимфа, которая в основном выполняет транспортную функцию, а также обеспечивает хранение воды и поддержание пищевых запасов.
Выделительная система характеризуется значительным разнообразием, будучи представленной видоизмененными метанефридиями.
Нервная система характеризуется повышающейся концентрацией ганглиев (по сравнению с кольчатыми червями) и состоит из парного надглоточного и подглоточного ганглиев, а также из брюшной нервной цепочки, составленной парными ганглиями каждого сегмента. Нервная система обеспечивает сложные двигательные и поведенческие реакции.
  Половая система и репродуктивный процесс членистоногих характеризуются такж,е большим разнообразием. Большинство членистоногих раздельнополо, но встречаются и гермафродиты, главным образом среди ракообразных. Среди насекомых довольно обычен партеногенез, хотя нормальная половая репродукция в определенные интервалы встречается даже среди партеногенетических видов. У большинства членистоногих мужские половые клетки переносятся в полость брюшка самок, где позднее они оплодотворяют яйцеклетки. Организмы многих видов развиваются с метаморфозами. Половая система и репродуктивный процесс членистоногих характеризуются такж,е большим разнообразием. Большинство членистоногих раздельнополо, но встречаются и гермафродиты, главным образом среди ракообразных. Среди насекомых довольно обычен партеногенез, хотя нормальная половая репродукция в определенные интервалы встречается даже среди партеногенетических видов. У большинства членистоногих мужские половые клетки переносятся в полость брюшка самок, где позднее они оплодотворяют яйцеклетки. Организмы многих видов развиваются с метаморфозами.
Рост членистоногих характеризуется периодичностью, т. к. происходит во время линек, каждая из которых контролируется гормонально.
Классификация членистоногих очень сложна, поскольку среди них существует много филогенетических линий. Однако в рамках одной из классификаций их классифицируют на три подтипа: Жаберноды-шащие (Branchiata) с классом Ракообразные (Crustacea), Хелицеровые (Chelicerata) с классами Меристомовые и Паукообразные (Arachnoidea) и Трахейнодышащие (Tracheata) с классами Многоножки (Myriapoda) и Насекомые (Insecta).
Наиболее известным представителем жабернодышащих является речной рак. Представителями хелицеровых являются фаланги, скорпионы (рис. 27), пауки, клещи, трахейнодышащих — насекомые, включая блох, клопов, вшей (рис. 28), двукрылых (рис. 29) и других организмов.
Значение членистоногих в природе исключительно велико. Они являются компонентами всех экологических систем. Многие из них играют очень важную роль в патологии человека, являясь паразитами, переносчиками или хозяевами паразитов (возбудителей болезней), а некоторые являются ядовитыми животными, многие известны в качестве вредителей полей, садов и огородов.
Считают, что членистоногие возникли в докембрии и берут начало от примитивных кольчатых червей, причем в их эволюции существенное значение имели ароморфозы, которые привели к развитию гетерономной метамерии, плотных хитинизированных покровов, членистого строения конечностей, крыльев у насекомых, мышечной системы, сердца, внутреннего оплодотворения, более совершенной нервной системы. Как показали современные исследования, развитие этих свойств происходит под контролем кластера генов Нох, которые идентифицированы у всех исследованных к настоящему времени членистоногих. Поэтому предполагают, что гены Нох были вовлечены в эволюцию членистоногих.
Предполагают также, что важное значение в эволюции членистоногих имело развитие плотного наружного скелета, предупреждающего высыхание животных. Кроме того, развитие членистых конечностей обеспечило устойчивость животных к обитанию в разных средах. Наконец, эволюция лимитировала размеры наземных членистоногих, а естественный отбор закрепил в качестве абсолютно необходимой линьку, обеспечивающую изменение форм и размеров животных, замену их покровов (кутикулы) в процессе развития.
Считают, что прародиной жаберно дышащих является море, а хелицеровых — мелководье морей. Трахейнодышащие возникли на суше.
Тип Мягкотелые (
Mollusca
). Мягкотелые являются вторым по численности типом после членистоногих (около 80 000 видов). Большинство этих животных обитают в соленых и пресных
Моллюски чрезвычайно полиморфны. Размеры тела взрослых форм колеблются от 1 мм  до 17 метров. Для них характерна билатеральная симметрия, но смещение органов очень часто искажает ее. Их тело сегментировано, не имеет конечностей и покрыто раковиной. Характерно разнообразие форм раковин (рис. 30) до 17 метров. Для них характерна билатеральная симметрия, но смещение органов очень часто искажает ее. Их тело сегментировано, не имеет конечностей и покрыто раковиной. Характерно разнообразие форм раковин (рис. 30)
У некоторых видов раковина распространяется лишь на часть тела, которое покрыто кожной складкой — мантией. Полость тела представлена в основном гемоцелью. Мягкотелые обладают всеми основными системами органов, формирующимися из экто-, эндо- и мезодермы.
Пищеварительная система представлена извитой трубкой. Дыхательная система представлена кожны- ми жабрами, которые расположены под кож-ной мантией. Кровеносная система не замкнута. Сердце состоит из желудочка и одного-двух предсердий. Кровь содержит гемоцианин, иногда гемоглобин.
Нервная система представлена 5 парами ганглиев, что делает ее еще примитивней.
Многие моллюски раздельнополые и яйцек ладущие, но встречаются виды-гермафродиты,
В составе типа Моллюски классифицируют пять классов.
Некоторые из этих животных имеют промысловое значение (устрицы, мидии, морские гребешки, осьминоги и другие). Отдельные представители этого типа имеют медицинское значение, т. к. являются промежуточными хозяевами патогенных трематод (ланцетовидного сосальщика, фас-циолы, описторхиса, шистосом).
Палеонтологические данные свидетельствуют о том, что самые ранние моллюски уже существовали в каменноугольном периоде. Считают, что моллюски родственны кольчатым червям и берут начало от предков, общих с членистоногими.
Тип Иглокожие (
Echinodermata
).
Иглокожие представлены 6000 видов животных, являющихся обитателями морей и океанов. Среди них наиболее известными являются морские ежи, морские звезды, голотурии и др. (рис. 31).
Для иглокожих характерны радиальная симметрия (личинки — билатерально симметричны), трехслойность, образование вторичного рта в период зародышевого развития, вследствие чего их называют вторичноротыми. Для них характерны также выраженный целом, наличие в коже соединительной ткани и эпидермиса, скелет мезодермального происхождения. Исключительно важным признаком этих животных является наличие у них так называемой амбулакральной системы, которая представляет собой полости, наполненные жидкостью и используемые для движения.
Кожный покров иглокожих состоит из слоя соединительной ткани и слоя эпидермиса, скелет — из известковых пластинок.
Пищеварительная система представлена кишечной трубкой. В переваривании пищи принимают участие ферменты.
Дыхательная система представлена кожными жабрами. Кровеносная система не замкнута. «Кровью» является морская вода, с помощью которой транспортируются питательные вещества в разные участки тела.
Нервная система очень примитивна, ганглии отсутствуют.
Развитие происходит с метаморфозами. В противоположность всем беспозвоночным, которые являются первичноротыми организмами (Protostomia), иглокожие являются вторичноротыми (Deutorostomia) животными, поскольку первичный рот (бластопор) в стадии гастру-лы превращается в анальное отверстие, а новый (вторичный) рот образуется на брюшной стороне.
Иглокожих классифицируют на несколько классов.
Иглокожие не имеют существенного хозяйственного интереса, хотя некоторых из них используют в пищу (трепанги, мидии и другие).
Иглокожие — очень древние животные. Предполагают, что они родственны хордовым и произошли от древнейших вторичноротых форм, родственных с гребневиками, но в своем развитии остановились на начальных этапах эволюции, вторично приобретя радиальную симметрию.
Тип Хордовые (
Chordata
).
Хордовые являются главным типом животного царства. В этом типе насчитывают более 42 000 видов, обитающих в разных средах.
Для хордовых характерен ряд специфических свойств. В частности, для них характерно наличие хорды (нотохорды), представляющей собой гибкую структуру стержневидной формы, тянущуюся вдоль тела в положении, занимаемом у позвоночных позвоночником.
Хорда является осевым скелетом. Состоя из вакуолизирован-ных клеток, она сохраняется всю жизнь лишь у низших организмов, но у высших в ходе развития замещается позвонками.
Для хордовых характерно также наличие нервного пучка в виде трубки над хордой и пищеварительной трубки под хордой. Далее, для них характерно наличие в зародышевом состоянии или в течение всей жизни многочисленных жаберных щелей, открывающихся наружу из глоточного района пищеварительной трубки и являющихся органами дыхания. Наконец, для них характерно расположение сердца или замещающего его сосуда на брюшной стороне.
Наряду со специфическими чертами для хордовых характерны билатеральная симметрия, трехслойность, вторичная полость тела. Как и полухордовые, хордовые являются вторичноротыми организмами.
Хордовых классифицируют на подтипы Бесчерепные (Acrania) и Черепные (Craniata), или Позвоночные (Vertebrata).
Бесчерепные представлены классом Ланцетники (Amphioxi). Эти животные являются обитателями прибрежных участков морей. Наиболее известный представитель этого подтипа — ланцетник (Branchiostoma lanceolatum). Размеры его составляют несколько сантиметров в длину (рис. 32).
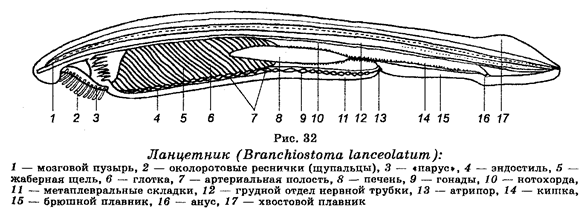
У ланцетников отсутствуют выраженные голова, мозг, челюсти, сердце. Осевой скелет у этих животных представлен хордой. Способны к свободной локомоции, раздельнополы. Оплодотворение происходит в воде.
Хозяйственного значения не имеют.
В эволюционном плане ланцетники представляют вторую стадию в эволюции хордовых, которая предшествовала эволюции позвоночных.
Позвоночные
(Vertebrata) имеют очень совершенные покровы тела в виде кожи, состоящей из двух слоев (многослойного эпидермиса и кориума), производными которой являются чешуя, перья, волосы, когти и ногти.
У позвоночных форм она заменена позвоночником. Во взрослом состоянии большинство позвоночных имеет костный скелет. Хорошо развиты голова, глаза, конечности.
Мышечная система дифференцируется на поперечно-полосатую и гладкую мускулатуру.
Пищеварительная система очень сложна. Кишечник разделен на три отдела. Имеются печень и поджелудочная железа.
Дыхательная система представлена жабрами или легкими.
Кровеносная система замкнута, представлена многокамерным сердцем, сосудами в виде артерий и вен. Клеточный состав крови очень дифференцирован. Имеется незамкнутая лимфатическая система.
Выделительная система очень совершенна. Она представлена парными почками и мочеточниками.
Характерной особенностью нервной системы, даже у низших позвоночных, является развитый мозг, а у высших форм — наличие мозговых полушарий. Нервная система подразделяется на центральную и периферическую. Развиты специализированные органы чувств (нос, глаза, уши). Развиты также железы внутренней секреции.
Для позвоночных характерны раздельнополость (кроме некоторых видов круглоротых), выраженный половой диморфизм. Мужские и женские половые железы парные, осеменение наружное или внутреннее. Для некоторых позвоночных характерно развитие с метаморфозом.
В пределах подтипа отмечается гигантское структурное и функциональное разнообразие организмов, что осложняет их классификацию и исключает пока единое мнение о количестве классов в этом подтипе. Однако наиболее распространенной является классификация, в которой выделяют классы Круглоротые (Cyclostomata), Хрящевые рыбы (Chonrichthyes), Костные рыбы (Osteichthyes), Земноводные, или Амфибии (Amphibia), Пресмыкающиеся, или Рептилии (Reptilia), Птицы (Aves) и млекопитающие, или Звери (Mammalia).
Поскольку у позвоночных тоже найдены гены кластеров Нох (4 кластера по 9-11 генов), то считают, что дубликация этих генов у примитивных хордовых, а затем у ранних позвоночных позволила эволюцию в направлении общего плана строения и сходных систем органов.
Класс Круглоротые
(Cyclostomata) представлены примитивными животными, обитающими в морских и пресных водах умеренных широт обоих полушарий. Насчитывают свыше 40 видов. Наиболее известными организмами этого класса являются миноги и миксины. Типичным представителем этого класса является речная минога (Lampetra fluviatilis).
Имеют промысловое значение.
Отсутствие у миног и миксин челюстей и парных конечностей позволяет считать их низшими позвоночными. Современные круг-лоротые являются потомками очень древней группы позвоночных животных, называемых бесчелюстными и произошедших от ископаемых остракодерм, живших 500 млн лет назад и представлявших собой первых позвоночных.
Предполагают, что острокодермы представляют собой предко-вую форму всех позвоночных и что хрящевой скелет Круглоротых возник в результате дегенерации костного скелета в ходе эволюции их предков.
 Класс Хрящевые рыбы
(Chondichthyes) представлен обитателями в основном морей и океанов. Насчитывают около 730 видов этих рыб. Наиболее известными представителями этого класса являются акулы и скаты (рис. 33). Класс Хрящевые рыбы
(Chondichthyes) представлен обитателями в основном морей и океанов. Насчитывают около 730 видов этих рыб. Наиболее известными представителями этого класса являются акулы и скаты (рис. 33).
Размеры хрящевых рыб составляют от нескольких миллиметров до нескольких метров в длину. Характерными особенностями их являются хрящевой скелет, кожа, покрытая плакоидными (зубовидными) чешуями, парные конечности — плавники, зубы, покрытые эмалью, 5-7 пар наружных жаберных щелей (у пластино-жаберных), отсутствие кожных костей и плавательного пузыря. Для большинства хрящевых рыб характерны также поперечный рот и головной мозг прогрессивного строения.
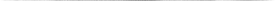 У этих животных размножение происходит путем откладки яиц, яйце-живорождения и живорождения. У этих животных размножение происходит путем откладки яиц, яйце-живорождения и живорождения.
Хрящевые рыбы имеют некоторое хозяйственное значение. Отдельные виды являются ядовитыми для человека.
Первые хрящевые рыбы обитали в древних морях уже около 300 млн лет назад, современные формы появились около 150 млн лет назад. Предполагают, что они представляют собой боковую филогенетическую ветвь.
Класс Костные рыбы
(Osteichthyes) в видовом составе довольно многочисленны (около 1500 видов). Являясь обитателями морских и пресных вод (рис. 34), они отличаются от хрящевых рыб тем, что имеют внутренний костный скелет, головные кости (черепную коробку, в которой размещен мозг), покров из костных чешуи незубовидного типа, плавательный пузырь (или легкое). Тело расчленено на голову, туловище и хвост. Внешние покровы представляют собой кожу.
Размеры костных рыб составляют от нескольких миллиметров до нескольких метров в длину.
Мускулатура имеет сегментарное строение. Развиты специализированные мышцы, приводящие в движение челюсти, глаза и другие органы.
Пищеварительная система представлена ротовым отверстием, ведущим в ротовую полость, пищеводом, желудком и кишкой, заканчивающейся задним проходом. Имеются печень с желчным пузырем и поджелудочная железа (слаборазвитая).
Плавательный пузырь заполнен газовой смесью (кислород, углекислота и азот) и служит гидростатическим аппаратом. Дыхательная система представлена жаберным аппаратом. Кровеносная система состоит из одного круга кровообращения (как у круглоротых). Двухкамерное сердце расположено в передней части полости тела, состоя из предсердия и желудочка. Выделительная система представлена первичными почками. Нервная система характеризуется более прогресссивным развитием по сравнению с круглоротыми, т. к. полушария переднего мозга более развиты. В среднем мозгу есть изгиб, характерный для всех позвоночных. Центры чувств расположены в разных отделах мозга. Из головного мозга выходит 10 пар нервов, начинающихся спинным и брюшным корешками. Вегетативная нервная система представлена двигательными волокнами блуждающего нерва. Она инНервирует все внутренние органы.
Костные рыбы раздельнополы. Оплодотворение внешнее, у некоторых видов внутреннее.
Костные рыбы имеют огромное хозяйственное значение, являясь источником продовольствия, иногда кормом для домашних животных.
Важно отметить, что рыбы некоторых видов служат промежуточными хозяевами для гельминтов, являющихся возбудителями описторхоза, дифиллоботриоза, а также сырьем для получения лекарств (рыбий жир), или являются ядовитыми животными, представляющими опасность для здоровья человека.
Костных рыб классифицируют на подклассы Лопастеперые (Sarcopterygii) и Лучеперые (Actinopterygii).
Предполагают, что костные рыбы произошли около 500 млн лет назад от предков, общих с круглоротыми. Их эволюция шла в направлении развития челюстей, жаберных дуг, парных плавников. Наибольшего разнообразия костные рыбы достигли к началу нашей эры, став одними из процветающих классов позвоночных. Одна из эволюционных ветвей лопастеперых представляла собой кистеперых рыб, которые дали затем начало первым наземным позвоночным. Современные рыбы на 95% представлены лучеперыми.
Класс Земноводные
(Amphibia) объединяет примерно 4000 видов. Земноводные занимают промежуточное положение между водными и наземными животными (рис. 35). Всю жизнь или, по крайней мере, в личиночном состоянии земноводные так или иначе связаны с водой. Они обитают вблизи водоемов, во влажных местах или водоемах во всех районах мира. У них впервые появляется голос. В качестве наземных животных они примитивны, причем их несовершенство отражается на строении и функциях всех их систем органов.
 Покровы земноводных представлены голой кожей, покрытой слизью. Покровы земноводных представлены голой кожей, покрытой слизью.
Скелет разделен на шейный и крестцовый отделы. В черепе еще сохраняются остатки хрящевой ткани, характерной для кистеперых рыб. Грудной клетки и ребер нет, но уже имеются пятипалые передние и задние конечности, развиты плечевой и тазовый пояса конечностей.
Мускулатура хорошо развита и состоит из мышц, двигательные функции которых специфичны.
Пищеварительная система состоит из ротовой полости, переходящей в глотку, пищевода, желудка и кишечника, заканчивающегося клоакой. Развит язык. Имеются печень, поджелудочная железа, а также слюнные железы.
Дыхательная система характеризуется тем, что у личиночных форм дыхание осуществляется с помощью жабр, а у взрослых — легкими, которые имеют вид тонкостенных мешков ячеистого строения. Однако дыхательные пути развиты недостаточно. В газообмене принимает участие также и кожа, содержащая большое количество капилляров.
Кровеносная система представлена трехкамерным сердцем, состоящим из двух предсердий и желудочка, артериями, венами и капиллярами. Имеется два круга кровообращения, но артериальная и венозная кровь еще частично смешиваются.
Выделительная система представлена парой примитивных почек в виде продолговатых тел, двумя мочеточниками, мочевым пузырем и клоакой. »
Нервная система состоит из головного мозга, в котором относительно развит продолговатый мозг. От головного мозга отходят 10 пар нервов. Органы чувств очень развиты. В глазах имеются уплощенный хрусталик и выпуклая роговица. Животные хорошо различают запахи. Среднее ухо содержит слуховую косточку.
Эндокринная система представлена рядом желез внутренней секреции, продукты которых регулируют окраску животных, завершение метаморфоза и другие функции.
Земноводные раздельнополы, их система органов размножения существенно изменена по сравнению с этой системой у рыб. У самцов имеется пара семенников, у самок — пара яичников. Развитие происходит с метаморфозом. Из яиц выходят личинки (в воде), которые развиваются в головастиков.
Хозяйственное значение земноводных ничтожно.
Возникнув в девоне около 300 млн лет назад, они оказались самыми первыми примитивными животными среди наземных позвоночных.
Первые прыгающие лягушки появились около 200 млн лет назад. Предполагают, что предками земноводных являются древние кистеперые рыбы. Парные плавники кистеперых развились в пятипалые конечности земноводных. Покровные кости черепа кистеперых рыб сходны с покровными костями черепа палеозойских земноводных. Предполагают также, что особенности дыхания и передвижения, пригодные для жизни на суше, у предков земноводных появились, когда они были еще настоящими водными животными. Эти особенности, а также отрыв земноводных от воды
И закрепление их на суше возникли, видимо, в связи с изменениями кормовых возможностей. Промежуточной формой между Древними кистеперыми рыбами и современными земноводными являются стегоцефалы, которые произошли от кистеперых рыб и расцвет которых приходился на каменноугольный и пермский периоды. Типичные бесхвостые и хвостатые земноводные появились в верхней юре и среднем меле. Расцвет земноводных имел место в каменноугольном периоде.
Класс Пресмыкающиеся
(Reptilia) — это первые настоящие наземные позвоночные. Количество видов в этом классе достигает 7000. Обитают во многих зонах земного шара. Характерной особенностью их является то, что они размножаются на суше яйцами, дышат исключительно легкими, а кожа имеет роговые покрытия. Для яиц характерно наличие защитной оболочки (скорлупы) и большое количество желтка, что уже само по себе означает первое приспособление к жизни на суше. У организмов этого класса развились также оболочки, окружающие эмбрион (одна из этих оболочек есть амнион). Благодаря этим важным репродуктивным приспособлениям пресмыкающихся относят к амниотам (вместе с птицами и млекопитающими).
У пресмыкающихся получили дальнейшее развитие пятипалые конечности. Впервые у них появляется кора больших полушарий.
Покровы тела представлены кожей, которая покрыта роговыми чешуйками, щитками или пластинками и не содержит слизистых желез. Такие свойства кожи обеспечивают независимость осмотического давления в теле от окружающей среды.
Тело состоит из головы, шеи, туловища, хвоста и конечностей (кроме змей). Скелет характеризуется прогрессивным развитием. Являясь костным, он подразделяется на головной (череп), шейный, грудной, поясничный, крестцовый и хвостовой отделы. Череп у одних видов почти монолитен, (кроме отверстий для глаз, ноздрей и теменного органа), у других дифференцирован на отдельные кости. В шейном отделе позвоночника имеются атлант и эпистрофей, что расширяет двигательные возможности головы. Конечности, которые по строению являются промежуточными между конечностями земноводных и млекопитающих, заканчиваются 5 пальцами.
Скелетная мускулатура намного мощнее, чем у земноводных.
Пищеварительная система значительно дифференцирована. Имеется зачаток слепой кишки.
Дыхательная система представлена трахеей, которая разветвляется на два бронха, входящие в легкие. Последние построены из тонкостенных мешков, имеющих ячеистую структуру. Кожного дыхания уже нет, что привело к усовершенствованию легких в качестве единственного органа дыхания и к участию в дыхании грудной клетки.
Кровеносная система также характеризуется дальнейшим совершенствованием. Сердце трехкамерное, желудочек разделен неполной перегородкой на венозную и артериальную половины. Настоящее четырехкамерное сердце встречается лишь у крокодилов, у которых правый и левый желудочек полностью обособлены, т. е. разделены тонкой перегородкой. У пресмыкающихся два круга кровообращения, но они еще не полностью разделены (даже у крокодилов), в результате чего кровь еще частично смешивается в спинной аорте (как и у земноводных).
Выделительная система представлена парой вторичных тазовых почек (метанефрос) и парой мочеточников, впадающих в клоаку, в которую одновременно открывается и мочевой пузырь. Особенность строения почек заключается в том, что при уменьшении относительной фильтрационной площади клубочков увеличена протяженность канальцев. Осморегулирующая функция почек почти не выражена, т. к. В теле пресмыкающихся нет избытка воды.
Нервная система также характеризуется прогрессивными чертами. В частности, головной мозг имеет черты, характерные для мозга наземных животных. В продолговатом мозгу сформирован изгиб, который обычно встречается у всех амниот. Сформирован также зачаток коры больших полушарий мозга. От головного мозга отходят черепномозговые нервы (12 пар). Характерно более совершенное зрение за счет возможности изменения кривизны хрусталика. В ресничном теле развита поперечно-полосатая мускулатура. В органах слуха увеличена улитка, в слуховой капсуле имеется второе отверстие. Органы обоняния отличаются большой развитостью носовых раковин, развитием вторичного нёба. Очень развит якобсонов орган, обеспечивающий восприятие запаха добычи.
Пресмыкающиеся обладают всеми эндокринными железами, характерными для высших животных. Температура тела пресмыкающихся зависит от среды.
Пресмыкающиеся раздельнополы, характеризуются выраженным половым диморфизмом. Одни пресмыкающиеся являются яйцекладущими, другие —живородящими. Широко развито яйце-живорождение. Пол генетически детерминируется лишь у некоторых видов ящериц, возможно, у всех змей. Однако у многих пресмыкающихся пол детерминируется не генетически, а факторами внешней среды, в частности, температурой. Например, у многих видов черепах действие высоких температур сопровождается появлением на свет лишь самок, низких температур — только самцов. Обратное положение отмечено у крокодилов и некоторых видов ящериц.
Животных этого класса подразделяют на отряды Чешуйчатые (Sguamata), Черепахи (Chelonia), Крокодилы (Crocodylia) и Первоящеры, или Клювоголовые (Prosauria, или Rhyncocephalia).
Значение змей определяется тем, что многие виды обладают ядовитыми железами и зубами. Хозяйственное значение определяется ценностью их кожи. Черепахи имеют хозяйственное значение, поскольку некоторые виды и их яйца съедобны. Крокодилы тоже имеют хозяйственное значение, но опасны в качестве хищников, нападающих на человека.
Пресмыкающиеся входят в главный ствол эволюции, являясь предковыми формами птиц и млекопитающих. Древнейшими пресмыкающимися были котилозавры, которые произошли от стегоцефалов в каменноугольном периоде. Развитие котилозавров дало начала нескольким эволюционным ветвям (ихтиозаврам, плезиозаврам). В качестве слепой боковой группы отделились анапсиды, современными потомками которых являются черепахи, возникшие в триасе.
Важным событием в эволюции пресмыкающихся было появление малых пресмыкающихся (диапсид), череп которых имел два отверстия в височной области. Одним из потомков этих пресмыкающихся является относительно мало модифицированная сейчас гаттерия. Ящерицы и змеи развились из предковых форм диапсид в результате дальнейшей модификации черепа. Другие диапсид-ные формы дали начало археозаврам («правящим рептилиям»), потомками которых являются крокодилы, появившиеся в конце триаса. В мезозое эта группа была процветающей в виде динозавров и летающих пресмыкающихся. Одна из ветвей диапсид могла быть предковой для птиц.
Предполагают, что очень ранняя боковая ветвь из ствола древних котилозавров дала линию, ведущую к терапсидам, представляющим собой пресмыкающихся, напоминающих млекопитающих. Остатки этих животных обнаруживают в перми и триасе. Они являются мостом между рептилиями и млекопитающими, первые из которых появились в конце триаса.
Класс Птицы
(Aves) — эта систематическая группа представлена позвоночными животными, биологические и анатомические свойства которых определяются наличием у них приспособлений для полета и передвижений по твердому субстрату. Известно около 9000 видов. Обитают по всему земному шару, но наибольшее количество видов сосредоточено в тропиках.
Наземная локомоция птиц является двуногой (задние конечности). Передние конечности видоизменены в крылья, имеющие рудименты трех пальцев. Туловище компактное с большой грудиной для крепления мышц крыльев. Кости черепа и скелета заполнены воздухом. Костный хвост уменьшен.
Скелет хорошо развит в области грудной клетки. Кости черепа сращены. Шейные позвонки очень подвижны. Поясничные, крестцовые и часть хвостовых позвонков также сращены.
Покровами тела служит тонкая кожа, не содержащая желез, кроме копчиковой железы у корня хвоста. Производными кожи являются перья на теле, роговые чешуи на конечностях и когти на концах пальцев.
Мускулатура очень развита, особенно мышцы груди, шеи и конечностей.
Пищеварительная система характеризуется отсутствием зубов, но есть зоб (расширение пищевода), который служит для размягчения пищи. Имеются крупная двухлопастная печень и хорошо развитый желчный пузырь.
Дыхательная система обеспечивает приспособление для полета. Поэтому легкие представляют собой уплотненный губчатый орган, дополненный тонкостенными воздушными мешками, располагающимися между органами, в полостях костей, под кожей, в мышцах. Для птиц характерно двойное дыхание, т. к. при взмахе крыльями воздух проходит через легкие. Это позволяет более интенсивный газовый обмен в полете (рис. 36).
Кровеносная система характеризуется тем, что сердце полностью разделено на правую венозную и левую артериальную половины, а имеющиеся два круга кровообращения полностью самостоятельны. Артериальная и венозная кровь не смешиваются. Температура тела постоянна (42—45°С).
Выделительная система характеризуется наличием пары вторичных почек и пары мочеточников, которые открываются непосредственно в клоаку, мочевого пузыря нет. Компоненты мочи вместе с экскрементами выходят наружу через клоаку.
Нервная система птиц характеризуется дальнейшим развитием головного мозга, в котором имеются относительно большие полушария и зрительные доли, хорошо развитый мозжечок. Имеются 12 пар черепно-мозговых нервов.
 Птицы — раздельнополы с выраженным половым диморфизмом. Птицы имеют важное хозяйственное значение,являясь источником мяса, яиц, пуха, пера. Отдельные виды диких птиц имеют промысловое значение. Они имеют также медицинское значение, т. к. могут болеть орнитозом, к которому чувствителен человек. Являясь дополнительными хозяевами ряда паразитов, поддерживают природные очаги таких опасных для человека болезней, как токсоплазмоз и листериоз. Птицы — раздельнополы с выраженным половым диморфизмом. Птицы имеют важное хозяйственное значение,являясь источником мяса, яиц, пуха, пера. Отдельные виды диких птиц имеют промысловое значение. Они имеют также медицинское значение, т. к. могут болеть орнитозом, к которому чувствителен человек. Являясь дополнительными хозяевами ряда паразитов, поддерживают природные очаги таких опасных для человека болезней, как токсоплазмоз и листериоз.
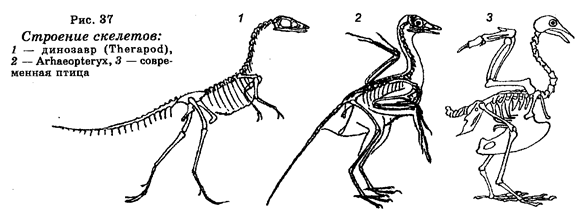
Таежные птицы служат резервуаром вируса таежного энцефалита, птицы Средней Азии — резервуаром вируса клещевого возвратного тифа. Доказано, что птицы могут транспортировать переносчиков возбудителей болезней на очень большие расстояния. Птицам присуща высокая экологическая пластичность.
Классификация птиц очень осложнена, т. к. нет единого мнения о количестве основных систематических единиц в этом классе.
Предками птиц были, вероятно, древнейшие бегающие пресмыкающиеся (рис. 37). Промежуточной формой между пресмыкающимися и птицами считают археоптерикса (Archaeopteryx), ископаемые остатки которого обнаружены в позднеюрских отложениях (возраст 85 млн лет). Найдены также ископаемые остатки птиц (Liaoning), которые имели сходство по ряду признаков как с археоптериксом, так и с современными птицами. У птиц размеры генома значительно уменьшены по сравнению с млекопитающими. Масса ДНК у птиц составляет 2,82,±0,33 пг на клетку, тогда как у млекопитающих она равна 8 пг на клетку. Предполагают, что редукция размеров генома птиц происходила постепенно на протяжении длительного периода и что на протяжении этого периода шел отбор на уменьшенные размеры генома. Предполагают также, что размеры генома обеспечивают одно из приспособлений к полету. В пользу этого предположения свидетельствует тот факт, что размеры геномов у нелетающих птиц являются значительно большими, чем у летающих.
Класс Млекопитающие
, или Звери
(Mammalia) — это наиболее организованные, в основном наземные позвоночные животные, находящиеся на высшей стадии развития и обитающие во всех географических зонах. Они представляют собой современную процветающую группу животных. В этом классе насчитывают около 3200 видов.
Для млекопитающих характерен ряд особенностей. Их детеныши выкармливаются молоком, которое вырабатывается специальными молочными железами. Плод развивается в матке. Благодаря совершенному развитию нервной системы и механизмов терморегуляции у них поддерживается постоянная температура тела, обеспечивающая их активность в любых климатических условиях.
Имеют волосяной или шерстный покров. Масса отдельных особей составляет от 2 г (землеройка-малютка) до 150 тонн (синий кит).
Покровы тела представлены кожей, которая состоит из многослойного эпидермиса и кориума, построенного из волокнистой соединительной ткани. Кожа снабжена многими потовыми и сальными железами. В подкожной клетчатке содержатся жировые клетки. Роговыми образованиями кожи являются волосы, ногти, когти, рога и копыта. У всех млекопитающих имеются молочные железы, которые представляют собой видоизмененные потовые железы.
Осевой скелет характерен четким разделением позвоночника на шейный, грудной, поясничный, крестцовый и хвостовой отделы. Количество шейных позвонков постоянно, у атланта на передней стороне имеются две поверхности. Скелет конечностей очень прочен. Бедренные кости выдерживают очень большие нагрузки, которые у некоторых видов достигают до 1500-2000 кг. Мышечная система очень развита, состоя из множеств специализированных мышц.
Пищеварительная система имеет все отделы, заканчиваясь самостоятельным заднепроходным отверстием. Развиты зубы (кроме отдельных видов, включая китообразных), среди которых различают резцы, клыки и коренные. По характеру пищи, используемой для кормления, различают растительноядных и плотоядных млекопитающих.
Дыхательная система представлена легкими, трахеей, бронхами, бронхиолами, альвеолами.
Кровеносная система очень совершенна. Сердце четырехкамерное, имеются два круга кровообращения, левая дуга аорты. Зрелые эритроциты лишены ядер.
Выделительная система характеризуется тем, что мочевой пузырь открывается в мочеиспускательный канал. Почки парные, отходящие от них мочеточники открываются в мочевой пузырь.
Нервная система достигает вершины в своем развитии. Особо развиты полушария головного мозга, извилины и борозды в коре головного мозга, мозжечок. Очень развиты органы чувств. Орган слуха состоит из наружного уха, наружного прохода, трех слуховых косточек и звуковоспринимающего аппарата. Обоняние связано с развитием пластинчатых носовых раковин и носового лабиринта. Развиты зрение и осязание.
Железы внутренней секреции очень развиты.
Размножение половое, половые железы у особей обоих полов парные. Выражен половой диморфизм. Оплодотворение внутреннее.
Млекопитающих классифицируют на подклассы Первозвери (Prototheria) с отрядом Однопроходные (Monotremata) и Настоящие звери (Theria) с инфраклассами Низшие звери (Metatheria) и Высшие звери (Eutheria) с большим количеством отрядов.
Однопроходные (утконос, ехидна, проехидна) являются обитателями Австралии и характеризуются тем, что подобно пресмыкающимся, они откладывают яйца.
Низшие звери, или Сумчатые (кенгуру, сумчатый волк, сумчатый крот и другие), являются обитателями Австралии и Южной Америки. Не имея плаценты, рожденных детенышей вынашивают в сумке.
Высшие звери являются плацентарными животными. Они чрезвычайно разнообразны (насекомоядные, рукокрылые, грызуны, хищные, ластоногие, китообразные, непарно- и парнокопытные, хоботные, приматы и другие). В эмбриональном онтогенезе питание плодов происходит через плаценту, детеныши рождаются развитыми, характеризуются двойной сменой зубов.
Для млекопитающих характерно чрезвычайное разнообразие в образе жизни. Различают наземных, подземных, водных и даже летающих животных (рукокрылых).
Роль млекопитающих в жизни человека, который сам относится к млекопитающим, чрезвычайно велика и разнообразна. Ее невозможно переоценить. Многие из них, особенно домашние животные, имеют огромное хозяйственное значение, являясь источником продовольствия для человека и сырья для промышленности. Значительное количество видов имеет промысловое значение. Например, основу пушной добычи составляет около 20 видов. Поскольку млекопитающие болеют общими с человеком заболеваниями, служат хозяевами паразитов или являются природными резервуарами возбудителей ряда трансмиссивных заболеваний человека, то очень велико их медицинское значение.

Происхождение млекопитающих обычно объясняют возникновением их от терапсид (древних палеозойских рептилий) в позднем триасе. Многие зоологи считают, что млекопитающие имеют по-лифилетический генез, т. е. каждый подкласс млекопитающих имеет отдельного терапсидного предка. Обладая высокоразвитой нервной системой и другими свойствами, млекопитающие выделились из позвоночных в качестве класса, завоевавшего сушу. Расцвет плацентарных приходится на палеоцен (55-65 млн лет назад). Начиная с третичного периода, они являются преобладающей (процветающей) формой жизни на Земле (рис. 38).
1. Каковы основные черты простейших? В чем заключается их значение в природе и в жизни человека?
2. Входят ли губки и кишечнополостные в основной ствол эволюции?
3. Что вы знаете о классификации гельминтов?
4. Насколько велики различия между плоскими и круглыми гельминтами?
б. Что вы знаете о происхождении круглых гельминтов?
6. На каких принципах основана классификация членистоногих, в чем она заключается и каковы основные черты эволюции животных этого типа?
7. Каково значение членистоногих в природе и жизни человека?
8. Какова древность иглокожих?
9. Каковы признаки, характерные для типа Хордовые? Как вы представляете классификацию Хордовых?
10. Каково эволюционное значение бесчерепных?
11. Каковы характерные черты позвоночных?
12. Каково филогенетическое значение круглоротых и рыб?
13. В чем заключается филогенетическое значение земноводных? С какими ароморфозами связана эволюция животных этого класса?
14. Назовите прогрессивные черты пресмыкающихся. Каковы происхождение и эволюционное значение пресмыкающихся?
15. Назовите прогрессивные черты в организации и приспособлении к полету птиц. Что вы знаете о происхождении животных этого класса? Каково их значение в жизни человека?
16. В чем заключаются главнейшие признаки, характерные для млекопитающих, и особенно их прогрессивные черты?
17. Какова роль млекопитающих в жизни человека?
18. Как и когда произошли млекопитающие?
19. Какое значение в понимании теории эволюции имеет рассмотрение основных свойств животных, принадлежащих к разным типам?
Выховский Б. Е. Зоология. В кн. «История биологии». М.: Наука. 1975. 24-57.
Хадорн Э.. Венер Р. Общая зоология. М.: Мир. 1989. 523 стр.
Carrol S. В. Homeotic genes and the evolution of arthropods and chordates. Nature, 1995, Vol. 376, 479-485.
Chiappe L. M. The first 86 million years of avian evolution. Nature, 1996, Vol. 378, 349-366.
Вирусы представляют собой субмикроскопические внеклеточные формы жизни.
Открытие вирусов принадлежит Д. И. Ивановскому, который в 1892 г. обнаружил возбудителя мозаичной болезни табака и его способность проходить через фильтры, не пропускающие бактерий. По этой причине вирусы первоначально получили название фильтрующихся ядовитых жидкостей. Повторив опыты Д. И. Ивановского и получив аналогичные результаты, голландский микробиолог М. Бейеринк в 1898 г. дал для возбудителя мозаичной болезни табака название «фильтрующаяся вирусная жидкость». Тогда же был описан вирусный возбудитель ящура крупного рогатого скота (Ф. Лефлер и П. Фрош, 1898), а в 1901 г. У. Рид, используя существовавшую со времен А. Пастера традицию называть инфекционное начало вирусом, назвал открытого им возбудителя желтой лихорадки просто вирусом (от лат. virus — яд). Начиная с 1931 г., когда был открыт способ культивирования вирусов в клетках эмбрионов цыплят, вирусы стали широко культивировать в лабораторных условиях, что значительно расширило возможности экспериментального изучения этих структур.
К настоящему времени вирусы открыты у организмов практически всех систематических групп растений и животных (у микоплазм, бактерий, листостебельных растений, простейших, гельминтов, насекомых, земноводных, пресмыкающихся, птиц, млекопитающих). Известно значительное количество вирусов, выделенных от человека. Можно сказать, что вирусы вездесущи.
В лабораторных условиях вирусы культивируют на куриных эмбрионах, в культивируемых соматических клетках, на переживающих лоскутах кожи и в эксплантантах органов, а также в организме восприимчивых животных. Важно отметить, что они не способны к росту на питательных средах, используемых для культивирования бактерий или соматических клеток, и в этом заключается одно из принципиальных отличий их от других организмов.
Вирусы не способны к воспроизведению в свободном состоянии. Их воспроизведение возможно только в клетках. Кроме того, оказавшись в клетках, они ведут себя как облигатные внутриклеточные паразиты, вызывая болезни организмов, в которых паразитируют. Следовательно, вирусам присущи две формы существования, а именно: внеклеточная, или покоящаяся, и внутриклеточная, или репродуцирующаяся.
В общем виде вирусы представляют собой субмикроскопические образования, состоящие из белка и нуклеиновой кислоты и организованные в форме вирусных частиц, часто называемых вирусными корпускулами, вирионами, вироспорами или нуклеокапсидами.
Размеры вирусных частиц составляют от 15—18 до 300—350 нм. Их можно увидеть только с помощью электронного микроскопа. Исключение составляют лишь вирус оспы и некоторые другие крупные вирусы, которые можно увидеть в высокопрецизионном световом микроскопе.
Одиночные вирусы тех или иных видов представляют собой образования различной формы (округлой, палочковидной или другой формы), внутри которых содержится нуклеиновая кислота (ДНК или РНК), заключенная в белковую оболочку (капсид).
Вирусный капсид построен из полипептидных цепей (разных белков), сложенных в несколько слоев. У отдельных вирусов капсид окружен дополнительной мембраной, построенной из сложных вирусоспецифических белков (гликопротеидов) и двойного слоя липидов, идентичных липидам плазматической мембраны клетки-хозяина. Эту вирусную мембрану часто называют супер-капсидом.
Функциональное значение капсида определается тем, что он предохраняет вирусный геном (нуклеиновую кислоту) от повреждений, а также содержит рецепторы, обеспечивающие адсорбцию вирусных частиц на поражаемых ими клетках.
Внутреннее содержимое вирусов, часто называемое геномом или вирусной хромосомой, состоящей из нескольких генов, представляет собой ДНК или РНК в разных формах. Например, геном бактериальных вирусов MI3 и XI74 представлен одноцепочечной кольцевой молекулой ДНК, тогда как у парвовирусов, к которым относят многие вирусы крупного рогатого скота, свиней, кошек, крыс и других животных, он представлен одноцепочечной линейной молекулой ДНК. Геном вирусов полиомы и SV40 представлен двухцепочечной кольцевой молекулой ДНК, но у аденовирусов двухцепочечная кольцевая ДНК имеет на 5'-концах цепей ковалентно присоединенный белок, а у вируса оспы двухцепочечная ДНК характеризуется тем, что концы ее цепей ковалентно «сшиты» фосфодиэфирными связями.
Геном вирусов мозаичной болезни табака, полиомиелита и некоторых бактериальных вирусов представлен одноцепочечной РНК, тогда как геномом реовируса служит двухцепочечная РНК-Воспроизводство вирусов происходит в клетках хозяина и состоит из нескольких стадий, а именно: адсорбция и проникновение вирусов в клетки, синтез вирусных белков, необходимых для репликации вирусной нуклеиновой кислоты, репликация нуклеиновой кислоты, синтез белков капсида, сборка вирусов и выход из клетки «готовых» вирусных частиц.
ДНК-содержащие вирусы имеют либо собственные ферменты репликации (в капсиде), либо в их геноме закодирована информация о синтезе вирусных ферментов, обеспечивающих репликацию вирусной нуклеиновой кислоты. Количество этих ферментов различно в применении к разным вирусам. Например, в геноме бактериального вируса Т4 закодирована информация о синтезе около 30 вирусных ферментов. Далее геном крупных вирусов кодирует нуклеазы, разрушающие ДНК клетки-хозяина, а также белки, воздействие которых на клеточную РНК-полимеразу сопровождается тем, что «обработанная» таким образом РНК-полимераза транскрибирует на разных стадиях вирусной инфекции разные вирусные гены. Напротив, малые по размерам ДНК-содержащие вирусы в большей мере зависят от ферментов клеток-хозяев. Например, синтез ДНК аденовирусов обеспечивается клеточными ферментами.
Репликация ДНК-содержащих вирусов начинается с 0-пункта их хромосомы, к которому прикрепляются белки клетки-хозяина, «притягивающие» к себе как вирусные, так и клеточные ферменты репликации.
У некоторых РНК-содержащих вирусов в геноме закодированы РНК-зависимые транскриптазы (обратные транскриптазы). Поэтому репликация генома РНК-содержащих вирусов катализируется собственной обратной транскриптазой, которая упаковывается в капсиде при каждой репликации вирусов в клетке-хозяине. Такие РНК-содержащие вирусы получили название ретровирусов (от лат. retro — возврат назад).
Классическим примером ретровирусов служит вирус саркомы кур Рауса (рис. 39).
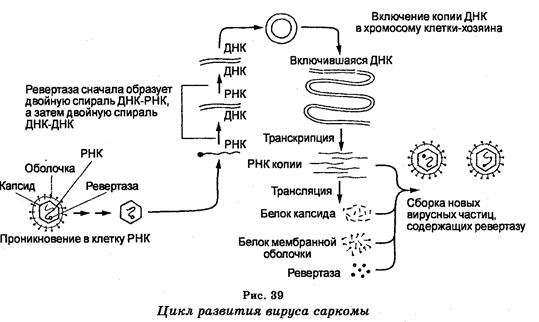
К ретровирусам относят вирус иммунодефицита человека (ВИЧ), являющийся причиной синдрома приобретенного иммунодефицита (СПИДа). Однако РНК вируса полиомиелита служит в качестве мРНК.
Классификация вирусов является сложной и часто противоречивой. Тем не менее ДНК-содержащие и РНК-содержащие вирусы обычно классифицируют раздельно на разные семейства. Например, среди ДНК-содержащих вирусов животных классифицируют семейства парвовирусов (вирусы крыс, кошек и др. животных), паповавирусов (вирусы бородавок человека, полном и другие), аде-новирусов (вирусы фарингитов и конъюнктивитов человека, вирусы млекопитающих), поксвирусов (вирусы оспы человека и животных), вирусы герпеса (вирусы герпеса, опоясывающего лишая человека, ларинготрахеита птиц и др.), иридовирусы (вирусы оспы свиней, африканской чумы свиней и др.).
Среди РНК-содержащих вирусов животных классифицируют семейства пикорнавирусов (риновирусы человека, вирус полиомиелита человека, полиовирусы животных, вирус ящура животных и др.), реовирусов (вирусы тендосиновита кур и др.), миксовиру-сов (вирусы гриппа, кори, бешенства человека, чумы плотоядных и др.), арбовирусов (вирусы клещевого и японского энцефалитов, желтой лихорадки человека и Др.). Подобные классификации существуют и в случае вирусов растений; что касается фагов, то их классифицируют на основе того, какие бактерии они лизируют. Например, кишечные фаги — это фаги, лизирующие кишечные бактерии, дизентерийные фаги — это фаги, лизирующие дизентерийные бактерии и т. д.
Поскольку вирусы обнаружены в клетках организмов разных систематических групп, мы рассмотрим здесь вирусы человека, животных (млекопитающих), растений и бактерий раздельно.
Вирусы человека и животных являются наиболее изученными по сравнению с вирусами другого происхождения. Они вызывают болезни, многие из которых характеризуются большей тяжестью лечения и высокой смертностью. Наиболее известными вирусными болезнями человека являются грипп, полиомиелит, бешенство, оспа, клещевой энцефалит и другие, а домашних животных — бешенство, ящур, чума, оспа, энцефаломиелит и др., отдельные из которых являются повальными болезнями.
Морфология вирусов человека и животных довольно разнообразна, но в общем они являются овальными образованиями, диаметр которых составляет несколько десятков нанометров (рис. 40).
Как уже отмечено, репродукция вирусов происходит только в живых клетках, т. е. синтез как вирусного генома (нуклеиновой кислоты), так и белков капсида происходит только в клетках. В клетках осуществляется также и сборка из вирусной нуклеиновой кислоты и белков новых инфекционных вирусных частиц.
 Вирусная инфекция человека (животных) начинается с того, что инфицирующий вирус связывается с цитоплазматической мембраной клеток, после чего в результате эндоцитоза проникает внутрь клеток. Внутри клеток вирусные частицы оказываются или в ядре или в цитозоле, где их нуклеиновая кислота освобождается от капсида. Например, ДНК вируса герпеса человека освобождается от капсида в ядре. Там же происходит и сборка новых нуклеокапси-дов, после чего они покидают клетку. Напротив, РНК некоторых РНК-содержащих вирусов освобождается от капсида в цитозоле, после чего она транслируется на рибосомах клетки-хозяина с образованием кодируемой вирусом РНК-полимеразы. Последняя синтезирует РНК, отдельные из копий которой выполняют роль мРНК, транслируемой в мембранные и капсидные белки. Сборка вирусных частиц идет затем на плазматической мембране, после чего они покидают клетку. Вирусная инфекция человека (животных) начинается с того, что инфицирующий вирус связывается с цитоплазматической мембраной клеток, после чего в результате эндоцитоза проникает внутрь клеток. Внутри клеток вирусные частицы оказываются или в ядре или в цитозоле, где их нуклеиновая кислота освобождается от капсида. Например, ДНК вируса герпеса человека освобождается от капсида в ядре. Там же происходит и сборка новых нуклеокапси-дов, после чего они покидают клетку. Напротив, РНК некоторых РНК-содержащих вирусов освобождается от капсида в цитозоле, после чего она транслируется на рибосомах клетки-хозяина с образованием кодируемой вирусом РНК-полимеразы. Последняя синтезирует РНК, отдельные из копий которой выполняют роль мРНК, транслируемой в мембранные и капсидные белки. Сборка вирусных частиц идет затем на плазматической мембране, после чего они покидают клетку.
Выход вирусных частиц из клеток характеризуется особенностями, определяемыми видом вирусов. Вновь образованные вирусные частицы могут выходить из клеток наружу постепенно. В этом случае структура клеток не нарушается, в них продолжается синтез новых вирусных частиц. Однако после инфекции клеток генетический материал вирусов может включаться в их геном и находиться там неопределенно долгое время в активном состоянии. В данном случае вирус ведет себя по типу так называемой латентной (скрытой) инфекции. Дальнейшее развитие латентных вирусов может наступить в результате какого-либо воздействия, после которого вирусы освобождаются из клеток с лизисом их или без лизиса.
Помимо способности вызывать инфекционные болезни человека и животных, отдельные виды вирусов обладают свойством индуцировать образование опухолей. Таким свойством обладают как ДНК-содержащие, так и РНК-содержащие вирусы (ретровирусы). Вирусы, способные вызывать опухоли, получили название опухо-леродных или онкогенных вирусов, а процесс изменения клеток и превращение их в раковые называют вирусной неопластической трансформацией.
В случае ДНК-содержащих вирусов человека и животных их свойство вызывать опухоли зависит от отношения вирусной ДНК к хромосомам клетки. Вирусная ДНК может оставаться подобно ллазмидам в клетке в автономном состоянии, реплицируясь вместе с клеточными хромосомами. При этом регуляция деления клеток не нарушается. Однако вирусная ДНК может включиться в одну или несколько хромосом клетки-хозяина. При таком исходе деление клеток становится нерегулируемым. Другими словами, инфицированные ДНК-содержащим вирусом клетки превращаются в раковые. Примером онкогенных ДНК-содержащих вирусов является вирус SV40, выделенный много лет назад из клеток обезьян. Онкогенное действие этих вирусов зависит от того, что отдельные вирусные гены действуют как онкогены, активизируя клеточную ДНК и побуждая клетки к вступлению в 8-фазу с последующим неконтролируемым делением. РНК-содержащие вирусы из-за включения их РНК в одну или несколько хромосом клетки-хозяина также обладают онкогенным действием. В геноме этих вирусов также есть онкогены, однако они существенно отличаются от онкогенов ДНК-содержащих вирусов тем, что в геноме клеток-хозяев присутствуют их гомологи в виде протоонкогенов. Когда РНК-содержащие вирусы инфицируют клетки, они «захватывают» в свой геном протоонкогены, которые представляют собой последовательности ДНК, контролирующие синтез белков (киназ, факторов роста, рецепторов факторов роста и др.), участвующих в регуляции клеточного деления. Однако известно, что существуют и другие способы превращения клеточных протоонкогенов в вирусные онкогены.
К настоящему времени установлено очень большое количество протоонкогенов. Примерами онкогенных РНК-содержащих вирусов являются вирус саркомы кур Рауса, а также вирусы сарком птиц, мышей, кошек, обезьян и других животных (табл. 1).
Данные об онкогенных вирусах имеют огромное значение не только для понимания многообразия жизни, но и для поисков эффективных путей лечения и профилактики злокачественных новообразований у человека и животных.
Наконец, как уже отмечено ВИЧ или HIV (Human immunodeficiency virus) способен вызвать СПИД или AIDS (acquired immunodeficiency syndrom), который в конечном итоге смертелен для человека. В отличие от других ретровирусов, ВИЧ в своем геноме содержит пять дополнительных открытых рамок чтения, которые кодируют белки, активирующие или репрессирующие вирусный белковый синтез и, возможно, другие функции. Летальный эффект этого вируса связан с тем, что, убивая специализированные хелперные клетки СДЧ (клетки Т), он повреждает иммунную систему человека, т. к. без этих клеток В-клетки не могут размножаться в ответ на поступление в организм нового антигена (см. § 96). Механизм инфицирующего и летального воздействия ВИЧ на клетки СДЧ в общем виде состоит в том, что при инфицировании клеток его капсидный белок связывается с трансмембранным клеточным белком, после чего вирусный капсид сливается с мембраной клетки, а вслед за этим вирусная РНК освобождается в клетку, где она после конверсии в двухцепочечную ДНК, включается в хромосому в качестве провируса. Белок, синтезируемый под контролем про-вируса, позволяет инфицированным Т-клеткам сливаться с неинфицированными Т-клетками, что ведет к гибели всех этих клеток. Следовательно, человек умирает от неспособности защититься иммунологически от тех инфекций, которые сами по себе не являются смертельными.
Таблица 1
Некоторые онкогены ретровирусов и функции, контролируемые этими протоонкогенами
| Онкоген
|
Вирус
|
Функция протоонкогена
|
| erb-A
|
Вирус эритробластоза птиц
|
Рецептор гормона щитовидной железы
|
| erb-B
|
Вирус эритролеикоза птиц
|
Тирозиновая протеинкиназа.
Рецептор фактора роста эпидермиса
|
| myb
|
Вирус миелобластоза птиц
|
Ядерный белок
|
| mos
|
Вирус миелоцитоматоза птиц
|
Ядерный фактор транскрипции
|
| fos
|
Вирус саркомы мышей
|
Протеинкиваза (серино-вая/треониновая)
|
| H-ras
|
Вирус остеосаркомы мышей
Вирус саркомы крыс
|
Ядерный фактор транскрипции G-белок
|
| sis
|
Вирус саркомы обезьян
|
В-цепь тромбоцитарного фактора роста
|
| yes
|
Вирус саркомы Рауса
|
Тирозиновая протеинкиназа
|
Впервые ВИЧ был выделен в 1959 г. в Заире, после чего второй случай выделения этого вируса (в США) датируется 1969 г. С тех пор был выделен ряд штаммов ВИЧ. Открытие ВИЧ позволило по-новому взглянуть на мир вирусов, с одной стороны, и на характер общения между людьми, с другой стороны.
Вирусы, растений также широко распространены в природе, вызывая болезни растений разной видовой принадлежности. Как уже отмечено, первым открытым вирусом оказался МБТ-вирус мозаичной болезни табака (рис. 41), повреждающий листья растений этого вида. Помимо вируса мозаичной болезни табака широко известны вирусы некроза табака, желтой карликовости картофеля, желтой мозаики репы, а также вирусы, поражающие многие другие культурные и дикие растения.
Форма вирусов растений в основном бывает палочковидной и округлой. Размеры вирусов палочковидной формы составляют 300-480 х 15 нм, а размеры тех, которые имеют округлую форму, равны 25-30 нм.
 Основными путями передачи вирусов от больных растений к здоровым является физический контакт между растениями, прививки растений, перенос вирусов через почву, а также перенос насекомыми. Основными путями передачи вирусов от больных растений к здоровым является физический контакт между растениями, прививки растений, перенос вирусов через почву, а также перенос насекомыми.
Вирусные болезни растений приносят значительный ущерб сельскому хозяйству.
Вирусы бактерий, или бактериофаги (фаги), поражают бактерии, принадлежащие к разным систематическим группам. В молекулярной биологии в начале ее развития выдающаяся роль в качестве экспериментальных моделей принадлежала бактериофагам так называемой Т-группы, к которым чувствительны клетки Е. coli. Эти бактериофаги сейчас
являются самыми изученными, и все, что известно о бактериофагах, обязано в основном Т-бактериофагам.
Бактериофаги Т-группы имеют форму барабанных палочек размером 100 х 25 нм. Их геном представлен ДНК. Они являются вирулентными фагами, т. к. после инфипирования ими бактериальных клеток последние лизируются с освобождением большого количества вновь синтезированных фаговых частиц.
В противоположность вирулентным фагам, известны так называемые фаги умеренного действия, или просто умеренные фаги. Типичным представителем таких фагов является фаг К, который тоже использовался и используется в качестве экспериментальной модели для выяснения многих вопросов молекулярной генетики. Фагу X присущи два важных свойства. Подобно вирулентным фагам он может инфицировать бактериальные клетки, размножаться вегетативно, продуцируя в клетках сотни копий и лизировать клетки с освобождением зрелых фагочастиц. Однако ДНК этого фага может включаться в бактериальную хромосому, превращаясь в про-фаг. При этом происходит так называемая лизогенизация бактерий, а бактерии, содержащие профаг, называют лизогенными. Лизогенные бактериальные клетки могут обладать профагом бесконечно долгое время, при этом не лизируясь. Лизис с освобождением новых фагочастиц отмечается после воздействия на лизоген-ные бактерии какого-либо фактора, например УФ-излучения, которое индуцирует развитие профага в фаг. Изучение лизогенных бактерий позволило получить ряд новых данных о роли разных белков в действии фаговых генов.
Бактериофаги используют в медицине в качестве лечебного и профилактического средства в случае отдельных бактериальных болезней.
Вопросы происхождения вирусов обсуждаются на протяжении всего времени, начиная с момента их открытия, поскольку необходимость рациональных объяснений эволюции вирусов определяется фундаментальным и практическим значением этих структур.
Одним из обсуждаемых вопросов является вопрос о времени появления вирусов. Поскольку вирусы не содержат ни рибосом, ни АТФ и поскольку они не могут существовать вне клеток, то считают, что они возникли позднее клеток.
Что же касается возникновения вирусов, то по этому поводу известно несколько гипотез. Первоначально считали, что они являются бактериями, включившимися в соматические клетки и ставшими там дегенеративными формами. В порядке обоснования этой гипотезы приводили данные о крупных вирусах, которые можно видеть в световом микроскопе с большим разрешением. Однако с началом развития молекулярной биологии на происхождение вирусов стали смотреть по-другому. Пререквизитом к появлению вирусов стали считать формирование последовательностей ДНК, способных к репликациям. Так как эти молекулярные события предположи-тельно представлялись сходными с событиями, предшествующими появлению плазмид (образование кольцевых молекул ДНК со свойствами репликонов из повторяющихся последовательностей ДНК на основе реципрокного кроссинговера), то в современной литературе четко обозначилась тенденция рассматривать происхождение, по крайней мере, бактериальных вирусов совместно с бактериальными плазмидами, причем дополнительным основанием к такому рассмотрению обычно считают сходство по ряду свойств между умеренными фагами и плазмидами.
Во-первых, как умеренные фаги, так и плазмиды (факторы переноса) способны к автономной репликации в цитоплазме и к включению в хромосому.
Во-вторых, факторы переноса детерминируют синтез специфических клеточных рецепторов, которые в отдельных случаях сходны с рецепторами для адсорбции фагов.
Наконец, факторы переноса, как и бактериальные вирусы, построены из нуклеиновой кислоты одного типа, зависят в своем метаболизме от клетки-хозяина и инфекциозны по отношению к клетке-хозяину.
Таким образом, бактериальные вирусы соответствуют ряду важных критериев, обычно используемых в определении плазмид. По этой причине плазмиды можно рассматривать в качестве бактериальных вирусов, у которых вместо механизма, обеспечивающего синтез белковой оболочки, получил развитие механизм, обеспечивающий их конъюгативность и эффективное распространение среди бактерий разных видов и родов.
Следовательно, бактериальные вирусы по отношению к плаз-мидам являются прогрессивными структурами, которые в ходе эволюции приобрели специализированные функции, необходимые для осуществления сложного цикла — воспроизводства инфекционных вирусных частиц и для существования за пределами клетки-хозяина (во внеклеточной форме). Первый бактериальный вирус, вероятно, возник, когда у плазмид появился ген (гены), кодирующий белок (белки) капсида.
Однако между бактериальными вирусами и плазмидами типа факторов переноса все же имеются очень важные различия.
Во-первых, факторы переноса, находясь в бактериях, контролируют на их поверхности синтез специализированных структур пиле и, вследствие чего бактериальные клетки обладают конъю-гативностью, т. е. способностью действовать в качестве генетических доноров. Бактериальные вирусы этой способностью не обладают.
Во-вторых, факторы переноса способны передаваться от клетки к клетке в результате клеточных контактов. Способ инфицирова-ния бактерий вирусами является совершенно иным и заканчивается фаголизисом инфицированной бактериальной клетки.
В-третьих, факторы переноса способны мобилизовать бактериальную хромосому на перенос от одной клетки к другой, причем также в результате клеточных контактов. Бактериальные вирусы этой способностью не обладают. Что же касается способности отдельных бактериальных вирусов переносить сегменты бактериальной хромосомы от одних клеток к другим, то механизм такого генетического обмена (трансдукции) не связан с клеточными контактами.
В-четвертых, сайты включения отдельных плазмид в хромосому более или менее разнообразны, тогда как интеграция профагов чаще происходит только в определенных сайтах хромосомы.
Наконец, плазмиды не обладают способностью лизировать бактерии, тогда как все фаги, включая также умеренные (в автономном состоянии), всегда вызывают лизис бактерий. Наличие перечисленных различий можно объяснить приобретением специфических преобразований бактериальными вирусами и плазмидами в ходе дивергентной эволюции.
Если в объяснениях происхождения бактериальных вирусов наметился несомненный успех, то происхождение вирусов животных и растений не имеет удовлетворительных объяснений, хотя их происхождение тоже иногда связывают с плазмидами.
Обсуждая происхождение вирусов, нельзя не привлечь внимание к одному важному моменту, заключающемуся в том, что вирусы оказывали и оказывают влияние на эволюцию организмов, в которых они паразитируют. Это влияние может выражаться как в их способности переносить генетическую информацию от одних организмов к другим горизонтально, так и в способах действовать в качестве мутагенов (см. гл. X).
1. Что собой представляют вирусы и к какой форме жизни их можно отнести?
2. Как организованы вирусы, каковы сходства и различия в организации их геномов?
3. Что понимают под ретровирусами и каковы особенности их структуры и жизненного цикла?
4. Доступны ли вирусы для классификации? Как классифицируют вирусы?
5. Назовите наиболее известные вирусы человека и болезни, вызываемые этими вирусами.
6. Назовите наиболее известные вирусы животных и чем они отличаются от вирусов человека?
7. Могут ли вирусы вызывать болезни растений?
8. Могут ли вирусы растений вызывать болезни животных?
9. Какова роль вирусов в качестве экспериментальных моделей в молекулярной биологии?
10. Что вы знаете об онкогенных вирусах?
11. Можете ли вы сформулировать гипотезу о происхождении вирусов?
12. Реально ли допущение влияния вирусов на эволюцию организмов, в которых они паразитируют? В случае положительного ответа сформулируйте доводы в пользу этого допущения.
Пехов А. П. Основы плазмидологии. М.: РУДН. 1996. 231 стр.
Филдс Б„ Найп Д. (ред.). Вирусология. М.: Мир. 1989. Т. I. 494 стр.
White D„ Ferner F. Medical Virology. Academic Press. 1994. 603 pp.
Wiedbrauk D., Farkas D. Molecular Methods for Virus Detection. Academic Press. 1995. 386 pp.

«Системой можно назвать только комплекс таких
изобретательно вовлеченных компонентов, у которых взаимодействие
и взаимоотношения принимают характер взаимодействия компонентов
для получения фокусированного результата».
П. К. АНОХИН
,
1978
В широком плане под системой (от греч. systema — целое, составленное из частей) обычно понимают какую-либо совокупность взаимосвязанных, расположенных в определенном порядке частей какого-либо единого целостного образования или совокупность принципов какой-либо теории. Различают неживые и живые системы. Неживыми системами являются системы аксиом и определений, системы счислений, используемые в математике, системы информации и другие. Напротив, живые системы являются категориями биологическими.
Живые системы характеризуются рядом особенностей, которые отличают их от живых систем. Важнейшая особенность живых систем заключается в том, что их жизнь невозможна без притока в них энергии, обмена веществ и обмена информацией. Можно сказать, что они взаимодействуют со средой и по этой причине являются открытыми системами. Далее для живых систем характерна способность к самовоспроизводству, саморегуляции и самовосстановлению, в основе которого лежит способность к восстановлению повреждений собственного генетического материала. Наконец, всем живым системам присуща строгая пространственно-временная организация, позволяющая их существование во времени и пространстве и основанная на единстве структурно-функциональных связей между их частями.
Живыми системами являются клетки, ткани, органы, системы органов, организмы, популяции организмов, экологические системы, биосфера в целом. Начав с описания представлений о сущности жизни, свойствах и уровнях организации живого, в этом разделе мы остановимся на строении и функциях клеток, а также на росте, размножении и индивидуальном развитии организмов.
Вопрос о сущности жизни является одним из давних вопросов в биологии, поскольку интерес к нему восходит еще к античным векам. Дававшиеся в разные времена определения жизни не могли быть исчерпывающими из-за отсутствия достаточных данных. Лишь развитие молекулярной биологии привело к новому пониманию сущности жизни, определению свойств живого и вычленению уровней организации, живого.
Всеобщим методологическим подходом к пониманию сущности жизни в настоящее время является понимание жизни в качестве процесса, конечным результатом которого является самообновление, проявляющееся в самовоспроизведении. Все живое происходит только из живого, а всякая организация, присущая живому, возникает только из другой подобной организации. Следовательно, сущность жизни заключается в ее самовоспроизведении, в основе которого лежит координация физических и химических явлений и которое обеспечивается передачей генетической информации от поколений к поколениям. Именно эта информация обеспечивает самовоспроизведение и саморегуляцию живых существ. Поэтому жизнь — это качественно особая форма существования материи, связанная с воспроизведением. Явления жизни представляют собой форму движения материи, высшей по сравнению с физической и химической формами его существования.
Живое построено из тех же химических элементов, что и неживое (кислород, водород, углерод, азот, сера, фосфор, натрий, калий, кальций и другие элементы). В клетках они находятся в виде органических соединений. Однако организация и форма существования живого имеет специфические особенности, отличающие живое от предметов неживой природы.
В качестве субстрата жизни внимание привлекают нуклеиновые кислоты (ДНК и РНК) и белки. Нуклеиновые кислоты — это сложные химические соединения, содержащие углерод, кислород, водород, азот и фосфор. ДНК является генетическим материалом клеток, определяет химическую специфичность генов. Под контролем ДНК идет синтез белков, в котором участвуют РНК.
Белки — это также сложные химические соединения, содержащие углерод, кислород, водород, азот, серу, фосфор. Молекулы белков характеризуются большими размерами, чрезвычайным разнообразием, которое создается аминокислотами, соединенными в полипептидных цепях в разном порядке. Большинство клеточных белков представлено ферментами. Они выступают также в роли структурных компонентов клетки. Каждая клетка содержит сотни разных белков, причем клетки того или иного типа обладают белками, свойственными только им. Поэтому содержимое клеток каждого типа характеризуется определенным белковым составом.
Ни нуклеиновые кислоты, ни белки в отдельности не являются субстратами жизни. В настоящее время считают, что субстратом жизни являются нуклеопротеиды. Они входят в состав ядра и цитоплазмы клеток животных и растений. Из них построены хроматин (хромосомы) и рибосомы. Они обнаружены на протяжении всего органического мира — от вирусов до человека. Можно сказать, что нет живых систем, не содержащих нуклеопротеидов. Однако важно подчеркнуть, что нуклеопротеиды являются субстратом жизни лишь тогда, когда они находятся в клетке, функционируют и взаимодействуют там. Вне клеток (после выделения из клеток) они являются обычными химическими соединениями. Следовательно, жизнь есть, главным образом, функция взаимодействия нуклеиновых кислот и белков, а живым является то, что содержит самовоспроизводящую молекулярную систему в виде механизма воспроизводства нуклеиновых кислот и белков.
В отличие от живого различают понятие «мертвое», под которым понимают совокупность некогда существовавших организмов, утративших механизм синтеза нуклеиновых кислот и белков, т. е. способность к молекулярному воспроизведению. Например, «мертвым» является известняк, образованный из остатков живших когда-то организмов.
Наконец, следует различать «неживое», т. е. ту часть материи, которая имеет неорганическое (абиотическое) происхождение и ничем не связана в своем образовании и строении с живыми организмами. Например, «неживым» является известняк, образованный из неорганических вулканических известняковых отложений. Неживая материя в отличие от живого не способна поддерживать свою структурную организацию и использовать для этих целей внешнюю энергию.
Обсуждая молекулы, рассматриваемые в качестве субстрата жизни, нельзя не отметить, что они подвергаются непрерывным превращениям во времени и пространстве. Достаточно сказать, что ферменты могут превратить любой субстрат в продукт реакции в исключительно короткое время. Поэтому определение нуклеопротеидов в качестве субстрата жизни означает признание последнего в качестве очень подвижной системы.
Как живое, так и неживое построены из молекул, которые изначально являются неживыми. Тем не менее живое резко отличается от неживого. Причины этого глубокого различия определяются свойствами живого, а молекулы, содержащиеся в живых системах, называют биомолекулами.
Для живого характерен ряд свойств, которые в совокупности «делают» живое живым. Такими свойствами являются самовоспроизведение, специфичность организации, упорядоченность структуры, целостность и дискретность, рост и развитие, обмен веществ и энергии, наследственность и изменчивость, раздражимость, движение, внутренняя регуляция, специфичность взаимоотношений со средой.
Самовоспроизведение (репродукция
). Это свойство является важнейшим среди всех остальных. Замечательной особенностью является то, что самовоспроизведение тех или иных организмов повторяется в неисчислимых количествах генераций, причем генетическая информация о самовоспроизведении закодирована в молекулах ДНК. Положение «все живое происходит только от живого» означает, что жизнь возникла лишь однажды и что с тех пор начало живому дает только живое. На молекулярном уровне самовоспроизведение происходит на основе матричного синтеза ДНК, которая программирует синтез белков, определяющих специфику организмов. На других уровнях оно характеризуется чрезвычайным разнообразием форм и механизмов, вплоть до образования специализированных половых клеток (мужских и женских). Важнейшее значение самовоспроизведения заключается в том, что оно поддерживает существование видов, определяет специфику биологической формы движения материи.
Специфичность организации
. Она характерна для любых организмов, в результате чего они имеют определенную форму и размеры. Единицей организации (структуры и функции) является клетка. В свою очередь клетки специфически организованы в ткани, последние — в органы, а органы — в системы органов. Организмы не «разбросаны» случайно в пространстве. Они специфически организованы в популяции, а популяции специфически организованы в биоценозы. Последние вместе с абиотическими факторами формируют биогеоценозы (экологические системы), являющиеся элементарными единицами биосферы.
Упорядоченность структуры
. Для живого характерна не только сложность химических соединений, из которого оно построено, но и упорядоченность их на молекулярном уровне, приводящая к образованию молекулярных и надмолекулярных структур. Создание порядка из беспорядочного движения молекул — это важнейшее свойство живого, проявляющееся на молекулярном уровне. Упорядоченность в пространстве сопровождается упорядоченностью во времени. В отличие от неживых объектов упорядоченность структуры живого происходит за счет внешней среды. При этом в среде уровень упорядоченности снижается.
Целостность (непрерывность) и дискретность (прерывность).
Жизнь целостна и в то же время дискретна как в плане структуры, так и функции. Например, субстрат жизни целостен, т. к. представлен нуклеопротеидами, но в то же время дискретен, т. к. состоит из нуклеиновой кислоты и белка. Нуклеиновые кислоты и белки являются целостными соединениями, однако тоже дискретны, состоя из нуклеотидов и аминокислот (соответственно). Репликация молекул ДНК является непрерывным процессом, однако она дискретна в пространстве и во времени, т. к. в ней принимают участие различные генетические структуры и ферменты. Процесс передачи наследственной информации тоже является непрерывным, но он дискретен, т. к. состоит из транскрипции и трансляции, которые из-за ряда различий между собой определяют прерывность реализации наследственной информации в пространстве и во времени. Митоз клеток также непрерывен и одновременно прерывен. Любой организм представляет собой целостную систему, но состоит из дискретных единиц — клеток, тканей, органов, систем органов. Органический мир также целостен, поскольку существование одних организмов зависит от других, но в то же время он дискретен, состоя из отдельных организмов.
Рост и развитие.
Рост организмов происходит путем прироста массы организма за счет увеличения размеров и числа клеток. Он сопровождается развитием, проявляющимся в дифференцировке клеток, усложнении структуры и функций. В процессе онтогенеза формируются признаки в результате взаимодействия генотипа и среды. Филогенез сопровождается появлением гигантского разнообразия организмов, органической целесообразностью. Процессы роста и развития подвержены генетическому контролю и нейро-гуморальной регуляции.
Обмен веществ и энергии
. Благодаря этому свойству обеспечивается постоянство внутренней среды организмов и связь организмов с окружающей средой, что является условием для поддержания жизни организмов. Живые клетки получают (поглощают) энергию из внешней среды в форме энергии света. В дальнейшем химическая энергия преобразуется в клетках для выполнения многих работ. В частности, для осуществления химической работы в процессе синтеза структурных компонентов клетки, осмотической работы, обеспечивающей транспорт разных веществ в клетки и вывод из них ненужных веществ, и механической работы, обеспечивающей сокращение мышц и передвижение организмов. У неживых объектов, например, в машинах химическая энергия превращается в механическую только в случае двигателей внутреннего сгорания.
Таким образом, клетка является изотермической системой. Между ассимиляцией (анаболизмом) и диссимиляцией (катаболизмом) существует диалектическое единство, проявляющееся в их непрерывности и взаимности. Например, непрерывно проходящие в клетке превращения углеводов, жиров и белков являются взаимными. Потенциальная энергия поглощаемых клетками углеводов, жиров и белков превращается в кинетическую энергию и тепло по мере превращения этих соединений. Замечательной особенностью клеток является то, что они содержат ферменты. Будучи катализаторами, они ускоряют протекание реакций, синтеза и распада в миллионы раз, при этом в отличие от органических реакций осуществляемых с использованием искусственных катализаторов (в лабораторных условиях), ферментативные реакции в клетках осуществляются без образования побочных продуктов.
В живых клетках энергия, полученная из внешней среды, накапливается в виде АТФ (аденозинмонофосфата). Теряя концевую фосфатную группу, что имеет место при передаче энергии другим молекулам, АТФ превращается в АДФ (аденозиндифосфат). В свою очередь получая фосфатную группу (за счет фотосинтеза или химической энергии), АДФ может снова превратиться в АТФ, т. е. стать главным носителем химической энергии. Такие особенности у неживых систем отсутствуют.
Обмен веществ и энергии в клетках ведет к восстановлению (замене) разрушенных структур, к росту и развитию организмов.
Наследственность и изменчивость
. Наследственность обеспечивает материальную преемственность между родителями и потомством, между поколениями организмов, что в свою очередь обеспечивает непрерывность и устойчивость жизни. Основу материальной преемственности в поколениях и непрерывности жизни составляет передача от родителей к потомству генов, в ДНК которых зашифрована генетическая информация о структуре и свойствах белков. Характерной особенностью генетической информации является ее чрезвычайная стабильность.
Изменчивость связана с появлением у организмов признаков, отличных от исходных, и определяется изменениями в генетических структурах. Наследственность и изменчивость создают материал для эволюции организмов.
Раздражимость.
Реакция живого на внешние раздражения является проявлением отражения, характерного для живой материи. Факторы, вызывающие реакцию организма или его органа, называют раздражителями. Ими являются свет, температура среды, звук, электрический ток, механические воздействия, пищевые вещества, газы, яды и др.
У организмов, лишенных нервной системы (простейшие и растения), раздражимость проявляется в виде тропизмов, таксисов и настий. У организмов, имеющих нервную систему, раздражимость проявляется в виде рефлекторной деятельности. У животных восприятие внешнего мира осуществляется через первую сигнальную систему, тогда как у человека в процессе исторического развития сформировалась еще и вторая сигнальная система. Благодаря раздражимости организмы уравновешиваются со средой. Избирательно реагируя на факторы среды, организмы «уточняют» свои отношения со средой, в результате чего возникает единство среды и организма.
Движение
. Способностью к движению обладают все живые существа. Многие одноклеточные организмы двигаются с помощью особых органоидов. К движению способны и клетки многоклеточных организмов (лейкоциты, блуждающие соединительнотканные клетки и др.), а также некоторые клеточные органеллы. Совершенство двигательной реакции достигается в мышечном движении многоклеточных животных организмов, которое заключается в сокращении мышц.
Внутренняя регуляция.
Процессы, протекающие в клетках, подвержены регуляции. На молекулярном уровне регуляторные механизмы существуют в виде обратных химических реакций, основу которых составляют реакции с участием ферментов, обеспечивающие замкнутость процессов регуляции по схеме синтез — распад — ресинтез. Синтез белков, включая ферменты, регулируется с помощью механизмов репрессии, индукции и позитивного контроля. Напротив, регуляция активности самих ферментов происходит по принципу обратной связи, заключающейся в ингиби-ровании конечным продуктом. Известно также регулирование путем химической модификации ферментов. В регуляции активности клеток принимают участие гормоны, обеспечивающие химическую регуляцию.
Любое повреждение молекул ДНК, вызванное физическими или химическими факторами воздействия, может быть восстановлено с помощью одного или нескольких ферментативных механизмов, что представляет собой саморегуляцию. Она обеспечивается за счет действия контролирующих генов и в свою очередь обеспечивает стабильность генетического материала и закодированной в нем генетической информации.
Специфичность взаимоотношений со средой.
Организмы живут в условиях определенной среды, которая для них служит источником свободной энергии и строительного материала. В рамках термодинамических понятий каждая живая система (организм) представляет собой «открытую» систему, позволяющую взаимно обмениваться энергией и веществом в среде, в которой существуют другие организмы и действуют абиотические факторы. Следовательно, организмы взаимодействуют не только между собой, но и со средой, из которой они получают все необходимое для жизни. Организмы либо отыскивают среду, либо адаптируются (приспосабливаются) к ней. Формами адаптивных реакций являются физиологический гомеостаз (способность организмов противостоять факторам среды) и гомеостаз развития (способность организмов изменять отдельные реакции при сохранении всех других свойств). Адаптивные реакции определяются нормой реакции, которая генетически детерминирована и имеет свои границы. Между организмами и средой, между живой и неживой природой существует единство, заключающееся в том, что организмы зависят от среды, а среда изменяется в результате жизнедеятельности организмов. Результатом жизнедеятельности организмов является возникновение атмосферы со свободным кислородом и почвенного покрова Земли, образование каменного угля, торфа, нефти и т. д.
Обобщая сведения о свойствах живого, можно заключить, что клетки представляют собой открытые изотермические системы, которые способны к самосборке, внутренней регуляции и к самовоспроизведению. В этих системах осуществляется множество реакций синтеза и распада, катализируемых ферментами, синтезируемыми внутри самих клеток.
Свойства, перечисленные выше, присущи только живому. Некоторые из этих свойств обнаруживаются и при исследовании тел неживой природы, однако у последних они характеризуются совершенно другими особенностями. Например, кристаллы в насыщенном растворе соли могут «расти». Однако этот рост не имеет тех качественных и количественных характеристик, которые присущи росту живого. Между свойствами, характеризующими живое, существует диалектическое единство, проявляющееся во времени и пространстве на протяжении всего органического мира, на всех уровнях организации живого.
В организации живого в основном различают молекулярный, клеточный, тканевой, органный, организменный, популяционный, видовой, биоценотический и глобальный (биосферный) уровни. На всех этих уровнях проявляются все свойства, характерные для живого. Каждый из этих уровней характеризуется особенностями, присущими другим уровням, но каждому уровню присущи собственные специфические особенности.
Молекулярный уровень
. Этот уровень является глубинным в организации живого и представлен молекулами нуклеиновых кислот, белков, углеводов, липидов, и стероидов, находящихся в клетках и, как уже отмечено, получивших название биологических молекул.
Размеры биологических молекул характеризуются довольно значительным разнообразием, которое определяется занимаемым ими пространством в живой материи. Самыми малыми биологическими молекулами являются нуклеотиды, аминокислоты и сахара. Напротив, белковые молекулы характеризуются значительно большими размерами. Например, диаметр молекулы гемоглобина человека составляет 6,5 нм.
Биологические молекулы синтезируются из низкомолекулярных предшественников, которыми являются окись углерода, вода и атмосферный азот и которые в процессе метаболизма превращаются через промежуточные соединения возрастающей молекулярной массы (строительные блоки) в биологические макромолекулы с большой молекулярной массой (рис. 42). На этом уровне начинаются и осуществляются важнейшие процессы жизнедеятельности (кодирование и передача наследственной информации, дыхание, обмен веществ и энергии, изменчивость и др.).
Физикохимическая специфика этого уровня заключается в том, что в состав живого входит большое количество химических элементов, но основной элементарный состав живого представлен углеродом, кислородом, водородом, азотом. Из групп атомов образуются молекулы, а из последних формируются сложные химические соединения, различающиеся по строению и функциям. Большинство этих соединений в клетках представлено нуклеиновыми кислотами и белками, макромолекулы которых являются полимерами, синтезированными в результате образования мономеров, и соединения последних в определенном порядке. Кроме того, мономеры макромолекул в пределах одного и того же соединения имеют одинаковые химические группировки и соединены с помощью химических связей между атомами их неспецифических частей (участков).
Все макромолекулы универсальны, т. к. построены по одному плану независимо от их видовой принадлежности. Являясь универсальными, они одновременно и уникальны, ибо их структура неповторима. Например, в состав нуклеотидов ДНК входит по одному азотистому основанию из четырех известных (аденин, гуанин, цитозин и тимин), вследствие чего любой нуклеотид или любая последовательность нуклеотидов в молекулах ДНК неповторимы по своему составу, равно как неповторима также и вторичная структура молекулы ДНК. В состав большинства белков входит 100-500 аминокислот, но последовательности аминокислот в молекулах белков неповторимы, что делает их уникальными.
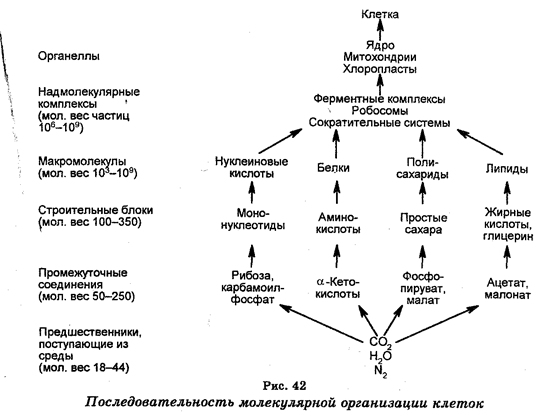
Объединяясь, макромолекулы разных типов образуют надмоле-кулярные структуры, примерами которых являются нуклеопроте-иды, представляющие собой комплексы нуклеиновых кислот и белков, липопротеиды (комплексы липидов и белков), рибосомы (комплексы нуклеиновых кислот и белков). В этих структурах комплексы связаны нековалентно, однако нековалентное связывание весьма специфично. Биологическим макромолекулам присущи непрерывные превращения, которые обеспечиваются химическими реакциями, катализируемыми ферментами. В этих реакциях ферменты превращают субстрат в продукт реакции в течение исключительно короткого времени, которое может составлять несколько миллисекунд или даже микросекунд. Так, например, время раскручивания двухцепочечной спирали ДНК перед ее репликацией составляет всего лишь несколько микросекунд.
Биологическая специфика молекулярного уровня определяется функциональной специфичностью биологических молекул. Например, специфичность нуклеиновых кислот заключается в том, что в них закодирована генетическая информация о синтезе белков. Этим свойством не обладают другие биологические молекулы.
Специфичность белков определяется специфической последовательностью аминокислот в их молекулах. Эта последовательность определяет далее специфические биологические свойства белков, т. к. они являются основными структурными элементами клеток, катализаторами и регуляторами различных процессов, протекающих в клетках. Углеводы и липиды являются важнейшими источниками энергии, тогда как стероиды в виде стероидных гормонов имеют значение для регуляции ряда метаболических процессов.
Специфика биологических макромолекул определяется также и тем, что процессы биосинтеза осуществляются в результате одних и тех же этапов метаболизма. Больше того, биосинтезы нуклеиновых кислот, аминокислот и белков протекают по сходной схеме у всех организмов независимо от их видовой принадлежности. Универсальными являются также окисление жирных кислот, глико-лиз и другие реакции. Например, гликолиз происходит в каждой живой клетке всех организмов-эукариотов и осуществляется в результате 10 последовательных ферментативных реакций, каждая из которых катализируется специфическим ферментом. Все аэробные организмы-эукариоты обладают молекулярными «машинами» в их митохондриях, где осуществляется цикл Кребса и другие реакции, связанные с освобождением энергии. На молекулярном уровне происходят многие мутации. Эти мутации изменяют последовательность азотистых оснований в молекулах ДНК.
На молекулярном уровне осуществляется фиксация лучистой энергии и превращение этой энергии в химическую, запасаемую в клетках в углеводах и других химических соединениях, а химической энергии углеводов и других молекул — в биологически доступную энергию, запасаемую в форме макроэнергетических связей АТФ. Наконец, на этом уровне происходит превращение энергии макроэргических фосфатных связей в работу — механическую, электрическую, химическую, осмотическую, механизмы всех метаболических и энергетических процессов универсальны.
Биологические молекулы обеспечивают также преемственность между молекулярным и следующим за ним уровнем (клеточным), т. к. являются материалом, из которого образуются надмолекуляр-ные структуры. Молекулярный уровень является «ареной» химических реакций, которые обеспечивают энергией клеточный уровень.
Клеточный уровень
. Этот уровень организации живого представлен клетками, действующими в качестве самостоятельных организмов (бактерии, простейшие и другие), а также клетками многоклеточных организмов. Главнейшая специфическая черта этого ^уровня заключается в том, что с него начинается жизнь. Будучи способными к жизни, росту и размножению, клетки являются ос-иовной формой организации живой материи, элементарными еди-Вицами, из которых построены все живые существа (прокариоты и эукариоты). Между клетками растений и животных нет принципиальных различий по структуре и функциям. Некоторые различия касаются лишь строения их мембран и отдельных органелл. Заметные различия в строении есть между клетками-прокариотами и клетками организмов-эукариотов, но в функциональном плане эти различия нивелируются, ибо везде действует правило «клетка от клетки». Надмолекулярные структуры на этом уровне формируют мембранные системы и органеллы клеток (ядра, митохондрии и др.).
Специфичность клеточного уровня определяется специализацией клеток, существованием клеток в качестве специализированных единиц многоклеточного организма. На клеточном уровне происходит разграничение и упорядочение процессов жизнедеятельности в пространстве и во времени, что связано с приуроченностью функций к разным субклеточным структурам. Например, у клеток эукариотов значительно развиты мембранные системы (плазматическая мембрана, цитоплазматическая сеть, пластинчатый комплекс) и клеточные органеллы (ядро, хромосомы, центриоли, митохондрии, пластиды, лизосомы, рибосомы).
Мембранные структуры являются «ареной» важнейших жизненных процессов, причем двухслойное строение мембранной системы значительно увеличивает площадь «арены». Кроме того, мембранные структуры обеспечивают отделение клеток от окружающей среды, а также пространственное разделение в клетках многих биологических молекул. Мембрана клеток обладает высокоизбирательной проницаемостью. Поэтому их физическое состояние позволяет постоянное диффузное движение некоторых из содержащихся в них молекул белков и фосфолипидов. Помимо мембран общего назначения в клетках существуют внутренние мембраны, которые ограничивают клеточные органеллы.
Регулируя обмен между клеткой и средой, мембраны обладают рецепторами, которые воспринимают внешние стимулы. В частности, примерами восприятия внешних стимулов являются восприятие света, движение бактерий к источнику пищи, ответ клеток-мишеней на гормоны, например, на инсулин. Некоторые из мембран одновременно сами генерируют сигналы (химические и электрические).'Замечательной особенностью мембран является то, что на них происходит превращение энергии. В частности, на внутренних мембранах хлоропластов происходит фотосинтез, тогда как на внутренних мембранах митохондрии осуществляется окислительное фосфорилирование.
Компоненты мембран находятся в движении. Построенным главным образом из белков и липидов, мембранам присущи различные перестройки, что определяет раздражимость клеток — важнейшее свойство живого.
Тканевой уровень
представлен тканями, объединяющими клетки определенного строения, размеров, расположения и сходных функций. Ткани возникли в ходе исторического развития вместе с многоклеточ-ностью. У многоклеточных организмов они образуются в процессе онтогенеза как следствие дифференциации клеток. У животных различают несколько типов тканей (эпителиальная, соединительная, мышечная, нервная, а также кровь и лимфа). У растений различают меристематическую, защитную, основную и проводящую ткани. На этом уровне происходит специализация клеток.
Органный уровень
. Представлен органами организмов. У простейших пищеварение, дыхание, циркуляция веществ, выделение, передвижение и размножение осуществляются за счет различных орга-нелл. У более совершенных организмов имеются системы органов. У растений и животных органы формируются за счет разного количества тканей. Для позвоночных характерна цефализация, защищающаяся в сосредоточении важнейших центров и органов чувств в голове.
Организменный уровень
. Этот уровень представлен самими организмами — одноклеточными и многоклеточными организмами растительной и животной природы. Специфическая особенность орга-низменного уровня заключается в том, что на этом уровне происходит декодирование и реализация генетической информации, создание структурных и функциональных особенностей, присущих организмам данного вида. Организмы уникальны в природе, потому что уникален их генетический материал, детерминирующий развитие, функции и взаимоотношение их с окружающей средой.
Популяционный уровень
. Растения и животные не существуют изолированно; они объединены в популяции. Создавая надорганиз-менную систему, популяции характеризуются определенным генофондом и определенным местом обитания. В популяциях начинаются и элементарные эволюционные преобразования, происходит выработка адаптивной формы.
Видовой уровень.
Этот уровень определяется видами растений, животных и микроорганизмов, существующими в природе в качестве живых звеньев. Популяционный состав видов чрезвычайно разнообразен. В составе одного вида может быть от одной до многих тысяч популяций, представители которых характеризуются самым различным местообитанием и занимают разные экологические ниши. Виды представляют собой результат эволюции и характеризуются сменяемостью. Ныне существующие виды не похожи на виды, существовавшие в прошлом. Вид является также единицей классификации живых существ.
Биоценотический уровень.
Представлен биоценозами — сообществами организмов разной видовой принадлежности. В таких сообществах организмы разных видов в той или иной мере зависят один от другого. В ходе исторического развития сложились биогеоценозы (экосистемы), которые представляют собой системы, состоящие из взаимозависимых сообществ организмов и абиотических факторов среды. Экосистемам присуще динамическое (подвижное) равновесие между организмами и абиотическими факторами. На этом уровне осуществляются вещественно-энергетические круговороты, связанные с жизнедеятельностью организмов.
Биосферный (глобальный) уровень.
Этот уровень является высшей формой организации живого (живых систем). Он представлен биосферой. На этом уровне осуществляется объединение всех вещественно-энергетических круговоротов в единый гигантский биосферный круговорот веществ и энергии.
Между разными уровнями организации живого существует диалектическое единство, живое организовано по типу системной организации, основу которой составляет иерархичность систем. Переход от одного уровня к другому связан с сохранением функциональных механизмов, действующих на предшествующих уровнях, и сопровождается появлением структуры и функций новых типов, а также взаимодействия, характеризующегося новыми особенностями, т. е. связан с появлением нового качества.
1. В чем заключается всеобщий методологический подход к пониманию сущности жизни? Когда он возник и в связи с чем?
2. Можно ли определить сущность жизни? Если да, то в чем заключается это определение и каковы его научные обоснования?
3. Возможна ли постановка вопроса о субстрате жизни?
4. Назовите свойства живого. Укажите, какие из этих свойств характерны для неживого и какие только для живого.
5. Какое значение для биологии имеет подразделение живого на уровни организации? Имеет ли такое подразделение практическое значение?
6. Какими общими чертами характеризуются разные уровни организации живого?
7. Почему нуклеопротеиды считают субстратом жизни и при каких условиях они выполняют эту роль?
8. Какое содержание вкладывают в понятия «мертвое» и «неживое»?
Верная Д. Возникновение жизни М.: Мир. 1969. 391 стр.
Опарин А. В. Материя, жизнь, интеллект. М.: Наука. 1977. 204 стр
Пехов А. П. Биология и научно-технический прогресс. М.: Знание. 1984. 64 стр.
Karcher S. J. Molecular Biology. Acad. Press. 1995. 273 pp.
Murphy M. P., O'Neill L. A. (Eds.) What is Life? The Next Fifty Years. Cambridge University Press. 1995. 203 pp.
Клетка — это основная единица живого (биологической активности), ограниченная полупроницаемой мембраной и способная к самовоспроизведению в среде, не содержащей живых систем. Жизнь начинается с клетки. Вне клеток нет жизни.
Первые исследования клеток восходят к XVII в., и, вероятно, принадлежат англичанину Роберту Гуку (1635-1703). Рассматривая под примитивным микроскопом срезы пробки (1665 г.), он обнаружил, что они состоят из ячеек, названных им клетками (от лат. cellula — ячейка, клетка). В дальнейшем ячеистое строение многих растений микроскопически наблюдали итальянец М. Маль-пиги (1628-1694) и англичанин Н. Грю (1641-1712), однако то, что они видели, сейчас мы называем клеточной стенкой клеток растений. В 1675 г. голландец А. Левенгук (1632-1723) впервые с помощью простого микроскопа увидел одноклеточные организмы (бактерии).
В 1825 г. чех Ян Пуркинье (1787-1869) увидел и описал внутреннее содержимое клетки, назвав его протоплазмой (от греч. protos первый, plasma — образование), а в 1831 г. англичанин Р. Броун (Г773-1858) обнаружил ядро клетки (от лат. nucleus, греч. сагуоn).
Важнейшим этапом в изучении клеток явились работы, обеспечивавшие фактическую основу для создания клеточной теории. В 1838 г. немецкий ботаник М. Шлейден (1804-1881) пришел к выводу, что ткани растений состоят из клеток, тогда как немецкий зоолог Т. Шванн (1810-1882) в 1839 г. к аналогичному выводу пришел, изучая строение клеток животных. Опираясь на данные о том, что клетки животных и растений имеют ядра, М. Шлейден и Т. Шванн в 1838—1839 гг. сформулировали клеточную теорию, содержавшую ряд важнейших положений, а именно:
а) Организмы состоят из клеток и продуктов их жизнедеятельности, причем клетки являются главной структурной единицей растений и животных;
б) Размножение клеток лежит в основе роста животных и растений.
Выдающийся вклад в последующее развитие клеточной теории принадлежит Р. Вирхову (1821-1902), сформулировавшему в 1855 г. очень важное положение «cellula e cellula» («каждая клетка из клетки»), означающее, что клетка может возникнуть лишь из предсу-. ществующей клетки и что других путей появления клеток не существует. Это положение имело не только фундаментальное значение, но и практическое, т. к. означало начало разработки основ клеточной патологии.
В дальнейшем важнейший вклад в развитие клеточной теории был обеспечен открытием хромосом и наблюдениями в 1879—1883 гг. деления клеток путем митоза (В. Флеминг, 1844-1905; В. Рут 1850-1924 и другие). Уже к концу XIX в. были описаны хромосомы, определено их гаплоидное и диплоидное число у ряда организмов, а также были определены и получили название фазы митоза. Тогда же состоялся синтез цитологии и генетики, а также вычленение самостоятельной проблематики под названием «Биология клетки».
В начале XX в. (1903) Р. Гертвиг (1850-1937) формулирует закон постоянства ядерно-плазменного отношения, а в 1905 г. Дж. Фармер и Дж. Мур вводят в научную литературу термин «мейоз», что способствовало лучшему пониманию деления и развития клеток. Но особенно прогресс учения о клетке был обеспечен введением в практику исследований фазово-контрастной и электронной микроскопии, а затем и метода меченых атомов. Уже в 50-е гг. нашего века были получены электронно-микроскопические изображения почти всех структур клетки.
Современный этап в развитии клеточной теории характеризуется дальнейшим обоснованием ее положений на основе результатов, полученных при изучении тонкого строения клеток, синтеза нуклеиновых кислот и белков, а также регуляции активности генов. Окончательное подтверждение получило важнейшее положение клеточной теории о том, что клетка является элементарной структурно-функциональной единицей живого, вне которой нет жизни, т. e. клетка является элементарной единицей структуры и функции многоклеточного организма. Клетки являются высокоорганизованными дифференцированными образованиями, а размножение клеток обеспечивает физическую основу генетической непрерывности между родительскими клетками и дочерними клетками. Установлено, что активность организмов зависит от активности его клеток и что рост, развитие и дифференцировка тканей зависят от образования новых клеток. Через клетки происходит поглощение, превращение, запасание и использование веществ и энергии. Структуры клеток являются ареной, на которой осуществляются многочисленные биологические реакции, в частности, ферментация, дыхание, фотосинтез, дупликация хромосом, причем эти процессы имеют место как у одноклеточных организмов, так и в клетках многоклеточных организмов. Можно сказать, что жизнь многоклеточных организмов основывается на жизни их клеток.
Таблица 2
Основные свойства прокариотических и эукариотических клеток
| Свойство
|
Прокариоты
|
Эукариоты
|
| Капсула
|
имеется у отдельных видов
|
отсутствует
|
| Клеточная стенка
|
имеется
|
имеется в клетках растений, отсутствует в клетках животных
|
| Плазматическая мембрана
|
имеется
|
имеется
|
| Ядерная мембрана
|
отсутствует
|
имеется
|
| Количество хромосом
|
одна или две
|
от нескольких до многих
|
| Количество групп сцепления
|
одна или две
|
от нескольких до многих
|
| Химический состав хромосом
|
ДНК
|
нуклеопротеид
|
| Митохондрии
|
отсутствуют
|
имеются
|
| Деление
|
прямое
|
непрямое (митоз)
|
В настоящее время различают прокариотические и эукариоти-ческие клетки. Прокариотическими являются одноклеточные организмы из мира растений, представленные в основном бактериями. Напротив, эукариотическими являются в основном одноклеточные организмы животной природы, а также клетки большинства (если не всех) многоклеточных животных и растений. Как прокариотические, так и эукариотические клетки характеризуются значительным структурно-функциональным разнообразием, что определяет между ними существенные различия (табл. 2).
Всеобъемлющим современным подходом к изучению клеток является системно-структурный подход.
Для изучения клеток используют микроскопическую технику в виде световой, фазово-контрастной, ультрафиолетовой, люминесцентной и электронной микроскопии. Последняя используется в сочетании с техникой ультратонких срезов. С целью получения трехмерных изображений клеток используют сканирующие электронные микроскопы. Для документации поведения живых клеток используют замедленную киносъемку.
В цитологических исследованиях очень эффективны цитохи-мические методы, основанные на том, что определенные реактивы (краски) избирательно окрашивают химические вещества цитоплазмы, а также ауторадиография, которая заключается во введении в клетки радиоактивных изотопов фосфора (32
Р), углерода (14
С) и водорода (3
H) с последующим обнаружением их клеточной локализации с помощью фотоэмульсий.
Для выделения клеточных компонентов используют дифференциальное центрифугирование, а для разделения биологических молекул — хроматографию и электрофорез. Рентгеноструктурный анализ позволяет определять пространственное расположение молекул различных веществ, расстояние между отдельными молекулами, объем, форму и другие свойства молекул. Метод ядерного магнитного резонанса позволяет исследования химической природы вещества.
Для изучения клеток используют также биохимические, генетические и иммунологические методы в сочетании с культивированием клеток на искусственных питательных средах. В последние годы в исследованиях клеток широко используют методы генетической инженерии.
Для прокариотических клеток характерна довольно простая структурно-функциональная организация. Вероятно, среди прока-риотов наиболее примитивно устроены микоплазмы, которые известны тем, что являются паразитами растений или возбудителями некоторых респираторных заболеваний человека и домашних животных. Считают также, что около '/д лабораторных культур соматических клеток заражены этими организмами.
Микоплазменные клетки имеют овальную форму, а их размеры составляют около 0,1-0,25 нм в диаметре (рис. 43). Для них характерно наличие тонкой наружной плазматической мембраны (толщина — около 8 нм), которая окружает цитоплазму, содержащую молекулу ДНК, достаточную для кодирования около 800 разных белков, РНК разных типов, рибосом диаметром порядка 20 нм. В их цитоплазме содержатся различные включения в виде белков, гранул ли-пидов и других соединений. Из-за недостаточной жесткости клеточной мембраны микоплазмы проходят через бактериальные фильтры.
Более сложными Прокариотическими клетками являются бактерии, цианобактерии и одноклеточные водоросли.
 Как отмечено в § 4, бактерии имеют разную форму, начиная от палочек и заканчивая округлыми формами микроскопических размеров. Размеры одиночной клетки Е. coli (рис. 44) составляют 1— 3 мкм в длину и 0,5—0,8 мкм в диаметре, объем — около 1 нм8
, а масса равна Ю-12
г. Как отмечено в § 4, бактерии имеют разную форму, начиная от палочек и заканчивая округлыми формами микроскопических размеров. Размеры одиночной клетки Е. coli (рис. 44) составляют 1— 3 мкм в длину и 0,5—0,8 мкм в диаметре, объем — около 1 нм8
, а масса равна Ю-12
г.
Одиночная клетка Е. coli окружена трехслойной клеточной оболочкой толщиной порядка 40 нм, представляющей собой «мешок» или «конверт», в котором заключено клеточное содержимое в виде, примерно, 2´10-18
г белка, 6´10-
16
г ДНК и 2´10-14
г РНК (в основном рибосомной РНК). В бактериальной клетке синтезируется около 2000 разных белков, большинство которых содержится в цитоплазме. Концентрация одних белков составляет 10-8
М, тогда как других — порядка 2´10-4
М (от 10 до 200 000 молекул на клетку).
«Конверт» состоит из трех частей (рис. 45), из которых две части представлены наружным и внутренним слоями, являющимися наружной и внутренней мембранами (соответственно) и построенными в основном из липополисахаридов. Внешняя поверхность наружной мембраны в основном состоит из липополисахаридов, которые прикрепляются к располагающимся там же липидам. В состав наружной мембраны входят также белки. Внутренняя мембрана, называемая цитоплазматической, состоит из многих белков, включенных в двойной фосфолипидный слой. Иногда мембраны образуют складки, называемые мезосомами. Предполагают, что они принимают участие в репликации бактериальной клетки.
Третьей частью «конверта» является пептидогликановый слой, который непосредственно представляет собой клеточную стенку толщиной порядка 40 нм, лежащую между наружной и внутренней ци-топлазматическими мембранами. Определяя форму бактериальной клетки, пептидогликановый слой в химическом плане является одиночной сложной молекулой, содержащей полисахаридные цепи, связанные с короткими пептидами. Внешняя мембрана прикреплена к пептидогликановому слою большим количеством (10е
) молекул ли-попротеида, белковый конец которых ковалентно прикреплен к ди-аминопимелиновой кислоте пептидогликана, тогда как их липид-ный конец «спрятан» во внешней мембране. На долю мембран и клеточной стенки приходится около 20% всего клеточного белка.
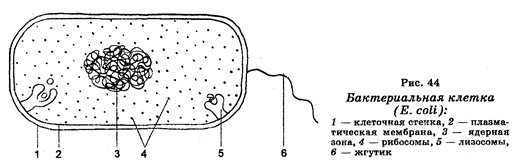

Пространство между наружной и внутренней мембраной, содержащее пептидогликановый слой, носит название периплазма-тического пространства. Растворение клеточных стенок сопровождается образованием так называемых протопластов, сохраняющих лишь внутреннюю мембрану. Эти структуры широко используют в экспериментальной работе.
На поверхности клеточной стенки у бактерий многих видов могут быть жгутики или жгутики и пили, а снаружи от клеточной стенки может иметься также капсула, как, например, у пневмококков. Эти структуры имеют диагностическое значение.
Основное вещество-бактерий представлено цитоплазмой, являющейся раствором белка, концентрация которого составляет 200 мг/мл. В цитоплазме бактерий имеется ядерная область, которую из-за отсутствия мембраны называют нуклеотидом. В этой области обнаруживают волокна диаметром 3—5 нм, представляющие собой скрученные двойные цепи одиночной кольцевой молекулы ДНК. Эти цепи ДНК рассматривают в качестве одиночной хромосомы. В большинстве случаев у бактерий действительно обнаруживают по одной кольцевой хромосоме, однако у бактерий ряда видов найдено по две кольцевых хромосомы (Rhodobacter sphaeroides, Brucella melitensis, Leptospira interrogans, Pseudomonas cepaeia), различающихся по размерам (одна из них является большой, другая малой), у Agrobacterium tumefaciens одна из двух хромосом является линейной.
Секвенирование ДНК ряда прокариотических организмов показало, что их геномы, вопреки простоте, характеризуются уникальностью. Например, Mycoplasma gallinarum обладает геномом, размер которого составляет 580 килооснований, тогда как Haemophilus influencae обладает геномом в 1,8 мегаоснований, но геномы этих прокариотов функционально различны. У Н. influencae количество кодирующих районов (генов) составляет 1743, из которых 1007 кодируют аминокислотный и липидный метаболизмы, биосинтез кофакторов и клеточного «конверта», синтез нуклеотидов и белков, репликацию и транскрипцию ДНК, продукцию энергии и транспорт веществ, причем на контроль метаболизма приходится 10% ДНК, транскрипции и трансляции — 17% ДНК, транспорта веществ — 12% ДНК и синтеза белков клеточного «конверта» — 8% ДНК. В противоположность Н. influencae, у которой аминокислотный биосинтез контролируется 68 генами, у М. galliriatum аминокислотный биосинтез контролируется всего лишь одним геном. Микоплазмы этого вида не имеют генов для цитохромов и ферментов цикла три-карбоновых кислот. Но они обладают рядом генов, которые кодируют адгезин, позволяющий им прекрепляться к соматическим клеткам животных и человека, в организмах которых они паразитируют.
В цитоплазме бактерий содержатся также рибосомы, которых очень много (по одним подсчетам около 10 000 на клетку, по другим — 15 000-30 000). Масса каждой рибосомы Е. coli составляет 2 х 7 х 10е
дальтон, а состав определяется 65% рибосомной РНК и 35% белками. Белковая часть представлена примерно 50 различными белками. В цитоплазме содержатся различные включения в виде гранул (капель) жира, гликогена, липидов, серы. У В. megaterium в цитоплазме содержится очень много гранул поли-р-оксимальной кислоты. В бактериальных клетках обнаруживают также гранулы высокополимерной фосфорной кислоты (метахроматические гранулы).
Сходным образом организованы и одноклеточные водоросли.
Наиболее сложная организация присуща эукариотическим клеткам животных и растений. Строение клеток животных и растений характеризуется принципиальным сходством, но форма, размеры и масса их чрезвычайно разнообразны и зависят от того, является ли организм одноклеточным или многоклеточным. Например, диа-томовые водоросли, эвгленовые, дрожжи, миксомицеты и простейшие являются одноклеточными эукариотами, тогда как организмы подавляющего большинства других типов являются многоклеточными эукариотами, количество клеток у которых составляет от нескольких (например, у некоторых гельминтов) до миллиардов (у млекопитающих) на организм. Организм человека состоит из около 10 различных клеток, которые различаются между собой по осуществляемым ими функциям.
В случае человека насчитывают более 200 типов разных клеток. Наиболее многочисленными клетками в организме человека являются эпителиальные клетки, среди которых различают орого-вевающие клетки (волос и ногтей), клетки, обладающие всасывательной и барьерной функциями (в желуд очно-кишечном тракте, мочеполовых путях, роговице, влагалище и других системах органов), клетки, выстилающие внутренние органы и полости (пневмо-циты, серозные клетки и многие другие). Различают клетки, обеспечивающие метаболизм и накопление резервных веществ (гепатоциты, жировые клетки). Большую группу составляют эпителиальные и соединительнотканные клетки, секретизирующие внеклеточный матрикс (амилобласты, фибробласты, остеобласты и другие) и гормоны, а также сократительные клетки (скелетных и сердечных мышц, радужной оболочки и других структур), клетки крови и иммунной системы (эритроциты, нейтрофилы, эозинофилы, базофилы, Т-лимфоциты и другие). Существуют также клетки, выполняющие роль сенсорных преобразователей (фоторецепторы, осязательные, слуховые, обонятельные, вкусовые и другие рецепторы). Значительное число клеток представлено нейронами и гли-альными клетками центральной нервной системы. Существуют также специализированные клетки хрусталика глаза, пигментные клетки и питающие клетки, далее следует назвать подовые клетки. Известны и многие другие типы клеток человека.
В природе не существует некой типичной клетки, ибо все они характеризуются чрезвычайным разнообразием. Тем не менее все эукариотические клетки существенно отличаются от прокариотических клеток по ряду свойств и прежде всего по объему, форме и размерам. Объем большинства эукариотических клеток превышает объем прокариотов в 1000-10 000 раз. Такой объем прокариотических клеток связан с содержанием в них различных органелл, осуществляющих всевозможные клеточные функции. Для эукариотических клеток характерно также наличие большого количества генетического материала, сосредоточенного в основном в относительно большом количестве хромосом, что обеспечивает им большие возможности в дифференцировке и специализации. Не менее важной особенностью эукариотических клеток является то, что им присуща компартментализация, обеспеченная наличием внутренних мембранных систем. В результате этого многие ферменты локализуются в определенных компартментах. Например, почти все ферменты, катализирующие синтез белков в животных клетках, локализованы в рибосомах, тогда как ферменты, катализирующие синтез фосфолипидов, в основном сосредоточены на клеточной ци-топлазматической мембране. В отличие от прокариотических клеток в эукариотических клетках имеется ядрышко.
Эукариотические клетки по сравнению с прокариотическими обладают более сложной системой восприятия веществ из окружающей среды, без чего невозможна их жизнь. Существуют и другие различия между эукариотическими и прокариотическими клетками.
Форма клеток бывает самой разнообразной и часто зависит также от выполняемых ими функций. Например, многие простейшие имеют овальную форму, тогда как эритроциты являются овальными дисками, а мышечные клетки млекопитающих вытянуты. Размеры эукариотических клеток являются микроскопическими (табл. 3).
Некоторые виды клеток характеризуются значительными размерами. Например, размеры нервных клеток у крупных животных достигают нескольких метров в длину, а у человека — до 1 метра. Клетки отдельных тканей растений достигают нескольких миллиметров в длину.
Считают, что чем крупнее организм в пределах вида, тем крупнее его клетки. Однако для родственных видов животных, различающихся по размерам, характерны и сходные по размерам клетки. Например, у всех млекопитающих сходны по размерам эритроциты.
Клетки различаются также и по массе. Например, одиночная клетка печени (гепатоцит) человека весит 19-9
г.
Соматическая клетка человека (типичная эукариотическая клетка) представляет собой образование, состоящее из множества структурных компонентов микроскопических и субмикроскопических размеров(рис. 46).
Использование электронной микроскопии и других методов позволило установить чрезвычайное разнообразие в структуре как оболочки и цитоплазмы, так и ядра. В частности, был установлен мембранный принцип строения внутриклеточных структур, исходя из которого различают ряд структурных компонентов клетки, а именно:
Таблица 3
Средние размеры клеток животных и растений
| Происхождение клетки
|
Диаметр (в мкм)
|
Объем (в мкм3
)
|
| Клетка печени человека
|
20
|
4000
|
| Малая клетка тимуса
|
6
|
120
|
| Клетка меристемы (корешок лука)
|
17
|
2600
|
| Клетка паренхимы плода растения
|
1000
|
1 х 108
|
1. Мембранная система.
2. Цитоплазматический мат-рикс (основное вещество клеток).
3. Клеточные органеллы (внутриклеточные компартменты).
4. Клеточные включения.
Мембранная система. Эта система представлена клеточной плазматической (цитоплазматической) мембраной, цитоплазматической (эндоплазматической) сетью (рети-кулом) и пластинчатым комплексом Гольджи.
а) Плазматическая (цитоплаз-матическая) мембрана
имеет толщину 8-12 нм и состоит из трех слоев, два из которых являются белковыми слоями толщиной по 3 нм каждый, а третий (внутренний) — двойным фосфолипидным слоем толщиной 6 нм (рис. 47). Плазматическая мембрана является полупроницаемой структурой. Через нее в клетку входят питательные вещества и выходят все «отходы» (продукты секреции). Она создает барьер проницаемости. В результате этого плазматическая мембрана регулирует обмен различными веществами между клеткой и внешней средой. В плазматической  мембране содержатся многие важные ферменты, системы активного транспорта ионов натрия и калия при помощи АТФазы, а также системы транспорта аминокислот. мембране содержатся многие важные ферменты, системы активного транспорта ионов натрия и калия при помощи АТФазы, а также системы транспорта аминокислот.
На поверхности плазматической мембраны обнаруживают ряд специальных образований в виде микроворсинок и ресничек. Микроворсинки очень часты в эпителии кишечника и почек. Реснички — это своеобразные выросты цитоплазмы. У эритроцитов мембрана является гладкой (элементарная мембрана). У некоторых одноклеточных организмов-эукариотов плазматическая мембрана также содержит реснички (микроворсинки), различные выпячивания, впячивания и выросты, переходящие в пузырьки. На внешней поверхности клеток животных обнаружены гликопротеиды как компоненты плазматической мембраны.
 Предполагают, что поверхностные гликопротеиды обеспечивают адгезионную способность клеток в тканях, и, следовательно, слипание однотипных клеток. В мембранах эритроцитов содержится гликопротеид, получивший название гликофорина (м. м. 30 000). Этот гликопротеид состоит из 130 аминокислотных остатков и большого количества (60% всей молекулы) остатков сахара. Кроме того, в эритроцитарной мембране содержится белок спектрин, молекулы которого формируют скелет мембраны. Предполагают, что поверхностные гликопротеиды обеспечивают адгезионную способность клеток в тканях, и, следовательно, слипание однотипных клеток. В мембранах эритроцитов содержится гликопротеид, получивший название гликофорина (м. м. 30 000). Этот гликопротеид состоит из 130 аминокислотных остатков и большого количества (60% всей молекулы) остатков сахара. Кроме того, в эритроцитарной мембране содержится белок спектрин, молекулы которого формируют скелет мембраны.
У клеток растений наружной структурой служит жесткая клеточная стенка, построенная из молекул целлюлозы, создающих очень прочные волокна, погруженные в матрикс из других поли-сахаридов и полимерного вещества лигнина. Клетки наружних слоев растений иногда покрыты очень тонким слоем восковидно-го вещества. На поверхности плазматических мембран имеются электрически заряженные группы, из-за которых поддерживается разность электрических потенциалов на мембранах. На поверхности плазматических мембран имеются также специфические рецепторы (участки распознания) для гормонов и других соединений. Кроме того, здесь же локализованы особые рецепторы, ответственные за индивидуальную тканевую совместимость. Считают, что рецепторные участки формируются гликопротеидами и ганглиозидами.
б) Цитоплазматическая (эндоплазматическая) сеть (рети-кулум) представлена пронизывающими однослойными мембранными полостями (трубочками, цистернами, вакуолями) разных размеров, заполненными белковыми гранулами (рис. 48).
Открыта К. Портером в 1945 г. Толщина трубочек и других структур этой сети равна 5-6 нм.
Различают гранулярный (шероховатый) эндоплазматический ретикулум, который выстлан множеством рибосом диаметром порядка 21-25 нм и молекулярной массой 4 х 106
, служащих центрами синтеза молекул белков, и агранулярный (гладкий) эндоплазматический ретикулум, на котором нет рибосом, но на котором синтезируются липиды и углеводы. Степень насыщенности гранулярной эндоплазмати-ческой сети рибосомами определяет степень интенсивности синтеза белков. У человека и животных агранулярной сетью богаты клетки коры надпочечников, яичников и семенников, печени, скелетных мышц.
Цитоплазматическая сеть без перерыва соединена с цитоплаз-матической мембраной, ядерной мембраной и пластинчатым комплексом Гольджи. Это позволяет синтезируемым белкам проходить в комплекс Гольджи, откуда после специальной обработки они выводятся из клетки или идут на построение лизосом.
Плазматическая мембрана, мембрана эндоплазматической сети, а также ядер, митохондрий и хлоропластов (см. ниже) представляют собой чрезвычайно сложные структуры, обладающие рядом важ-.нейших биологических свойств. Многие мембраны содержат ферменты, транспортные системы, с помощью которых осуществляется перенос молекул питательных веществ и неорганических ионов внутрь клеток и внутри клеток, а также вывод из клеток продуктов жизнедеятельности. Мембранные структуры способны к самовосстановлению, если в них по каким-то причинам возникают повреждения.
В процессе искусственного растирания или гомогенизации клеток с экспериментальными целями образуются мелкие частицы диаметром 50—150 нм, состоящие из фрагментов эндоплазматическо-го ретикулума и плазматической мембраны. Эти структуры получили название микросом и их широко используют в лабораторной работе для решения тех или иных вопросов молекулярной организации клеток.
 в) Комплекс Гольджи
. Этот комплекс, называемый еще пластинчатым, был открыт итальянцем Камилло Гольджи еще в 1898 г. Он присутствует во всех клетках, кроме эритроцитов и сперматозоидов, и представляет собой систему дискообразных однослойных мембран (мембранных пузырьков или цистерн), локализующихся рядом с гладким эндоплазмати-ческим ретикулом и ядром (рис. 49). Часто в клетках обнаруживают несколько таких комплексов (диктиосом), размеры которых составляют 30-60 нм. Структурными молекулами, поддерживающими структуру цистерн, являются ферменты, вовлеченные в процессинг оли-госахаридов, белки, являющиеся аутоантителами, а также белки, являющиеся компонентами цитоскелета. Основная функция комплекса Гольджи заключается в том, что он является местом упаковки (уплотнения) белков, поступающих с рибосом, а также присоединения к белкам углеводов (образования гликопротеидов), а к полисахаридам — сульфатных групп с последующим транспортом их к другим клеточным структурам или за пределы клетки (экзоцитоз). В клетках печени этот комплекс участвует в выделении в кровь липопротеидов. Как отмечено выше, он участвует также и в формировании лизосом. в) Комплекс Гольджи
. Этот комплекс, называемый еще пластинчатым, был открыт итальянцем Камилло Гольджи еще в 1898 г. Он присутствует во всех клетках, кроме эритроцитов и сперматозоидов, и представляет собой систему дискообразных однослойных мембран (мембранных пузырьков или цистерн), локализующихся рядом с гладким эндоплазмати-ческим ретикулом и ядром (рис. 49). Часто в клетках обнаруживают несколько таких комплексов (диктиосом), размеры которых составляют 30-60 нм. Структурными молекулами, поддерживающими структуру цистерн, являются ферменты, вовлеченные в процессинг оли-госахаридов, белки, являющиеся аутоантителами, а также белки, являющиеся компонентами цитоскелета. Основная функция комплекса Гольджи заключается в том, что он является местом упаковки (уплотнения) белков, поступающих с рибосом, а также присоединения к белкам углеводов (образования гликопротеидов), а к полисахаридам — сульфатных групп с последующим транспортом их к другим клеточным структурам или за пределы клетки (экзоцитоз). В клетках печени этот комплекс участвует в выделении в кровь липопротеидов. Как отмечено выше, он участвует также и в формировании лизосом.
Цитоплазматический матрикс.
Этот структурный компонент является основным веществом (цитоплазмой, гиалоплазмой) клетки. Первые электронномикроскопические изображения цитоплазмы были получены шведским ученым Ф. Шестрандом еще в 1955 г. Различают эктоплазму — вещество, располагающееся ближе к цитоплазматической мембране (твердое тело) и эндоплазму, отстоящую к центру клетки от эктоплазмы (более жидкое состояние) и представляющую собой цитозоль. Консистенция цитозоля приближается к гелю, В нем растворены многие ферменты и белки, обеспечивающие связывание и транспорт питательных веществ, микроэлементов и кислорода. Здесь же находятся аминокислоты и нуклеотиды, а также различные метаболиты (промежуточные продукты биосинтеза и распада макромолекул). Наконец, в цитозоле присутствуют различные коферменты, а также АТФ, АДФ, ионы ряда неорганических солей (K+
, Mg2+
, Са2+
, C1-
, НСО-2
3
, НРО4
-2
), тРНК.
В цитоплазме содержатся микрофиламенты (нити) толщиной 4-5 нм и микротрубочки, представляющие собой полые цилиндрические структуры диаметром 25 нм, а также филаменты промежуточных размеров. Эти структуры составляют жесткую конструкцию (каркас) в клетке, называемую цитоскелетом и определяющую внешний вид и форму клеток. Микрофиламенты состоят из белка, сходного с сократительным белком актином.
Объединяясь, микрофиламенты формируют пучки, в которые входят дополнительные белки (анкерин, спектрин и другие). Основная функция микрофиламентов заключается в обеспечении сократительных процессов клеток, в упрочении мембран. Микротрубочки построены из белков a- и b-тубулина, а также g-тубулина. Для микротрубочек характерен ряд функций. Формирование микротрубочек происходит в интерфазе клеток в так называемых центрах организации микротрубочек (ЦОМ), которые «окружают» центриоли, в результате чего предполагают, что центриоли являются ЦОМ (рис. 50). В каждом ЦСУ содержится по 10-13 молекул g-тубулина и примерно по 7 молекул других белков, включая ди-мер a/b-тубулина. Эти белки формируют структуру, которая образует микротрубочный «ансамбль». Их значение до конца не выяснено, но предположительно заключается в том, что они обеспечивают перемещение клеточных органелл, включая хромосомы, внутри клеток.
В составе цитоплазмы обнаруживают ферменты, полисахариды, АТФ, тРНК, ионы Са, Na, К и других химических элементов.
 Основная функция цитоплазматиче-ского матрикса заключается в том, что он является внутренней средой клетки, поддерживающей мембранные системы, орга-неллы и включения. В нем осуществляются гликолиз, активация аминокислот и другие реакции. Цитоскелет выполняет опорную функцию. Микрофиламенты способствуют упрочению мембранной системы, а микротрубочки, как отмечено выше, обеспечивают перемещение клеточных органелл и транспорт химических соединений из одних отсеков клетки в другие. Цитоскелет имеет значение также в делении клетки. Основная функция цитоплазматиче-ского матрикса заключается в том, что он является внутренней средой клетки, поддерживающей мембранные системы, орга-неллы и включения. В нем осуществляются гликолиз, активация аминокислот и другие реакции. Цитоскелет выполняет опорную функцию. Микрофиламенты способствуют упрочению мембранной системы, а микротрубочки, как отмечено выше, обеспечивают перемещение клеточных органелл и транспорт химических соединений из одних отсеков клетки в другие. Цитоскелет имеет значение также в делении клетки.
Клеточные органеллы
. Эти структуры представлены ядром, хромосомами, ядрышком, центриолями, митохондриями, рибо-сомами, лизосомами. Они характерны за некоторыми исключениями как для клеток животных, так и для клеток растений.
а) Ядро в клетке имеет форму и размеры, зависящие от формы, размеров и функций клетки. В клетках эукариотических организмов содержится, как правило, по одному ядру, реже по два и более. В зрелом состоянии эритроциты млекопитающих и клетки ситовидных трубочек покрытосеменных растений лишены ядер, тогда как клетки скелетных мышц позвоночных и млечных сосудов растений являются многоядерными. Для инфузорий характерно наличие двух ядер — одно небольшое (микронуклеус) и одно крупное политенное (макронуклеус).
Обычно ядро имеет округлую, палочковидную, четковидную вытянутую и другие формы. Размеры его колеблются от 2 до 100 мкм, а объем составляет около 65 мкм3
. Особенно крупные ядра характерны для половых клеток (размером до 500 мкм). Отношение объема ядра к объему цитоплазмы называют ядерно-плазменным отношением, которое у клеток всех типов обычно постоянно.
Строение ядра характеризуется чрезвычайной сложностью, хотя принципиально одинаково в клетках всех эукариотических организмов. В случае животных клеток ядро располагается в центре клетки. Напротив, в клетках растений оно имеет пристеночную локализацию. Однако положение, форма и размеры ядра могут меняться в зависимости от интенсивности метаболизма. В ядре  содержатся хромосомы и ядрышки (см. ниже). Благодаря им ядро плотно заполнено ДНК, РНК и белками. Содержимое ядра иногда называют нуклеоплазмой или кариоплазмой. Она отделена от цитоплазмы ядерной мембраной, построенной из двух слоев (наружного и внутреннего) толщиной по 7 нм каждый и имеющей поры диаметром 40—100 нм. Поры занимают около 5% площади ядра. Через ядерные поры синтезируемая в ядре РНК выходит в цитоплазму, где она участвует в трансляции генетической информации (синтезе белков). содержатся хромосомы и ядрышки (см. ниже). Благодаря им ядро плотно заполнено ДНК, РНК и белками. Содержимое ядра иногда называют нуклеоплазмой или кариоплазмой. Она отделена от цитоплазмы ядерной мембраной, построенной из двух слоев (наружного и внутреннего) толщиной по 7 нм каждый и имеющей поры диаметром 40—100 нм. Поры занимают около 5% площади ядра. Через ядерные поры синтезируемая в ядре РНК выходит в цитоплазму, где она участвует в трансляции генетической информации (синтезе белков).
Ядерная мембрана и ядерные поры объединены с мембранной системой клетки, в результате чего клетка, по существу, канализирована различными проходами, обеспечивающими двухстороннее движение веществ в клетке, начиная от плазматической мембраны. В порах обнаружены глобулярные и фибриллярные белковые структуры. Количество пор зависит от вида клеток и обычно увеличивается в 8-периоде. Между мембранными слоями имеется так называемое перинуклеарное пространство шириной 20—60 нм. Наружная ядерная мембрана часто переходит в эндоплазматическую сеть. Когда клетки делятся, ядерная оболочка исчезает, а после деления восстанавливается. Ядерное вещество (кариоплазма) — плотный коллоид.
Таблица 4
Количество хромосом в соматических клетках
отдельных организмов
| Вид животных м растений
|
Количество хромосом (в диплоидиом наборе)
|
| Малярийный плазмодий
|
Plasmodium malariae
|
2
|
| Плодовая мушка
|
Drosophila melanogaster
|
8
|
| Комнатная муха
|
Musca domestica
|
12
|
| Сазан
|
Cyrpinus carpio
|
104
|
| Лягушка зеленая
|
Rana esculenta
|
26
|
| Голубь
|
Columba livia
|
80
|
| Кролик
|
Lepus cuniculus
|
44
|
| Шимпанзе
|
Antropopitecus pan
|
48
|
| Человек
|
Homo sapiens
|
46
|
| Картофель
|
Solanum tuberosum
|
48
|
| Рожь
|
Secale cereale
|
14
|
| Пшеница (мягкая)
|
Triticum
|
42
|
| Кукуруза
|
Zea mays
|
20
|
Важнейшая функция ядра заключается в том, что оно является центром управления в клетке, в нем происходит синтез ДНК, РНК и ядерных белков.
б) Хромосомы
. Хромосомы располагаются в ядре. Их название введено в литературу В. Вальдейлом в 1883 г. Они имеют форму палочек, нитей, петель. Для каждой хромосомы характерны индивидуальные особенности, касающиеся длины и положения перетяжки (центромеры) по длине хромосомы. Каждая из митотичес-ких индивидуальных хромосом состоит из двух сестринских хроматид, удерживаемых центромерой. В зависимости от локализации центромер различают метацентрические, субметацентричес-кие, акроцентрические и телоцентрические хромосомы (рис. 51). Количество хромосом постоянно в ядрах соматических клеток, где они находятся в парах. Диплоидный набор хромосом называют кариотипом (от греч. caryon — ядро, type — форма, тип). Для разных организмов характерны разные по количеству диплоидные наборы хромосом (табл. 4). В зависимости от строения хромосомы человека классифицируют на группы (рис. 52).
 Когда ядра соматических метафазных клеток окрашивают основными красителями, то при микроскопии препаратов в ядерной зоне обнаруживают глыбки и гранулы окрашенного вещества, получившего название хроматина. Именно из этого вещества во время деления клеток организуются хромосомы. При изучении таких микроскопических препаратов отмечают, что некоторые районы хромосом окрашены очень интенсивно, Другие слабее. Интенсивно окрашивающийся хроматин получил название гетерох-роматина, менее окрашивающийся — эухроматина. Последний содержит области, на которых очень активно синтезируется РНК. Когда ядра соматических метафазных клеток окрашивают основными красителями, то при микроскопии препаратов в ядерной зоне обнаруживают глыбки и гранулы окрашенного вещества, получившего название хроматина. Именно из этого вещества во время деления клеток организуются хромосомы. При изучении таких микроскопических препаратов отмечают, что некоторые районы хромосом окрашены очень интенсивно, Другие слабее. Интенсивно окрашивающийся хроматин получил название гетерох-роматина, менее окрашивающийся — эухроматина. Последний содержит области, на которых очень активно синтезируется РНК.
Исследование хромосом типа ламповых щеток, содержащихся в овоцитах многих животных и политенных хромосомах, обнаруживаемых в клетках насекомых, показало, что они имеют петельную структуру. Эта структура характерна, вероятно, для организации хроматина у всех организмов.
В клетках многих видов организмов гетерохроматин концентрируется в основном в районах, фланкирующих центромеры.
Половые хромосомы часто почти полностью представлены гете-рохроматином. Гетерохроматином богаты районы повторов последовательностей ДНК (см. гл. X).
Когда гетерохроматин исследуют под электронным микроскопом, то отмечают, что он построен из плотноупакованных хрома-тиновых нитей диаметром 25 нм. Эухроматин составлен из менее плотноупакованных нитей, но такого же диаметра. Гетерохроматин сохраняется в высококонденсированной форме на протяжении всего клеточного деления, тогда как эухроматин менее конденсирован и невидим в интерфазе при исследовании хромосом в световом микроскопе. Имеющиеся данные показывают, что большинство исследованных генов эукариот локализовано в эухроматиновых (менее конденсированных) районах хромосом, тогда как гетерохроматин (высококонденсированный) генетически не активен.
Химический состав хроматина довольно прост. Он состоит из ДНК (15%), белков (75%), а также некоторого количества РНК (10%). Различают хромосомные белки двух типов — основные белки (положительно заряженные при нейтральном рН), называемые гисто-нами, и гетерогенные белки, по большей части кислые (отрицательно заряженные при нейтральном рН), называемые негистоновыми. Молекулярная масса гистонов составляет 11 000—21 000. Гистоны являются основными белками по той причине, что содержат позитивно заряженные аминокислоты аргинин и лизин в количестве от 20% до 30% аминокислотных остатков белка. Группы R аргинина и лизина позволяют гистонам действовать в качестве поликатионов и взаимодействовать с ДНК, которая является полианионной благодаря ее отрицательно заряженным фосфатным группам. Другими словами, по причине протонирования боковых групп R-остатков аргинина и лизина гистоны, соединяясь с отрицательно заряженной двухцепочечной ДНК, образуют комплексы ДНК-гистоны.
Гистоны присутствуют в хроматине всех высших эукариотов в количествах, эквивалентных количествам ДНК. У всех высших растений и животных они представлены пятью главными белками, обозначенными символами HI, Н2а, Н2в, НЗ и Н4. Количество ДНК и гистонов в хромосомах эквивалентно. Они присутствуют в клетках всех типов, кроме сперматозоидов некоторых организмов. В последних обнаруживают основные белки, называемые протами-нами. Все пять гистонов присутствуют в молярных соотношениях 1 HI : 2 Н2а : 2 Н2в : 2 НЗ : 2 Н4. Для них характерны различия по молекулярной массе и аминокислотному составу. Гистон HI богат лизином (29%), гистоны Н2а и Н2в богаты как лизином, так и аргинином, а гистоны НЗ и Н4 богаты аргинином. Для гистонов НЗ и Н4 характерно сходство последовательностей аминокислот; для других гистонов заметное сходство отсутствует. Данные о константности гистонов Н2а, Н2в, НЗ и Н4 в клетках организмов разных видов позволяют предположить, что гистоны имеют значение в упаковке ДНК. Кроме того, они не специфически вовлечены в регуляцию экспрессии генов (см. гл. XII).
Негистоновая белковая фракция хроматина состоит из различных очень гетерогенных белков. Состав этой фракции широко варьирует в клетках разных организмов даже одного и того же вида. Предполагают, что негистоновые белки также принимают участие в регуляции экспрессии специфических генов.
 Каждая хроматина содержит одиночную гигантскую двухцепочечную молекулу ДНК, вытянутую по всей длине хромосомы. Это заключение основано на данных о размерах самых больших молекул ДНК в ядрах клеток дрозофилы и других организмов, а также на данных об общем количестве ДНК, присутствующей в самых больших хромосомах дрозофилы. Наиболее крупные молекулы ДНК, выделенные из хромосом, имеют контурную длину порядка 1,2 см, что соответствует молекулярной массе 2,4—3,2 х 1010
, т. е. от a
/y до '/4 размера самой крупной молекулы соответствующей хромосомы. На концах хромосом человека имеются повторы азотистых оснований, получившие название теломер. Количество теломерных повторов связывают с возрастом организма. Каждая хроматина содержит одиночную гигантскую двухцепочечную молекулу ДНК, вытянутую по всей длине хромосомы. Это заключение основано на данных о размерах самых больших молекул ДНК в ядрах клеток дрозофилы и других организмов, а также на данных об общем количестве ДНК, присутствующей в самых больших хромосомах дрозофилы. Наиболее крупные молекулы ДНК, выделенные из хромосом, имеют контурную длину порядка 1,2 см, что соответствует молекулярной массе 2,4—3,2 х 1010
, т. е. от a
/y до '/4 размера самой крупной молекулы соответствующей хромосомы. На концах хромосом человека имеются повторы азотистых оснований, получившие название теломер. Количество теломерных повторов связывают с возрастом организма.
Количество ДНК и гистонов в хроматине эквивалентно. Будучи специфически связанными с ДНК, гистоны принимают участие в формировании основных структурных субъединиц хроматина (хромосом) — нуклеосом (рис. 53), которые являются элементарными единицами организации хроматина. Каждая нуклеосома представлена сегментом ДНК диаметром около 30 нм и длиной 146-240 пар оснований, намотанной 1,8 раз вокруг гистонового стержня, состоящего из пары гистонов. Нуклеосома закручена в сложную нить 2 раза при участии гистона HI в качестве кросслинкера, что дает структуру с упаковочным отношением 25:1, близким к интерфазному хроматину. Чтобы сформировалась митотическая хромосома из нити длиной 30 нм, необходима дальнейшая компактизация с помощью специфических негистоновых белков, приводящая к формированию скелета хромосомы. Установлено, что нуклеосомы являются репрессорами инициации транскрипции в эукариотических клетках. Нарушение структуры нуклеосом ведет к освобождению транскрипции от репрессии.
В метафазных хромосомах ДНК организуется в петли длиной 30 000-40 000 пар оснований, а каждая петля хроматина прикрепляется к гистоновому скелету хромосомы на его основании в результате воздействия ДНК + белок или ДНК + РНК.
Гистон HI связан с линкерной ДНК, принимая участие в стабилизации сверхскрученных ДНК между нуклеосомами, но способ этой связи полностью еще не выяснен. Длина сегментов ДНК (линкеров), которые соединяют нуклеосомы, составляет от 15 до 100 нуклеотидных пар в зависимости от типа клетки. Результаты Х-дифракции кристаллов изолированных нуклеосом указывают на то, что они имеют дисковидную структуру и состоят из двух симметричных половин. ДНК каждой половины намотана в форме суперспирали на поверхность гистоновых стержней.
Для хромосом характерно наличие отдельных сайтов, которые определают их хрупкость, что создает условия для нарушений структуры хромосом, сопровождающихся хромосомными мутациями (см. гл. X).
в) Ядрышко
— это очень мелкая структура диаметром 1—5 мкм, которая локализуется в ядре. Количество ядрышек бывает разным — от одного до нескольких. Центральная часть в ядрышке представлена фибриллярной частью (цепи, ДНК- ядрышковые организаторы и рибонуклеопротеиды) и гранулярной (формирующиеся субъединицы рибосом). Ядрышко является местом синтеза рРНК. Матрицей для синтеза рРНК является ДНК ядрышкового организатора. Синтезируемая рРНК объединяется с белком и образующиеся рибонуклеопротеиды служат затем материалом для сборки из них субъединиц рибосом. Последние через ядерные поры направляются в цитоплазму, участвуя там в формировании рибосом, на которых происходит затем синтез белков.
г) Центриоли
представляют собой гранулярные цилиндры диаметром около 0,15 мкм и длиной 0,5 мкм, локализующиеся парами в клетке около ядра. Эти пары называют диплосомами (центросома-ми). Структурно они представляют систему мембранных трубочек. Диплосома окружена другими микротрубочками, называемыми центросферой. Обе эти структуры образуют клеточный центр.
Функцией клеточного центра является организация цитоскеле-та клеток. Кроме того, центриоли участвуют в делении клеток, расходясь к полюсам клетки. Микротрубочки, протягивавшиеся взаимно, образуют митотическое веретено.
 В клетках растений этой органел-лы нет. В клетках растений этой органел-лы нет.
д) Митохондрии присутствуют во всех клетках организмов, которые используют для дыхания кислород. В одной клетке может находиться 50-5000 митохондрий. Они имеют форму палочек, нитей или гранул (рис. 54), а их размеры достигают 7 мкм.
Митохондрии образованы двумя мембранами — наружной и внутренней, между которыми образуется пространство шириной 10—20 нм. Внутренняя мембрана формирует складки (кристы), которые погружены в матрикс, представляющий собой молекулярное содержимое митохондрий. Наружная мембрана проницаема для низкомолекулярных соединений. Проникновение веществ во внутреннее пространство (матрикс) митохондрий контролируется внутренней мембраной. Содержимое митохондрии составляют белки, фосфолипиды, ДНК, РНК и рибосомы, которых значительно меньше, чем рибосом, локализованных в цитоплазме. Наличие этих структур обеспечивает собственную митохондриальную белоксинтезирующую систему. Митохондрии способны к самовоспроизведению путем деления или почкования в период 8-фазы и других фаз клеточного цикла.
Митохондрии имеются также у простейших. В частности, ти-паносомы содержат по одной митохондрии.
Митохондрии представляют собой «силовые станции», в которых происходят основные процессы получения и накопления энергии путем окисления молекул пищи (окислительное фосфорилиро-вание) для образования АТФ. В матриксе содержатся ферменты, а реакции окисления проходят на поверхности внутренней мембраны. Энергетические потребности клеток определяют уровень размножения митохондрии.
В клетках растений вместо митохондрии содержатся пластиды. Среди пластид различают хлоропласты, которые содержат хлорофилл, лейкопласты (бесцветные пластиды), в которых происходит накопление крахмала, и хромопласты, в которых происходит синтез пигмента плодов. У пурпурных бактерий имеются хроматофоры.
Для хлоропластов характерно мембранное строение (рис. 55). Встречаясь в клетках в количестве около 40 экземпляров на клетку, они имеют своеобразную форму двояковыпуклых линз и размеры  5-10 мкм. Каждый хлоропласт окружен двойной мембраной. Наружная мембрана является гладкой, состоя из белковых субъединиц, а внутренняя — складчатой. Внутренняя мембрана содержит хлорофилл, а также ферменты, синтезирующие АТФ и органические соединения с помощью АТФ. Кроме того, в хлоро-пластах имеется некоторое количество ДНК и РНК небольших по размерам рибосом, что обеспечивает собственную хло-ропластную белоксинтезирующую систему. 5-10 мкм. Каждый хлоропласт окружен двойной мембраной. Наружная мембрана является гладкой, состоя из белковых субъединиц, а внутренняя — складчатой. Внутренняя мембрана содержит хлорофилл, а также ферменты, синтезирующие АТФ и органические соединения с помощью АТФ. Кроме того, в хлоро-пластах имеется некоторое количество ДНК и РНК небольших по размерам рибосом, что обеспечивает собственную хло-ропластную белоксинтезирующую систему.
Для пластид характерны переходы из одного типа в другой. Например, зеленые хлоропласты листьев переходят в хромопласты (осенью, когда изменяется окраска листьев), а лейкопласты зеленеющих клубней картофеля переходят в хлоропласты.
е) Рибосомы
представляют собой гранулы, расположенные в ци-топлазматическом матриксе и связанные с мембранами цитоплаз-матической сети. Их размеры составляют 15—Збнм в диаметре. Они построены на одну треть из белка и на две трети из РНК (двух субъединиц). Количество рибосом в клетках очень большое. Например, в одной бактериальной клетке (кишечной палочке) их содержится около 6000 экземпляров. Рибосомы образуют группы, называемые полирибосомами.
Функции рибосом заключаются в том, что на них осуществляется синтез белков.
ж) Лизосомы
представляют собой мельчайшие пузырьковидные образования, окруженные однослойной мембраной и содержащие ферменты. Их размеры составляют 0,2-0,8 мкм. Лизосомы обеспечивают изоляцию гидролитических ферментов клетки.
В лизосомах обнаружено более 40 различных гидролитических ферментов (протеиназ, нуклеаз, липаз, гликоидаз и др.), осуществляющих внутриклеточное расщепление макромолекул, проникающих в клетки посредством фагоцитоза и пиноцитоза. Лизосомы чрезвычайно разнообразны по строению и частным функциям. Предполагают, что они образованы мембранами комплекса Гольджи и представляют собой систему удаления из клеток конечных продуктов обмена. Повреждение лизосом сопровождается растворением клеток. Следовательно, благодаря лизосомам происходит защита клеток от собственных ферментов. В эволюционном плане они являются аналогами пищеварительных вакуолей одноклеточных организмов.
з) Пероксисомы
(иногда их называют микротельцами) представляют собой образованные цистернами эндоплазматической сети пузырьки размером 0,3-1,5 мкм, ограниченные однослойной мембраной. Эти пузырьки содержат каталазу и некоторые оксидазы. Обильными по содержанию пероксисом являются эпителии печени и почек. Функция пероксисом заключается в том, что в них происходит разрушение клеточных перекисей и холестерина. Кроме того они содержатся также в клетках зеленых листьев растений. Известны также глиоксисомы, содержащиеся в проростках семян масличных растений.
Включения
. В клетках животных включения представлены жировыми каплями, гранулами гликогена, зимогеновыми гранулами (депо ферментов в клетках поджелудочной железы). Особенно богаты жиром у млекопитающих клетки соединительной ткани. Гликогена много в клетках поперечно-полосатых мышц, печени и в нейронах. Встречаются также белковые включения в цитоплазме яйцеклеток, печени, в теле простейших. Следует отметить, что некоторые клетки животных содержат в качестве включений пигменты. Например, в клетках коркового вещества надпочечников накапливаются липохромы. Пигментами являются также гемоглобин и меланин, а также ретинин (в зрительном пурпуре сетчатки глаза).
В клетках растений, например, таких как картофель, злаковые, наиболее частым включением является крахмал. В качестве включений в клетках растений встречаются также жиры, содержание которых очень большое в семенах масличных культур. Очень широко распространены пигменты.
Участки (компартменты) цитоплазматического матрикса, лишенные мембран и клеточных органелл, получили название цитозолл.
Клеточное вещество является сложным полифазным коллоидом, т. е. представляет собой систему из двух несмешивающихся фаз. Одна из этих фаз структурно является цитоплазматическим матриксом и выполняет роль водной фазы с переходами от жидкого до твердого состояния, тогда как другая является мембранной системой и выполняет роль относительно жидкой фазы. Цитоплазма практически бесцветна, имеет характер раствора.
В элементном составе клетки насчитывают более 70 элементов, среди которых наиболее частыми являются кислород, углерод, водород, азот. На долю кислорода приходится 65% общей массы, на долю углерода — 18%, водорода — 10%, азота — 3%. После этих элементов идут кальций, фосфор, калий, сера, натрий, хлор. Поскольку все эти элементы встречаются в клетках в большом количестве, часто их называют макроэлементами. Марганец, медь, иод, кобальт и другие, обнаруживаемые в микроколичествах, называют микроэлементами.
Химические элементы, входящие в состав клеток и обладающие биологическими функциями, называют биогенными.
Как правило, содержание катионов и анионов отличается от содержания их в той среде, в которой находятся клетки. Например, концентрация К+
в мышечных клетках в несколько десятков раз выше, чем в крови. Концентрация солей в клетках определяет буферность ее содержимого, под которой понимают уровень концентрации водородных ионов в клетках (рН).
Химические элементы участвуют в построении вещества клеток в виде ионов (катионов и анионов) или химических соединений. Важными являются катионы К+
, Na+
, Са2+
, Mg2+
. Что касается анионов, то ими являются Н2
РО4
-
, С1-
и HCO3
-
.
Соединяясь химическими связями, группы атомов образуют так называемые малые органические молекулы, которыми являются аминокислоты, нуклеотиды, сахара и жирные кислоты. Из этих малых молекул в клетках формируются макромолекулы в виде белков, нуклеиновых кислот, углеводов и липидов.
Клетки построены как из неорганических, так и органических соединений.
Неорганическими соединениями клетки являются вода и минеральные соли.
Вода составляет около 70% массы клетки. У отдельных организмов, например медуз, содержание превышает 95% . Для водных организмов характерна чрезвычайная приспособленность к воде, поскольку высокая теплоемкость воды представляет собой непрерывно действующий «тепловой» буфер, который обеспечивает в общем постоянную температуру тела независимо от температуры воздуха. В случае растений очень прочное сцепление молекул воды способствует переносу растворенных питательных веществ из корней в листья при транспирации. Наконец, на молекулярном уровне у наземных и водных животных, равно как и у растений, вода определяет ряд важных свойств макромолекул.
Таблица 4а
Химические элементы в клетках человека
(в % к сухой массе)
| Кислород
|
65
|
Марганец
|
0,0003
|
| Углерод
|
18
|
Медь
|
0,0002
|
| Водород
|
10
|
Иод
|
0,0004
|
| Азот
|
3
|
Кобальт
|
Следы
|
| Кальций
|
1,5
|
Цинк
|
Следы
|
| Фосфор
|
1
|
Молибден
|
Следы
|
| Калий
|
0,35
|
Никель
|
Следы
|
| Сера
|
0,25
|
Алюминий
|
Следы
|
| Натрий
|
0,15
|
Барий
|
Следы
|
| Хлор
|
0,15
|
Стронций
|
Следы
|
| Магний
|
0,05
|
Титан
|
Следы
|
| Железо
|
0,004
|
Литий и др.
|
Следы
|
Табл. 4б
Основные химические соединения в клетках человека
(в % к сырой массе)
| Вода
|
75-85
|
| Белки
|
10-20
|
| Нуклеиновые кислоты
|
1-2
|
| Липиды
|
1-5
|
| Углеводы
|
0,2-2
|
В теле человека вода составляет 60%, из которой 40% приходится на внутриклеточную, а 20% — на экстраклеточную воду. Плазма крови содержит 5% экстраклеточной воды (рис. 56).
Вода имеет исключительно важное значение для жизнедеятельности клеток, представляя собой среду, в которой осуществляются важнейшие реакции, лежащие в основе синтеза и распада веществ. Кроме того она является растворителем различных химических веществ. Вещества, хорошо растворимые в воде получили название гидрофильных (от греч. hydros — вода, phileo — люблю),  плохо растворимые называют гидрофобными (от греч. hydros- вода, phobos — боязнь). В воде хорошо растворяются хлористый натрий, сахара, простые спирты, альдегиды, кетоны. Под влиянием растворенных веществ вода может изменять свои свойства, в частности, могут изменяться температура замерзания, температура кипения, давление пара и осмотическое давление воды. Эта особенность воды имеет очень важное биологическое значение. Например, рыбы в пресной воде при температуре ее замерзания сохраняют свою активность, причем по той причине, что концентрация веществ, растворенных в крови рыб, является большей, чем в воде, и это исключает переохлаждение, а затем и замерзание их крови. плохо растворимые называют гидрофобными (от греч. hydros- вода, phobos — боязнь). В воде хорошо растворяются хлористый натрий, сахара, простые спирты, альдегиды, кетоны. Под влиянием растворенных веществ вода может изменять свои свойства, в частности, могут изменяться температура замерзания, температура кипения, давление пара и осмотическое давление воды. Эта особенность воды имеет очень важное биологическое значение. Например, рыбы в пресной воде при температуре ее замерзания сохраняют свою активность, причем по той причине, что концентрация веществ, растворенных в крови рыб, является большей, чем в воде, и это исключает переохлаждение, а затем и замерзание их крови.
Для воды характерно то, что она обладает некоторой способностью к обратимой ионизации, в ходе которой она распадается на ионы водорода (Н+
) и ионы гидроксила (ОН-
).
Для изменения концентрации ионов Н+
в любом водном растворе используют так называемую шкалу рН, с помощью которой обозначают концентрацию водородных ионов (Н+
) в водных растворах, кислотность которых находится между 1,0 МН+
и 1,0 МОН-
. Так значение рН для нейтрального раствора составляет 7,0, тогда как растворы, имеющие рН выше 7,0, — это щелочные растворы, а меньше 7,0 — это кислые растворы. Например, рН питьевой воды составляет 9,0, нашатырного спирта — 12,0, черного кофе — 5,0 лимонного сока — 2,0, а желудочного сока — 1,0. Величины рН характерны для всех внутриклеточных и внеклеточных жидкостей в организме, причем постоянство концентрации водородных ионов поддерживается буферными системами, которые у млекопитающих представлены фосфатной и бикарбонатной системами. Величины рН всех жидкостей организмов исключительно постоянны. Их изменения чрезвычайно неблагоприятны для организмов, поскольку даже небольшие сдвиги рН характеризуются значительным падением каталитической активности ферментов.
В воде под влиянием ферментов происходят реакции гидролиза (от греч. hydros — вода, lysis — расщепление) белков и других соединений. Вода принимает участие также в выведении из клеток продуктов обмена. Наконец, она поддерживает тепловой режим клетки.
Минеральные соли
входят в состав цитоплазмы. Встречаются калиевые, натриевые, магниевые соли, соли серной, соляной, фосфорной и других кислот. Важнейшая роль минеральных солей заключается в определении ими кислотно-щелочного состояния протоплазмы. Они необходимы также для размножения клеток.
Органическими (углеродсодержащими) соединениями клетки являются белки, нуклеиновые кислоты, углеводы, липиды и АТФ. Как уже отмечено, молекулы этих соединений часто называют биологическими молекулами, а из-за их крупных размеров — макромолекулами.
Поскольку все органические соединения клетки содержат углерод, то принято считать, что жизнь на Земле построена на углеродистой основе. Замечательной особенностью углерода является то, что его атомы очень легко образуют ковалентные связи с другими атомами, в результате чего он больше других элементов способен образовывать большие молекулы. До некоторой степени такой способностью, но меньшей, обладает и кремний, что явилось основанием к известным предположениям о существовании жизни на других планетах, но на кремниевой основе.
Белки
, или, как их еще называют, протеины (от греч. protos — первостепенный), являются наиболее сложными химическими соединениями, характеризующимися большой молекулярной массой. В состав всех известных белков входят углерод, водород, азот и кислород. В большинстве белков находят серу, а в некоторых белках — фосфор, железо, цинк и медь. Будучи макромолекулами, они представляют собой линейные полимеры, в которых мономерами являются аминокислоты, каждая из которых состоит из аминогруппы (-NH2
), карбоксильной группы (-СООН), атома водорода и R-группы, присоединенной к атому углерода, который называют tt-углеродным атомом. Благодаря наличию аминогрупп и карбоксильных групп аминокислоты способны реагировать друг с другом и образовывать между собой ковалентные связи. В частности, аминокислоты соединяются одна с другой посредством соединения аминогруппы одной аминокислоты с карбоксильной группой другой аминокислоты. Возникающую между аминокислотами связь называют пептидной (амидной), а несколько соединений аминокислот называют пептидом. Т.к. первоначально образующийся дипептид содержит реакционноспособные аминогруппу и карбоксильную группы, то к нему способны присоединиться другие аминокислоты, образуя полипептид (белок). Обычно цепь из трех аминокислот называют трипептидом, а цепь из многих соединенных аминокислот называют полипептидной цепью. Следовательно, пептиды — это цепочки аминокислот. Белок может состоять из одной полипептидной цепи или нескольких. Например, миоглобин состоит из одной цепи, тогда как гемоглобин — из двух цепей одного типа и двух цепей другого типа.
Для полипептидных цепей характерна неразветвленная структура. Молекула белка представляет собой, по существу, неопределенно длинные цепи аминокислот, связанные пептидными связями:
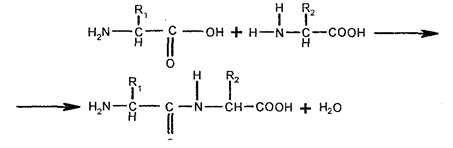
где R-группы (боковые группы, цепи) являются радикалами, каждый из которых состоит из гидроксильной (ОН-), сульфгидриль-ной (SH-) и других групп и которые являются частью молекулы. Боковые R-группы, будучи разными по структуре, электрозаряду и растворимости в воде, определяют различия между аминокислотами.
Для аминокислот характерна ассиметричность, в результате чего различают L- и D-аминокислоты. В составе клеточных белков имеются только L-аминокислоты, известно 20 L-аминокислот. Эти аминокислоты часто называют стандартными (основными, нормальными), т. к. известны и еще другие аминокислоты, которые присутствуют в организмах, но не обнаруживаются в большинстве их белков. Такие нестандартные аминокислоты встречаются в коллагене (4-гидроксипролин и 5-гидроксилизин), миозине (N-метил-лизин), в протромбине (g-карбоксиглутаминовая кислота) и в эластине (десмозин). Предполагают, что L-аминокислоты существуют около 2 млрд лет. Из них построены белки всех известных к настоящему времени организмов. Различия между разными L-аминокис-лотами определяются боковыми R-группами, присоединенными к альфауглероду. Кроме пептидов, образующих белки, существует много пептидов, встречающихся в организмах животных и человека в виде свободных соединений, не связанных с белками. Такими пеп-тида^яи являются некоторые гормоны (инсулин, глюкагон и другие). Поскольку R-группы характеризуются разной степенью полярности (разной способностью взаимодействовать с водой при рН около 7,0), то аминокислоты классифицируют на аминокислоты, содержащие неполярные R-группы (аланин, валин, лейцин, изолейцин, пролин), полярные незаряженные R-группы (глицин, серии, треонин, цисте-ин, тирозин, аспарагин, глутамин), отрицательно заряженные (кислые) R-группы (аспарагиновая и глутаминовая кислоты), положительно заряженные (основные) R-группы (лизин, аргинин, гистидин).
Белки различаются между собой по молекулярной массе, которая для большинства из них лежит в пределах 6000-1 000 000.
Белки различаются по составу на простые и сложные. Простые белки состоят только из аминокислот, содержа 50% углерода, 7% водорода, 23% кислорода, 16% азота. В состав некоторых простых белков может входить сера в небольшом количестве.
Сложные белки помимо аминокислот содержат в своем составе другие соединения как органические, так и неорганические. Эту небелковую часть молекулы сложного белка называют простетической группой. Сложными белками являются нуклеопротеиды, липопротеиды, фосфопротеиды, металлопротеиды и гликопротеиды (табл. 5).
Таблица 5
Перечень некоторых сложных белков
| Вид сложного белка
|
Простатическая группа
|
| Нуклеопротеиды
:
|
| Рибосомы
|
РНК
|
| РНК-содержащие вирусы
|
| Гликопротеиды
:
|
| g-глобулин
|
Гексозамин, галактоза, сиаловая кислота
|
| Липопротеиды
:
|
| Липопротеиды плазмы
|
Фосфолипиды, холестерин, нейтральные липиды
|
| Фосфопротеиды
:
|
| Казеин
|
Фосфатная группа + остаток серина
|
| Гемопротеиды
:
|
| Гемоглобин
|
Железопротопорфирин
|
| Цитохром
|
Железопротопорфирин
|
| Флавопротеиды
:
|
| Сукцинатдегидрогеназа
|
Флавиннуклеотид
|
| Оксидаза D-аминокислот
|
Флавиннуклеотид
|
| Металлопротеиды
:
|
| Ферритин
|
| Тирозиноксидаза
|
| Алкогольдегидрогеназа
|
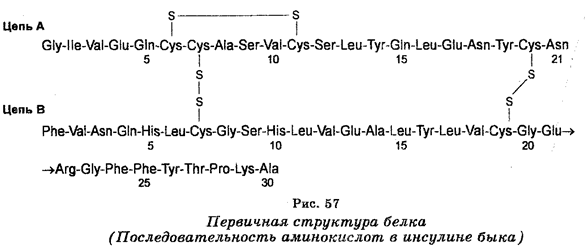
Белки различаются также по структуре, которая зависит от количества входящих в их состав аминокислот (аминокислотных остатков) и последовательности (чередования) аминокислот в поли-пептиде. Одни белки построены из одной (рибонуклеаза, лизоцим), двух (бычий инсулин), трех (хемотрипсин), четырех (гемоглобин человека) и более полипептидных цепей.
Различают первичную, вторичную, третичную и четвертичную структуру белков. Первичная структура определяется отношением одного аминокислотного остатка к другому в полипептиде, т. е. последовательностью аминокислотных остатков в полипептиде (рис. 57). Вторичная структура определяется скрученностью полипептидных цепей в виде спиралей. Третичная структура характерна для полипептидных спиральных структур, образующих клубки (фибриллы). Что касается четвертичной структуры, то она возникает, когда происходит соединение между собой нескольких молекул.
В зависимости от конформации (пространственной структуры), различают глобулярные (полипептидные цепи образуют клубки) и фибриллярные белки (полипептидные цепи формируют фибриллы). Глобулярными белками являются почти все ферменты, антитела, гормоны, гемоглобин, сывороточный альбумин, тогда как фибрил-лярными являются коллаген сухожилий и костей, кератинин волос и другие.
На долю белков в цитоплазме приходится 50-70% от общего количества органических соединений. Что же касается функций белков, то они исключительно разнообразны (табл. 6).
Белки являются, прежде всего, строительным материалом, т. к. входят в состав практически всех клеточных структур. Кроме того, обладая способностью образовывать волокна, многие белки выполняют опорную функцию. Например, основу кожи, хрящей и сухожилий у млекопитающих составляет фибриллярный белок коллаген, который при кипячении в воде превращается в желатин, а связок — эластин. Химический состав волос, ногтей (когтей) и перьев определяется в основном кератином. Шелковые нити и паутина построены из белка фибреина.
Таблица 6
Некоторые белки человека и животных и их функции
| Тип белка
|
Функция
|
| Структурные белки:
|
| Гликопротеиды
|
Образуют клеточные стенки
|
| Белки мембран
|
Образуют мембраны
|
| Белки вирусного капсида
|
Образуют вирусный капсид
|
| Коллаген
|
Образуют фиброзную соединительную ткань
|
| Эластин
|
Связки
|
| Кератин
|
Кожа, перья, ногти, копыта
|
| Ферменты
|
Катализируют реакции синтеза и распада
|
| Гормоны
:
|
| Инсулин
|
Регулирует обмен глюкозы
|
| Аденокортикотропный
|
Регулирует синтез кортикостероидов
|
| Гормон роста
|
Стимулирует рост костей
|
| Двигательные белки:
|
| Миозин
|
Нити в миофибриллах
|
| Актин
|
Движущиеся нити в миофибриллах
|
| Дипеин
|
Движение жгутиков и ресничек простейших
|
| Транспортные белки
:
|
| Гемоглобин
|
Переносит кислород (позвоночные)
|
| Гемоцианин
|
Переносит кислород (беспозвоночные)
|
| Миоглобин
|
Переносит кислород в мышцах
|
| Защитные белки:
|
| Антитела
|
Контролируют чужеродные белки
|
| Комплемент
|
Комплексируются с антителом + антигеном
|
| Фибриноген
|
Предшественник фибрина
|
| Тромбин
|
Фактор свертываемости крови
|
| Токсины
:
|
| Ботулистический токсин
|
Пищевые отравления
|
| Змеиный яд
|
Гидролиз фосфоглицеридов
|
| Рицин
|
Токсический белок клещевины
|
Многие белки являются ферментами (энзимами). Ферменты локализуются в митохондриях, цитоплазме, лизосомах, пероксисо-мах, на мембранах клеток и органелл. Они катализируют все протекающие в клетках реакции. Считают, что ферменты повышают скорость реакции минимум в 1 млн раз. Каждая реакция обеспечивается собственным ферментом. Например, липаза расщепляет жиры, амилаза расщепляет крахмал. В настоящее время известно более 2000 разных ферментов. В зависимости от катализируемых реакций их классифицируют на гидролазы (реакции гидролиза), нуклеазы (расщепление нуклеиновых кислот), трансферазы (перенос функциональных групп), оксидоредуктазы (окислительно-восстановительные реакции), липазы (образование связей за счет АТФ) и др.
Важнейшей особенностью белков является то, что в клетках многоклеточных организмов тысячи белков функционально связаны между собой и переносят информацию от плазматической мембраны к геному. Например; фермент в метаболическом пути «читает» концентрацию субстрата и продуцирует соответствующий уровень продукта, а рецептор на клеточной поверхности «читает» концентрацию его лиганда и продуцирует определенный уровень комплекса рецептор — лиганд.
Белки обладают регуляторной способностью. Те животные белки, которые являются гормонами, обладают способностью регулировать физиологические процессы, протекающие в клетках. Например, инсулин, являющийся белковым гормоном, продуцируемым клетками поджелудочной железы, регулирует в организме метаболизм глюкозы. Белковыми являются также гормоны, продуцируемые клетками гипоталамической части мозга и гипофизом и имеющие важное значение в росте и развитии организмов. Пара-тиреоидный гормон регулирует транспорт ионов Са^ и фосфатов. Однако заметим, что не все гормоны имеют белковую природу. У растений также известны отдельные белки, обладающие гормональной активностью. Например, такой активностью обладает индоли-луксусная кислота, которая стимулирует рост растений. Репрессорные белки участвуют в регуляции экспрессии генов.
Белки обладают двигательной и сократительной функциями, обеспечивая на молекулярном, уровне движение хромосом и сперматозоидов, на других уровнях — движение простейших, двигательные реакции у растений; сокращение скелетных мышц у многоклеточных животных (мышечные белки антин и миозин). Они выполняют также роль механической опоры. Например, высокая упругость кожи обусловлена наличием в ней коллагена.
Для белков характерна транспортная функция,. В частности, они являются транспортерами гормонов, аминокислот, липидов, саха-ров, ионов кислорода.
Белки являются источниками энергии, если происходит их распад до аминокислот. Теряя аминогруппы (дезаминируясь), белки становятся источником энергия во время, когда в клетках наступает истощение углеводных и липидных ресурсов.
Наконец, белками являются различные токсины, продуцируемые паразитами животной и растительной природы, а также змеиные яды и токсичные белки растений (рицин и другие), представляющие угрозу для животных и человека. В то же время важнейшей функцией белков является их защитная функция, поскольку иммунными антителами у животных и человека служат высокоспецифичные белки иммуноглобулины. Защитными белками можно считать также фибриноген и тромбин, которые из-за участия в свертывании крови предохраняют организм от ее потери. Кровь некоторых рыб Антарктики содержит специфический антифриз-ный белок, предотвращающий ее замерзание. Противовирусное соединение интерферон также имеет белковую природу. Наконец, некоторые белки обладают запасной пищевой функцией в семенах растений. Такие белки используются в качестве пищевых зародышами в начальный их период развития. Пищевыми белками являются альбумин (главный белок яиц птиц) и казеин (главный молочный белок).
Оценивая роль белков в жизни клеток, тканей и организмов, нельзя также не отметить, что они обладают видовой специфичностью, а это ведет к одному основополагающему заключению, сводящемуся к признанию положения «организмы делаются белками».
Нуклеиновые кислоты
являются органическими соединениями, содержащими углерод, кислород, водород, азот и фосфор. Различают дезоксирибонуклеиновую и рибонуклеиновую кислоты (ДНК и РНК). Важнейшая биологическая роль нуклеиновых кислот заключается в том, что они являются хранителями генетической информации (см. главу X).
Отсылая читателя к главе X, где изложены основные сведения о нуклеиновых кислотах как генетическом материале, здесь кратко рассмотрим сведения об аденозинтрифосфорной кислоте (АТФ), которая представляет собой нуклеотид, образованный присоединением к аденозинмонофосфорной кислоте (АМФ), содержащейся в РНК, двух дополнительных молекул фосфорной кислоты (НдРО^). Другими словами, в составе АТФ содержатся аденин, рибоза и три молекулы фосфорной кислоты. АТФ синтезируется в мито-хондриях.
Выдающаяся роль АТФ определяется ее чрезвычайной важностью в обеспечении клеток энергией, которая освобождается в результате воздействия на АТФ фермента АТФ-азы, сопровождаемого в начале отщеплением одной молекулы фосфорной кислоты и образованием аденозиндифосфатной кислоты (АДФ), а затем еще двух молекул фосфорной кислоты и переходом АДФ в аденозинмо-нофосфорную кислоту (АМФ). Фосфорно-кислородные связи в АТФ (их две) называют макроэргическими, обозначая их символом Р.
Синтез АТФ происходит в митохондриях.
Углеводы
— это органические соединения углерода, водорода и кислорода с общей формулой (СН2
)n
, где n — представляет собой число от трех до семи. Такое название этим соединениям дал в 1844 г. К. Шмидт (1822-1894). Содержание их в клетках очень значительно. Например, содержание крахмала доходит иногда до 90% сухой массы (картофель, семена злаковых).
Различают полисахариды (С6
Н10
О5
)n
, дисахариды (C12
H22
O11
) и простые сахара — моносахариды (С6
Н12
О6
), являющиеся малыми органическими молекулами. Две молекулы моносахарида продуцируют одну молекулу дисахарида и одну молекулу воды, тогда как из n молекул моносахарида образуется одна молекула полисахарида и (n - 1) молекул воды. Следовательно,
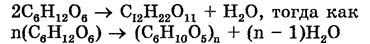
В зависимости от количества атомов углерода в молекуле моносахарида различают триозы (3 атома углерода), тетрозы (4 атома углерода), пентозы (5 атомов углерода) и гексозы (6 атомов углерода).
Среди триоз важное значение для животных и человека имеют такие моносахариды, как глицерин и его производные, молочная и пировиноградная кислоты.
Наиболее известной тетрозой является эритроза — промежуточный продукт фотосинтеза.
Пентозами являются рибоза и дезоксирибоза.
Среди гексоз важнейшее значение для организмов имеют глюкоза, являющаяся первичным источником энергии, а также фруктоза и галактоза. Глюкоза, или виноградный сахар, является основной частью ряда ди- и полисахаридов. В природных условиях в свободном состоянии она встречается в клетках практически всех растений. Что касается животных, то она также широко распространена, обнаруживаясь в структурных компонентах, в крови. В результате окисления глюкозы происходит ее распад до разных производных Сахаров, а затем и до CO2
и H2
O. При этом освобождается энергия и образуется восстановительная способность, запасаемая в молекулах АТФ и НАДФ. Фруктоза (плодовый сахар) в свободном состоянии встречается в плодах растений. Особенно много ее в сахарном тростнике и сахарной свекле, во фруктах, а также меде. Галактоза встречается в составе лактозы, которой много в молоке.
Дисахариды, трисахариды и тетрасахариды часто называют оли-госахаридами, среди которых очень важными для жизни организмов являются сахароза, лактоза и мальтоза. Сахароза, известная в быту как сахар, в больших количествах содержится в сахарном тростнике и сахарной свекле. Молекулы сахарозы состоят из остатков D-глюкозы и D-фруктозы. Лактоза, или молочный сахар, в большом количестве содержится в молоке. Она состоит из глюкозы и галактозы. Мальтоза встречается в составе крахмала и гликогена. Она состоит из двух молекул глюкозы и этим определяется ее биологическая важность.
Полисахариды обладают свойствами полимеров. Будучи образованными сотнями или даже тысячами моносахаридных единиц, они являются либо линейными полимерами (целлюлоза), либо разветвленными (гликоген).
Полисахаридами, состоящими из большого количества моноса-харидов, и наиболее известными и биологически важными у растений являются целлюлоза и крахмал, которые состоят из монотонно повторяющихся остатков D-глюкозы. Являясь основным структурным элементом клеточных стенок, целлюлоза обеспечивает прочность клеток зеленых растений. Известен также полисаха-рид хитин, содержащийся в клеточных стенках грибов и в скелете членистоногих. Он обеспечивает прочность их скелета. Считают, что целлюлоза является самым распространненным углеводом среди всех углеводов, известных на Земле. Крахмал содержится в большом количестве в клубнях картофеля и семенах злаковых (особенно кукурузы и пшеницы). Он построен из двух полимеров D-глюкозы (а-амилазы и пектина). Крахмал является резервным углеводом в клетках растений. В клетках животных содержится полисахарид гликоген, который тоже состоит из очень большого количества остатков D-глюкозы. Накапливаясь в печени, мышцах и других органах, он является источником глюкозы, поступающей в кровь. Этот углевод обнаруживают также в грибах.
Полисахариды, гиалуроновая кислота и пектины создают прослойку между клетками животных и растений, соответственно. Такой полисахарид, как гепарин синтезируется клетками легких, печени и других тканей и секретируется в кровь.
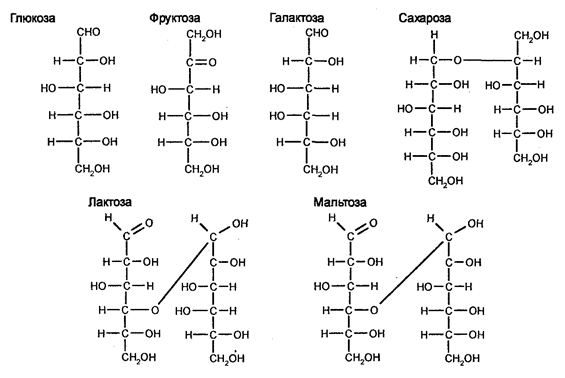
Известны соединения, представляющие собой комплекс сахаров с белками. Например, небольшие олигосахаридные группы, прикрепляясь через 0-гликоидную связь к -ОН-группам остатков се-рина, треонина, оксилизина или через N-гликозидную связь к амидному азоту аспарагина формируют гликопротеиды, обладающие рядом биологических функций (ферментативных, структурных, регулирующих). Если концевое звено полисахарида ковален-тно присоединению О-гликоидной связи к сериновому остатку в белке, то образующееся сложное соединение называют пептидо-гликаном. Это соединение обладает структурной функцией.
Углеводы обладают структурной функцией, причем самым распространенным структурным углеводом является целлюлоза. Другими структурными углеводными элементами являются гликоза-миногликаны (кислые мукополисахариды) и протеогликаны.
Углеводы являются важнейшим источником энергии в организме, которая освобождается в результате окислительно-восстановительных реакций. Установлено, что окисление 1 г углевода сопровождается образованием энергии в количестве 4,2 ккал. Целлюлоза не переваривается в желудочно-кишечном тракте позвоночных из-за отсутствия гидролизующего фермента. Она переваривается лишь в организме жвачных животных (крупный и мелкий рогатый скот, верблюды, жирафы и другие). Что касается крахмала и гликогена, то в желудочно-кишечном тракте млекопитающих они легко расщепляются ферментами-амилазами. Гликоген в желудочно-кишечном тракте расщепляется до глюкозы и некоторого количества мальтозы, но в клетках животных он расщепляется гликогенфосфорилазой с образованием глюкозо-1-фосфата. Наконец, углеводы служат своеобразным питательным резервом клеток, запасаясь в них в виде гликогена в клетках животных и крахмала в клетках растений.
Липиды
(от греч. lipos — жир), или жиры являются соединениями, состоящими из жирных кислот и глицерола. К этим липидам относят также жироподобные вещества (воска). Жирные кислоты — это органические кислоты. Наиболее встречаемыми жирными кислотами в жирах животных и растений являются пальмитиновая (CH3
(CH2
)15
COOH), стеариновая (CH3
(СH2
)16
COOH) и олеиновая (СН3
-СН2
)7
СН-СН(СН2
)7
-СООН) жирные кислоты. Одна молекула глицерола и три молекулы жирной кислоты образуют одну молекулу липида и три молекулы воды. Например, стеарин образуется в результате реакции между одной молекулой глицерола и тремя молекулами стеариновой кислоты:
Для липидов характерно то, что они не растворимы в воде. Растворителями для них являются эфир, бензин, хлороформ и другие органические растворители.
Липиды встречаются почти во всех клетках, но в основном в небольших количествах, хотя некоторые клетки содержат эти соединения в очень больших количествах, доходящих до 90% их сухой массы. Они обнаруживаются в нервной ткани, мужских половых клетках, в семенах растений. В бараньем жире глицерол связан в основном со стеариновой кислотой, тогда как в говяжьем жире — с пальмитиновой и стеариновой кислотами. Напротив, в растительных маслах и жире лошади глицерол связан с олеиновой кислотой.
Липиды в сочетании с другими соединениями образуют более сложные соединения. Например, известны фосфолипиды (глицерол + жирная кислота + фосфатная группа), липопротеиды (комплексы липидов с белками) и другие. Названные выше липиды определяют в качестве омыляемых, т. к. нагревание их совместно со щелочами сопровождается их гидролизом с образованием мыл. Между тем известны липиды, не способные образовывать мыла. Эти липиды называют неомыляемыми или стероидами, среди которых наиболее распространенными являются стеролы (стероид-ные спирты). В тканях животных наиболее часто обнаруживается холестерол.
Липиды обладают рядом важнейших свойств в жизни клеток. Прежде всего, поскольку углеводы могут переводиться в липиды, то последние выполняют роль накопителей энергии, ибо окисление липидов сопровождается выделением энергии. Например, окисление 1 г жира сопровождается выделением энергии в количестве 9,5 ккал. При окислении образуются также углекислый газ и вода. Очень тонкий слой жира в плазматической мембране клеток выполняет защитную роль.
Очень важное значение в построении клеточных структур липиды приобрели в составе фосфолипидов, которые являются одним из основных строительных материалов мембран клеток. Важную биологическую роль в жизни клеток и организмов играют также липопротеиды. Липиды способны к запасанию в организмах в больших количествах и этим обеспечивают терморегуляцию организмов, являясь материалом для образования эндогенной воды в результате его окисления, что имеет важное значение для жизни многих животных пустынь (верблюды, мелкие млекопитающие). Являясь предшественниками в синтезе ряда гормонов, они принимают участие в регуляции важных функций организмов. Воска предохраняют кожу позвоночных от воды, у птиц они придают водоотталкивающие свойства перьям. У многих видов растений воска покрывают листья. Питаясь фитопланктоном, содержащим воска, киты и лососевые рыбы используют их в качестве главного источника липидов. Помимо соединений, рассмотренных в этом параграфе, в клетках содержатся также и другие соединения. Чтобы проявлялась каталитическая активность ферментов, для многих из них необходимо присутствие кофакторов небелковой природы, которые либо непосредственно участвуют в каталитическом процессе, либо являются промежуточными переносчиками функциональных групп от субстрата непосредственно к ферменту. Если кофакторами ферментов являются органические соединения, то их называют кофермента-ми. Предшественниками многих таких органических соединений (ко-ферментов) служат витамины, которые тоже являются органическими соединениями, присутствующими в небольших количествах в клетках растений и животных и попадающих в организм человека с пищей. К настоящему времени известно более 10 различных вята-минов, которые классифицируют на водорастворимые и жирорастворимые витамины. Водорастворимыми являются витамины B2
(тимин), B2
(рибофлавин), В6
(пиридоксин), В12
никотиновая, панто-теновая и фолиевая кислоты, биотин и витамин С (аскорбиновая кислота). Жирорастворимыми являются витамины А, Д, Е и К.
Водорастворимые витамины в качестве коферментов участвуют в катализировании многих реакций, в частности таких, как декарбоксилирование a-кетокислот (витамин В1
), окислительно-восстановительных реакций (витамин В2
, никотиновая кислота), реакций гидроксилирования (витамин С), карбоксилирования (витамин К), переносе ацильных (пантотеиновая кислота) и многих других реакций.
Жирорастворимые витамины выполняют самые различные функции. В частности витамин А принимает участие в формировании зрительного процесса в виде альдегида витамина А, связанного с белком опсином, витамин Д регулирует обмен кальция, витамин Е участвует в защите липидов клеточных мембран от разрушения кислородом, а витамин К является кофактором реакций карбоксилирования.
Кофакторами ферментов являются также микроэлементы. В частности, для каталитического действия многих ферментов (цитохромоксидазы, каталазы, пероксидазы) необходимы ионы железа. Для действия цитохромоксидазы и лизиноксидазы необходима медь. Ионы Zn2+
присутствуют в НАД- и НАДФ-зависимых дегидрогена-зах. Другим ферментам необходимы ионы марганца (аргиназа), никеля (уреаза) или атомы молибдена и ванадия (флавиндегидрогена-зы). Некоторые микроэлементы участвуют в регуляторных реакциях. Например, хром участвует в регуляции усвоения глюкозы клетками животных тканей, а олово необходимо для кальцификации костей. Бор и алюминий необходимы для развития растений.
Наконец, в клетках в очень небольших количествах встречаются аминокислоты в свободном состоянии (свыше 150), которые не встречаются в составе белков.
Размножение или пролиферация (от лат. proles — потомство, ferre — нести) клеток — это процесс, который приводит к росту и обновлению клеток. Данный процесс характерен как для одноклеточных, так и многоклеточных организмов.
Клетки-организмы (одноклеточные организмы) размножаются простым делением надвое (бактерии, саркодовые), множественным делением (споровики и др.) или другим путем. Поэтому у бактерий и одноклеточных животных удвоение клеток представляет собой размножение их как самостоятельных организмов, поскольку из исходной формы (организма) образуется две новые клетки, каждая из которых является организмом. Каждая дочерняя клетка (организм) получает полную генетическую информацию, несомую исходной клеткой-организмом.
Соматические клетки многоклеточных организмов размножаются путем сложного деления, которое получило название митоти-ческого деления и которое в общем виде представляет собой механизм, посредством которого одиночные клетки репродуцируют себя. Образовавшиеся в результате деления дочерние клетки подобны исходной (материнской) клетке, отличаясь от последней лишь меньшими размерами. Однако вслед за делением дочерние клетки мгновенно начинают расти и быстро достигают размеров материнской клетки.
Биологический смысл митотического деления состоит в том, что оно является ключевым событием в точной репликации всех хромосом еще до того, как произойдет деление ядра и клетки. В результате митоза дочерние клетки после деления получают хромосомы в точно таком же количестве, какое имела их родительская (материнская) клетка. Следовательно, митотическое деление есть особый способ упорядоченного деления клеток, при котором каждая из двух дочерних клеток получает хромосомы в точно таком же количестве и точно такого же строения, что и хромосомы, которые имела материнская клетка. При каждом митозе образуется копия каждой хромосомы и действует точный механизм их распределения между дочерними клетками.
В митотическом делении клетки различают две стороны — разделение исходного ядра на два дочерних ядра (равное деление хромосом), называемое кариокинезом (от греч. caryon — ядро, kinesis — движение) и представляющее собой, по существу, хромосомный цикл, и следующее затем разделение цитоплазмы с образованием двух дочерних клеток, называемое цитокинезом (от греч. cytos — клетка, kinesis — движение) и представляющее собой цитоплазматический цикл. Каждая из дочерних клеток содержит одно дочернее ядро.
Кариокинез и цитокинез протекают синхронно, причем в кариоки-незе имеет место чередование синтеза ДНК с митозом, тогда как цитокинез чередуется с ростом клеток (удвоением в числе клеточных компонентов).
Существенной особенностью митотического деления является то, что оно в значительной мере сходно у всех организмов. Совокупность процессов, происходящих в клетке от одного деления до другого, получило название митотического цикла.
Митотический цикл состоит из двух стадий — стадии покоя или интерфазы и стадии деления или митоза (от греч. rnitos — нить), обозначаемого символом м. Термины «митоз» и «кариокинез» — синонимы. Интерфаза доступна для оценки качественно и количественно, точно так же доступен для измерения и митоз. В частности, для измерения интенсивности количества митозов используют так называемый митотический индекс, под которым понимают число митозов на 1000 клеток. Данные о митотическом индексе имеют важное практическое значение, особенно в медицинской практике (в оценке интенсивности регенерации органов, действия лекарственных веществ и т. д.).
Интерфаза предшествует митозу, и функциональное содержание ее заключается в том, что в ней происходит синтез ДНК (рис. 58), причем ее длительность составляет не менее 90% в течение всего клеточного цикла. Различают три последовательных периода интерфазы, а именно: пресинтетический, синтетический и постсинтетический.
 Пресинтетический период (G1
), который часто называют еще первым интервалом (от англ. gap — интервал), является начальным периодом интерфазы. В этот период ДНК еще не синтезируется, однако происходит накопление РНК и белков, в том числе и белков, необходимых для синтеза ДНК. Увеличивается количество митохондрий. Обычно этот период длится 12-24 часа. Пресинтетический период (G1
), который часто называют еще первым интервалом (от англ. gap — интервал), является начальным периодом интерфазы. В этот период ДНК еще не синтезируется, однако происходит накопление РНК и белков, в том числе и белков, необходимых для синтеза ДНК. Увеличивается количество митохондрий. Обычно этот период длится 12-24 часа.
Синтетический период (S) следует за G1
-периодом и характеризуется тем, что в этот период в клетке происходит синтез (репликация) ДНК, в результате чего количество ее удваивается. В этот период продолжается также синтез РНК и белков. Очень важно, что к концу этого периода каждая из хромосом удваивается и состоит уже из двух сестринских хроматид, удерживаемых центромерои. Можно сказать, что наиболее фундаментальной особенностью S-периода является репликация генов и удвоение набора генов каждой дуплицированной хромосомы. Длительность S-периода обычно составляет около 5 часов.
Постсинтетический период (G2
) характеризуется остановкой синтеза ДНК и накоплением энергии. Однако продолжается синтез РНК и белков, формирующий нити веретена деления. Длительность G2
-периода составляет 3—6 часов.
Митоз совершается на протяжении четырех последовательных фаз, а именно: профазы, метафазы, анафазы и телофазы (рис. 59.).
 В профазе в начале происходит конденсация и спирализация (скручивание) хромосом, в результате чего они становятся видимыми при микроскопии окрашенных препаратов. Увеличивается диаметр каждого завитка. Ядерная мембрана растворяется под действием ферментов, ядрышко исчезает. Центросома делится на две центриоли, после чего последние расходятся к полюсам клетки. Отмечается также фосфорилирование отдельных клеточных белков. Затем между полюсами начинает формироваться ахромати-новая фигура, похожая на веретено. Оно состоит из белка и РНК. К концу этой фазы ахроматиновая фигура вытягивается вдоль клетки, становясь веретеном. Структурно веретено представляет собой двухполюсную структуру, построенную из микротрубочек и различных белков. Хроматиды (сестринские хроматиды) удерживаются вместе центромерои. Длительность профазы составляет примерно 30-60 минут. В профазе в начале происходит конденсация и спирализация (скручивание) хромосом, в результате чего они становятся видимыми при микроскопии окрашенных препаратов. Увеличивается диаметр каждого завитка. Ядерная мембрана растворяется под действием ферментов, ядрышко исчезает. Центросома делится на две центриоли, после чего последние расходятся к полюсам клетки. Отмечается также фосфорилирование отдельных клеточных белков. Затем между полюсами начинает формироваться ахромати-новая фигура, похожая на веретено. Оно состоит из белка и РНК. К концу этой фазы ахроматиновая фигура вытягивается вдоль клетки, становясь веретеном. Структурно веретено представляет собой двухполюсную структуру, построенную из микротрубочек и различных белков. Хроматиды (сестринские хроматиды) удерживаются вместе центромерои. Длительность профазы составляет примерно 30-60 минут.
В метафазе хромосомы располагаются на экваторе веретена. Они имеют вид толстых образований, плотно свернутых спиралью, что облегчает подсчет и изучение их структуры с помощью микроскопа. Будучи прикрепленными к нитям веретена центромерои, к которой прикрепляются особые белковые комплексы (кине-тофоры), связанные с отдельными микротрубочками хроматиды, пока удерживаются вместе, но плечи их уже разъединены. Длительность метафазы составляет 2—10 минут.
В анафазе наступает разделение кинетохоров, а затем и продольное разделение хромосом, в результате чего каждая сестринская хро-матида имеет собственную центромеру и становится дочерней хромосомой. Хромосомы удлиняются и двигаются к соответствующим полюсам веретена. Анафаза длится 2-3 минуты. Репликация хромосомных концов (теломер), длина которых составляет 2—20 кб., требует теломеразы.
В телофазе (от греч. telos — конец) дочерние хромосомы достигают полюсов, вытягиваются и деспирализуются. Кинетохорные трубочки исчезают. Образуется ядерная оболочка, вновь появляется ядрышко. Длительность составляет 20-30 минут.
На заключительном этапе клеточного деления происходит ци-токинез, который начинается еще в анафазе. Этот процесс заканчивается образованием в экваториальной зоне клетки перетяжки, которая разделяет делящуюся клетку на две дочерние клетки. Перетяжка обеспечивается сокращением кольца, сформированного филаментами актиновой природы.
В отличие от соматических клеток животных в клетках растений из-за ригидности их стенок вместо образования сократительного кольца формируется пластинка между будущими дочерними клетками. На каждой из сторон этой пластинки откладывается целлюлоза, после чего она становится клеточной стенкой.
Каждое клеточное деление является непрерывным процессом, поскольку ядерные и цитоплазматические фазы, вопреки различиям в содержании и по значению, координированы во времени.
Упорядоченность клеточных делений у эукариотов зависит от координации событий в клеточном цикле. У эукариот эта координация осуществляется путем регуляции трех переходных периодов в клеточном цикле, а именно: вступление в митоз, выход из митоза и прохождение через пункт, называемый «Старт», который вводит инициацию синтеза ДНК ( S-фазу) в клетке.
Продолжительность митотических циклов разных клеток различна и составляет от нескольких часов до нескольких дней. Однако она зависит от типа тканей, физиологического состояния, внешних факторов (температура, свет).
Клеточный цикл эукариотических клеток регулируется последовательной активацией циклинзависимых киназ (СДК) путем взаимодействия их с белками-циклинами. Комплекс циклин-СДК оказывается полностью активированным фосфорилированием треонинового остатка в Т-петле СДК, осуществляемым специфической СДК-активирующей киназой (САК). При этом комплекс циклин-СДК вовлекается в инициацию как митоза, так и репликации ДНК. Регуляция митоза зависит от регуляции СДК.
Существуют и другие регуляторы клеточного цикла. В частности известны регуляторы, ингибирующие СДК. Такими ингибиторами являются белки р21, р16 и р27. Они ингибируют функции киназ также путем связывания с ними.
Хромосомная ДНК в клетках организмов-эукариотов реплици-руется лишь один раз в клеточном цикле. Поэтому давно возник вопрос о механизме, ограничивающем лишь один раунд репликации ДНК в клеточном цикле. Предполагают существование так называемого лицензирующего фактора репликации (licensing factor), который позволяет репликацию. В подтверждение этого взгляда установлены лицензирующие белки MSM, которые обычно связаны с хромосомами, но с началом S-фазы освобождаются от этой связи, позволяя репликацию ДНК, а после того, как синтез ДНК завершается, вновь связывается с хромосомами.
Разные ткани характеризуются разной митотической активностью. Поэтому в зависимости от митотической активности различают стабильные, растущие и обновляющиеся ткани. Стабильные ткани — это ткани, в которых клетки не делятся, а количество клеточной ДНК постоянно. Например, клетки центральной и периферической нервной системы не делятся. В этих клетках происходят лишь возрастные изменения. Растущие ткани — это ткани, в которых клетки живут всю жизнь, но среди последних имеются такие, которые делятся посредством митоза. В результате этого наступает увеличение размеров органов. Примером растущих тканей являются ткани почек, желез внутренней секреции, скелетная и сердечная мускулатуры. Обновляющиеся ткани — это ткани, в которых многие клетки подвержены митозам, в результате чего погибающие клетки компенсируются вновь образующимися. Примерами обновляющихся тканей являются клетки желудочно-ки-шечного, дыхательного и мочеполового трактов, эпидермиса, костного мозга, семенников и др. Для митозов характерны суточные колебания, волны.
У высших организмов митотическое деление клеток обеспечивает их рост с последующим увеличением массы тела и дифференциацией клеток. По мере индивидуального развития человека количество его клеток увеличивается, достигая у взрослого человека более чем 10 клеток и оставаясь затем константным.
Как уже отмечено, митохондрии и хлоропласты способны к делению в клетках эукариотов, но контроль их деления не ясен. Установлено лишь, что в геноме клеток растений существует ген, который, возможно, принимает участие в контроле деления хло-ропластов.
Для деления клеток млекопитающих и птиц характерно то, что оно имеет определенные ограничения количества клеточных удвоений. Например, фибробласты плодов человека удваиваются лишь на протяжении 50 генераций, тогда как фибробласты от людей в возрасте 40 и 80 лет подвергаются примерно 40 и 30 удвоениям соответственно, если их культивируют в стандартных условиях. Это явление получило название старения клеток. Считают, что в организме также большинство клеток стареет, например, клетки печени живут около 18 месяцев, эритроциты — 4 месяца, в результате чего в них накапливаются липиды, кальций, пигмент «изнашивания» и они гибнут. Подсчитано, что организм взрослого человека ежедневно теряет около 1-2% своих клеток в результате их гибели. После смерти клетки в ней происходит коагуляция протоплазмы, распад митохондрий и других органелл в результате ауто-лиза (активации внутриклеточных ферментов).
Для объяснения природы старения клеток предложено несколько гипотез, в которых придается значение ошибкам биосинтетических механизмов клеток, механизмам защиты от злокачественного перерождения нормальных клеток или другим причинам. Однако ни одна из известных гипотез не является исчерпывающей в объяснении феномена старения клеток.
Установлено, что для клеток во многих случаях характерен апоптоз, под которым понимают генетическую программу, в результате которой клетки совершают суицид. Можно сказать, что апоптоз — это эволюционно сохраняемый процесс. С помощью этого процесса многоклеточные организмы освобождаются от излишних или потенциально вредных клеток. Этот феномен отличен от старения клеток. На примере нематоды Caenorhabditie elegans было выяснено, что клеточный суицид контролируется генным набором, состоящим из трех генов, контролирующих синтез белка СЕД-3, СЕД-4 и СЕД-9, регулирующих апоптоз. У млекопитающих выявлены белки-2, которые регулируют апоптозную смерть клеток. Полагают, что апоптоз имеет значение в этиологии многих наследственных болезней (болезнь Альцгеймера и Др.), аутоиммунных нарушений, сердечно-сосудистых болезней, возрастных нарушений и даже СПИДа.
Однако погибающие клетки замещаются новыми. Считают, что клеточное содержание организма человека обновляется примерно каждые семь лет. Особенно сильно замещение клеток происходит в крови за счет интенсивного образования кровяных клеток в кроветворных тканях. Применительно к другим видам клеток процесс обновления происходит с очень высокой скоростью. Например, эпителий желудка и кишечника крыс обновляется каждые 72 и 38 часов соответственно, эпителий тонкого кишечника человека — каждые 7-8 дней. Однако нервные клетки функционируют (живут) на протяжении всей жизни организмов.
Наряду с делением клеток путем митоза известен амитоз (от греч. а — не, mytosia — деление ядра), под которым понимают прямое деление ядра клетки. При амитозе сохраняется интерфазное состояние ядра, ядрышко, ядерная мембрана. Ядро клетки делится на две части без формирования веретена, в результате чего образуется двухъядерная клетка. Амитоз встречается иногда в клетках скелетной мускулатуры, кожного эпителия, соединительной ткани. Однако считают, что амитоз является аномальным механизмом в размножении клеток.
Считают, что митотический цикл у высших организмов является результатом эволюции разделительного механизма эукарио-тов. В пользу этого предположения свидетельствуют результаты сравнения разделительных механизмов бактерий, некоторых водорослей, дрожжей, простейших и млекопитающих. Это сравнение показывает, что усложнение митотического аппарата происходит по мере усложнения организации и функций организмов, принадлежащих к разным систематическим группам.
Для клеток многоклеточных организмов характерна специализация и объединение, в результате которых они образуют структуры, получившие название тканей, из которых формируются органы. Впервые термин «ткань» был использован англичанином Н. Грю еще в 1671 г. С тех пор эти системы стали предметом изучения ученых — гистологов многих поколений. В наше время под тканью понимают систему объединенных клеток и их производных, выполняющих сходные специализированные функции. К этому следует добавить, что ткани являются результатом развития живых форм в ходе филогенеза и онтогенеза.
Клетки объединяются в составе тканей с помощью разных механизмов — «прикрепительных» и «коммуникационных». «Прикрепительный» механизм заключается в том, что клетки с помощью рецепторов адгезии (адгезинов) могут присоединяться к так называемому внеклеточному матриксу, представляющему собой сеть органических молекул (фибриллярных белков) и лигандов, погруженных в полисахаридный гель. Основным белком во внеклеточном матриксе является коллаген, полимерные формы которого сосредоточены в коже, сухожилиях, хрящах, кровеносных сосудах, внутренних органах и т. д. Важнейшей особенностью молекул коллагена является то, что им присуща трехцепочечная спиральная структура. Они могут связываться между собой межклеточными соединениями в виде адгезионного соединения или разных клеточных контактов (десмосом) или контактов между межклеточным матриксом и клетками (полудесмосом).
Помимо «прикрепительных» соединений для клеток в тканях характерны «коммуникационные» соединения, наиболее распространенные из которых получили название щелевых контактов. Различают несколько видов таких контактов. Они могут быть представлены щелями между плазматическими мембранами соседних клеток, заполненными рыхлой сетью органических молекул (внеклеточным матриксом), что обеспечивает щелевой контакт клеток. Далее, щелевые контакты могут иметь вид выпячиваний (выроста) плазматической мембраны одной клетки в плазматическую мембрану другой клетки и слипанием этих выпячиваний. Щелевые контакты позволяют малым молекулам переходить из одних клеток в другие. В случае нервных клеток имеют место синапсы, позволяющие передачу электрических и химических сигналов от одной клетки к другой. Важно подчеркнуть, что любой из названных межклеточных контактов основан на межмембранных связях.
Механизм объединения клеток растений является другим. Поскольку у них нет плазматической мембраны, но есть клеточная стенка, которая содержит каналы, то соединение соседних клеток обеспечивается соединением их цитоплазматическими мостиками (плазмодесмами), представляющими собой цитоплазму, проникающую через каналы.
Организация тканей связана с наличием у клеток обмена информацией, который достигается выделением клетками химических веществ, выполняющих функцию сигналов для других клеток, наличием на поверхностной мембране клеток сигнальных молекул, влияющих на другие клетки при их контакте, и щелевых контактов, позволяющих обмен малыми молекулами.
Химическая сигнализация осуществляется с помощью сигнальных молекул, в частности, гормонов, выделяемых эндокринными клетками и воздействующих через кровь на клетки-мишени, а также с помощью локальных химических медиаторов, действующих только на ближайшие (соседние) клетки. В случае нервной системы клетки секретируют нейромедиаторы. Примерами белковых гормонов являются инсулин, соматотропин, адренокортикотропный гормон, тогда как стероидными гормонами являются эстрадиол, тестостерон, кортизол и другие. Сигнальными молекулами являются также некоторые олигопептиды (соматостатин, вазопрессин и др.), адреналин и нейромедиаторы (глицин, ацетилхолин и др.). Примером локальных сигнальных молекул является гистамин, выделяемый клетками соединительной ткани (тучными клетками). Сигнальные молекулы еще называют лигандами. Они связываются со специфическими белковыми рецепторами на поверхности клеток-мишеней, в результате чего акт связывания генерирует сигнал, влияющий на поведение клеток, в частности на их кооперацию, ведущую к образованию тканей. Сигнальными молекулами, синтезируемыми на мембранной поверхности клеток, являются простагландины. Они очень быстро синтезируются и очень быстро разрушаются.
Образование тканей (гистогенез) у животных происходит из эктодермы, энтодермы, мезодермы и мезенхимы в период эмбриогенеза, а основными элементами тканей, как отмечено выше, являются клетки и их производные в виде неклеточных структур. Таким образом, ткань можно определить в виде сообщества клеток и их производных со специализированными функциями.
В рамках классификации тканей, основанной на морфофунк-циональном принципе, у животных и человека различают 5 типов тканей, а именно: эпителиальную, соединительную, мышечную и нервную ткани, а также кровь и лимфу.
Эпителиальная тканъ
, или эпителий, состоит из клеток, покрывающих поверхность тела, внутренние поверхности внутренних органов (желудок, мочевой пузырь и др.), поверхности серозных оболочек (брюшина, плевра, перикард), а также из клеток, образующих некоторые железы (слюнные железы, поджелудочная железа и др.). Поэтому различают покровный и железистый (секреторный) эпителий. Из эктодермы развивается эпителий кожи, из энтодермы — эпителий желудка, кишечника, легких и др., а из мезодермы — эпителий почек, серозных оболочек и других структур.
 Среди покровных эпителиальных тканей различают плоский, кубический, призматический и ресничный эпителий (рис. 60). Среди покровных эпителиальных тканей различают плоский, кубический, призматический и ресничный эпителий (рис. 60).
Плоский эпителий представлен уплощенными клетками, которые образуют поверхностный слой кожи и выстилают ротовую полость, пищевод и влагалище. Как правило, плоский эпителий является многослойным, образует слизистые оболочки пищевода, влагалища, эпидермис кожи и др.
Кубический эпителий представлен кубовидными клетками, которые выстилают почечные канальцы, наружную поверхность яичника и другие органы.
Призматический эпителий представлен клетками цилиндрической формы, им выстлан желудок, кишечник, матка и другие органы.
Ресничный эпителий представлен клетками, на поверхности которых имеются реснички. Биение этих ресничек обусловливает перемещение слизи и других веществ по эпителиальному слою.
Железистый эпителий представлен клетками призматической или кубической формы, которые продуцируют секрет. Они функционируют либо как одноклеточные железы, секретируя разные секреты, либо формируют многоклеточные железы, получившие название эндокринных желез, т. к. они выделяют продукты своей деятельности (гормоны) в кровь и лимфу.
Соединительные ткани
представлены собственно соединительной, костной и хрящевой тканями, развивающимися из мезенхимы. Они состоят из клеток и межклеточного вещества. Исходя из структуры и свойств межклеточного вещества, различают несколько типов этой ткани.
Волокнистая соединительная ткань представляет собой волокна (коллаген) и межклеточное вещество (протеогликаны и гликопротеиды), окружающие соединительнотканные клетки (фибробласты, макрофаги, тучные клетки) и являющиеся продуктом этих клеток. Эта ткань образует строму многих внутренних органов, основу слизистых оболочек, соединяет кожу с мышцами, участвует в формировании надкостницы.
 Костная ткань формирует скелет организма. Она состоит из костных клеток (остеоцитов, остеобластов и остеокластов) и выделяемого ими основного вещества кости, содержащего белки, из которых преобладающим является коллаген, и соли кальция (рис. 61). Костная ткань формирует скелет организма. Она состоит из костных клеток (остеоцитов, остеобластов и остеокластов) и выделяемого ими основного вещества кости, содержащего белки, из которых преобладающим является коллаген, и соли кальция (рис. 61).
Хрящи также формируют скелет (в эмбриональном состоянии). У взрослых хрящевой скелет имеется лишь у акул и скатов. Хрящевая ткань состоит из клеток (хондриоцитов, прехондроблас-тов и хондробластов) и межклеточного вещества (в основном коллагена).
Соединительные ткани выполняют опорную, трофическую, защитную и другие функции.
Кровь и лимфа
являются тканями, которые начинают развиваться уже в эмбриональном периоде жизни организмов из мезенхимы, а затем из так называемых полипотентных стволовых клеток крови (СКК). У человека развитие первых клеток крови идет синхронно с сосудами, развивающимися вначале  в стенке желчного мешка, а затем в печени, красном костном мозге, тимусе, селезенке, лимфатических узлах эмбриона. Образование крови и лимфы происходит и на протяжении всего постэмбрионального периода. Важнейшими функциями крови являются трофическая, дыхательная и транспортная. в стенке желчного мешка, а затем в печени, красном костном мозге, тимусе, селезенке, лимфатических узлах эмбриона. Образование крови и лимфы происходит и на протяжении всего постэмбрионального периода. Важнейшими функциями крови являются трофическая, дыхательная и транспортная.
Кровь является очень сложным образованием, составляющим у человека примерно 5-9% массы тела. В ее составе различают плазму и форменные элементы — эритроциты, лейкоциты и тромбоциты (кровяные пластинки).
Плазма крови состоит на 90—93% из воды, в которой содержатся белки, углеводы, жиры и минеральные вещества.
Эритроциты, или красные кровяные тельца (шарики), представляют собой безъядерные овальные клетки, диаметр которых составляет 7,1-7,9 мкм (рис. 62). 1 мл крови мужчины содержит 3,9— 5,5 х 109
эритроцитов, а 1 мл крови женщины — 3,7—4,9 х 109
. Основной функцией эритроцитов является транспортировка кислорода и углекислоты.
Лейкоциты (белые кровяные клетки) подразделяют на гранулоциты и агранулоциты. В составе гранулоцитов на основе отношения их к красителям различают нейтрофилы, эозинофилы и базофилы. В составе агранулоцитов различают лимфоциты и моноциты. Лимфоцитов в крови довольно много (20—35%). Они очень полиморфны. Их размеры составляют 4,5—10 мкм. Поскольку для них характерно разное происхождение, то различают Т-лимфоци-ты, образование которых происходит в тимусе, и В-лимфоциты, образующиеся в красном костном мозге. Эти лимфоциты различаются и по функциям (см. § 96).
Моноциты являются клетками размером 18-22 мкм. Их доля среди лейкоцитов составляет 6—7%. Эти клетки постоянно мигрируют в соединительную ткань, где они дают начало макрофагам.
Лейкоциты выполняют защитную функцию (участвуют в формировании иммунитета).
Тромбоциты (красные кровяные пластинки) — это безъядерные тельца размером 2—3 мкм, содержание которых в 1 мл крови человека равно 3 х Ю8
. Являясь составной частью тромбоксилазы, они принимают участие в свертывании крови.
Лимфа, подобно крови, также состоит из жидкой части и форменных элементов. Жидкой частью является лимфоплазма, а форменные элементы представлены в основном лимфоцитами. В лимфе встречаются также моноциты, но в небольшом количестве. Основная функция лимфы заключается в регулировании циркуляции лимфоцитов, а также оттока различных жидкостей и находящихся в ней метаболитов от органов.
Мышечная ткань
образована мышечными клетками (миоцита-ми), являющимися структурно-функциональными единицами многоядерных мышечных волокон — миофибрилл. Эти волокна образуются в результате слияния миоцитов. Установлено, что слияние обеспечивается несколькими белками (кадгеринами, интегринами, меятринами). Различают гладкую и поперечно-полосатую мышечную ткань (рис. 63), которые различаются между собой по строению миофибрилл. Гладкие мышцы построены из вытянутых сигароподобных клеток (миоцитов). Они формируют мышечные слои стенок сосудов, бронхов, желудка, кишечника и т. д. Поперечно-полосатая мышечная ткань представлена скелетной мышечной тканью. Скелетные мышцы прикрепляются к костям. Сердечная мышечная ткань представлена сократительными кардиомиоцитами. Сократительная способность мышц обеспечивается по той причине, что сократительные структуры (миофибриллы) содержат миозин и актин.

Нервная ткань формируется из эктодермы и представлена нейронами (нейроцитами), которые являются клетками, проводящими электрические импульсы, и клетками нейроглии (рис. 64).
Нейрон состоит из тела, в котором содержится ядро, и отходящих от тела двух или более отростков. Те отростки, которые проводят нервные импульсы от тела нейрона к периферии, получили название аксонов, а те, которые проводят импульсы к телу нейрона, названы дендритами. Нейроглия представлена клетками, выстилающими полости головного и спинного мозга и образующими оболочки нейронов и их отростков, а также клетками, встречающимися на поверхности тела нейронов и нервных ганглиев, в нервных окончаниях. Нервными волокнами являются отростки нервных клеток и глиальные оболочки.
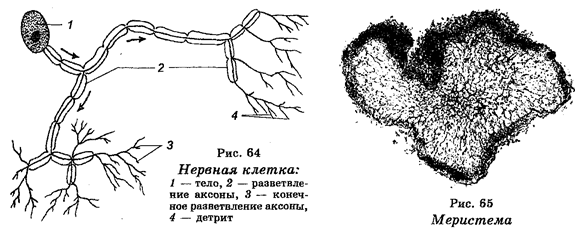
Нервная ткань
составляет основной компонент нервной системы, главные функции которой заключаются в регуляции функционирования тканей и органов, а также координации связи организмов с окружающей средой.
Клетки почти всех высших растений также специализированы и организованы в ткани. У растений различают меристематичес-кую (образовательную), покровную (защитную), основную и проводящую ткани.
 Меристематические ткани представлены мелкими клетками с крупными ядрами, в которых очень высок уровень метаболизма (рис. 65). Эти клетки способны к делению, что обеспечивает рост растений в течение длительного периода. Кроме того, они дают начало тканям остальных типов, т. к. происходит их дифференциация в ткани других типов. Меристема имеется в зародыше, на кончиках корней, а также в тех частях растения, которые очень быстро растут, и в камбии. Меристемы осевых органов растений обеспечивают их рост в длину, тогда как меристемы стебля и корня ответственны за их рост в толщину. В частности деление клеток камбия сопровождается ростом стебля в толщину. Слои клеток древесины, выросшие в течение сезона (весна, лето и осень), образуют так называемое годичное кольцо прироста. Меристематические ткани представлены мелкими клетками с крупными ядрами, в которых очень высок уровень метаболизма (рис. 65). Эти клетки способны к делению, что обеспечивает рост растений в течение длительного периода. Кроме того, они дают начало тканям остальных типов, т. к. происходит их дифференциация в ткани других типов. Меристема имеется в зародыше, на кончиках корней, а также в тех частях растения, которые очень быстро растут, и в камбии. Меристемы осевых органов растений обеспечивают их рост в длину, тогда как меристемы стебля и корня ответственны за их рост в толщину. В частности деление клеток камбия сопровождается ростом стебля в толщину. Слои клеток древесины, выросшие в течение сезона (весна, лето и осень), образуют так называемое годичное кольцо прироста.
Покровные ткани представлены плотно сомкнутыми клетками, располагающимися на внешней поверхности растений (рис. 66). К этим тканям относят эпидерму листьев, а также пробковые слои стебля и корней. Они выполняют защитную функцию, предохраняя от высыхания или механических повреждений лежащие глубже тонкостенные клетки.

Основные ткани представлены различными по форме клетками, образующими основную массу тела растений (мягкие части листьев, цветков, плодов, сердцевину стеблей и корней, а также кору). Главная функция этих тканей заключается в синтезе и накоплении питательных веществ. В частности, часть этих тканей представлена хлоропластосодержащими клетками, в которых происходит фотосинтез.
Проводящие ткани (рис. 67) представлены ксилемой (древесина) и флоэмой (луб). Клетки ксилемы дают начало длинным клеткам, называемым трахеидами. Соединяясь между собой концами, трахеиды образуют сосуды древесины. После растворения в них поперечных стенок они превращаются в длинные целлюлозные трубки, по которым и проходит вода. Ксилема проводит воду и растворенные в ней соли от корня к листьям, что представляет собой восходящий (транспирационный) ток.
Флоэма образуется так же, как и ксилема, но с той лишь разницей, что поперечные стенки не устраняются, а сохраняются. Однако в них образуются отверстия, обеспечивающие «проход» органических веществ от листьев к корням. Следовательно, флоэма обеспечивает нисходящий ток, т. е. движение органических веществ от листьев к корням.
Происхождение, специализация и объединение клеток в ткани есть категории исторические, ибо они возникли в ходе филогенеза. Однако объяснение этих категорий чрезвычайно затруднено, поскольку между клетками прокариотических и эукариотических организмов существуют важные различия. Тем не менее известно несколько гипотез.
На основании изучения ископаемых остатков бактерий и циано-бактерий предполагают, что предковой клеточной формой была примитивная прокариотическая клетка, возникшая около 3,5 х 109
лет назад. Клетки этого типа для обеспечения своего существования и размножения в начале использовали органические молекулы небиологического происхождения. Первым актом в формировании примитивных клеток было образование мембраны, окружавшей вещество клетки.
В последующем у примитивных прокариотических клеток стали развиваться механизмы синтеза и энергетического обеспечения. Предполагают, что первые прокариотические клетки обладали наиболее простыми каталитическими системами, в результате чего обеспечение их энергией основывалось на брожении. В последующем отдельные виды прокариотических клеток переключились с брожения на дыхание, что способствовало более эффективному получению энергии. Таким образом, эволюционные изменения прокариотических клеток шли по линии развития у них различных метаболических путей. Их геном развивался в направлении формирования «голых» молекул ДНК.
Эволюционные изменения эукариотических клеток шли в направлении увеличивающегося разнообразия в форме, размерах, структуре и функциях с одновременной компартментализацией биохимических систем и сохранением общего для всех Клеток аэробного метаболизма. Считают, что эукариотические клетки возникли менее 1 млрд лет назад из прокариотических клеток, причем для объяснения их происхождения выдвинуто три гипотезы.
В соответствии с одной их этих гипотез (гипотезой клеточного симбиоза), которая является наиболее распространенной, предполагают, что эукариотическая клетка является симбиотической структурой, состоящей из нескольких клеток разных типов, объединенных общей клеточной мембраной. В частности, предполагают, что пластиды клеток современных зеленых растений происходят от бактерий, бывших предками современных цианобактерий и способных к аэробному фотосинтезу, а митохондрии эукариотических клеток ведут начало от аэробных бактерий, которые вступали в симбиоз с примитивными анаэробными клетками, способными к фотосинтезу, что привело к образованию клеток, способных к существованию в атмосфере кислорода и использованию кислорода путем дыхания. Относительно ядра предполагают, что оно является рудиментом также какого-то древнего внутриклеточного симбионта, утратившего после включения в исходную клетку свою цитоплазму. В пользу этой гипотезы свидетельствуют данные о симбиотических взаимоотношениях некоторых современных организмов. Например, одноклеточная зеленая водоросль хлорелла (Chlorella) обитает в цитоплазме зеленой парамеции (Paramecium bussaria). Из-за способности к фотосинтезу она снабжает парамеции питательными веществами. Пластиды и митохондрии содержат собственную систему генетической информации о синтезе белков в виде ДНК, мРНК, рРНК, тРНК и соответствующих ферментов. Для хлоропластов, митохондрии и клеток-прокариот характерно сходство способов репродукции (все они одинаково репродуцируются путем простого деления надвое). Наконец, мутации митохон-дриальных генов назависимы от мутаций ядерных генов.
В соответствии с другой гипотезой считают, что эукариотичес-кая клетка произошла от прокариотической клетки, содержавшей несколько геномов, прикрепленных к клеточной мембране. В результате инвагинаций клеточной мембраны образовывались мезо-сомы, способные первоночально к фотосинтезу. Однако в дальнейшем произошла специализация этих органелл, в результате чего одна из них, утратив дыхательную и фотосинтетическую функцию, развилась в ядро, другие, наоборот, развив эти функции, стали митохондриями у животных и пластидами у растений. В пользу этой гипотезы свидетельствуют данные о двойном строении мембран ядра, митохондрии и пластид.
В соответствии с третьей гипотезой, основанной на мысли о том, что все живые формы произошли от предковых анаэробных ферментативных гетеротрофов, эукариоты представляют собой сублинию бесстеночных (анаэробных) прокариотов, которые развили способность к эндоцитозу. Посредством «заглатывания» других прокариотов, которые дали им дополнительные метаболические способности и которые, в конце концов, дегенерировали в органеллы, примитивная клетка (уркариот) стала эукариотической клеткой. Таким образом, прокариоты древнее, проще и примитивнее клеток-эукариот.
В соответствии с четвертой гипотезой предполагают, что эука-риотические клетки возникли из прокариотической клетки, содержавшей много геномов, которые распадались на части, давшие начало структурам с разными функциями. В последующем шло кло-нирование структур со сходными функциями, после чего они покрывались двойными мембранами, что привело к образованию ядра, митохондрии, а позднее и мембранной сети. В пользу этой гипотезы свидетельствуют данные о сходстве генетического кода, содержащегося в ядерной и митохондриальной ДНК, а также о сходстве в регуляции дыхательной функции ядром и митохондриями.
Как отмечено выше, симбиотическая гипотеза происхождения эукариотических клеток сейчас наиболее популярна. Однако, разделяя эту гипотезу, нельзя не отметить, что митохондрии и хло-ропласты вопреки их сходству с современными бактериями-аэробами и цианобактериями (соответственно) все же существенно отличаются от них. В частности, в митохондриях и хлоропластах намного меньше ДНК. Следовательно, здесь митохондрии и хлоропласты в ходе эволюции претерпели значительные изменения в направлении своих размеров.
Геном эукариотических клеток впоследствии развивался в направлении объединения молекул ДНК с белками и формирования хроматина и хромосом разной формы и в разном количестве. Специализация хроматина проявилась в формировании эухроматина и гетерохроматина, в формировании аутосом и половых хромосом. Что касается количества хромосом, то объяснить их эволюционную тенденцию пока трудно, поскольку многие примитивные организмы содержат в своих клетках большее число хромосом, чем организмы, занимающие высшие эволюционные ступени. Однако несомненно, что количественные и структурные изменения карио-типов в течение эволюции играли важную роль в видообразовании. Параллельно с этим происходило усложнение структуры и функции клеточных компонентов, развитие регуляторных механизмов.
Несомненно эволюционное значение митоза. Считают, что точность разделения и распределения хромосом в результате митоза является условием, обеспечивающим многоклеточность. Однако происхождение самого митоза не имеет достаточных объяснений. Предполагают лишь, что он развился из примитивного митоза, представляющего собой механизм, при котором расхождение реп-лицировавшихся хромосом происходило после вытягивания и разрыва веретена без разрушения ядерной мембраны (см. выше).
Объяснения эволюции тканей связаны со сложностями, которые обусловлены одинаковым строением тканей, принадлежащих живым организмам, находящимся на разных ступенях эволюционной лестницы. Например, мышечные волокна членистоногих, некоторых моллюсков и позвоночных имеют одинаковое строение. Между тем эти организмы филогенетически разделены очень большими «расстояниями». Аналогичная ситуация имеет место и при сравнении тканей растений из разных таксономических групп.
Начала тканеобразования в эволюционном плане уже прослеживаются у самых простых организмов. Например, у вольвокса отмечается формирование колоний, состоящих иногда более чем из 50 000 клеток, причем часть клеток уже специализирована. В частности, клетки, располагающиеся по краям колониальной формы, ответственны за образование новых колоний. У цианобактерий при нерасхождении разделившихся клеток образуются клеточные нити, в которых часть клеток специализирована на фиксации азота, чем обеспечиваются потребности в азоте и других клеток.
Идя вверх по эволюционной лестнице, можно видеть, что у губок уже отмечается около пяти специализированных типов клеток, специализация которых связана с выполнением разных функций в процессе фильтрации воды и поглощения отфильтрованных пищевых частиц.
У кишечнополостных тело состоит из двух слоев — эктодермы и энтодермы, представляющих собой наружный и внутренний эпителиальные слои. Наружные эпителиальные клетки являются стрекательными клетками, содержащими ядовитую жидкость, тогда как внутренние эпителиальные клетки секретируют пищеварительные ферменты и обеспечивают пищеварение. Поэтому предполагают, что первыми сформировались слои эпителиальных клеток и их роль в эволюции многоклеточных аналогизируется с ролью клеточных стенок и мембран одноклеточных организмов.
Значительный вклад в понимание эволюции тканей принадлежит А. А. Заварзину (1886-1945), который считал, что одни и те же факторы эволюции обеспечили не только разнообразие организмов, но и однообразие строения их тканей. Сходство в строении тканей у филогенетически далеко отстоящих животных А. А. За-варзин называл законом параллельных рядов тканевой эволюции. Работы А. А. Заварзина и его учеников заложили основы эволюционной гистологии.
1. Насколько велико значение методов исследования в изучении клеток? Какие из этих методов вы знаете?
2. Сформулируйте основные положения клеточной теории. Как Вы считаете, какова роль этой теории в биологии?
3. Почему клетку определяют в качестве элементарной единицы жизни и в чем заключаются доказательства того, что клетка действительно является элементарной единицей жизни? Что представляют собой межклеточные структуры?
4. Назовите два процесса, которые являются общими для всех живых систем.
б. Назовите принципиальные различия между клетками-прокариота-ми и клетками-эукариотами. Является ли одноклеточность признаком прокариот?
6. Назовите и охарактеризуйте компоненты мембранной системы клеток животных. Есть ли мембранная система в клетках растений?
7. Охарактеризуйте цитоплазматический матрикс и клеточные орга-неллы. Что собой представляет цитозоль? Есть ли у клеток скелет? Как организован цитоскелет и каковы его компоненты?
8. Каковы структура и роль клеточного ядра? Есть ли различия между ядрами клеток животных и клеток растений?
9. Каковы структура и функции митохондрий? Все ли клетки обладают митохондриями?
10. Сформулируйте определения клеточного цикла и митоза. С какой скоростью протекают митозы в клетках разных тканей?
11. Что собой представляют лизосомы и какова их роль? Что произойдет с клетками, если лизосомы подвергнутся разрушению?
12. Каково значение ферментов в жизни клеток? Все ли белки являются ферментами и в чем заключается их действие?
13. Каковы фазы митоза и сущность процессов, происходящих в эти фазы?
14. В какой фазе происходит разделение центромеры и освобождение сестринских хроматид?
15. Определите, какая весовая часть ядра приходится на хроматин клетки (примерно), допуская, что диаметр ядра составляет б мкм, а плотность 1,1 г/см"3
?
16. Считая, что хромосомы человека состоят на 15% из ДНК, определите массу всех хромосом его диплоидных клеток.
17. Что вы можете сказать о происхождении митоза?
18. Что вы знаете об элементарном составе клеток?
19. Что понимают под биологическими молекулами?
20. Какой представляется вструктура белков и что вы знаете о функциях белков?
21. Как вы понимаете происхождение клеток-прокариот?
22. Как вы понимаете происхождение клеток-эукариот?
23. Каково ваше мнение относительно развития генома эукариотичес-ких клеток?
24. Каковы причины гибели клеток? Существует ли генетический механизм, контролирующий гибель клеток?
25. Что вы знаете об эпителиальных тканях и их функциях?
26. Назовите основные группы мышечной ткани и что составляет основу их классификации?
27. Каковы основные клеточные элементы собственно соединительных тканей?
28. Что такое нервная ткань и из каких компонентов она состоит?
29. Как вы представляете строение нервного волокна?
30. Почему кровь и лимфу считают тканями?
31. Какова функциональная роль лимфоцитов?
32. Как вы понимаете происхождение клеток-прокариотов и клеток-эукариотов?
33. Применима ли эволюционная теория к учению о тканях?
Альберте В., Брей Д., Льюис Дж„ Рэфф М., Роберте К., Уотсон Дж. Молекулярная биология клетки. М.: Мир. 1994. Т. 1. 615 стр.; 1994. Т. 2. 540 стр.
Вермель Е. М. История учения о клетке. М
.:
Наука
.
1970. 259
стр
.
Kaufman
Р
.
В
., Wu W. Handbook of Molecular and Cellular Methods in Biology and Medicine. CRC Press L.
1995. 496
pp. Lackie J. M„ Dow
3.
A. The Dictionary of Cell Biology. Academic
Press
. 1995. 380
pp
.
Обмен веществ и энергии (метаболизм) — это совокупность химических реакций, протекающих в клетках или в целостном организме и заключающихся в синтезе сложных молекул и новой протоплазмы (анаболизм) и в распаде молекул с освобождением энергии (катаболизм). Энергия необходима для биосинтеза (образования нового вещества), осмотической работы (поглощения и секреции клетками разных веществ), механической работы (при движении) и других реакций.
Обмен веществ и энергии — это важнейшее свойство живого, проявляющееся на разных уровнях организации живого. Благодаря обмену веществ и энергии происходят рост и размножение, формируются другие важнейшие свойства клеток и организмов. Характерная особенность метаболических функций животных и растительных клеток заключается в том, что они являются ферментативными и сходны между собой, поскольку клетки всех орга низмов обладают всеми молекулами, играющими центральную роль в метаболизме и обеспечивающими переход энергии одного вида в энергию другого вида. Кроме того в основе регуляции метаболических путей лежат общие механизмы. Благодаря этому энергетические процессы у всех живых существ сходны. Жизнь существует и продолжается лишь благодаря энергии (рис. 68).
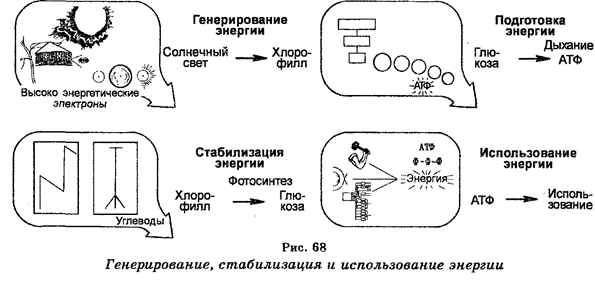
Основными метаболическими процессами являются анаболизм (ассимиляция) и катаболизм (диссимиляция).
Анаболизм, или ассимиляция (от лат. assimilatio — уподобление), представляет собой эндотермический процесс уподобления поступающих в клетку веществ веществам самой клетки. Она является «созидательным» метаболизмом.
Важнейшим моментом ассимиляции является синтез белков и нуклеиновых кислот. Частным случаем анаболизма является фотосинтез, который представляет собой биологический процесс, при котором органическое вещество синтезируется из воды, двуокиси углерода и неорганических солей под влиянием лучистой энергии Солнца. Фотосинтез в зеленых растениях является автотрофным типом обмена.
Катаболизм, или диссимиляция (от лат. dissimilis — расподобление), является экзотермическим процессом, при котором происходит распад веществ с освобождением энергии. Этот распад происходит в результате переваривания и дыхания. Переваривание представляет собой процесс распада крупных молекул на более мелкие молекулы, тогда как дыхание является процессом окислительного катаболизма простых Сахаров, глицерина, жирных кислот и дезаминированных аминокислот, в результате которого происходит освобождение жизненно необходимой химической энергии. Эта энергия используется для пополнения запасов аденозинтри-фосфата (АТФ), который является непосредственным донором (источником) клеточной энергии, универсальной энергетической «валютой» в биологических системах. Пополнение запасов АТФ обеспечивается реакцией фосфата (Ф) с аденозиндифосфатом (АДФ), а именно:
АДФ + Ф + энергия
® АТФ
Когда АТФ разлагается на АДФ и фосфат, энергия клетки освобождается и используется для работы в клетке. АТФ представляет собой нуклеотид, состоящий из остатков аденина, рибозы и трифосфата (трифосфатных групп), тогда как аденозиндифосфат (АДФ) имеет лишь две фосфатные группы. Богатство АТФ энергией определяется тем, что его трифосфатный компонент содержит две фос-фоангидридные связи. Энергия АТФ превышает энергию АДФ на 7000 ккал/моль. Этой энергией обеспечиваются все биосинтетические реакции в клетке в результате гидролиза АТФ до АДФ и неорганического фосфата. Итак, цикл АТФ-АДФ является основным механизмом обмена энергии в живых системах.
К живым системам применимы два закона термодинамики.
В соответствии с первым законом термодинамики (законом сохранения энергии) энергия на протяжении химических и физических процессов не создается, не исчезает, а просто переходит из одной формы в другую, пригодную в той или иной мере для выполнения работы, т. е. использование энергии для выполнения какой-либо работы или переход энергии из одной формы в другую не сопровождается изменением (уменьшением или увеличением) общего количества энергии. Имея в виду глобальные категории, можно сказать, что вопреки любым физическим или химическим изменениям во Вселенной, количество энергии в ней останется неизменным.
В соответствии со вторым законом термодинамики физические и химические процессы протекают в направлении необратимого перехода полезной энергии в хаотическую, неупорядоченную форму и установления равновесия между упорядоченным состоянием и хаотическим, неупорядоченным. По мере приближения к установлению равновесия между упорядоченностью и неупорядоченностью и к остановке процесса происходит уменьшение свободной энергии, т.е. той порции общей (полезной) энергии, которая способна производить работу при постоянной температуре и постоянном давлении. Когда количество свободной энергии уменьшается, то повышается та часть общей внутренней энергии системы, которая является мерой степени случайности и неупорядоченности (дезорганизации) и называется энтропией. Другими словами, энтропия есть мера необратимого перехода полезной энергии в неупорядоченную форму. Таким образом, естественная тенденция любой системы направлена на повышение энтропии и уменьшение свободной энергии, которая является самой полезной термодинамической функцией. Живые организмы являются высокоупорядоченными системами. Для них характерно содержание очень большого количества информации, но они бедны энтропией.
Если Вселенная представляет собой реакционные системы, под которыми понимают совокупность веществ, благодаря которым протекают физические и химические процессы, с одной стороны, и окружающую среду, с которой реакционные системы обмениваются информацией, с другой стороны, то в соответствии со вторым законом термодинамики в ходе физических процессов или химических реакций энтропия Вселенной увеличивается. Метаболизм живых организмов не сопровождается возрастанием внутренней неупорядоченности, т. е. для живых организмов не характерны возрастные энтропии. В любых условиях все организмы, начиная от бактерий и заканчивая млекопитающими, сохраняют упорядоченный характер своего строения. Однако для самой энтропии характерно то, что она возрастает в окружающей среде, причем непрерывное возрастание энтропии в окружающей среде обеспечивается существующими в среде живыми организмами. Например, для извлечения свободной энергии анаэробные организмы используют глюкозу, которую они получают из окружающей среды и окисляют молекулярным кислородом, проникающим тоже из среды. При этом конечные продукты окислительного метаболизма (СО2
и H2
O) поступают в среду, что и сопровождается возрастанием энтропии среды, которое частично происходит из-за рассеивания тепла. Возрастание энтропии в этом случае повышается, кроме того за счет возрастания количества молекул после окисления (C6
H12
O6
+ 6O2
® 6СО2
+ 6Н2
О), т. е. образование из 7 молекул 12 молекул. Как видно, молекулярная неупорядоченность ведет к энтропии.
Для живых существ первичным источником энергии является солнечная радиация, в частности видимый свет, который состоит из электромагнитных волн, встречающихся в виде дискретных единиц, называемых фотонами или квантами света. В живом мире одни живые существа способны улавливать световую энергию, другие получают энергию в результате окисления пищевых веществ.
Энергия видимого света улавливается зелеными растениями в процессе фотосинтеза, который осуществляется в хлоропластах их клеток. Благодаря фотосинтезу живые существа создают упорядоченность из неупорядоченности, а световая энергия превращается в химическую энергию, запасаемую в углеводах, являющихся продуктами фотосинтеза. Таким образом, фотосинтезирующие организмы извлекают свободную энергию из солнечного света. В результате этого клетки зеленых растении обладают высоким содержанием свободной энергии.
Получение энергии в результате окисления неорганических веществ происходит при хемосинтезе.
Животные организмы получают энергию, уже запасенную в углеводах, через пищу. Следовательно, они способствуют увеличению энтропии среды. В митохондриях клеток этих организмов энергия, запасенная в углеводах, переводится в форму свободной энергии, подходящей для синтеза молекул других веществ, а также для обеспечения механической, электрической и осмотической работы клеток. Освобождение энергии, запасенной в углеводах, осуществляется в результате дыхания — аэробного и анаэробного. При аэробном дыхании расщепление молекул, содержащих запасенную энергию, происходит путем гликолиза и в цикле Кребса. При анаэробном дыхании действует только гликолиз. Таким образом, жизнедеятельность клеток животных организмов обеспечивается в основном энергией, источником которой служат реакции окисления-восстановления «топлива» (глюкозы и жирных кислот), в процессе которых происходит перенос электронов от одного соединения (окисление) к другому (восстановление). С окислительно-восстановительными реакциями сопряжено фосфорилирование. Эти реакции протекают как при фотосинтезе, так и дыхании.
Организм — открытая саморегулирующая система, она поддерживает и реплицирует себя посредством использования энергии, заключенной в пище, либо генерируемой Солнцем. Непрерывно поглощая энергию и вещества, жизнь не «стремится» к равновесию между упорядоченностью и неупорядоченностью, между высокой молекулярной оранизацией и дезорганизацией. Напротив, для живых существ характерна упорядоченность как в их структуре и функциях, так и в превращении и использовании энергии. Таким образом, сохраняя внутреннюю упорядоченность, но получая свободную энергию с солнечным светом или пищей, живые оранизмы возвращают в среду эквивалентное количество энергии, но в менее полезной форме, в основном в виде тепла, которое, рассеиваясь, уходит во Вселенную.
Процессы обмена веществ и энергии подвержены регуляции, причем существует множество регулирующих механизмов. Главнейшим механизмом регуляции метаболизма является контроль количества ферментов. К числу регулирующих механизмов относят также контроль скорости расщепления субстрата ферментами, а также контроль каталитической активности ферментов. Метаболизм подвержен так называемому обратному аллостерическому контролю, заключающемуся в том, что во многих биосинтетических путях первая реакция может быть ингибирована (подавлена) конечным продуктом. Можно сказать, что такое ингибирование происходит по принципу обратной связи. В регуляции обмена веществ и энергии имеет значение и то, что метаболические пути синтеза и распада почти всегда разобщены, причем у эукариотов это разобщение усиливается компартментализацией клеток. Например, местом окисления жирных кислот в клетках являются митохондрии, тогда как их синтез происходит в цитозоле. Многие реакции метаболизма подвержены некоторой регуляции со стороны так называемого энергетического статуса клетки, показателем которого является энергетический заряд, определяемый суммой молярных фракций АТФ и АДФ. Энергетический заряд в клетке всегда постоянен. Синтез АТФ ингибируется высоким зарядом, тогда как использование АТФ стимулируется таким же зарядом.
Благодаря содержанию растворов солей, Сахаров и других осмотически активных веществ, клетки характеризуются наличием в них определенного осмотического давления. Например, давление в клетках животных (морских и океанических форм) достигает 30 атм и более. В клетках растений осмотическое давление является еще большим. Разность концентрации веществ внутри и снаружи клетки называют градиентом концентрации.
Поступление веществ в клетки животных, равно как и удаление их из клеток, связано с проницаемостью клеточной мембраны для молекул или ионов, а также со свойствами веществ. Клеточная мембрана регулирует обмен различными веществами между клеткой и средой. Поддержание мембраны и ее проницаемость обеспечиваются клеточной энергией.
Известно несколько путей поступления веществ в клетки. В частности, различают пассивный, катализируемый и активный транспорт веществ в клетки, а также проникновение веществ в клетки путем эндоцитоза в виде фагоцитоза и пиноцитоза. Пассивный, катализируемый и активный транспорт обеспечивают проникновение в клетки лишь малых молекул, тогда как эндоцитоз ответственен за поступление в клетки макромолекул (белков, по-линуклеотидов, полисахаридов) и разных твердых частиц, включая бактерии. В то же время клетки способны секретировать различные вещества в окружающую их среду. Этот процесс называют экзоцитозом.
Пассивный транспорт веществ в клетки обеспечивается диффузией через мембрану по градиенту концентрации. Молекулы обычно переходят из области высокой концентрации в область более низкой концентрации.
Количество работы, затрачиваемой на обеспечение транспорта молекул в клетку против градиента концентрации, можно определить, исходя из допущенияпростой реакции, в которой Ац есть концентрация молекул за пределами клетки, a Ai есть концентрация молекул внутри клетки. Эту реакцию можно описать константой равновесия в виде уравнения:
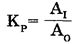
Между тем константа равновесия Кр
связана со свободной энергией реакции отношением в виде G = RTInКр
, где R есть около 2 кал/моль, а Т есть 25°С (температура, при которой протекают многие биологические реакции). Допуская, что совместная энергия гидролиза АТФ к АДФ обеспечивает эту реакцию с 50-процентной эффективностью, можно далее допустить, что транспортная система будет располагать примерно 3500 калориями (из общего количества энергии в 7000 калорий) на 1 моль АТФ, гидролизуемого при определенных физиологических условиях. Следовательно, константа равновесия будет равна:
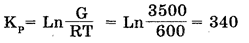
Важнейший вывод их этих заключений состоит в том, что работа, необходимая для транспорта какой-либо молекулы, не зависит от абсолютных концентраций. Она зависит от отношений между концентрациями внутри и вне клетки.
Когда транспортируются незаряженные молекулы, то пассивный транспорт определяется только градиентом концентрации, т. е. разностью концентрации вещества на разных сторонах мембраны. Если же молекулы транспортируемого вещества заряжены, то к влиянию градиента концентрации добавляется влияние электичес-ких потенциалов по обе стороны мембраны. Градиент концентрации и электрический градиент в совокупности составляют электрохимический потенциал, который позволяет транспорт в клетку только положительно заряженных ионов.
Можно сказать, что пассивный транспорт веществ в клетки осуществляется обычной диффузией через клеточную мембрану, причем скорость диффузии вещества зависит от его растворимости в мембране, коэффициента диффузии в мембране и от разности концентрации веществ в клетке и за ее пределами (в среде). Этим путем в клетку проникают вода, двуокись углерода и молекулы органических веществ, способные хорошо растворяться в жирах. Вещества в клетку проникают через поры, имеющиеся в клеточной мембране. Пассивный транспорт не зависит от энергии, обеспечиваемой АТФ.
Известна катализируемая, или так называемая «облегченная» диффузия, при которой скорость диффузии разных веществ, например, Сахаров, аминокислот и нуклеозидов через мембрану повышается с помощью белков (ферментов). Как и обычная диффузия, «облегченная» диффузия тоже зависит от градиента концентрации, однако здесь имеются подвижные «переносчики», роль которых выполняют ферменты. Находясь в составе мембраны, ферменты действуют в качестве «переносчиков» молекул веществ, проникая (диффундируя) на противоположную сторону мембраны, где они освобождаются от переносимых веществ. Поскольку «облегченная» диффузия веществ является переносом по градиенту концентрации, она тоже непосредственно не зависит от энергии, обеспечиваемой АТФ.
Активный транспорт веществ в клетку отличается от пассивного (диффузии) тем, что вещество переносится против градиента концентрации, т. е. из области низкой концентрации в область более высокой концентрации. Активный транспорт связан со способностью мембраны поддерживать разность электрических потенциалов (помимо поддержания разности в концентрациях веществ внутри и снаружи клетки), под которыми понимают различия между электрическими потенциалами внутри и вне клетки, а также с затратами энергии на работу в виде перемещения веществ против электрохимического градиента, т. е. «вверх».
Энергия для транспорта обеспечивается фосфоэнолпируватом, фосфатная группа которого и часть химической энергии которого передаются белкам, часть которых используется всеми сахарами, транспортируемыми фосфотрансферазной системой, а часть специфична для отдельных Сахаров. Конечный белок содержится в мембране и ответственен за транспорт и фосфорилирование Сахаров.
 Активный транспорт особенно эффективен в случае переноса ионов. Реакции, обеспечивающие активный транспорт, происходят в мембране и сопряжены с реакциями, дающими свободную энергию. Ферменты, катализирующие эти реакции, также локализованы в мембране. Примером активного транспорта веществ является транспорт ионов натрия и калия (рис. 69), который определяет клеточный мембранный потенциал. Концентрация ионов натрия (Na+
) внутри большинства клеток является меньшей, чем в среде, тогда как концентрация ионов калия (К+
) внутри клеток является в 10—20 раз большей, чем в среде. В результате этого ионы Nа+
стремятся проникнуть из среды в клетку, а ионы К+
, наоборот, выйти из клетки в среду. Поддержание концентрации этих ионов в клетке и в окружающей среде обеспечивается благодаря наличию в клеточной мембране системы, которая является ионным «насосом» и которая откачивает ионы Na+
из клетки в среду и накачивает ионы К+
в клетку из среды. Работа этой системы, т.е. движение ионов против электрохимического градиента, обеспечивается энергией, которая генерируется гидролизом АТФ, причем фермент АТФ-аза, катализирующий эту реакцию, содержится в самой мембране и, как считают, выполняет роль натриево-калиевого «насоса», генерирующего мембранный потенциал. Энергия, освобождаемая при гидролизе одной молекулы АТФ, обеспечивает транспорт за пределы клетки трех ионов Na+
и внутрь клетки двух ионов К+
. Активный транспорт особенно эффективен в случае переноса ионов. Реакции, обеспечивающие активный транспорт, происходят в мембране и сопряжены с реакциями, дающими свободную энергию. Ферменты, катализирующие эти реакции, также локализованы в мембране. Примером активного транспорта веществ является транспорт ионов натрия и калия (рис. 69), который определяет клеточный мембранный потенциал. Концентрация ионов натрия (Na+
) внутри большинства клеток является меньшей, чем в среде, тогда как концентрация ионов калия (К+
) внутри клеток является в 10—20 раз большей, чем в среде. В результате этого ионы Nа+
стремятся проникнуть из среды в клетку, а ионы К+
, наоборот, выйти из клетки в среду. Поддержание концентрации этих ионов в клетке и в окружающей среде обеспечивается благодаря наличию в клеточной мембране системы, которая является ионным «насосом» и которая откачивает ионы Na+
из клетки в среду и накачивает ионы К+
в клетку из среды. Работа этой системы, т.е. движение ионов против электрохимического градиента, обеспечивается энергией, которая генерируется гидролизом АТФ, причем фермент АТФ-аза, катализирующий эту реакцию, содержится в самой мембране и, как считают, выполняет роль натриево-калиевого «насоса», генерирующего мембранный потенциал. Энергия, освобождаемая при гидролизе одной молекулы АТФ, обеспечивает транспорт за пределы клетки трех ионов Na+
и внутрь клетки двух ионов К+
.
Система Na+
+ К+
—АТФ-аза помогает поддерживать ассиметрическое распределение ионов калия при высокой концентрации последнего в клетках. Ионы калия участвуют в регуляции многих клеточных функций, включая поток солей и воды из почечных клеток, освобождение инсулина из панкреатических клеток, частоту сердцебиений.
Установлено, что энергетически выгодный транспорт ионов Na+
внутрь клеток оказывает также влияние на транспорт сахаров и аминокислот в клетки. В частности, с транспортом ионов Na+
сопряжен транспорт глюкозы. Чтобы создать градиент концентрации ионов Na+
, благоприятный для транспорта ионов К+
и глюкозы внутрь клеток, ионная «насосная» система благодаря энергии активно откачивает ионы Na+
из клетки за ее пределы.
Определенная роль в транспорте веществ принадлежит белоксвязывающим системам, представляющим четвертый способ транспорта. Речь идет о белках, локализованных в периплазматическом пространстве. Эти белки специфически связывают сахара, аминокислоты и ионы, перенося их затем к специфическим молекулам-носителям, локализованным в клеточной мембране. Источником энергии для этих систем является АТФ.
Эндоцитоз
, как отмечено выше, обеспечивает перенос в клетки крупных частиц и молекул. В рамках эндоцитоза различают фагоцитоз и пиноцитоз.
Фагоцитоз (от греч. phagos — пожирающий и cytos — клетка) представляет собой процесс, заключающийся в том, что клетки-лейкоциты (макрофаги и нейтрофилы) захватывают (обволакивают) твердые частицы (фрагменты клеток, бактерии) путем выпячиваний своей клеточной мембраны и образования пузырьков, сливающихся затем с плазматической мембраной и открывающихся внутрь клетки. Вошедшие внутрь клеток частицы поступают в лизосомы, где с помощью клеточных (лизосомных) ферментов разрушаются и усваиваются затем клетками. Фагоцитоз широко распространен среди одноклеточных организмов. У многоклеточных (млекопитающих) он выполняется специализированными клетками (лейкоцитами).
У простейших фагоцитоз является формой питания, в результате которого твердые частицы проникают в лизосомы, где и перевариваются, образуя продукты, служащие пищей. Биологическое значение фагоцитоза у млекопитающих заключается в том, что он обеспечивает иммунную (фагоцитарную) защиту организма (см. гл. XVII).
Пиноцитоз (от греч. pino — пить и cytos — клетка) представляет собой процесс, при котором клетки поглощают жидкости и находящиеся в них высокомолекулярные вещества путем впячива-ний плазматической мембраны и образования пузырьков (каналь-цев), куда поступает жидкость. Канальцы после заполнения жидкостью отшнуровываются, поступают в цитоплазму и доходят до лизосом, где их стенки перевариваются, в результате чего содержимое (жидкость) канальцев освобождается и подвергается дальнейшей обработке лизосомными ферментами.
Пиноцитоз часто встречается у одноклеточных животных, у многоклеточных он наблюдается в клетках кровеносной и лимфатической систем, в клетках злокачественных опухолей, а также в клетках тканей, для которых характерен повышенный уровень обмена веществ.
Экзоцитоз
— это процесс секретирования клетками различных веществ, причем известны регулируемый и конститутивный пути экзоцитоза. Примером регулируемого экзоцитоза является экзоци-тоз инсулина. Клетки поджелудочной железы, продуцирующие инсулин, упаковывают его вначале в так называемые секреторные пузырьки, которые после внеклеточного сигнала сливаются с плазматической мембраной, а затем открываются в межклеточное пространство, освобождая гормон. Подобным образом происходит эк-зоцитоз других гормонов, нейротрансмиттеров и многих ферментов. Напротив, конститутивный путь экзоцитоза присущ многим белкам, непрерывно синтезируемым клетками и упаковываемым в эк-зоцитозные пузырьки в комплексе Гольджи, после чего эти пузырьки перемещаются к плазматической мембране, где и открываются в межклеточное пространство, освобождаясь от белкового содержимого.
С помощью экзоцитоза из клетки удаляются также частицы, оказавшиеся непереваренными путем фагоцитоза. У большинства клеток циклы эндоцитоз-экзоцитоз непрерывны.
Фотосинтез
— это синтез органических соединений в листьях зеленых растений из воды и углекислого газа атмосферы с использованием солнечной (световой) энергии, адсорбируемой хлорофиллом в хлоропластах. Благодаря фотосинтезу происходит улавливание энергии видимого света и превращение ее в химическую энергию, сохраняемую (запасаемую) в органических веществах, образуемых при фотосинтезе (рис. 70). Значение фотосинтеза гигантское. Отметим лишь, что он поставляет топливо (энергию) и атмосферный кислород, необходимые для существовария всего живого. Следовательно, роль фотосинтеза является планетарной.
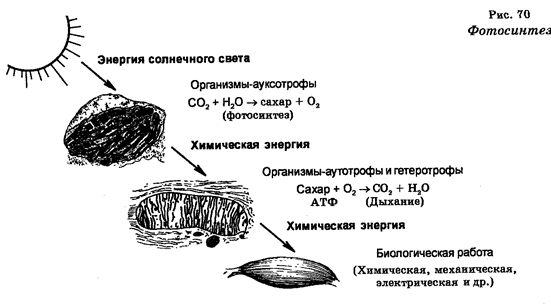
Планетарность фотосинтеза определяется также тем, что благодаря круговороту кислорода и углерода (в основном) поддерживается современный состав атмосферы, что в свою очередь определяет дальнейшее поддержание жизни на Земле. Можно сказать далее, что энергия, которая запасается в продуктах фотосинтеза, есть по существу основной источник энергии, которым сейчас располагает человечество.
Химию фотосинтеза описывают следующими уравнениями:
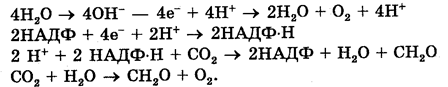
Как отмечено выше, фотосинтез происходит в хлоропластах зеленых растений.
Фотосинтез начинается с улавливания и поглощения света пигментом хлорофиллом, содержащимся в хлоропластах клеток зеленых растений. Когда свет падает на молекулу хлорофилла, то один из ее электронов оказывается в возбужденном состоянии. Другими словами, он переходит на более высокий энергетический уровень. Возбужденные электроны передаются затем другими молекулами, в результате чего повышается свободная энергия молекулы-акцептора, а «брешь», образованная в молекуле хлорофилла, заполняется электроном, поступающим из воды. Последняя при этом окисляется, в результате чего выделяется молекулярный кислород. Таким образом, в молекулах хлорофилла световая энергия переводит электроны на более высокий энергетический уровень. Хлорофилл является промежуточным соединением на пути электронов от низкоэнергетического уровня в молекулах воды к высокоэнергетическому уровню в конечном акцепторе электронов.
В переходе электронов на высокий энергетический уровень участвуют две содержащиеся в хлоропластах фотосистемы, образованные хлорофиллом и особыми белками — фотосистема I, активируемая далеким красным светом (-700 нм) и фотосистема II, активируемая красным светом с более высокой энергией (-650 нм), т. е. этот переход происходит в два этапа при использовании света. Реакции, протекающие на этих этапах, получили название световых. Обе фотосистемы связаны между собой системой переноса электронов.
На уровне фотосистемы I молекулы хлорофилла передают свои электроны, богатые энергией, через ферредоксин к никотин-ами-даденин-динуклеотидфосфату (НАДФ), который в результате этого восстанавливается в НАДФЧН ив восстановленной форме уже сам способен самостоятельно поставлять электроны, необходимые для образования глюкозы путем восстановления атмосферной СОу После перехода электронов в НАДФЧН из молекулы хлорофилла в последних остаются своеобразные «бреши».
На уровне фотосистемы II богатые энергией возбужденные электроны хлорофилла передаются системе переноса электронов, а образовавшиеся в молекулах хлорофилла «бреши» после «ушедших» электронов замещаются бедными энергией электронами, которые поступают от воды, окисляющейся с образованием молекулярного кислорода. Пройдя через ряд соединений, составляющих цепь переноса электронов, электроны из фотосистемы II, богатые энергией, в конечном итоге замещают утраченные электроны в хлорофилле из фотосистемы I.
В цепи переноса электронов осуществляется несколько окислительно-восстановительных реакций, в каждой из которых электроны переходят на более низкий энергетический уровень.
Часть энергии, теряемой при переходе через цепь переноса электронов, идет на обеспечение синтеза АТФ из АДФ и неорганического фосфата. Считают, что синтез молекул АТФ связан также с фотосистемой I, в которой имеется циклический поток электронов, заключающийся в том, что электроны, захваченные акцептором, возвращаются хлорофиллу через цитохром В. При этом энергия, высвобождающаяся в реакциях систем переноса электронов, в которых электроны двигаются «вниз», запасается путем синтеза молекул АТФ.
В результате световых реакций фотосинтеза образуются высокоэнергетические АТФ и восстановленный НАДФ, которые снабжают энергией последующие, так называемые темновые реакции, протекающие без света и приводящие, в конце концов, к восстановлению атмосфертной COg до Сахаров. Источником энергии здесь является АТФ, а восстанавливающим агентом — НАДФ-Н, синтезируемые в реакциях фотосинтетического переноса электронов. Процесс восстановления COg начинается с катализируемой ри-булозобисфосфаткарбоксилазой фиксации молекул этого соединения молекулами акцептора и сопровождается вступлением атомов углерода в ряд последовательных реакций, что приводит к образованию на каждые шесть фиксированных молекул COg одной молекулы глюкозы, причем связывание одной молекулы COg обеспечивается затратой трех молекул АТФ и двух молекул НАДФ-Н.
Как отмечено выше, энергия и электроны, необходимые для темновых реакций, поставляются АТФ и восстановленным НАДФ, образованными в световых реакциях.
Таким образом, химическая энергия, генерированная световыми
реакциями, стабилизируется в молекулах глюкозы в процессе темновых
реакций. В конечном итоге из глюкозы образуется крахмал, который является ее высокомолекулярным полимером, в котором оказываются запасенными по существу как атомы углерода, так и энергия. Полимеризуясь, глюкоза образует также целлюлозу. Подсчитано, что в листьях зеленых растений Земли и в фитопланктоне водоемов ежегодно синтезируется около 150 млрд тонн органических веществ и выделяется в атомсферу около 200 млрд тонн кислорода.
Фотосинтез имеет большую древность. Предполагают, что круговорот углерода, т. е. фотосинтез, существовал уже 3,5 х 109
лет назад.
Хемосинтез
— это синтез органических веществ с помощью энергии, генерируемой окислением неорганических соединений, например, аммиака, оксида железа, сероводорода. Хемосинтез был открыт С. Н. Виноградским в 1889-1890 гг. Его осуществляют бактерии разных видов. Рассмотрим некоторые из наиболее известных примеров, начав с нитрифицирующих бактерий, роль которых была показана С. Н. Виноградским.
Нитрифицирующие бактерии являются обитателями почвы. Они получают энергию окислением аммиака, образующегося в почве в результате разложения белков (остатков животных и растений). Реакция окисления аммиака может быть описана следующим уравнением:
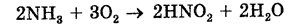
В этой реакции выделяется энергия в количестве бй2 кДж. Образующаяся в ходе этой реакции азотистая кислота окисляется нитрифицирующими бактериями другого вида до азотной кислоты с выделением энергии в количестве 101 кДж. Эта реакция описывается следующим уравнением:
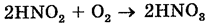
Энергия, освобождаемая в этих реакциях, используется для синтеза органических веществ.
Серобактерии получают энергию, окисляя сероводород. Этот процесс можно описать следующим уравнением:
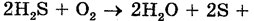 энергия энергия
Образующаяся в результате этой реакции свободная сера накапливается в цитоплазме серобактерий. Если недостает далее сероводорода, то происходит окисление свободной серы в бактериальной цитоплазме с дальнейшим освобождением энергии:
 энергия энергия
Эта энергия используется для синтеза органических веществ из углекислого газа.
Хемосинтезирующие бактерии окисляют также соединения железа и марганца. Считают, что образование залежей железных и марганцевых руд является результатом деятельности микроорганизмов в прошлые геологические эпохи (В. И. Вернадский).
У растений источником энергии является солнечный свет, причем ответственными за производство АТФ являются хлоропласты. Энергия, которая оказывается запасенной в основном в углеводах, используется в дальнейшем клетками растений для обеспечения различных биологических реакций. Что же касается клеток животных, то энергия поступает к ним с пищей (сахарами и жирными кислотами). Чтобы эта энергия могла использоваться в процессе жизнедеятельности клеток, она должна быть подготовлена для использования. Каковы же механизмы, с помощью которых энергия, запасенная в глюкозе, трансформируется в клетках в доступную для использования форму АТФ?
Подготовка энергии к использованию, т. е. генерирование (извлечение) энергии из пищевых веществ осуществляется в процессе дыхания, под которым понимают окисление (расщепление) молекул-энергоносителей, т. е. «топливных» молекул, при котором роль конечного акцептора электронов выполняет О у а донором электронов является органическое или неорганическое соединение. Процесс подготовки энергии к использованию протекает в три последовательные стадии (рис. 71).
На первой стадии поступающие в клетки крупные молекулы полисахаридов гидролизуются до простых Сахаров. На этой стадии происходит разложение и других энергоносителей. В частности, жиры разлагаются на глицерол и жирные кислоты, белки гидро-лизуются до аминокислот. Однако на этой стадии высвобождение запасенной в пищевых веществах энергии все еще не происходит.
На второй стадии происходит распад малых молекул до еще более простых структур, играющих уже ключевую роль в метаболизме. Глюкоза превращается в ацетильную часть ацетил-КоА, являющегося производным кофермента А. В результате этих реакций образуются молекулы АТФ, но их еще мало. На уровне ацетил-КоА в метаболический путь могут вступать также жирные кислоты и аминокислоты.
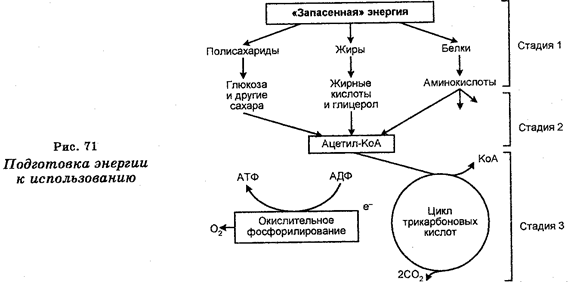
Наконец, на третьей стадии происходит полное окисление ацетильного компонента ацетил-КоА до СОу На этой стадии образуется основная часть АТФ.
Процесс генерирования энергии в животных клетках (извлечения ее из субстрата) осуществляется с участием митохондрий и начинается с гликолиза (от греч. glycos — сахар и lysis — растворение), который представляет собой окисление глюкозы, заканчивающееся превращением этого углевода в пировиноградную кислоту и образованием АТФ. Уже давно установлено, что для дыхания в качестве акцептора электронов необходим кислород. Однако на первых этапах расщепления Сахаров кислорода не требуется. Окисление глюкозы начинается в анаэробных условиях дыхания (при отсутствии кислорода) с частичного расщепления ее шестиуглеродной молекулы и заканчивается образованием двух трехуглеродных молекул пировиноградной кислоты (рис. 72). Превращения глюкозы можно описать следующим уравнением:
C
6
H12
O6
+ 2Ф + 2АДФ ® 2СН3
СНОНСООН + 2АТФ + 2Н2
О
У большинства организмов гликолиз служит одним из центральных метаболических путей и состоит из десяти последовательных химических реакций, протекающих в цитоплазме (цитозол). Вначале глюкоза превращается через глюкозо-6-фосфат во фрукто-зо-1,6-дифосфат в результате фосфорилирования, катализируемого гексокиназой и фосфофруктозокиназой. Поскольку эти реакции еще сами нуждаются в АТФ, они являются подготовительными в образовании АТФ. В частности, на превращение каждой молекулы глюкозы в этих реакциях затрачивается по две молекулы АТФ.
На втором этапе фруктозо-1,6-дифосфат превращается с помощью альдолазы в дигидроксиацетонфосфат и глицераль-дегид-3-фосфат, которые взаимопревращаемы в реакциях, катализируемых триозофосфатизомеразой. Затем глицеральдегид-3-фосфат окисляется и фосфорилизуется, в результате чего превращается в высокоэнергетическое фосфатное соединение 1,3-дифосфоглицерат (1,3-БФГ). Это превращение катализируется глицеральдегид-3-фосфатдегидрогеназой.
 Поскольку 1,3-БФГ-ацилфосфат обладает высоким потенциалом переноса фосфатной группы, эта особенность используется для генерирования АТФ. Поэтому дальше происходит перенос фосфатной группы от ацилфосфатной группы 1,3-БФГ-ацилфосфата на АДФ, катализируемый фосфоглицераткиназой. В результате этого образуется молекула АТФ и 3-фосфоглицерат, т. е. на этом этапе происходит образование энергии. Поскольку 1,3-БФГ-ацилфосфат обладает высоким потенциалом переноса фосфатной группы, эта особенность используется для генерирования АТФ. Поэтому дальше происходит перенос фосфатной группы от ацилфосфатной группы 1,3-БФГ-ацилфосфата на АДФ, катализируемый фосфоглицераткиназой. В результате этого образуется молекула АТФ и 3-фосфоглицерат, т. е. на этом этапе происходит образование энергии.
Последний этап гликолиза связан с превращением 3-фосфоглицерата в пируват и образованием второй молекулы АТФ. Этот этап осуществляется в реакциях трех типов. Первая реакция заключается во внутримолекулярной перестройке, связанной с превращением 3-фосфоглицера-та в 2-фосфоглицерат, катализируемым фосфоглицеромутазой.
В результате этой реакции происходит перемещение фосфатной группы. Во второй реакции происходит дегидратация 2-фосфоглицерата, катализируемая енолазой, в результате чего образуется фосфоенолпируват. При этом повышается потенциал переноса фосфатной группы енолфосфат обладает высоким потенциалом переноса фосфатной группы. В третьей (заключительной) реакции этого этапа гликолиза происходит перенос фосфатной группы, от фосфоенолпирувата к АДФ, катализируемый пируваткиназой. Это приводит к образованию пирувата и АТФ (второй молекулы).
При анаэробных условиях пировиноградная кислота превращается в молочную кислоту (лактат) или в этиловый спирт (этанол), или в пропионовую кислоту. Этот анаэробный процесс называют еще брожением. В данном случае речь идет о молочнокислом, спиртовом и пропионовом брожении (соответственно). Молочная кислота образуется из пирувата при метаболизме ряда микроорганизмов, а также в клетках мышц многоклеточных организмов. Суммарная реакция превращения глюкозы в лактат имеет следующий вид:
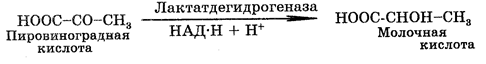
НАД×Н образуется в результате окисления глицеральдегид-3-фосфата, который используется при восстановлении пирувата.
В процессе превращения пировиноградной кислоты в лактат происходит регенерирование НАД+
, что поддерживает непрерывность гликолиза в анаэробных условиях. Этиловый спирт образуется из пирувата при метаболизме дрожжей и некоторых других микроорганизмов спиртового брожения. Суммарная реакция превращения глюкозы в этанол имеет следующий вид:
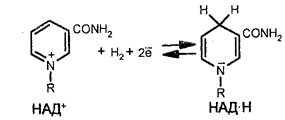
Глюкоза + 2Pi + 2АДФ + 2Н+
® 2 этанол + 2СО2
+ 2АТФ + 2Н2
0.
Восстановление ацетальдегида в этиловый спирт сопровождается регенерированием НАД+
.
Анаэробное дыхание с точки зрения производительности не является эффективным процессом, т. к. при анаэробном превращении глюкозы в этанол или лактат освобождается лишь небольшое количество энергии. Большая часть энергии, запасенная в глюкозе, продолжает затем оставаться запасенной уже в молекулах этанола.
Как видно, последовательность реакций, в процессе которых глюкоза превращается в пируват, сходна в клетках всех видов у всех организмов. Биологическое значение гликолиза заключается в том, что он генерирует молекулы АТФ. В результате распада глюкозы образуются строительные блоки, используемые для синтеза клеточных структур. Оба эти процесса регулируются скоростью превращения глюкозы в пируват. Однако роль пирувата в генерировании энергии обмена веществ различна в разных клетках и разных организмах.
У аэробных организмов гликолиз, осуществляемый в цитозоле выполняет роль своего рода процесса-прелюдии к дальнейшему окислению, ибо при аэробном дыхании (в присутствии кислорода) окисление идет дальше и осуществляется уже в митохондриях в так называемом цикле Кребса (цикле трикарбоновых кислот или цикле лимонной кислоты) и в цепи переноса электронов, цикл Кребса является конечным путем окисления топливных молекул, причем не только глюкозы и других углеводов, но и жирных кислот и аминокислот (рис. 73). Следовательно, «топливом» для окисления в митохондриях являются пируват и жирные кислоты. Включение в этот окислительный путь осуществляется на уровне кофермента (ацетил-КоА), т. е. происходит с образования ацетил-КоА в митохондриаль-ном матриксе, в результате окислительного декарбоксилирования пирувата или распада жирных кислот до двухуглеродных групп. Ацетил-КоА обладает высоким потенциалом переноса ацетильных групп. Следовательно, топливные молекулы вступают в цикл Кребса в виде ацетил-КоА. Непрерывность же снабжения окислительных процессов «топливом» обеспечивается запасанием животными клетками липидов, являющихся главным ресурсом жирных кислот, а также гликогена, являющегося источником глюкозы.
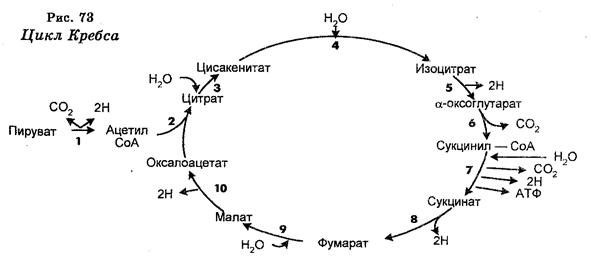
Цикл Кребса действует только в аэробных условиях и начинается с конденсации ацетил-КоА (C2
) и оксалоацетата (C4
с образованием цитрата (С6
), изомеризация которого приводит к изоцитрату (C6
). Затем следует окислительное декарбоксилирование изоцитрата и образование a-оксоглутарата (С5
), после чего последний подвергается окислительному декарбоксилированию (выделяется вторая молекула CO2
в сукцинил-КоА (С4
). В следующей реакции происходит расщепление тиоэфирной связи сукцинил-КоА в присутствии пирофосфата (Pi), в результате чего образуется сукцинат и генерируется высокоэнергетические фосфатные связи в форме ГТФ и АТФ.
Сукцинат потом окисляется в фумарат (С4
), а последний гидратируется в малат. В последующей реакции происходит окисление малата, что приводит к регенерированию оксалоацетата (C4
). Следовательно, в цикл Кребса вступают два атома углерода в виде ацетил-КоА и такое же количество атомов углерода покидают этот цикл уже в виде CO2
в последовательных реакциях декарбоксилирования, которые катализируются дегидрогеназами.
 
В результате четырех окислительно-восстановительных реакций цикла Кребса происходит перенос трех пар электронов над НАД и одной пары электронов на ФАД. Восстановленные этим путем переносчики электронов НАД и ФАД подвергаются затем окислению уже в цепи переноса электронов, в результате которого генерируется одиннадцать молекул АТФ. Одна высокоэнергетическая связь генерируется непосредственно в цикле Кребса. Таким образом, на каждый двухуглеродный фрагмент, полностью окисляемый до Н2
О и СО2
, генерируется двенадцать высокоэнергетических фосфатных связей.
Цикл Кребса подвержен регуляции; его скорость зависит от потребности в АТФ других метаболических реакций. Важное значение имеет регуляция синтеза цитратсинтазы, изоцитратдегидроге-назы и оксоглутаратдегидрогеназы.
Биологическое значение цикла Кребса заключается не только в том, что он является завершающим этапом в генерировании энергии, но и в том, что он «поставляет» промежуточные продукты для биосинтеза.
Цикл Кребса действует только в аэробных условиях по той причине, что для него необходимы НАД и ФАД, регенерирование которых происходит при переносе электронов НАД×Н и ФАД×H2
на О2
по цепи транспорта электронов, сопровождаемом одновременным образованием АТФ (рис. 74).
Поскольку у аэробных организмов единственным акцептором электронов является О2
, а электроны не переносятся от топливных молекул и продуктов их реакций прямо на O2
, топливные молекулы и продукты их распада переносят электроны к пиримидиннуклеотидам или флавинам, являющимся переносчиками.
Главным акцептором электронов при окислении топливных молекул является никотинамидадениндинуклеотид (НАД+
, реакционноспособной частью которого является никотинамидное кольцо. Последнее присоединяет ион водорода и два электрона. Восстановленной формой этого переносчика является НАД×Н. Окисление последнего дает три молекулы АТФ. Вторым акцептором электронов является ФАД (флавинадениндинуклеотид), ре-акционноспособной частью которого является изоаллоксазиновое кольцо, которое тоже присоединяет два электрона. Восстановленной формой ФАД является ФАД×H2
. Окисление последнего дает две молекулы АТФ.
Таким образом, главными переносчиками являются НАД×Н и ФАД-Hg, которые содержат по паре электронов с высоким потенциалом и которые доставляют свои высокоэнергетические электроны к О3
по цепи транспорта электронов, также локализованной в митохондриях.
Этот перенос сопровождается образованием АТФ из АДФ и пи-рофосфата (Рi), происходит на митохондриальных мембранах и носит название окислительного фосфорилирования. Оно было открыто в 1931 г. В. А. Энгельгардтом (1894-1984). Следовательно, окислительное фосфорилирование — это процесс образования АТФ, сопряженного с переносом электронов по цепи транспорта (переносчиков) от НАД×Н или ФАД×H2
к O2
через многие другие переносчики, в частности питохромы. В процессе окислительного фосфорилирования генерируется 32 молекулы АТФ из всех 36 молекул АТФ, генерируемых в процессе окисления глюкозы до СО2
и Н2
О.
Многоступенчатость транспорта электронов от НАД×Н или ФАД×Н2
к О2
по цепи многочисленных переносчиков сопровождается выбросом протонов из митохондриального матрикса и генерированием на внутренней митохондриальной мембране протон-движущей силы (мембранного потенциала), измеряемой в милливольтах. На внутренней поверхности митохондриальной мембраны протондвижущая сила равна 220 микровольтам.
В процессе обратного перехода протонов в митохондриальный матрикс происходит синтез АТФ.
Следовательно, окисление НАД×Н и ФАД×H2
и фосфорилирование АДФ в АТФ сопряжены по той причине, что они обеспечиваются протонным градиентом через внутреннюю мембрану митохондрий. Это сопряжение называют дыхательным контролем.
Потенциальные возможности окисления в митохондриях очень большие, т. к. последние обеспечивают производство почти всего АТФ в клетках млекопитающих.
Благодаря фотосинтезу и дыханию световая энергия Солнца конвертируется в форму, которая может использоваться клетками для обеспечения всех выполняемых ими функций (рис. 75).
 Основными видами биологической работы в клетках являются транспорт веществ через мембраны, биологический синтез и механическая работа. Обеспечение этих видов биологической работы в клетках основано на цикле АТФ-АДФ. Для обеспечения энерготребующих функций клеток используются высокоэнергетические связи АТФ. В результате реакции в конечном итоге освобождается неорганический фосфат. АДФ рефосфорилируется в АТФ в процессе реакций катаболизма. Основными видами биологической работы в клетках являются транспорт веществ через мембраны, биологический синтез и механическая работа. Обеспечение этих видов биологической работы в клетках основано на цикле АТФ-АДФ. Для обеспечения энерготребующих функций клеток используются высокоэнергетические связи АТФ. В результате реакции в конечном итоге освобождается неорганический фосфат. АДФ рефосфорилируется в АТФ в процессе реакций катаболизма.
Большое место в катаболизме занимает биосинтез различных соединений, который в клетках происходит непрерывно. Больше того, клетки обладают гигантской биосинтетической способностью в отношении всех веществ. Например, одиночная клетка Е. coli способна за время от одного деления до другого (в процессе одного клеточного цикла) синтезировать огромное количество молекул различных соединений (табл. 7).
Таблица 7
Биосинтетическая способность Е. coli
| Химическое соединение
|
Сухой вес,(в%)
|
Молекулярная масса
|
Количество молекул на клетку
|
Количество молекул, синтезируемых за 1 сек
|
Количество молекул АТФ, необходимых для синтеза в течение секунды
|
Количество требуемой энергии (в %) к общей затрачиваемой энергии
|
| ДНК
|
5
|
2000000000
|
1
|
0,00083
|
60000
|
2,5
|
| РНК
|
10
|
1000000
|
15
|
12,5
|
75000
|
3,1
|
| Белки
|
70
|
60000
|
1700000
|
1,400
|
2120000
|
88
|
| Липиды
|
10
|
1000
|
15000000
|
12500
|
87500
|
3,7
|
| Полиса-хариды
|
5
|
200000
|
39000
|
32,5
|
65000
|
2,7
|
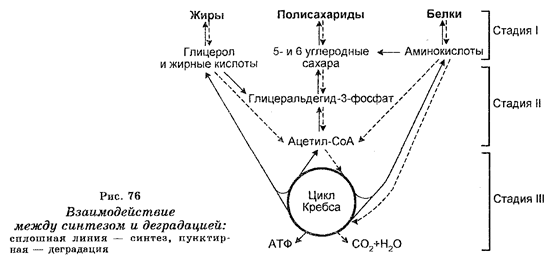
Центральное место в биосинтезе принадлежит синтезу белков (см. гл. XII). Синтез белков, нуклеиновых кислот и других химических соединений необходим для поддержания живых клеток. Во все биосинтезы вовлечен АТФ.
Больше того, между биосинтезом и деградацией химических соединений наблюдается взаимодействие, причем это взаимодействие обеспечивается АТФ (рис. 76).
Одним из обычных примеров механической работы является мышечное сокращение, в котором существенную роль играет АТФ.
По характеру ассимиляции различают автотрофные, гетеротрофные и миксотрофные организмы.
Автотрофные (от греч. autos — сам, trophe — пища), или самопитающиеся организмы, — это организмы, способные синтезировать органические соединения из неорганических (углекислого газа, воды и неорганических соединений азота и серы). В зависимости от источника потребляемой энергии автотрофы классифицируют на фотосинтезирующие и хемосинтезирующие организмы.
Первые используют световую энергию, тогда как вторые — энергию экзотермических химических реакций (в ходе превращения неорганических соединении), т. е. энергию, образующуюся при окислении различных неорганических соединений (водорода, сероводорода, аммиака и др.).
Фотосинтезирующими организмами являются растения, в листьях которых осуществляется фотосинтез.
Зеленые растения образуют углеводы, которые передвигаются из листьев в корни, где вступают в реакции с аммиаком и образуют аминокислоты.
Хемосинтезирующими организмами являются микроорганизмы — нитрифицирующие, серобактерии, водородные бактерии и железобактерии. Свободный азот усваивают азотфиксирующие бактерии.
Гетеротрофные (от греч. heteros — другой, trophe — пища) организмы — это организмы, которые нуждаются в готовых органических соединениях. Ими являются животные, а также микроорганизмы. Гетеротрофные организмы получают энергию путем окисления органических соединений (см. § 24).
Для животных характерен голозойный способ гетеротрофного питания, заключающийся в потреблении пищи в виде твердых частиц с последующей ее механической и химической переработкой.
Напротив, для микроорганизмов характерен осмотическим способ гетеротрофного питания. При этом способе питание происходит растворенными питательными веществами путем поглощения их всей поверхностью тела.
Миксотрофные (от лат. mixtus — смешанный) организмы — это организмы, способные как к синтезу органических веществ, так и к использованию их в готовом виде.
Например, эвглена зеленая на свету является автотрофом, в темноте — гетеротрофом.
По характеру диссимиляции различают аэробные и анаэробные организмы. Аэробные (от греч. aer — воздух) организмы для дыхания (окисления) используют свободный кислород. Аэробами является большинство ныне живущих организмов. Напротив, анаэробы окисляют субстраты, например, сахара в отсутствие кислорода, следовательно, для них дыханием является брожение. Анаэробами являются многие микроорганизмы, гельминты. Например, динитри-фицирующие анаэробные бактерии окисляют органические соединения, используя нитриты, являющиеся неорганическим окислителем.
Автотрофы и гетеротрофы связаны между собой питанием (пищевыми цепями) и энергетически, в результате чего существование одних из них зависит от других и наоборот.
Например, кислородные потребности аэробоз полностью зависят от автотрофов (зеленых растений). Последние используют СОу поставляемый в окружающую среду гетеротрофами. Все живые существа обладают системами, обеспечивающими превращение энергии и способны понижать энтропию.
Жизнедеятельность организмов с различными типами питания создает круговороты веществ в природе (см. гл. XVIII).
Обсуждение вопросов, касающихся происхождения типов обмена, очень осложнено незнанием первых этапов в развитии жизни. Поэтому формулируемые гипотезы не доступны экспериментальной проверке. Тем не менее предполагают, что когда возникла жизнь и когда ресурсы «первичного бульона» были исчерпаны, то у первых клеток возникла необходимость синтезировать ферменты, катализирующие образование органических молекул. Следовательно, селективные преимущества далее приобрели клетки, способные к биосинтезу. Со временем у клеток возникли различные метаболические пути. Предполагают, что центральным был метаболизм в виде гликолиза, ведущего к синтезу АТФ.
Считают, что первыми организмами, сходными, вероятно, с бактериями, были гетеротрофные анаэробы, способные использовать органические вещества абиогенного происхождения. Формирование цепи транспорта электронов позволило анаэробным бактериям использовать в качестве источника энергии те органические соединения, которые не подвергаются сбраживанию. Первые ге-теротрофы дали начало автотрофам, которые тоже были анаэробами. Позднее среди автотрофов появились организмы, способные осуществлять фотосинтез, что повело около 3,5—2 млрд лет назад к превращению СО2
в органическое соединение и к накоплению в атмосфере кислорода.
Предполагают, что первой стадией в возникновении фотосинтеза у примитивных организмов явился простейший синтез АТФ. Более поздние организмы возможно использовали водород для синтеза углеводов, после чего появились организмы, которые оказались способными использовать воду в качестве источника водорода и продуцировать кислород, т. е. уже быть фотосинтезирующими. Предполагают, что первые автотрофы использовали ферментные системы, развитые гетеротрофами. Поэтому на фотосинтез следует смотреть как на процесс, усложнявшийся в ходе исторического развития. В конечном итоге фотосинтезирующие организмы заселили все водоемы, а потом и сушу. Углеводы являются начальным продуктом фотосинтеза и сырым материалом для всех других структур, синтезируемых в растениях.
Оценивая значение фотосинтеза, можно заключить, что его эволюция привела к двум очень важным результатам. Первый результат заключается в том, что благодаря фотосинтезу возникло огромное количество видов организмов, которые оказались независимыми от пищи (корма) органического происхождения. Это создало условие для их процветания. Что же касается второго результата, то он заключается в том, что благодаря кислороду, освобождаемому в процессе биосинтеза, многие организмы стали извлекать из потребляемой пищи (корма) больше энергии, стали лучше компенсироваться их энергетические затраты.
Благодаря обилию кислорода и органических молекул возник процесс адаптации цепи транспорта электронов с НАДФ на кислород, что способствовало появлению аэробного типа диссимиляции. Предполагают далее, что первыми аэробами были зеленые растения и хемосинтезирующие микроорганизмы.
Особый вопрос связан с происхождением ферментов. Несомненно, что они являются эволюционным приобретением организмов. Предполагают, что в ходе эволюции ферменты усложнялись постепенно. Однако убедительных объяснений механизма возникновения первых ферментов нет, также как и нет объяснений характера первых ферментативных реакций.
1. Какова роль обмена веществ и энергии в жизни живых существ?
2. Что такое энергия и каковы ее формы?
3. Применимы ли к живым системам законы термодинамики?
4. Как организмы используют энергию? б. Какова связь между световой энергией и пигментами растений? Что происходит, когда свет падает на хлорофилл?
6. Почему энергия, запасенная в глюкозе, не может использоваться прямым образом для обеспечения биологических реакций?
7. Какова роль дыхания в подготовке энергии к использованию?
8. Какова роль гликолиза, цикла Кребса и цепи переноса электронов в «улучшении» энергии? Каково биологическое значение цикла Кребса?
9. Можете ли вы написать суммарное уравнение для превращения глюкозы в лактат?
10. Какова природа энергетической «валюты»?
11. Как используется АТФ в биологической работе?
12. Какова роль АТФ в транспорте ионов через клеточную мембрану?
13. Как вы понимаете роль АТФ в росте клеток и биосинтезе?
14. Полное окисление глюкозы дает 636 ккал энергии (глюкоза + О2
® Н2
О + СО2
+ 636 ккал). В процессе гликолиза глюкозы продуцируются две молекулы АТФ (глюкоза — 2 АТФ + 2 пировиноградная кислота). Распад одной молекулы АТФ до одной молекулы АДФ дает 8 ккал (АТФ ® АДФ + РО4
+ 8 ккал). Сколько энергии остается в двух молекулах пировиноградной кислоты, если допустить, что гликолиз эффективен на 100%?
15. Сколько молекул АТФ необходимо для синтеза в одну секунду ДНК, РНК, белка, липидов и полисахаридов в клетках человека, если допустить, что их около 3 х 102
в организме взрослого человека и что их биосинтетическая способность является такой же, как и в Е. coli (см. табл. 7).
16. Как много энергии необходимо для транспорта 3 эквивалентов Nа+
из клетки в жидкую среду и 2 эквивалентов К+
из жидкой среды в клетку при условии, что концентрация Na+
составляет в клетках 10 мМ, в среде — 100 мМ, тогда как концентрация K+
равна 100 мМ в клетке и б мМ в среде?
17. Опишите свойства автотрофов и гетеротрофов с точки зрения ввода и использования энергии.
18. Что представляют собой АТФ и АДФ?
19. Каковы пути поступления веществ в клетки?
20. Каким образом можно демонстрировать существование «насоса» Na+
-К+
в эукариотических клетках?
21. Какие виды биологической работы осуществляются в клетках?
22. Приведите примеры биосинтетической способности клеток.
23. Дайте определения и перечислите основные свойства автотрофных, гетеротрофных, и миксотрофных организмов.
24. Если концентрация какой-либо аминокислоты в клетке составляет 10-
3
, то как долго это количество аминокислот будет поддерживать синтез белка при частоте, количественно выражающейся в 1´10-13
г вновь синтезированного белка в течение 30 минут, затрачиваемых на удвоение клеток?
25. Что вы знаете о происхождении разных типов метаболизма?
26. Что вы понимаете под энтропией?
Альберте Б.. Брей Д., Льюис Дж., Рэфф М„ Роберте К., Уотсон Дж. Молекулярная биология клетки. М.: Мир. 1994. Т
.
1. 615
стр
.
Стройтер
Л
.
Биохимия
.
М
.:
Мир
.
1985.
Т
.
2. 240
стр
.
Bray D. Protein molecules as computational elements in living cells. Nature
, 1995,
Vol
. 376, 307-312.
Размножение — это свойство организмов производить потомство или способность организмов к самовоспроизведению. Являясь важнейшим свойством живого, размножение обеспечивает непрерывность жизни, продолжение видов
Процесс размножения исключительно сложен и связан не только с передачей генетической информации от родителей к потомству, но и с анатомическими и физиологическими свойствами организмов, с их поведением, гормональным контролем. Размножение организмов сопровождается процессами их роста и развития.
Для живых существ характерно чрезвычайное разнообразие в способах размножения. Тем не менее различают два основных способа размножения — бесполое и половое (рис. 77). Бесполое размножение, или апомиксис (от греч. аро — без, mixis — смешение), представляет собой процесс, в котором участвует лишь один родитель (клетка или многоклеточный организм). Напротив, в половом размножении участвует два родителя, каждый из которых имеет собственную репродуктивную систему и продуцирует половые клетки (гаметы), которые после слияния образуют зиготу (оплодотворенное яйцо), дифференцирующуюся затем в эмбрион. Следовательно, при половом размножении имеет место смешение наследственных факторов, т. е. процесс, называемый амфимикси-сом (от греч. amphi — с обеих сторон, mixis — смешение).
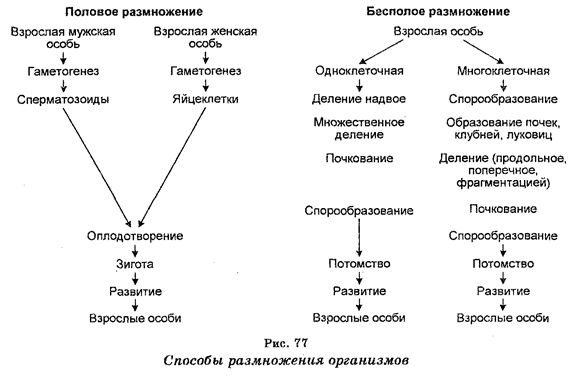
Бесполое размножение характерно для организмов многих видов как растений, так и животных. Оно встречается у вирусов, бактерий, водорослей, грибов, сосудистых растений, простейших, губок, кишечнополостных, мшанок и оболочников.
Наиболее простая форма бесполого размножения характерна для вирусов. Их репродуктивный процесс связан с молекулами нуклеиновых кислот, со способностью этих молекул к самоудвоению и основан на специфичности относительно слабых водородных связей между нуклеотидами.
Применительно к другим организмам, размножающимся бесполым путем, различают вегетативное размножение и размножение спорообразованием.
Вегетативное размножение — это размножение, при котором из части, отделившейся от материнского организма, развивается новый организм. Этот вид размножения характерен как для одноклеточных, так и многоклеточных организмов, но имеет у них разное проявление.
У одноклеточных организмов вегетативное размножение представлено такими формами, как деление, множественное деление и почкование. Деление путем простой перетяжки с образованием при этом из одного родительского организма двух дочерних присуще бактериям и сине-зеленым водорослям (цианобактериям). Напротив, размножение делением бурых и зеленых водорослей, а также одноклеточных животных (саркодовых, жгутиковых и инфузорий) происходит путем митотического деления ядра с последующей перетяжкой цитоплазмы.
Размножение путем множественного деления (шизогонии) заключается в делении ядра с последующим разделением цитоплазмы на части. В результате такого деления из одной клетки образуется несколько дочерних организмов. Примером множественного деления является размножение малярийного плазмодия (Р. vivax) в эритроцитах человека. В этом случае у плазмодиев происходит повторяющееся много раз деление ядра без цитокинеза, после чего следует и цитокинез. В результате этого один плазмодий дает начало 12-24 дочерним организмам.
У многоклеточных растительных организмов вегетативное размножение путем деления осуществляется черенками, луковицами, листьями, корневищами. Но это по существу искусственное размножение, используемое в сельскохозяйственной практике. Размножение высших растений в искусственных условиях возможно и из одной клетки. Организмы, развивающиеся из одной клетки, обладают всеми свойствами исходного многоклеточного организма. Это размножение получило название клонального микроразмножения. В качестве одной из форм вегетативного размножения могут служить прививки, или трансплантации, многих культурных растений, заключающиеся в пересадке почки или части побега от одного растения к другому. Конечно, это тоже способ размножения, который в природе не встречается, но в сельском хозяйстве используется очень широко.
У многоклеточных животных вегетативное размножение происходит путем фрагментации их тела на части, после чего каждая часть развивается в новое животное. Такое размножение характерно для губок, кишечнополостных (гидр), немертин, плоских червей, иглокожих (морских звезд) и некоторых других организмов. Близкой формой к вегетативному размножений животных фрагментацией является полиэмбриония животных, заключающаяся в том, что на определенной стадии развития эмбрион разделяется на несколько частей, каждая из которых развивается в самостоятельный организм. Полиэмбриония встречается у броненосцев. Однако последние размножаются половым путем. Поэтому полиэмбриония является скорее своеобразной стадией в половом размножении, а потомство, возникающее в результате полиэмбрионии, представлено монозиготными близнецами.
Почкование заключается в том, что на материнской клетке образуется бугорок (вырост) с ядром, который затем отделяется и становится самостоятельным организмом. Почкование встречается как у одноклеточных растений, например, у дрожжей, так и у одноклеточных животных, например, инфузорий отдельных видов.
Размножение спорообразованием связано с образованием специализированных клеток — спор, которые содержат ядро, цитоплазму, покрыты плотной оболочкой и способны к длительному существованию в неблагоприятных условиях, что способствует, кроме того, и их расселению. Наиболее часто такое размножение встречается у бактерий, водорослей, грибов, мхов, папоротникообразных.
У некоторых зеленых водорослей из отдельных клеток могут формироваться так называемые зооспоры.
Среди животных размножение снорообразованием отмечается у споровиков, в частности, у малярийного плазмодия.
У организмов многих видов бесполое размножение может чередоваться с половым размножением (см. § 31).
Половое размножение встречается как у одноклеточных, так и у многоклеточных растений и животных.
Как отмечено в главах II и III, половое размножение у бактерий осуществляется путем конъюгации, служащей аналогом полового процесса и являющейся системой рекомбинации этих организмов, тогда как у простейших половое размножение происходит тоже путем конъюгации либо путем сингамии и аутогамии.
У многоклеточных организмов (растений и животных) половое размножение связано с образованием зародошевых или половых клеток (гамет), оплодотворением и образованием зигот.
Половое размножение является значительным эволюционным приобретением организмов. С другой стороны, оно способствует пересортировке генов, появлению разнообразия организмов и повышению их конкурентоспособности в непрерывно меняющихся условиях окружающей среды.
У одноклеточных организмов половое размножение существует в нескольких формах. У бактерий половое размножение можно анало-гизировать с имеющими место у них конъюгацией, заключающейся в передаче плазмидной или хромосомной ДНК от клеток-доноров (содержащих плазмиды) к клеткам-реципиентам (не содержащих плазмиды), а также с трансдукцией бактерий, заключающейся в передаче генетического материала от одних бактериальных клеток к другим фагам. Конъюгация встречается также у инфузорий, у которых во время этого процесса происходит переход ядер от одних особей к другим, после чего следует деление последних.
Одной из распространенных форм полового размножения у одноклеточных животных, например, у паразитических споровиков, является копуляция. Половое размножение у них заключается в слиянии двух особей, которые являются гаметами, в одну, являющуюся споровой формой, из которой затем развивается новый организм. Независимо от способа слияния генетического материала у одноклеточных организмов существенной особенностью этого слияния является то, что оно сопровождается генетической рекомбинацией.
У многоклеточных растений и животных половое размножение происходит через образование женских и мужских половых клеток (яйцеклеток и сперматозоидов), последующее оплодотворение яйцеклетки сперматозоидом и образование зиготы. У растений половые клетки продуцируются в специализированных репродуктивных структурах, у животных они продуцируются в половых железах, называемых гонадами (от греч. gone — семя).
Между соматическими и половыми клетками животных существует важное различие. Оно заключается в том, что соматические клетки способны к делению, т. е. репродуцируют себя и кроме того из них образуются половые клетки. Напротив, половые клетки не делятся, но они «начинают» репродукцию целого организма.
Диплоидные соматические клетки, из которых образуются мужские половые клетки, называют сперматогониями, а диплоидные соматические клетки, из которых образуются женские половые клетки — овогониями. Процесс образования (роста и дифференциации) мужских и женских половых клеток носит название гаметогенеза.
Гаметогенез основан на мейозе (от греч. meiosis — понижать), который представляет собой процесс редукционного деления ядер клеток, сопровождающегося понижением числа хромосом на ядро. Мейоз происходит в специализированных клетках репродуктивных органов живых существ, размножающихся половым путем (рис. 78). Например, у папоротникообразных мейоз встречается в специализированных клетках спорангий, располагающихся на нижней поверхности листьев этих растении и развивающихся в споры, а затем в га-метофиты. Последние существуют раздельно, продуцируя в конечном итоге мужские и женские гаметы. У цветковых растений мейоз осуществляется в специализированных клетках семяпочек, которые развиваются в споры.

Последние продуцируют гаметофит с одной яйцеклеткой.
 Кроме того у этих растений мейоз происходит также в специализированных клетках пыльников, которые также развиваются в споры, продуцирующие в конечном итоге пыльцу с двумя мужскими гаметами. У земляных червей, которые являются гермафродитами и содержат мужские половые органы в одном сегменте тела, а женские — в другом и которые характеризуются способностью к перекрестному оплодотворению между разными особями, имеется способность одновременно к сперматогенезу и овогенезу. Кроме того у этих растений мейоз происходит также в специализированных клетках пыльников, которые также развиваются в споры, продуцирующие в конечном итоге пыльцу с двумя мужскими гаметами. У земляных червей, которые являются гермафродитами и содержат мужские половые органы в одном сегменте тела, а женские — в другом и которые характеризуются способностью к перекрестному оплодотворению между разными особями, имеется способность одновременно к сперматогенезу и овогенезу.
У млекопитающих мейоз осуществляется в специализированных клетках семенников и яичников, в которых продуцируются мужские и женские гаметы соответственно. Выявлены белки — индукторы мейоза.
В процессе мейоза дипдоидное число хромосом (2п), которое характерно для соматических клеток (клеточных ядер) и незрелых зародышевых клеток, изменяется до гаплоидного числа (In), характерного для зрелых зародышевых клеток. Таким образом, в результате гаметогенеза половые клетки получают лишь половину хромосом соматических клеток (рис. 79).
Поведение хромосом в период гаметогенеза у животных является одинаковым как у мужских, так и у женских особей. Однако пол различается во времени происхождения разных стадий мейоза, что особенно заметно у человека. У мужских особей в постпубертатный период полный процесс мейоза завершается примерно в течение двух месяцев, в то время как у женских особей первое мейотическое деление начинается еще в фетальном яичнике и не завершается, пока не начнется овуляция, которая наступает у них примерно в пятнадцать лет.
У высших животных в случае мужских особей мейоз сопровождается образованием четырех функционально активных гамет (рис. 80). Напротив, у женских особей каждый овоцит II порядка дает лишь одну яйцеклетку. Другие ядерные продукты женского мейоза представляют собой три редукционных тельца, которые не участвуют в размножении и дегенерируют.
Мейоз состоит из двух делений клеточного ядра, которые называют мейотическими. Первое мейотическое деление ядра разделяет членов каждой пары гомологичных хромосом после того, как они спарились одна с другой (синапсис) и обменялись генетическим материалом (кроссинговер). В результате этого разделения образуется два гаплоидных ядра. Второе мейотическое деление разделяет две продольные половины хромосом (хроматиды) в каждом из этих ядер, продуцируя четыре гаплоидных ядра.
В процессе гаметогенеза происходит также дифференциация яйцеклеток (овогенез) и сперматозоидов (сперматогенез), являющаяся пререквизитом их функций. Будучи высокоспециализированными структурами, яйцеклетки животных намного крупнее сперматозоидов, обычно неподвижны и содержат питательный материал, который обеспечивает развитие эмбриона в начальном периоде после оплодотворения. Сперматозоиды большинства животных обладают жгутиком, обеспечивающим независимость их движения.
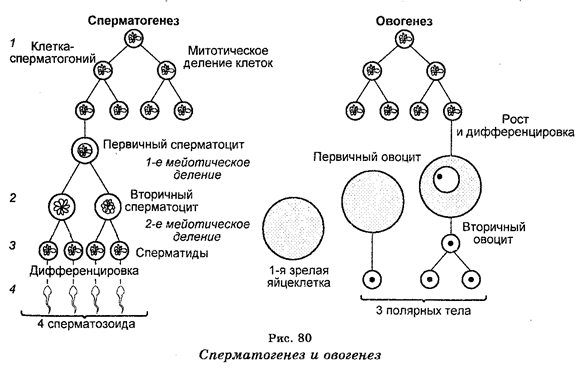
Мейоз имеет выдающееся биологическое значение. Благодаря мейозу в клетках организмов поддерживается постоянное число хромосом независимо от количества поколений. Следовательно, мейоз поддерживает постоянство видов. Наконец, в мейозе в результате кроссинговера происходит рекомбинация генов, которая является одним из факторов эволюции, хотя его значение является и меньшим по сравнению с мутагенезом.
Сперматогенез
— это процесс образования зрелых мужских половых клеток. Сперматозоиды развиваются в мужских половых железах (семенниках, или тестисах) из специализированных соматических клеток (рис. 81). Такими специализированными клетками служат так называемые примордиальные зародышевые клетки, которые мигрируют к тестисам в раннем периоде эмбриогенеза мужского индивидуума. Следовательно, примордиальные клетки являются прародителями (предшественниками) зрелых половых клеток. У млекопитающих после достижения половой зрелости сперматозоиды образуются практически всю жизнь.
Семенники человека состоят из многочисленных канальцев, стенки которых сформированы слоями клеток, которые находятся на разных стадиях развития сперматозоидов. Наружный слой канальцев образован крупными клетками, называемыми сдерма-тогониями. Эти клетки содержат диплоидный набор хромосом и являются в тестисах потомками примордиальных зародышевых клеток.
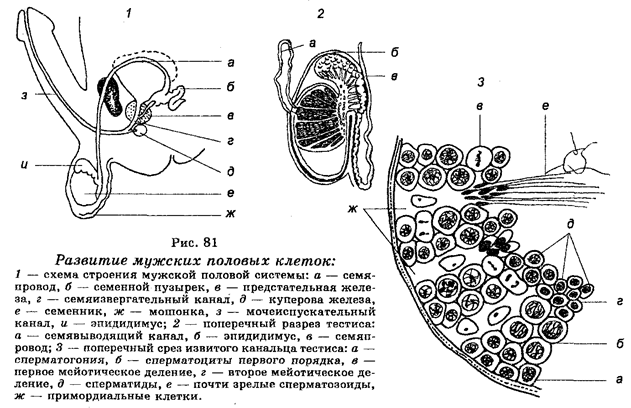
В период половой зрелости индивида часть сперматогоний перемещается во внутренний слой канальцев, где в результате мейоза они развиваются в клетки, получившие название сперматоцитов первого порядка (сперматоцитов I), затем в спер-матоциты второго порядка (сперматоциты II) и, наконец, в сперматиды, являющиеся гаплоидными зародышевыми клетками, дифференцирующимися в конечном итоге в зрелые сперматозоиды. Таким образом, в обобщенном виде можно сказать, что сперматогенез инициируется в диплоид-ных соматических клетках (сперматогони-ях), после чего следует период созревания зародышевых клеток, в котором происходит два деления ядер путем мейоза, приводящего к образованию сперматид.
 Мейоз в сперматогенезе протекает в несколько стадий (фаз). Между делениями имеются две интерфазы. Таким образом, мейотическое деление можно представить в виде серии следующих одно за другим событий, а именно: интерфаза I ® первое мейотическое деление (ранняя профаза I, поздняя профаза I, метафаза I, анафаза I, телофаза I) ® интерфаза II (интерокинез) ® второе мейотическое деление (профаза II, метафаза II, анафаза II, телофаза II). Процесс мейоза очень динамичен, поэтому микроскопические различия между разными стадиями отражают скорее не характер самих стадий, а скорее свойства хромосом на разных стадиях (рис. 82). Интерфаза I характеризуется тем, что в ней происходит репликация хромосом (удвоение ДНК), которая к началу ранней профазы I почти полностью завершается. Мейоз в сперматогенезе протекает в несколько стадий (фаз). Между делениями имеются две интерфазы. Таким образом, мейотическое деление можно представить в виде серии следующих одно за другим событий, а именно: интерфаза I ® первое мейотическое деление (ранняя профаза I, поздняя профаза I, метафаза I, анафаза I, телофаза I) ® интерфаза II (интерокинез) ® второе мейотическое деление (профаза II, метафаза II, анафаза II, телофаза II). Процесс мейоза очень динамичен, поэтому микроскопические различия между разными стадиями отражают скорее не характер самих стадий, а скорее свойства хромосом на разных стадиях (рис. 82). Интерфаза I характеризуется тем, что в ней происходит репликация хромосом (удвоение ДНК), которая к началу ранней профазы I почти полностью завершается.
Первое мейотическое деление начинается в первичном сперматоците и характеризуется длинной профазой, которая состоит из переходящих одна в другую профазы I и профазы II. В профазе I различают пять главных стадий — лептонему, зигонему, пахинему, диплонему и диакинез.
На стадии лептонемы хромосомы в ядре представлены в виде тонких спирализованных нитей, содержащих многочисленные темноок-рашенные гранулы (хромомеры). Расщепления хромомер и нитей не отмечают, но считают, что хромосомы на уровне этой стадии являются двойными, т. е. диплоидными. Гомологи каждой хромосомной пары объединяются хромомерами вдоль их длины по принципу застежки.
На стадии зигонемы характерно установление синапсов между гомологичными хромосомами, в результате чего образуются спаренные хромосомы (биваленты). Хромосомы Х и Y ведут себя по сравнению с аутосомами несколько по-другому. Они конденсируются в темноокрашиваемые гетерохроматиновые тела, спариваемые в результате наличия гомологичных районов на их концах.
На стадии пахинемы, которая является наиболее долгой во времени стадией в мейотической профазе, происходит конденсация бивалентов и разделение каждой хроматиды надвое, в результате чего каждый бивалент представляет собой сложную спиральную структуру, состоящую из четырех сестринских хроматид (тетрад). В конце этой стадии начинается разделение спаренных хромосом-бивалентов. Теперь гомологичные хромосомы могут наблюдаться рядом. Поэтому в некоторых препаратах можно видеть четыре хромосомы, которые образуются в результате дупликации каждого гомолога, формирующего сестринские хроматиды. На этой стадии происходят обмены между гомологами и формирование хиазм.
На стадии диплонемы происходит укорачивание, утолщение и взаимное отталкивание сестринских хроматид, в результате чего хроматиды в биваленте почти разъединены. Разделение является неполным по той причине, что в каждой паре хромосом еще не расщеплена центромера. Что касается бивалентов, то они удерживаются на различных местах вдоль их длины с помощью хиазм, которые являются структурами, сформированными между гомологичными хроматидами в результате предыдущего кроссинговера между синаптически связанными гомологами. В хороших препаратах можно наблюдать от одной до нескольких хиазм в зависимости от длины бивалента. Каждая наблюдаемая на этой стадии хиазма представляет собой результат обмена, который встречался между несестринскими хроматидами в течение стадии пахинемы. Поскольку сжатие и отталкивание бивалентов усиливается, хиазмы двигаются к концам хромосом, т. е. происходит терминализа-ция хромосом. В конце диплонемы наступает деспирализация хромосом; гомологи продолжают отталкиваться друг от друга.
На стадии диакинеза, которая сходна с диплотеной, продолжается укорочение бивалентов и наступает ослабление (уменьшение) хиазм, вследствие чего формируются дискретные единицы в виде хроматид (четырех). Непосредственно после завершения этой стадии происходит растворение ядерной мембраны.
В метафазе I биваленты достигают наибольшей конденсации. Становясь овальными, они располагаются в экваториальной части ядра, где формируют экваториальные пластинки мейотической метафазы I. Форма каждого бивалента определяется числом и локализацией хиазм. У мужчин число хиазм на бивалент в метафазе I составляет обычно 1—5. Бивалент XY становится палочковидным в результате одиночной терминально расположенной хиазмы.
В анафазе I начинается движение противоположных центромер к противоположным полюсам клетки. В результате этого происходит разделение гомологичных хромосом. Каждая хромосома состоит теперь из двух хроматид, удерживаемых центромерой, которая не делится и остается интактной. Этим анафаза I мейоза отличается от анафазы митоза, при котором центромера подвергается разделению. Важно заметить, что благодаря кроссинговеру каждая хро-матида является генетически различной.
В стадии телофазы I хромосомы достигают полюсов, чем заканчивается первое мейотическое деление. После телофазы I наступает короткая интерфаза (интеркинез), в которой хромосомы деспи-рализуются и становятся диффузными, или телофаза I переходит прямо в профазу II второго мейотического деления. Ни в одном, ни в другом случае репликации ДНК не отмечается. После первого мейотического деления клетки называют спермат&цитами II порядка. Количество хромосом в каждой такой клетке снижается от 2п до п, но содержание ДНК еще не изменяется.
Второе мейотическое деление осуществляется в течение нескольких фаз (профаза II, метафаза II, анафаза II, телофаза II) и сходно с митотическим делением. В профазе II хромосомы вторичных спер-матоцитов остаются у полюсов. В метафазе П центромера каждой из двойных хромосом делится, обеспечивая каждую новую хромосому собственной центромерой. В анафазе II начинается формирование веретена, к полюсу которого двигаются новые хромосомы. В телофазе II второе мейотическое деление заканчивается, в результате чего каждый сперматоцит II порядка дает два спермати-да, из которых дифференцируются затем сперматозоиды. Как и во вторичном сперматоците, число хромосом в сперматиде является гаплоидным (п). Однако хромосомы сперматид являются одиночными, тогда как хромосомы вторичных сперматоцитов II являются двойными, будучи построенными из двух хроматид. Следовательно, ядро каждого сперматида имеет одиночный набор негомологичных хромосом. Вторичное мейотическое деление является делением митотического типа (экваториальным делением). Оно разделяет двойные сестринские хроматиды и отличается от редукционного деления, в котором гомологичные хромосомы разделены. Единственное существенное отличие от классического митоза заключается в том, что здесь имеется гаплоидный набор хромосом.
 Итак, первое мейотическое деление сперматоцитов I порядка приводит к образованию двух вторичных сперматоцитов (II порядка). Обе хро-матиды структур, образующиеся в результате редукционного деления, являются сестринскими хроматида-ми. Последние возникают в результате репликации, предшествующей первому мейотическому делению. Второе мейотическое деление каждого вторичного сперматоцита приводит к образованию четырех сперма-тидов. Таким образом, в типичном мейозе клетки делятся дважды, тогда как хромосомы только один раз (рис. 83). Итак, первое мейотическое деление сперматоцитов I порядка приводит к образованию двух вторичных сперматоцитов (II порядка). Обе хро-матиды структур, образующиеся в результате редукционного деления, являются сестринскими хроматида-ми. Последние возникают в результате репликации, предшествующей первому мейотическому делению. Второе мейотическое деление каждого вторичного сперматоцита приводит к образованию четырех сперма-тидов. Таким образом, в типичном мейозе клетки делятся дважды, тогда как хромосомы только один раз (рис. 83).
Конечный этап в сперматогенезе связан с дифференциацией, которая заканчивается тем, что каждый из сравнительно больших, сферических неподвижных сперматидов превращается в небольшой вытянутый подвижный сперматозоид.
 У большинства взрослых (сексуально зрелых) мужских особей животных сперматогенез происходит в семенниках постоянно или периодически (сезонно). Например, у насекомых для завершения цикла сперматогенеза требуется лишь несколько дней, тогда как у млекопитающих этот цикл затягивается на недели и даже месяцы. У взрослого человека сперматогенез проходит в течение всего года. Время развития примитивных сперматогоний В зрелые сперматозоиды составляет около 74 дней. У большинства взрослых (сексуально зрелых) мужских особей животных сперматогенез происходит в семенниках постоянно или периодически (сезонно). Например, у насекомых для завершения цикла сперматогенеза требуется лишь несколько дней, тогда как у млекопитающих этот цикл затягивается на недели и даже месяцы. У взрослого человека сперматогенез проходит в течение всего года. Время развития примитивных сперматогоний В зрелые сперматозоиды составляет около 74 дней.
Мужские половые клетки, продуцируемые организмами разной видовой принадлежности, характеризуются подвижностью и чрезвычайным разнообразием в размерах и по структуре (рис. 84). Например, длина сперматозоидов D. melanogaster составляет 1,76 мм, что в 300 раз превышает длину сперматозоидов человека. Больше того, длина сперматозоидов D. bifurca составляет более 28 мм, что превышает длину самих насекомых этого вида в двадцать раз.
Каждый сперматозоид человека состоит из трех отделов — головки, средней части и хвоста (рис. 85). В головке сперматозоида располагается ядро. В нем содержится гаплоидный набор хромосом. Головка снабжена акросомой, которая содержит литические ферменты, необходимые сперматозоиду для вступления в яйцеклетку. В головке локализуется также две центриоли — прокси-мальная, которая побуждает деление оплодотворенной сперматозоидом яйцеклетки, и дистальная, которая дает начало аксимальному стержню хвоста. В средней части сперматозоида располагаются ба-зальное тельце хвоста и митохондрии. Хвост (отросток) сперматозоида образован внутренним аксимальным стержнем и внешним футляром, имеющим цитоплазматическое происхождение. Сперматозоиды человека характеризуются значительной подвижностью.
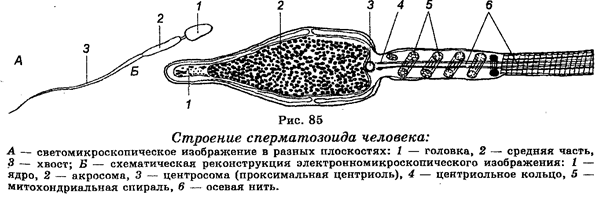
Овогенез — это процесс формирования яйцеклеток. Его функциями являются обеспечение гаплоидного набора хромосом в ядре яйцеклетки и обеспечение питательных потребностей зиготы. Овогенез в своем проявлении в основном сопоставим со сперматогенезом.
У млекопитающих и человека овогенез начинается еще во внутриутробном периоде (до рождения). Овогонии, представляющие собой мелкие клетки с довольно крупным ядром и локализованные в фолликулах яичников, начинают в фолликулах дифференцироваться в первичные овоциты. Последние формируются уже на третьем месяце внутриутробного развития, после чего они вступают в профазу первого мейотического деления. Ко времени рождения девочки все первичные овоциты уже находятся в профазе первого мейотического деления. Первичные овоциты остаются в профазе вплоть до наступления половой зрелости женского индивида. Когда при наступлении половой зрелости фолликулы яичника созревают, мейотическая профаза в первичных овоцитах возобновляется. Первое мейотическое деление для каждой развивающейся яйцеклетки завершается вскоре перед временем овуляции этой яйцеклетки. В результате первого мейотического деления и неравномерного распределения цитоплазмы одна образовавшаяся клетка становится вторичным овоцитом, другая — полярным (редукционным) тельцем.
Вторичное мейотическое деление у человека наступает, когда вторичный овоцит (развивающаяся яйцеклетка) проходит из яичника в фаллопиеву трубу. Однако это деление не завершается до тех пор, пока во вторичный овоцит не проникает ядерное содержимое сперматозоида, что происходит обычно в фаллопиевой трубе. Когда ядро сперматозоида проникает во вторичный овоцит, то последний делится, в результате чего образуется овотида (зрелая яйцеклетка) с пронуклеусом, содержащим одиночный набор из 23 материнских хромосом. У некоторых других видов образуются яйцеклетки, определяющие как мужской пол, так и женский. Важно подчеркнуть, что здесь происходит также расщепление и рекомбинация генов, основу которой создает расхождение хромосом. Другая клетка, образующаяся в результате второго мейотического деления у человека, является вторым полярным тельцем, не способным к дальнейшему развитию. В это время подвергается делению надвое и полярное (редукционное) тельце. Таким образом, развитие одного овоцита первого порядка сопровождается образованием одной овотиды и трех редукционных телец. В яичниках таким путем на протяжении жизни обычно созревает 300-400 ово-цитов, но в месяц созревает лишь один овоцит. В течение дифференциации яйцеклеток формируются мембраны, уменьшается в размере их ядро.
У некоторых видов животных овогенез протекает быстро и непрерывно и приводит к образованию большого количества яйцеклеток.
Вопреки сходству со сперматогенезом овогенез характеризуется некоторыми специфическими особенностями. Питательный материал (желток) первичного овоцита не распределяется поровну между четырьмя клетками, которые образуются в результате мей-отических делений. Основное количество желтка сохраняется в одной большой клетке, тогда как полярные тельца содержат очень мало этого вещества. Первые и вторые полярные тельца получают в результате делений те же самые хромосомные наборы, что и вторичные овоциты, но они не становятся половыми клетками. Поэтому яйцеклетки намного богаче питательным материалом по сравнению со сперматозоидами. Особенно сильно это различие проявляется в случае яйцекладущих животных.
Яйцеклетки млекопитающих имеют овальную или несколько вытянутую форму (рис. 86) и характеризуются типичными чертами клеточного строения. Они содержат все структуры, характер ные для соматических клеток, однако внутриклеточная организация яйцеклетки очень специфична и определяется тем, что яйцеклетка является также и средой, которая обеспечивает развитие зиготы. Одна из характерных особенностей яйцеклеток заключается в сложности строения их оболочек. У очень многих животных различают первичную, вторичную и третичную оболочки яйцеклеток. Первичная оболочка (внутренняя) формируется еще на стадии овоцита. Представляя собой поверхностный слой овоцита, она ные для соматических клеток, однако внутриклеточная организация яйцеклетки очень специфична и определяется тем, что яйцеклетка является также и средой, которая обеспечивает развитие зиготы. Одна из характерных особенностей яйцеклеток заключается в сложности строения их оболочек. У очень многих животных различают первичную, вторичную и третичную оболочки яйцеклеток. Первичная оболочка (внутренняя) формируется еще на стадии овоцита. Представляя собой поверхностный слой овоцита, она
имеет сложную структуру, т. к. пронизана выростами прилегающих к ней фолликулярных клеток. Вторичная (средняя) оболочка полностью сформирована фолликулярными клетками, а третичная (наружная) образована веществами, представляющими собой продукты секреции желез яйцеводов, через которые проходят яйцеклетки. У птиц, например, третичными оболочками яйцеклеток служат белковая, подскорлуповая и скорлуповая оболочки. Для яйцеклеток млекопитающих характерно наличие двух оболочек. Структура внутриклеточных компонентов яйцеклеток специфична в видовом отношении, а иногда имеет даже индивидуальные особенности.
Оплодотворение — это процесс объединения мужской и женской гамет, который приводит к формированию зиготы и последующему развитию нового организма. В процессе оплодотворения происходит установление диплоидного набора хромосом в зиготе, что определяет выдающееся биологическое значение этого процесса.
В зависимости от видовой принадлежности организмов у животных, размножающихся половым путем, различают наружное и внутреннее оплодотворение.
Наружное оплодотворение происходит в окружающей среде, в которую поступают мужские и женские половые клетки. Например, наружным является оплодотворение у рыб. Выделяемые ими мужские (молока) и женские (икра) половые клетки поступают в воду, где и происходит их «встреча» и объединение. Данные об оплодотворении у морских ежей свидетельствуют о том, что уже через 2 секунды после соприкосновения сперматозоидов и яйцеклетки наступают изменения в электрических свойствах плазменной мембраны яйцеклетки. Слияние содержимого гамет наступает через 7 секунд.
Внутреннее
оплодотворение обеспечивается переносом сперматозоидов из мужского организма в женский в результате полового акта. Такое оплодотворение встречается у млекопитающих, причем центральным моментом здесь является исход встречи между половыми клетками. Считают, что в яйцеклетку этих животных проникает ядерное содержимое лишь одного сперматозоида. Что касается цитоплазмы сперматозоида, то у одних животных она поступает в яйцеклетку в небольшом количестве, у других совсем не поступает в яйцеклетку.
У человека оплодотворение происходит в верхней части фаллопиевой трубы, причем в оплодотворении, как и у других млекопитающих, участвует лишь один сперматозоид, ядерное содержимое которого поступает в яйцеклетку. Иногда в фаллопиевой трубе может оказаться не одна, а две или более яйцеклеток, в результате чего возможно рождение двоен, троен и т. д. Например, в XVIII в. зарегистрирован случай рождения в России одной матерью (женой крестьянина Федора Васильева) 16 двоен, 7 троен и 4 четверней (всего 69 детей).
В результате оплодотворения в оплодотворенной яйцеклетке восстанавливается диплоидный набор хромосом. Яйцеклетки способны к оплодотворению в течение примерно 24 часов после овуляции, тогда как оплодотворяющая способность сперматозоидов сохраняется до 48 часов.
В механизмах оплодотворения многое еще остается неясным. Предполагают, что проникновение в яйцеклетку ядерного материала лишь одного из множества сперматозоидов связано с изменениями электрических свойств плазматической мембраны яйцеклетки. По вопросу о причинах активации сперматозоидом метаболизма яйцеклетки существует две гипотезы. Одни исследователи считают, что связывание сперматозоида с внешними рецепторами на поверхности клеток представляет собой сигнал, который через мембрану поступает внутрь яйцеклетки и активирует там инозитолтрифосфат и ионы кальция. Другие полагают, что сперматозоиды содержат специальный инициирующий фактор.
Оплодотворенная яйцеклетка дает начало зиготе, развитие организмов через образование зигот называют зигогенезом. Экспериментальные разработки, выполненные в последние годы, показали, что оплодотворение яйцеклеток млекопитающих, включая человека, возможно и в пробирке, после чего зародыши, развившиеся в пробирке, могут быть имплантированы в матку женщины, где они могут развиваться дальше. К настоящему времени известны многочисленные случаи рождения «пробирочных» детей (см. раздел VI). Установлено также, что оплодотворить яйцеклетку человека способны не только сперматозоиды, но и сперматиды. Наконец, возможно оплодотворение яйцеклеток (лишенных искусственно ядер) млекопитающих ядрами их соматических клеток (см. § 35).
В отличие от зигогенеза, многие животные организмы способны к размножению в естественных условиях путем партеногенеза (от греч. parthenos — девственница и genesis — рождение). Различают облигатный и факультативный партеногенез. Облигатный партеногенез — это размножение организмов из неоплодотворенной яйцеклетки. Такой партеногенез служит способом размножения животных более 90 видов, включая некоторых позвоночных. Примером облигатного партеногенеза является размножение кавказской скальной породы ящерицы, представленной только женскими особями. Напротив, факультативный партеногенез заключается в том, что яйцеклетки способны развиваться как без оплодотворения, так и после оплодотворения. Факультативный партеногенез в свою очередь бывает женским и мужским. Женский партеногенез част у пчел, муравьев, коловраток, у которых из неоплодотворенных яйцеклеток развиваются самцы. Мужской партеногенез встречается у некоторых изогамных водорослей.
У растений также известны случаи, когда зародыш развивается из не оплодотворенной яйцеклетки. Как отмечено выше, это явление получило название апомикса. Оно очень широко встречается у многих покрытосеменных растений, в т. ч. у культивируемых, таких, как свекла, хлопчатник, лен, табак и другие.
Наряду с естественным партеногенезом различают искусственный (индуцированный) партеногенез, который можно вызвать раздражением яйцеклеток с помощью физических или химических факторов, что ведет к активации яйцеклеток и, как следствие, к развитию неоплодотворенных яиц. Искусственный партеногенез наблюдали в случае животных, принадлежащих ко многим систематическим группам — иглокожим, червям, моллюскам и даже некоторым млекопитающим.
Известна форма партеногенеза, получившая название андроге-неза (от греч. andros — мужчина, genesis — рождение). Если в яйцеклетке инактивировать ядро и если после этого в нее проникнет несколько сперматозоидов, то из такой яйцеклетки в результате слияния мужских (сперматозоидных) ядер развивается мужской организм. Широко известны эксперименты В. Л. Астаурова (1904-1974), который показал андрогенез на тутовом шелкопряде. Эти опыты заключались в следующем. В яйцеклетках шелкопряда одного вида (Bombyx mandarina) с помощью высокой температуры инактивировали ядра, а затем такие яйцеклетки оплодотворяли сперматозоидами шелкопряда другого вида (В. mori). Проникнув в яйцеклетки, последние сливались между собой, что давало начало новым организмам, которые по своим свойствам оказались отцовскими организмами (В. mori). Скрещивания этих организмов с самками В. mori давало потомство, принадлежащее к В. mori.
Роль партеногенеза и его форм в природе невелика, т. к. он не обеспечивает широких адаптивных возможностей организмов. Однако его использование имеет практическое значение. В частности, Б. Л. Астауровым был разработан способ получения партеноге-нетического потомства у тутового шелкопряда, что широко используется в промышленном производстве шелка.
В отличие от зигогенеза и партеногенеза существует гиногенез (от греч. gyne — женщина), который представляет собой псевдогамию, заключающуюся в том, что сперматозоид встречается с яйцеклеткой и активирует ее, но ядро сперматозоида не сливается с ядром яйцеклетки. В этом случае позволяющее потомство состоит только из женских особей. У отдельных видов круглых червей, рыб и земноводных гиногенез служит нормальной формой размножения, давая потомство, состоящее только из самок. Гиногенез можно вызвать и искусственно с помощью факторов, способных разрушать клеточные ядра (радиации, температуры и др.). В частности, описаны случаи искусственного гиногенеза у тутового шелкопряда, у некоторых видов рыб и амфибий. Получение таких форм может иметь некоторое практическое значение в случае хозяйственно полезных видов.
Как было отмечено выше, оплодотворение у цветковых (покрытосеменных) имеет существенную отличительную особенность в виде двойного оплодотворения (С. Г. Навашин, 1896), которое сводится к тому, что в зародышевом мешке гаплоидная яйцеклетка и дипло-идная центральная клетка оплодотворяются спермиями, в результате чего образуется диплоидный зародыш и триплоидная клетка, развивающаяся в клетки эндосперма (см. гл. II).
Партеногенез, андрогенез и гиногенез являются формами нарушения полового размножения. Предполагают, что эти формы возникли в ходе эволюции в результате частных эволюционных приспособлений.
Организмам, размножающимся только половым путем, характерно чередование гаплоидной и диплоидной фаз в их развитии. У многих организмов, включая млекопитающих, это чередование имеет регулярный характер, и на нем основано сохранение видовых признаков организмов. Диплоидия способствует накоплению разных аллелей. Напротив, для организмов, которые могут размножаться как половым, так и бесполым путем, характерно чере дование (смена) поколений, когда одно или несколько бесполых поколений организмов сменяется поколением организмов, размножающихся половым путем. дование (смена) поколений, когда одно или несколько бесполых поколений организмов сменяется поколением организмов, размножающихся половым путем.
Различают первичное и вторичное чередование поколений. Первичное чередование поколений отмечается у организмов, развивших в ходе эволюции половой прогресс, но сохранивших способность к бесполому размножению, и заключается в регулярном чередовании полового и бесполого поколений (рис. 87). Оно встречается у животных (простейших), у водорослей и у всех высших растений. У простейших классическим примером первичного чередования поколений является бесполое размножение малярийного плазмодия в организме человека (шизогония) и половое — в организме малярийного комара. У растений половое поколение представлено гаметофитом, бесполое — спорофитом. Механизм первичного чередования заключается в том, что на растениях спорофитного поколения развиваются споры, которые на основе мейоза дают гаплоидные мужские и женские гаметофиты. На последних развиваются спермии и яйцеклетки. Оплодотворение яйцеклетки дает начало диплоидному спорофиту. Таким образом, клетки гаметофита содержат гаплоидный набор хромосом, а спорофита — диплоидный набор, т. е. у растений чередование поколений связано со сменой гаплоидного и диплоидного состояний.
Если проследить за соотношением между спорофитом и гаметофитом у растений разного уровня организации, то можно увидеть, что в ходе эволюции развитию подвергался спорофит, тогда как для гаметофиты была характерной редукция. Например, у мхов преобладающим является гаметофит (гаплоидное поколение), на котором живет спорофит. Но уже у папоротникообразных преобладающим является спорофит (диплоидное поколение) в виде хорошо развитого растения со стеблями и корнями, а гаметофит представлен слоем клеток, которые образуют пластину, прикрепляющуюся к почве с помощью ризоидов. Далее, у голосеменных гаметофит уменьшается до небольших количеств клеток, а у покрытосеменных мужской гематофит представлен лишь двумя клетками, женский — семью, тогда как спорофитом у голосеменных являются деревья (сосна, ель и другие), а покрытосеменных — деревья, кустарники, травы.
Между гаметофитом и спорофитом могут быть как сходства по морфологии и продолжительности жизни, так и различия по этим признакам. В первом случае это называют изоморфным чередованием поколений, во втором — гетероморфным.
Вторичное чередование поколений широко встречается у животных. Оно отмечается в формах гетерогонии и метагенеза. Гетерогония заключается в первичном чередовании полового процесса и партеногенеза. Например, у трематод половое размножение регулярно сменяется партеногенезом. У многих других организмов гетерогония зависит от сезона. Так, коловратки, дафнии и тли осенью размножаются путем зигогенеза (путем оплодотворения яйцеклеток и образования зигот), а летом — путем партеногенеза. Метагенез заключается в чередовании полового размножения и вегетативного (бесполового). Например, гидры размножаются обычно почкованием, но при понижении температуры образуют половые клетки. У кишечнополостных на некоторых стадиях развития происходит переход от полового размножения к вегетативному. У некоторых морских кишечнополостных полипоидное поколение правильно чередуется с медузоидным. Для полипоидного поколения характерно размножение так называемой стробиляцией (поперечными перетяжками), для медузоидного — половым путем (оплодотворение яиц, образование личинок и развитие полипов).
Для мужских и женских особей животных характерны различия в специфических фенотипических чертах (размеры, строение тела, окраска и другие свойства), а также в поведении. Различия между самками и самцами по их свойствам называют половым диморфизмом. У животных он встречается уже на низших ступенях эволюционного развития, например у круглых гельминтов, членистоногих, и достигает наибольшего выражения у позвоночных животных, у которых внешние различия между самцами и самками очень выразительны. У растений тех видов, для которых характерно наличие мужских и женских особей, также имеет место половой диморфизм, однако выражен он очень незначительно.
Если у животных мужские и женские половые клетки продуцируются одной и той же особью, имеющей как мужские, так и женские половые железы, то это явление носит название гермафродитизма. Термин «гермафродитизм» является сочетанием греческих имен Гермеса (бог мужской красоты) и Афродиты (богини женской красоты). Различают истинный и ложный гермафродитизм. Истинный гермафродитизм чаще всего встречается у организмов, находящихся на низких уровнях эволюции, например у плоских и кольчатых червей, а также у моллюсков. У плоских червей мужские и женские половые железы функционируют на протяжении всей жизни особи. Напротив, у моллюсков половые железы продуцируют яйцеклетки и сперматозоиды попеременно. Однако явление истинного гермафродитизма встречается и у более организованных существ. В частности, оно встречается у млекопитающих. Например, у свиней иногда отмечается развитие яичников на одной стороне тела, а развитие тестисов на другой, либо происходит развитие комбинированных структур (овотестисов), причем в обоих случаях имеет место синтеза функционально активных яйцеклеток и сперматозоидов. Таких животных относят к «промежуточному» полу, причем большинство особей промежуточного сексуального типа является особями женского пола с двумя ХХ-хромосомами. Некоторые из них характеризуются агрессивным поведением, а это свидетельствует о том, что хотя их тектикулярная ткань не содержит зародышевых клеток, секреция тестостерона, влияющая на поведение, все же имеет место. Аналогичное явление отмечено у коз.
Истинный гермафродитизм встречается и у человека, возникая в результате нарушений развития. Генотипами гермафродитов являются 46ХХ или 46XY, причем большинство случаев относится к XX (около 60%). Генотипы XX чаще всего встречаются у гермафродитов негроидных африканских популяций, тогда как генотипы XY чаще среди японцев. У гермафродитов обоих типов отмечена тенденция в сторону билатеральной ассиметрии гонад. Среди истинных гермафродитов встречаются также хромосомные мозаики, у которых соматические клетки содержат пару хромосом XX, другие — пару хромосом XY.
Известен и ложный гермафродитизм, когда индивидуумы обладают наружными половыми органами и вторичными половыми признаками, характерными для обоих полов, но продуцируют половые клетки лишь одного типа — мужские или женские.
Для большинства цветковых растений характерны гермафро-дитные цветки, которые обычно называют обоеполыми, поскольку в каждом цветке имеется пестик и тычинки. По этой причине плоды развиваются со всех цветков. Обоеполыми являются пшеница, вишня, яблоня и многие другие виды растений. Помимо обоеполых в ходе эволюции развились растения с разделением полов в пределах одного и того же вида, т. е. возникли однодомность и двудомность растений. Растения, содержащие одновременно пестичные (женские) и тычиночные (мужские) цветки получили название однодомных. У однодомных растений плоды развиваются только из пестичных цветков. Однодомными являются кукуруза, огурец, тыква и другие. Напротив, двудомными являются растения, содержащие либо пестичные, либо тычиночные цветки (в пределах одного и того же вида). У двудомных растений плодоносящими бывают только те, которые имеют пестичные цветки (женские особи). Двудомными являются тополь, клубника и другие виды древесных и травянистых растений.
Гермафродитизм у человека представляет одно из патологических состояний. Что касается растений, то знания об их гермафродитизме чрезвычайно важны для практики сельского хозяйства.
Онтогенез (от греч. ontos — существо, genesis — развитие) — это полная история (цикл) развития индивидуального организма (животного или растения), начинающаяся с образования давших ему начало половых клеток и заканчивающаяся его смертью. Представления об онтогенезе (индивидуальной истории развития организма) основаны на данных о росте организма, дифференцировке его клеток и морфогенезе. Следовательно, онтогенез есть категория индивидуальная.
В противоположность онтогенезу видовой категорией является филогенез (от греч. phyle — племя, genesis — развитие) под которым со времен Э. Геккеля, впервые обосновавшего этот термин, понимают историю возникновения и развития вида (животных или растений). Между онтогенезом и филогенезом существует тесная связь, которая отражена в так называемом биогенетическом законе (Э. Геккель, Ф. Мюллер), который, как показали исследования, в принципе справедлив. Поскольку онтогенез индивидуума определяется определенными чертами филогенетического развития вида, к которому принадлежит данный индивидуум, то можно сказать, что онтогенез является основой филогенеза, с одной стороны, и результатом филогенеза — с другой.
Изучение фундаментальных основ онтогенеза имеет важное значение для понимания биологии и эволюции организмов. Однако, чтобы лучше узнать современное состояние учения об онтогенезе, рассмотрим вначале, как понимали рост и развитие организма в прошлые времена на примере организма человека.
Первые представления о росте и развитии восходят ко временам античного мира. Еще Гиппократ (460-377 гг. до н. э.) предполагал, что яйцеклетки уже содержат полностью сформированный организм, но в очень уменьшенном виде. Это представление затем нашло продолжение в учении о преформизме (от лат. preformatio — предобразование), которое особенно популярным оказалось в XVII—XVIII вв. Сторонниками преформизма были Гарвей, Мальпиги и многие другие видные биологи и медики того времени. Для преформистов спорный вопрос заключался лишь в том, в каких половых клетках преформирован организм — женских или мужских. Тех, кто отдавал предпочтение яйцеклеткам, называли овистами, а тех, кто большое значение придавал мужским половым клеткам, называли анималькулистами. Преформизм — это метафизическое учение от начала до конца, ибо оно отрицало развитие. Решающий удар преформизму нанес Ш. Бонне (1720-1793), который открыл в 1745 г. партеногенез на примере развития тлей из неоплодотворенных яиц. После этого преформизм уже не мог оправиться и стал терять свое значение.
В античном мире возникло и другое учение, противоположное преформизму и получившее впоследствии название эпигенеза (от греч. epi — после, genesis — развитие). Как и преформизм, эпигенез большое распространение получил также в XVII—XVIII вв. В распространении эпигенеза большое значение имели взгляды К. Ф. Вольфа (1733—1794), обобщенные в его книге «Теория развития» (1759). К. Ф. Вольф считал, что в яйце нет ни преформи-рованного организма, ни его частей, и что яйцо состоит из перво-ночально однородной массы. В отличие от преформистов взгляды К- Ф. Вольфа и других сторонников эпигенеза для своего времени были прогрессивны, т. к. содержали мысль о развитии. Однако в дальнейшем появились новые моменты. В частности, в 1828 г. К. Бэр опубликовал свой труд «История развития животных», в котором показал, что содержимое яйца не однородно, т. е. структурировано, причем степень структурированности возрастает по мере развития зародыша. Таким образом, К. Бэр показал несостоятельность как преформизма, так и эпигенеза.
В наше время рост организма понимают в качестве постепенного увеличения его массы в результате увеличения количества клеток. Рост можно измерить, построив на основе результатов измерений кривые размеров организма, массы, сухой массы, количества клеток, содержания азота и других показателей. Что же касается дифференциации клеток, то это процесс, благодаря которому одни клетки становятся морфологически, биохимически и функционально отличными от других клеток. Размножение и дифференцировка одних клеток всегда координированы с ростом и дифференцировкой других. Оба эти процесса происходят на протяжении всего жизненного цикла организма. Поскольку дифференцирующиеся клетки изменяют свою форму, а в изменения формы вовлекаются группы клеток, то это сопровождается мор-фогенезом, представляющим собой совокупность процессов, определяющих структурную организацию клеток и тканей, а также общую морфологию организмов. Таким образом, рост является результатом количественных изменений в виде увеличения количества клеток (массы тела) и качественных — в виде дифференци-ровки клеток и морфогенеза.
Понятия о росте организмов (размножении клеток), дифферен-цировке клеток и о морфогенезе позволяют сформулировать заключение о развитии как основополагающей особенности онтогенеза.
Развитие — это качественные изменения организмов, которые определяются дифференцировкой клеток и морфогенезом, а также биохимическими изменениями в клетках и тканях, обеспечивающими в ходе онтогенеза прогрессивные изменения индивидов. В рамках современных представлений развитие организма понимают в качестве процесса, при котором структуры, образовавшиеся ранее, побуждают развитие последующих структур. Процесс развития детерминирован генетически и теснейшим образом связан со средой. Следовательно, развитие определяется единством внутренних и внешних факторов. Онтогенез в зависимости от характера развития организмов типируют на прямой и непрямой, в связи с чем различают прямое и непрямое развитие.
Прямое развитие организмов в природе встречается в виде неличиночного и внутриутробного развития, тогда как непрямое развитие наблюдается в форме личиночного развития.
Под личиночным развитием понимают непрямое развитие, поскольку организмы в своем развитии имеют одну или несколько личиночных стадий. Личиночное развитие широко распространено в природе и характерно для насекомых, иглокожих, амфибий. Личинки этих животных ведут самостоятельный образ жизни, подвергаясь затем превращениям. Поэтому это развитие называют еще развитием с метаморфозами (см. ниже).
Неличиночное развитие характерно для организмов, развивающихся прямым образом, например для рыб, пресмыкающихся и птиц, яйца которых богаты желтком (питательным материалом). Благодаря этому в яйцах, откладываемых во внешнюю среду, проходит значительная часть онтогенеза, метаболизм зародышей обеспечивается развивающимися провизорными органами, представляющими собой зародышевые оболочки (желточный мешок, амнион, аллантоис).
Внутриутробное развитие также характерно для организмов, развивающихся прямым путем, например для млекопитающих, включая человека. Поскольку яйцеклетки этих организмов очень бедны питательными веществами, то все жизненные функции зародышей обеспечиваются материнским организмом посредством образованных из тканей матери и зародыша провизорных органов, среди которых главным является плацента. Эволюционно внутриутробное развитие является самой поздней формой, однако оно наиболее выгодно для зародышей, т. к. наиболее эффективно обеспечивает их выживание.
Онтогенез подразделяют на проэмбриональный, эмбриональный и постэмбриональный периоды. В случае человека, а иногда и высших животных, период развития до рождения часто называют пре-натальным или антенатальным, после рождения — постнатальным. В пределах пренатального периода выделяют начальный (первая неделя развития), зародышевый и плодный периоды. Развивающийся зародыш до образования зачатков органов называют эмбрионом, после образования зачатков органов — плодом.
Проэмбриональный (от греч. pro — до, embryon — зародыш) период в индивидуальном развитии организмов связан с образованием половых клеток в процессе гаметогенеза. Как отмечено выше, мужские половые клетки животных по своей структуре не имеют существенных отличий от других (соматических) клеток, тогда как яйцеклетки характеризуются важной отличительной чертой, заключающейся в том, что они содержат очень много желтка. Учитывая количество желтка и топографию его в яйцеклетках, последние классифицируют на три типа, а именно:
1. Изолецитальные клетки. Эти яйцеклетки содержат немного желтка, который локализован равномерно по всей клетке. Изолецитальные яйцеклетки продуцируются иглокожими (морскими ежами), низшими хордовыми (ланцетниками), млекопитающими.
2. Телолецитальные яйцеклетки. Эти яйцеклетки содержат большое количество желтка, который сосредоточен на одном из полюсов — вегетативном. Такие яйцеклетки продуцируются моллюсками, земноводными, рептилиями, птицами. Например, яйцеклетки лягушки состоят из желтка на 50%, яйцеклетки кур (в обиходе куриные яйца) — на 95% . На другом полюсе (анимальном) телоле-ЦИтальных яйцеклеток сосредоточены цитоплазма и ядро.
3. Центролецитальные яйцеклетки. В этих яйцеклетках желтка немного, но он занимает центральное положение. Периферию таких яйцеклеток занимает цитоплазма. Примером центролециталь-ных яйцеклеток являются яйцеклетки, продуцируемые членистоногими.
Для проэмбрионального периода характерно также то, что в этот период в гаметах происходят метаболические процессы, связанные с накоплением интенсивно синтезируемых молекул РНК.
Эмбриональный период или эмбриогенез (от греч. embryon — за-родыш, genesis — развитие), начинается со слияния ядер мужской и Женской половых клеток, который представляет собой процесс оплодотворения яйцеклеток. У организмов, для которых характерно внутриутробное развитие, эмбриональный период заканчивается рождением потомства, а у организмов, для которых характерны личиночный и неличиночный типы развития, эмбриональный период завершается выходом потомства из яйцевых или зародышевых оболочек соответственно.
В пределах эмбрионального периода различают стадии зиготы, дробления, бластулы, гаструлы, образования зародышевых листков, гистогенез и органогенез. Как отмечено выше, с учетом фактора времени у млекопитающих и человека зародыш до момента формирования зачатков органов называют эмбрионом, а после этого вплоть до рождения называют плодом. У человека развитие эмбриона (зародыша) заканчивается к концу второго месяца. Начиная с 9-й недели, следует плодный период, характеризующийся дальнейшим ростом и развитием организма (плода) во внутриутробном состоянии вплоть до рождения.
Зигота
. У млекопитающих зигота образуется в результате оплодотворения, начинающегося с того, что одна из мужских половых клеток достигает яйцеклетки и инициирует ее развитие. В активированной мужской половой клеткой яйцеклетке происходит ряд физических и химических процессов, включая перемещение протоплазмы, что ведет к установлению билатеральной симметрии яйцеклетки, а также перестройку плазматической мембраны, что исключает слияние с яйцеклеткой других (дополнительных) мужских половых клеток. Затем следует слияние плазматических мембран яйцеклетки и спермия с последующим разрушением ядерных мембран, что обеспечивает слияние ядер двух клеток. Ядра клеток сливаются, при этом восстанавливается диплоидный набор хромосом. Оплодотворение яйцеклетки сопровождается активированием в ней синтеза белков. Таким образом, образуется по существу одноклеточный организм.
Дробление
. Образование морулы. Дробление представляет собой начальный период развития зиготы (оплодотворенного яйца). Поскольку яйцеклетки обладают центриолями, то оно заключается в делении зиготы путем митоза, которое начинается, например у человека, через 30 часов после осеменения. У человека деление начинается с движения оплодотворенной яйцеклетки по фаллопиевой трубе и заключается в появлении на поверхности яйцеклетки борозды. Первая борозда приводит к образованию двух клеток — двух бластомеров, вторая — четырех бластомеров, третья — восьми бластомеров и т. д. (рис. 88). Группа клеток, образовавшаяся в результате последовательных дроблений зиготы, получила название морулы (от греч. morum — тутовая ягода).
Стадию морулы проходят все многоклеточные животные, размножающиеся половым путем. В зависимости от видовой принад лежности деление идет по-разному. Различают радиальное (позвоночные, иглокожие), билатеральное (грибневики, некоторые хордовые) и спиральное дробление (немертины, кольчатые черви, многие моллюски), причем эти формы дробления зависят от плоскостей дробления. Поэтому их морулы состоят из разного количества клеток. Кроме того, из части клеток образуется структура, называемая трофобластом, клетки которого питают зародыш, а благодаря ферментам обеспечивают также внедрение последнего в стенку матки. У человека прикрепление морулы к стенке матки происходит на 7-й день после оплодотворения. Позднее клетки трофобласта отслаиваются от зародыша и образуют пузырек, который заполняется жидкостью тканей матки. лежности деление идет по-разному. Различают радиальное (позвоночные, иглокожие), билатеральное (грибневики, некоторые хордовые) и спиральное дробление (немертины, кольчатые черви, многие моллюски), причем эти формы дробления зависят от плоскостей дробления. Поэтому их морулы состоят из разного количества клеток. Кроме того, из части клеток образуется структура, называемая трофобластом, клетки которого питают зародыш, а благодаря ферментам обеспечивают также внедрение последнего в стенку матки. У человека прикрепление морулы к стенке матки происходит на 7-й день после оплодотворения. Позднее клетки трофобласта отслаиваются от зародыша и образуют пузырек, который заполняется жидкостью тканей матки.
Характерная особенность дробления заключается в том, что при нем значительного роста клеток не происходит. Поэтому биологическое значение этой стадии заключается в том, что из крупной клетки, которой является яйцеклетка, образуются более мелкие клетки, в которых уменьшено отношение цитоплазмы к ядру. В результате этого происходит изменение топологии цитоплазматических комплексов в бластомерах, что создает новое цитоплазматическое окружение для ядер.
Дробление зиготы завершается образованием многоклеточной структуры, получившей название бластулы (от греч. blastos — росток). Эта структура имеет форму пузырька, состоящего из одного слоя клеток, называемого бластодермой. Теперь эти клетки называют эмбриональными. По размерам бластула сходна с яйцеклеткой. В период дробления увеличивается количество ядер, возрастает общее количество ДНК. В конце стадии бластулы синтезируется также небольшое количество мРНК и тРНК, но новые рибосомы и рибосомная РНК до начала гаструляции еще не обнаруживаются, либо если обнаруживаются, то в ничтожных количествах.
Гаструляция. Гаструляция (от греч. gastre — полость сосуда) — это следующий за образованием бластулы процесс движения эмбриональных клеток, который сопровождается формированием двух или трех (в зависимости от вида животных) слоев зародыша или так называемых зародышевых листков (рис. 89).
Гаструляция характеризуется увеличением интенсивности обмена по сравнению с дроблением в 2-3 раза. Резко возрастает синтез мРНК, рРНК, рибосом и белков.
Развитие (гаструляция) изолецитальных яиц происходит путем инвагинации (впячивания) вегетативного полюса внутрь бластулы, в результате чего противоположные полюса почти сливаются, а бла-стоцель (полость бластулы) почти исчезает либо полностью исчезает. Внешний слой клеток зародыша получил название эктодермы (от греч. ectos — снаружи, derma — кожа) или наружного зародышевого листка, тогда как внутренний — энтодермы (от греч. entos — внутри) или внутреннего зародышевого листка. Образующаяся при этом полость получила название гастроцели, или первичной кишки, вход в которую называют бластопором (первичным ртом).
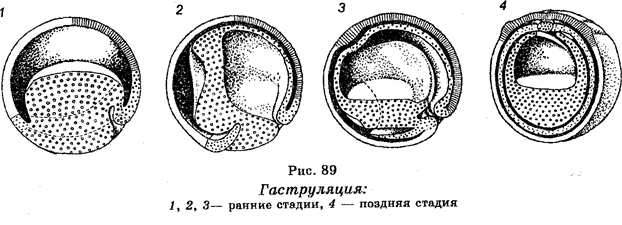
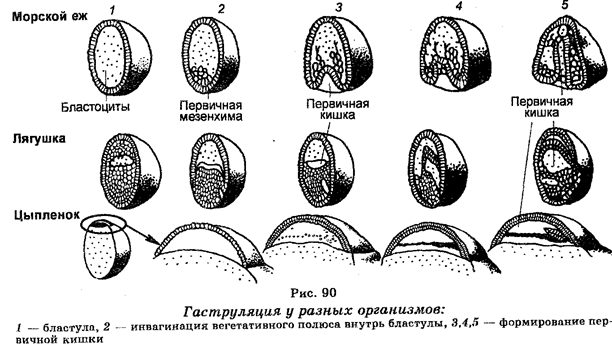
Развитие двух зародышевых листков характерно для губок и кишечнополостных. Однако для хордовых в период гаструляции характерно развитие третьего зародышевого листка — мезодермы (от греч. mesos — средний), образующегося между эктодермой и энтодермой (рис. 90).
Гаструляция является необходимым пререквизитом для последующих стадий развития, поскольку она приводит клетки в положение, открывающее возможность формировать органы. Дифференцированный на три эмбриональных закладки зародышевый материал дает начало всем тканям и органам развивающегося зародыша.
Развитие (дифференцировка) зародышевых листков в ходе эмбриогенеза сопровождается тем, что из них формируются различные ткани и органы. В частности, из эктодермы развиваются эпидермис кожи, ногти и волосы, сальные и потовые железы, нервная система (головной мозг, спинной мозг, ганглии, нервы), рецептор-ные клетки органов чувств, хрусталик глаза, эпителий рта, носовой полости и анального отверстия, зубная эмаль. Из энтодермы развиваются эпителий пищевода, желудка, кишек, желчного пузыря, трахеи, бронхов, легких, мочеиспускательного канала, а также печень, поджелудочная железа, щитовидная, паращитовидная и зобная железы. Из мезодермы развиваются гладкая мускулатура, скелетные и сердечные мышцы, дерма, соединительная ткань, кости и хрящи, дентин зубов, кровь и кровеносные сосуды, брыжейка, почки, семенники и яичники. У человека первыми обособляются головной и спинной мозг. Через 26 дней после овуляции длина человеческого зародыша составляет около 3,5 мм. При этом уже видны зачатки рук, но зачатки ног только вступают в развитие. Через 30 дней после овуляции длина зародыша равна уже 7,5 мм. В это время уже можно различить сегментацию зачатков конечностей, глазные бокалы, полушария головного мозга, печень, Желчный пузырь и даже разделение сердца на камеры.
У восьминедельного зародыша человека при длине его около 40 мм и весе около 5 г появляются почти все структуры тела. Органогенез заканчивается к концу эмбрионального периода. В это время эмбрион по внешнему виду приобретает черты сходства с человеком.
Длина 12 недельного человеческого плода составляет уже около 87 мм, а масса около 45 г. Продолжается дальнейший рост и развитие плода. Например, на 4-м месяце развития появляются волосы, а на 20-й неделе начинают образовываться клетки крови.
Если дефинитивное ротовое отверстие образуется на месте первичного рта (бластопора), то этих животных называют первичноро-тыми (черви, моллюски, членистоногие). Если же дефинитивный рот образуется в противоположном месте, то этих животных называют вторичноротыми (иглокожие, хордовые).
Для обеспечения связи зародыша со средой у него развиваются так называемые провизорные органы, которые существуют временно. В зависимости от типа яйцеклеток провизорными органами являются разные структуры. У рыб, рептилий и птиц провизорным органом является желточный мешок. У млекопитающих желточный мешок закладывается в начале эмбриогенеза, но не развивается. Позднее он редуцируется. В ходе эволюции у рептилий, птиц и млекопитающих развились зародышевые оболочки, обеспечивающие защиту и питание эмбрионов (рис. 91). У млекопитающих, в т. ч. у человека, эти зародышевые оболочки являются листками ткани, развивающимися из тела эмбриона. Существуют три такие оболочки — амнион, хорион и аллантоис. Наружная оболочка эмбриона называется хорионом. Она врастает в матку. Место наибольшего врастания в матку называется плацентой. Зародыш с плацентой связан через пуповину или пупочный канатик, в котором имеются кровеносные сосуды, обеспечивающие плацентарное кровообращение. Амнион развивается из внутреннего листка, а аллантоис развивается между амнионом и хорионом. В пространстве между эмбрионом и амнионом, которое называется амниоти-ческой полостью, содержится жидкость (амниотическая). В этой жидкости находится эмбрион, а затем и плод до самого рождения. Метаболизм плода обеспечивается через плаценту.
В основе формообразующего взаимодействия частей эмбриона лежат определенным образом скоординированные процессы обмена веществ. Закономерностью развития является гетерохронность, под которой понимают разное во времени образование закладок органов и разную интенсивность их развития. Быстрее развиваются те органы и системы, которые должны раньше начать функционировать. Например, у человека зачатки верхних конечностей развиваются быстрее, чем зачатки нижних.
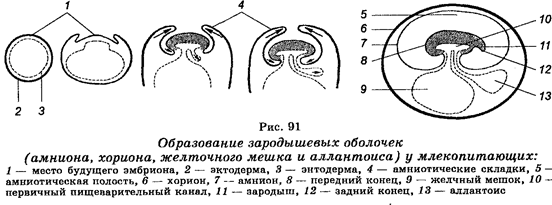
Большое влияние на развитие зародыша и плода оказывают условия жизни матери. Зародыш чрезвычайно чувствителен к разным воздействиям. Поэтому различают так называемые критические периоды, т. е. периоды, в которых зародыши, а потом и плоды наиболее чувствительны к повреждающим факторам. В случае человека критическими периодами эмбрионального онтогенеза являются первые дни после оплодотворения, время образования плаценты и роды, а повреждающими факторами являются алкоголь, токсические вещества, недостаток кислорода, вирусы, бактерии, патогенные простейшие, гельминты и другие факторы. Эти факторы обладают терратогенным действием и ведут к уродствам, нарушениям нормального развития.
 Еще со времен Гиппократа (V в. д. н. э.) обсуждается вопрос о причинах, которые инициируют рождение плода. В частности, сам Гиппократ предполагал, что развитие плода инициирует собственное рождение. Новейшие экспериментальные работы английских исследователей, выполненные на овцах, показали, что у овец инициация окотов контролируется комплексом гипоталамус + гипофиз + надпочечники плодов. Повреждения ядер гипоталамуса, удаление передней доли гипофиза или надпочечников пролонгирует беременности овец. Напротив, введение овцам аденокортикотроп-ного гормона (секрета передней доли гипофиза) или кортизола (секрета надпочечников) сокращает сроки их беременностей. Еще со времен Гиппократа (V в. д. н. э.) обсуждается вопрос о причинах, которые инициируют рождение плода. В частности, сам Гиппократ предполагал, что развитие плода инициирует собственное рождение. Новейшие экспериментальные работы английских исследователей, выполненные на овцах, показали, что у овец инициация окотов контролируется комплексом гипоталамус + гипофиз + надпочечники плодов. Повреждения ядер гипоталамуса, удаление передней доли гипофиза или надпочечников пролонгирует беременности овец. Напротив, введение овцам аденокортикотроп-ного гормона (секрета передней доли гипофиза) или кортизола (секрета надпочечников) сокращает сроки их беременностей.
Довольно частым нарушением развития является разделение зародыша на очень ранней стадии развития, что сопровождается развитием однояйцевых (моно-зиготных) близнецов (рис. 92). Известны также и так называемые сиамские близнецы, представляющие собой неразделенные организмы. Неразделенность встречается разной — от незначительного соединения до почти полного слияния двух организмов с разделенными головами или ногами. Иногда из двух сиамских близнецов один нормален, но другой чрезвычайно изменен, будучи прикрепленным к нормальному, являясь, по существу, паразитом.
 Итак, в процессе развития высших эукариотов одиночная оплодотворенная клетка-зигота в ходе дальнейшего развития в результате митоза дает начало клеткам разных типов — эпителиальным, нервным, костным, клеткам крови и другим, которые характеризуются разнообразием морфологии и макромолекулярного состава. Однако для клеток разных типов характерно и то, что они содержат одинаковые наборы генов, но являются высокоспециализированными, выполняя лишь одну или несколько специфических функций, т. е. одни гены «работают» в клетках, другие неактивны. Например, только эритроциты специфичны в синтезе и хранении гемоглобина. Итак, в процессе развития высших эукариотов одиночная оплодотворенная клетка-зигота в ходе дальнейшего развития в результате митоза дает начало клеткам разных типов — эпителиальным, нервным, костным, клеткам крови и другим, которые характеризуются разнообразием морфологии и макромолекулярного состава. Однако для клеток разных типов характерно и то, что они содержат одинаковые наборы генов, но являются высокоспециализированными, выполняя лишь одну или несколько специфических функций, т. е. одни гены «работают» в клетках, другие неактивны. Например, только эритроциты специфичны в синтезе и хранении гемоглобина.
Точно так лишь клетки эпидермиса синтезируют кератин. Поэтому давно возникли вопросы о генетической идентичности ядер соматических клеток и о контрольных механизмах развития оплодотворенных яйцеклеток как пререквизита в познании механизмов, лежащих в основе дифференцировки клеток.
Начиная с 50-х годов во многих лабораториях были выполнены эксперименты по успешной пересадке ядер соматических клеток в яйцеклетки, искусственно лишенные собственных ядер. Исследование ДНК из ядер разных дифференцированных клеток показало, что почти во всех случаях геномы содержат одинаковые наборы последовательностей нуклеотидных пар. Известны лишь исключения, когда эритроциты млекопитающих теряют свои ядра в течение последней стадии дифференцировки. Но к этому времени пулы стойких гемоглобиновых мРНК уже синтезированы, так что ядра больше не нужны эритроцитам. Другими примерами служат гены иммуноглобулинов и Т-клеток, модифицируемые в ходе развития.
Одним из крупных этапов в направлении познания контрольных механизмов эмбрионального онтогенеза стали результаты экспериментов, выполненных в 1960—70 гг. английским исследователем Д. Гёрдоном с целью выяснить, обладают ли ядра соматических клеток способностью обеспечивать дальнейшее развитие яйцеклеток, если эти ядра ввести в яйцеклетки, из которых предварительно удалены собственные ядра. На рис. 93 приведена схема одного из этих экспериментов, в котором ядра соматических клеток головастика пересаживали в яйцеклетки лягушки с предварительно удаленными ядрами. Эти эксперименты показали, что ядра соматических клеток действительно способны обеспечивать дальнейшее развитие яйцеклеток, т. к. они оказались способными оплодотворять яйцеклетки и «заставляли» их развиваться дальше. Этим была показана возможность клонирования животных.
Позднее другими исследователями были выполнены эксперименты, в которых было показано, что перенос отдельных бластомеров из 8- и 16-дневных эмбрионов овец одной породы в безъядерную половину яйцеклетки (после рассечения последней пополам) другой породы сопровождалось формированием жизнеспособных эмбрионов с последующим рождением ягнят.
В начале 1997 г. английскими авторами было показано, что введение в искусственно лишенные ядра яйцеклеток овец ядер соматических клеток (клеток эмбрионов, плодов и вымени взрослых овец), а затем имплантация оплодотворенных таким образом яйцеклеток в матку овец сопровождается возникновением беременности с последующим рождением ягнят.
Оценка этих результатов показывает, что млекопитающих можно размножать асексуальным путем, получая потомство животных, клетки которых содержат ядерный материал отцовского или материнского происхождения в зависимости от пола овцы-донора, в таких клетках лишь цитоплазма и митохондрии имеют материнское происхождение. Это заключение имеет чрезвычайно важное общебиологическое значение, расширяет наши взгляды на потенциал размножения животных. Но важно также добавить, что речь идет о генетических манипуляциях, которые в природе отсутствуют. С другой стороны, в практическом плане эти генетические манипуляции представляют собой прямой путь клонирования высокоорганизованных животных с заданными свойствами, что имеет важное экономическое значение (см. раздел VI). В медицинском плане этот путь, возможно, будет использован в будущем для преодоления мужского бесплодия.
Итак, генетическая информация, необходимая для нормального развития эмбриона, не теряется в течение дифференцировки клеток. Другими словами, соматические клетки обладают свойством, получившим название тотипотентности, т. е. в их геноме содержится вся информация, полученная ими от оплодотворенной яйцеклетки, давшей им начало в результате дифференциации. Наличие этих данных с несомненностью означает, что дифференциация клеток подвержена генетическому контролю.
Установлено, что интенсивный белковый синтез, следующий за оплодотворением, у большинства эукариотов не сопровождается синтезом мРНК. Изучение овогенеза у позвоночных, в частности У амфибий, показало, что интенсивная транскрипция происходит еще в течение профазы I (особенно диплотены) мейоза. Поэтому генные транскрипты в форме молекул мРНК или про-мРНК сохраняются в яйцеклетках в бездействующем состоянии. Установлено, что у эмбриональных клеток имеет место так называемое ассимет-ричное деление, заключающееся в том, что деление эмбриональной клетки дает начало двум клеткам, из которых лишь одна наследует белки, принимающие участие в транскрипции. Таким образом, неравное распределение транскрипционных факторов между дочерними клетками ведет к экспрессии в них разных наборов генов после деления, т. е. к дифференциации клеток.
У амфибий и, возможно, у большинства позвоночных, генетические программы, контролирующие раннее развитие (до стадии бластулы), устанавливается еще в течение овогенеза. Более поздние стадии развития, когда начинается клеточная дифференциация (примерно со стадии гаструлы) нуждаются в новых программах для экспрессии генов. Таким образом, дифференцировка клеток связана с перепрограммированием генетической информации в том или ином направлении.
Характерная особенность дифференцировки клеток заключается в том, что она необратимо ведет к тому или иному типу клеток. Этот процесс носит название детерминации и также находится под генетическим контролем, а как сейчас предполагают, дифференциация и детерминация клеток регулируется взаимодействием клеток на основе сигналов, осуществляемых пептидными ростовыми факторами через тирозинкиназные рецепторы. Вероятно, существует много таких систем. Одна из них заключается в том, что дифференциация мышечных и нервных клеток регулируется нейрорегу-линами, представляющими собой мембранные белки, действующие через один или более тирозинкиназных рецепторов.
Генетический контроль детерминации
демонстрируется также существованием так называемых гомейотропных или гомеозисных мутаций, которые, как показано у насекомых, вызывают изменения при детерминации в специфических имагинальных дисках. В результате некоторые части тела развиваются не на своих местах. Например, у дрозофил мутации трансформируют детерминацию антенного диска в диск, который развивается в аппендикс конечности, протянутой от головы. У насекомых из рода Ophthalmoptera структуры крыльев могут развиваться из диска для глаз. У мышей показано существование генного кластера (комплекса) Нох, который состоит из 38 генов и контролирует развитие конечностей.
Самостоятельное значение имеет вопрос о регуляции активности генов в период эмбрионального развития. Считают, что в ходе дифференцировки гены действуют в разное время, что выражается в транскрипции в разных дифференцированных клетках разных мРНК, т. е. имеет место репрессия и дерепрессия генов. Например, количество генов, транскрибируемых в РНК в бластоцитах морского ежа, равно 10% , в клетках печени крыс — тоже 10% , а в клетках тимуса крупного рогатого скота — 15%. Предполагают, что в контроле транскрипционного статуса генов принимают участие негистоновые белки. В пользу этого предположения свидетельствуют следующие данные. Когда хроматин клеток в фазе транскрибируется в системе in vitro, то синтезируется только гистоновая мРНК, а вслед за нею и гистоны. Напротив, когда используют хроматин клеток из О^-фазы, то никакой гистоновой мРНК не синтезируется. Когда же негистоновые белки удаляются из хроматина G1
-фазы и замещаются негис-тоновыми хромосомными белками, синтезированными в фазе S, то после транскрипции такого хроматина in vitro синтезируется гистоновая мРНК. Больше того, когда негистоновые белки происходят из G^-фазы клеток, а ДНК и гистоны из S-фазы клеток, никакой гистоновой мРНК не синтезируется. Эти результаты показывают, что содержащиеся в хроматине негистоновые белки определяют возможность транскрипции генов, кодирующих гистоны. Поэтому считают, что негистоновые хромосомные белки могут играть важную роль в регуляции и экспрессии генов у эукариот.
Имеющиеся данные позволяют считать, что в регуляции транскрипции у животных принимают участие белковые и стероидные гормоны. Белковый (инсулин) и стероидные (эстрогон и тестосте-рон) гормоны представляют собой две сигнальные системы, используемые в межклеточных коммуникациях. У высших животных гормоны синтезируются в специализированных секреторных клетках. Освобождаясь в кровяное русло, они поступают в ткани, поскольку молекулы белковых гормонов имеют относительно крупные размеры, то они не проникают в клетки. Поэтому их эффекты обеспечиваются белками-рецепторами, локализованными в мембранах клеток-мишеней, и внутриклеточными уровнями циклического АМФ (цАМФ). Напротив, стероидные гормоны являются малыми молекулами, вследствие чего легко проникают в клетки через мембраны. Оказавшись внутри клеток, они связываются со специфическими рецепторными белками, которые имеются в цитоплазме только клеток-мишеней. Как считают, комплексы гормон + белковый рецептор, концентрируясь в ядрах клеток-мишеней, активирует транскрипцию специфических генов через взаимодействие с определенными негистоновыми белками, которые связываются с промоторными районами специфических генов. Следовательно, связывание комплекса гормон + белок (белковый рецептор) с негистоновыми белками освобождает промоторные районы для движения РНК-полимеразы. Обобщая данные о генетическом контроле эмбрионального периода в онтогенезе организмов, можно заключить, что его ход контролируется дифференциальным включением и выключением действия генов в разных клетках (тканях) путем их дерепрессии и репрессии.
После появления организма на свет начинается его постэмбриональное развитие (постнатальное для человека), которое у разных организмов протекает от нескольких дней до сотен лет в зависимости от их видовой принадлежности. Следовательно, продолжительность жизни — это видовой признак организмов, не зависящий от уровня их организации (см. ниже).
В постэмбриональном онтогенезе различают ювениальный и пу-бертатный периоды, а также период старости, заканчивающийся смертью.
Ювенильный период
. Этот период (от лат. juvenilis — юный) определяется временем от рождения организма до полового созревания. У разных организмов он протекает по-разному и зависит от типа онтогенеза организмов. Для этого периода характерно либо прямое, либо непрямое развитие.
В случае организмов, для которых характерно прямое развитие (многие беспозвоночные, рыбы, пресмыкающиеся, птицы, млекопитающие, человек), вылупившиеся из яйцевых оболочек или новорожденные сходны со взрослыми формами, отличаясь от последних лишь меньшими размерами, а также недоразвитием отдельных органов и несовершенными пропорциями тела (рис. 94).

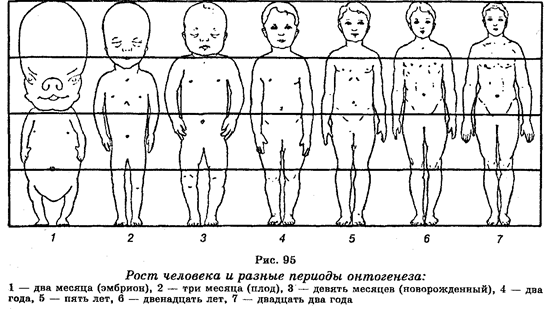
Характерной особенностью роста в ювенильный период организмов, подверженных прямому развитию, является то, что происходит увеличение количества и размеров клеток, изменяются пропорции тела. Рост человека в разные периоды его онтогенеза показан на рис. 95. Рост разных органов человека неравномерен. Например, рост головы заканчивается в детстве, ноги достигают пропорциональной величины примерно к 10 годам. Наружные половые органы очень быстро растут в возрасте 12—14 лет. Различают определенный и неопределенный рост. Определенный рост характерен для организмов, которые к определенному возрасту прекращают свой рост, например, насекомые, млекопитающие, человек. Неопределенный рост характерен для организмов, которые растут всю жизнь, например, моллюски, рыбы, земноводные, рептилии, многие виды растений.
В случае непрямого развития организмы претерпевают превращения, называемые метаморфозами (от лат. metamorphosis — превращение). Они представляют собой видоизменения организмов в процессе развития. Метаморфозы широко встречаются у кишечно-полостных (гидры, медузы, коралловые полипы), плоских червей (фасциолы), круглых червей (аскариды), моллюсков (устрицы, мидии, осьминоги), членистоногих (раки, речные крабы, омары, креветки, скорпионы, пауки, клещи, насекомые) и даже у некоторых хордовых (оболочечники и земноводные). При этом различают полные и неполные метаморфозы. Наиболее выразительные формы метаморфозов наблюдают у насекомых, которые подвергаются как неполным, так и полным метаморфозам.
 Неполное превращение — это такое развитие, при котором из яйцевых оболочек выходит организм, строение которого сходно со строением взрослого организма, но размеры намного меньше. Такой организм называют личинкой. В процессе роста и развития размеры личинок увеличиваются, но имеющийся хитипизированный покров мешает дальнейшему увеличению размеров тела, что приводит к линьке, т. е. сбрасыванию хитинизированного покрова, под которым находится мягкая кутикула. Последняя расправляется, и это сопровождается увеличением размеров животного. После нескольких линек животное достигает зрелости. Неполное превращение характерно, например, в случае развития клопов (рис. 96). Неполное превращение — это такое развитие, при котором из яйцевых оболочек выходит организм, строение которого сходно со строением взрослого организма, но размеры намного меньше. Такой организм называют личинкой. В процессе роста и развития размеры личинок увеличиваются, но имеющийся хитипизированный покров мешает дальнейшему увеличению размеров тела, что приводит к линьке, т. е. сбрасыванию хитинизированного покрова, под которым находится мягкая кутикула. Последняя расправляется, и это сопровождается увеличением размеров животного. После нескольких линек животное достигает зрелости. Неполное превращение характерно, например, в случае развития клопов (рис. 96).
Полное превращение — это такое развитие, при котором из яйцевых оболочек освобождается личинка, существенно отличающаяся по строению от взрослых особей. Например, у бабочек и многих насекомых личинками являются гусеницы. Гусеницы подвержены линьке, причем могут линять по нескольку раз, превращаясь затем в куколки. Из последних развиваются взрослые формы (имаго), которые не отличаются от исходных (рис. 97).
У позвоночных метаморфозы встречаются среди земноводных и костных рыб. Для личиночной стадии характерно наличие провизорных органов, которые либо повторяют признаки предков, либо имеют явно приспособительное значение. Например, для головастика, являющегося личиночной формой лягушки и повторяющего признаки исходной формы, характерны рыбообразная форма, наличие жаберного ды хания, одного круга кровообращения. Приспособительными признаками головастиков являются их присоски, длинный кишечник. Для личиночных форм характерно также и то, что по сравнению со взрослыми формами, они оказываются приспособленными к жизни в совершенно иных условиях, занимая другую экологическую нишу и другое место в цепи питания. Например, личинки лягушек имеют жаберное дыхание, тогда как взрослые формы — легочное. В отличие от взрослых форм, которые являются плотоядными существами, личинки лягушек питаются растительной пищей. хания, одного круга кровообращения. Приспособительными признаками головастиков являются их присоски, длинный кишечник. Для личиночных форм характерно также и то, что по сравнению со взрослыми формами, они оказываются приспособленными к жизни в совершенно иных условиях, занимая другую экологическую нишу и другое место в цепи питания. Например, личинки лягушек имеют жаберное дыхание, тогда как взрослые формы — легочное. В отличие от взрослых форм, которые являются плотоядными существами, личинки лягушек питаются растительной пищей.
Последовательность событий в развитии организмов часто называют жизненными циклами, которые могут быть простыми и сложными. Наиболее простые циклы развития характерны, например, для млекопитающих, когда из оплодотворенной яйцеклетки развивается организм, который снова продуцирует яйцеклетки и т. д. Сложными биологическими циклами являются циклы у животных, для которых характерно развитие с метаморфозами. Знания о биологических циклах имеют практическое значение, особенно в случаях, когда организмы являются возбудителями или переносчиками возбудителей болезней животных и растений.
Развитие и дифференциация, связанные с метаморфозами, являются результатом естественного отбора, благодаря которому многие личиночные формы, например, гусеницы насекомых и головастики лягушек адаптированы к среде лучше, чем взрослые половозрелые формы.
Пубертатный период
. Этот период называют еще зрелым, и он связан с половой зрелостью организмов. Развитие организмов в этот период достигает максимума.
На рост и развитие в постэмбриональный период большое влияние оказывают факторы среды. Для растений решающими факторами являются свет, влажность, температура, количество и качество питательных веществ в почве. Для животных первостепенное значение имеет полноценное кормление (наличие в корме белков, углеводов, липидов, минеральных солей, витаминов, микроэлементов). Важны также кислород, температура, свет (синтез витамина Д).
Рост и индивидуальное развитие животных организмов подвержены нейрогуморальной регуляции со стороны гуморальных и нервных механизмов регуляции. У растений обнаружены гормонопо-добные активные вещества, получившие название фитогормонов. Последние влияют на жизненно важные отправления растительных организмов.
 В клетках животных в процессе жизнедеятельности синтезируются химически активные вещества, влияющие на процессы жизнедеятельности. Нервные клетки беспозвоночных и позвоночных вырабатывают вещества, получившие название нейросекретов. Железы эндокринной, или внутренней, секреции также выделяют вещества, которые получили название гормонов. Эндокринные железы, в частности, те, которые имеют отношение к росту и развитию, регулируются нейросекретами. У членистоногих регуляция роста и развития очень хорошо показана на примере влияния гормонов на линьку. Синтез личиночного секрета клетками регулируется гормонами, накапливающимися в мозге. В особой железе у ракообразных вырабатывается гормон, тормозящий линьку. Уровни этих гормонов определяют периодичность линек. У насекомых установлена гормональная регуляция созревания яиц, протекание диапаузы. В клетках животных в процессе жизнедеятельности синтезируются химически активные вещества, влияющие на процессы жизнедеятельности. Нервные клетки беспозвоночных и позвоночных вырабатывают вещества, получившие название нейросекретов. Железы эндокринной, или внутренней, секреции также выделяют вещества, которые получили название гормонов. Эндокринные железы, в частности, те, которые имеют отношение к росту и развитию, регулируются нейросекретами. У членистоногих регуляция роста и развития очень хорошо показана на примере влияния гормонов на линьку. Синтез личиночного секрета клетками регулируется гормонами, накапливающимися в мозге. В особой железе у ракообразных вырабатывается гормон, тормозящий линьку. Уровни этих гормонов определяют периодичность линек. У насекомых установлена гормональная регуляция созревания яиц, протекание диапаузы.
У позвоночных железами внутренней секреции являются гипофиз, эпифиз, щитовидная, паращитовидная, поджелудочная, надпочечники и половые железы, которые тесно связаны одна с другой (рис. 98). Гипофиз у позвоночных вырабатывает гонадотропный гормон, стимулирующий деятельность половых желез. У человека гормон гипофиза влияет на рост. При недостатке развивается карликовость, при избытке — гигантизм. Эпифиз продуцирует гормон, который влияет на
сезонные колебания в половой активности животных. Гормон щитовидной железы влияет на метаморфоз насекомых и земноводных. У млекопитающих недоразвитие щитовидной железы ведет к задержке роста, недоразвитию половых органов. У человека из-за дефекта щитовидной железы задерживается окостенение, рост (карликовость), не наступает полового созревания, останавливается психическое развитие (кретинизм). Надпочечники продуцируют гормоны, оказывающие влияние на метаболизм, рост и дифференци-ровку клеток. Половые железы продуцируют половые гормоны, которые определяют вторичные половые признаки. Удаление половых желез ведет к необратимым изменениям ряда признаков. Например, у кастрированных петухов прекращается рост гребня, теряется половой инстинкт. Кастрированный мужчина преобрета-ет внешнее сходство с женщиной (не растет борода и волосы на коже, отлагается жир на груди и в области таза, сохраняется тембр голоса и т. д.).
Фитогормонами растений являются ауксины, цитокинины и гиб-береллины. Они регулируют рост и деление клеток, образование новых корней, развитие цветков и другие свойства у растений.
На всех периодах онтогенеза организмы способны к восстановлению утраченных или поврежденных частей тела. Это свойство организмов носит название регенерации, которая бывает физиологической и репаративной.
Физиологическая регенерация
— это замена утерянных частей тела в процессе жизнедеятельности организма. Регенерации этого типа очень распространены в животном мире. Например, у членистоногих она представлена линькой, которая связана с ростом. У рептилий регенерация выражается в замещении хвоста и чешуи, у птиц — перьев, когтей и шпор. У млекопитающих примером физиологической регенерации может быть ежегодное сбрасывание оленями рогов.
Репаративная регенерация
— это восстановление части тела организма, отторгнутой насильственным путем. Регенерация этого типа возможна у многих животных, но ее проявления различны. Например, она часта у гидр и связана с размножением последних, поскольку из части регенерирует весь организм. У других организмов регенерации проявляются в виде способности отдельных органов к восстановлению после утраты ими какой-либо части. У человека достаточно высокой регенеративной способностью обладают эпителиальная, соединительная, мышечная и костная ткани.
Растения многих видов также способны к регенерации. Данные о регенерации имеют большое значение не только в биологии. Их широко используют в сельском хозяйстве, в медицине, в частности, в хирургии.
Старость как этап онтогенеза. Старость является предпоследним этапом онтогенеза животных, причем ее длительность определяется общей продолжительностью жизни, которая является видовым признаком и которая у разных животных является разной. Наиболее точно старость изучена у человека.
Известны самые различные определения старости человека. В частности, одно из наиболее популярных определений заключается в том, что старость есть накопление последовательных изменений, сопровождающих повышение возраста организма и увеличивающих вероятность его болезней или смерти. Науку о старости человека называют геронтологией (от греч. geron — старик, старец, logos — наука). Ее задачей является изучение закономерностей возрастного перехода между зрелостью и смертью.
Научные исследования в геронтологии распространяются на разные области, начиная с исследований изменения активности клеточных ферментов и заканчивая выяснением влияния психологических и социологических смягчений в стрессах среды на поведение старых людей.
В случае человека различают физиологическую старость, старость, связанную с календарным возрастом, и преждевременное старение, обусловленное социальными факторами и болезнями. В соответствии с рекомендациями ВОЗ пожилым возрастом человека следует считать возраст порядка 60-75 лет, а старым в 75 лет и более.
Старость человека характеризуется рядом внешних и внутренних признаков.
Среди внешних признаков старости наиболее заметными являются снижение плавности движений, изменение осанки, снижение эластичности кожи, массы тела, упругости и эластичности мышц, появление на лице и других участках тела морщин, выпадение зубов. Так, например, по обобщенным данным человек в возрасте 30 лет теряет 2 зуба (в результате выпадения), в 40 лет — 4 зуба, в 50 лет — 8 зубов, а в 60 лет — уже 11 зубов. Заметным изменениям подвергается первая сигнальная система (притупляется острота органов чувств). Например, максимальное расстояние, при котором здоровые люди различают те или иные одинаковые звуки, в 20-30 лет составляет 12 м, в 50 лет — 10 м, в 60 лет — 7 м, а в 70 лет — только 4 м. Заметно изменяется также вторая сигнальная система (изменяется речевая интонация, голос становится глухим).
Среди внутренних признаков в первую очередь следует назвать такие признаки, как обратное развитие (инволюция) органов. Отмечается уменьшение размеров печени и почек, уменьшается количество нефронов в почках (к 80-ти годам почти наполовину), что снижает функциональные возможности почек и отражается на водно-элекролитном обмене. Снижается эластичность кровеносных сосудов, уменьшается перфузия кровью тканей и органов, повышается периферическая сопротивляемость сосудов. В костях накапливаются неорганические соли, изменяются (обызвествляются) хрящи, снижается способность органов к регенерации. Происходят существенные изменения в клетках, замедляется деление и восстановление их функционального тонуса, уменьшается содержание воды, снижается активность клеточных ферментов, нарушается координированность между ассимиляцией и диссимиляцией. В головном мозге нарушается синтез белков, в результате чего образуются аномальные белки. Повышается вязкость клеточных мембран, нарушается синтез и утилизация половых гормонов, происходят изменения в структуре нейронов. Наступают структурные изменения белков соединительной ткани и изменения эластичности этой ткани. Ослабляются иммунологические реакции, увеличивается возможность аутоим-мунных реакций. Снижаются функции эндокринных систем, в частности, половых желез. Поведение других признаков в старости показано на рис. 99.
 Стремления понять природу старения организма возникли давно. В Древней Греции Гиппократ считал, что старение связано с неумеренностью в пище, недостаточным пребыванием на свежем воздухе. Аристотель считал, что старение связано с расходом тепловой энергии организмом. Значение пищи как фактора старения отмечал также Гален. Но долгое время для объективного понимания этой проблемы не хватало научных данных. Лишь в XIX в. в изучении старения наметился некоторый прогресс, стали формулировать теории старения. Стремления понять природу старения организма возникли давно. В Древней Греции Гиппократ считал, что старение связано с неумеренностью в пище, недостаточным пребыванием на свежем воздухе. Аристотель считал, что старение связано с расходом тепловой энергии организмом. Значение пищи как фактора старения отмечал также Гален. Но долгое время для объективного понимания этой проблемы не хватало научных данных. Лишь в XIX в. в изучении старения наметился некоторый прогресс, стали формулировать теории старения.
Одной из первых наиболее известных теорий старения организма человека является теория немецкого врача Х. Гуфеланда (1762-1836), который отмечал в долголетии значение трудовой деятельности. До нас дошло его высказывание о том, что ни один лентяй не дожил до преклонного возраста. Еще более известной является эндокринная теория старения, которая берет начало от опытов, выполненных еще в середине прошлого века Бертольдом (1849), который показал, что пересадка семенников от одних животных к другим сопровождается развитием вторичных половых признаков. Позднее французский физиолог Ш. Броун Секар (1818-1894) на основе результатов впрыскиваний себе экстрактов из семенников утверждал, что эти инъекции производят благотворное и омолаживающее действие. В начале XX в. уже сложилось убеждение в том, что наступление старости связано с угасанием деятельности желез внутренней секреции, в частности, половых желез. В 20-30-е гг. на основе этого убеждения в разных странах было проделано много операций по омолаживанию пожилых или старых людей. Например, Г. Штейнах в Австрии перевязывал у мужчин семенные канатики, что вело к прекращению внешней секреции половых желез и, якобы, к некоторому омоложению. С. А. Воронов во Франции пересаживал семенники от молодых животных к старым и от обезьян к мужчинам, а Тушнов в СССР омолаживал петухов, вводя им гистолизаты половых желез. Все эти операции приводили к некоторым эффектам, но лишь временным. После названных воздействий процессы старения продолжались, причем еще интенсивнее.
В начале нашего века возникла микробиологическая теория старения, творцом которой был И. И. Мечников, который различал физиологическую старость и патологическую. Он считал, что старость человека является патологической, т. е. преждевременной. Основу представлений И. И. Мечникова составляло учение об ортобиозе (Orthos — правильный, bios — жизнь), в соответствии с которым основной причиной старения является повреждение нервных клеток продуктами интоксикации, образующимися в результате гниения в толстом кишечнике. Развивая учение о нормальном образе жизни (соблюдение правил гигиены, регулярный труд, воздержание от вредных привычек), И. И. Мечников предлагал также способ подавления гнилостных бактерий кишечника путем употребления кисломолочных продуктов.
В 30-е гг. широкое распространение получила теория о роли центральной нервной системы в старении. Творцом этой теории является И. П. Павлов, который установил интегрирующую роль центральной нервной системы в нормальном функционировании организмов. Последователи И. П. Павлова в экспериментах на животных показали, что преждевременное старение вызывается нервными потрясениями и продолжительным нервным перенапряжением.
Заслуживает упоминания теория возрастных изменений соединительной ткани, сформулированная в те годы А. А. Богомольцем (1881—1946). Он считал, что физиологическую активность организма обеспечивает соединительная ткань (костная ткань, хрящи, сухожилия, связки и волокнистая соединительная ткань) и что изменения коллоидного состояния клеток, потеря их тургора и т. д. определяют возрастные изменения организмов. Современные данные указывают на значение накопления кальция в соединительной ткани, т. к. он способствует потере ее упругости, а также уплотнению сосудов.
Для современных подходов к познанию сущности и механизмов старения характерно широкое использование данных физико-химической биологии и, в частности, достижений молекулярной генетики. Наиболее распространенные современные представления о механизмах старения сводятся к тому, что в процессе жизни в клетках организма накапливаются соматические мутации, в результате которых происходит синтез дефектных белков или нере-парируемые сшивки ДНК с белком. Поскольку дефектные белки играют дезинтегрирующую роль в клеточном метаболизме, то это ведет к старению. В случае культивируемых фибробластов показано, что связанные со старыми клетками белки и мРНК подавляют синтез ДНК в молодых фибробластах.
Известна также гипотеза, в соответствии с которой старение является результатом изменения митохондриальных метаболитов с последующим нарушением функций ферментов.
У человека показано существование генов, определяющих сроки развития наследственных дегенеративных процессов, связанных со старением. Ряд исследователей считает, что причиной старения являются изменения в системе иммунологической защиты организма, в частности, аутоиммунные реакции на структуры организма, имеющие жизненное значение. Наконец, в объяснениях механизмов старения специалисты большое внимание уделяют повреждениям белков, связанным с образованием свободных радикалов. Наконец, иногда придают значение освобождающимся после распада лизосом гидролазам, которые разрушают клетки.
Однако исчерпывающей теории старения все же еще не создано, поскольку ясно, что ни одна из этих теорий самостоятельно объяснить механизмы старения не может.
Смерть. Смерть является завершающим этапом онтогенеза. Вопрос о смерти в биологии занимает особое место, ибо чувство смерти «... совершенно инстинктивно присуще человеческой природе и всегда составляло одну из величайших забот человека» (И. И. Мечников, 1913). Больше того, вопрос о смерти стоял и стоит в центре внимания всех философских и религиозных учений, хотя философия смерти в разные исторические времена представлялась по-разному. В античном мире Сократ и Платон доказывали бессмертие души, тогда как Аристотель отрицал платоновскую идею бессмертия души, верил в бессмертность человеческого духа, продолжающего жить после смерти человека.
Цицерон и Сенека также признавали будущую жизнь, но Марк Аврелий считал смерть естественным явлением, которое следует принимать безропотно. В прошлом веке И. Кант и И. Фихте (1762-1814) тоже верили в будущую жизнь, а А. Г. Гегель придерживался убеждений, по которым душа поглощается «абсолютным существом», хотя природа этого «существа» не раскрывалась.
В соответствии со всеми известными религиозными учениями земная жизнь человека продолжается и после его смерти, и человек должен неустанно готовиться к этой будущей смерти. Однако естествоиспытатели и философы, не признающие бессмертия, считали и считают, что смерть представляет собой, как неоднократно подчеркивал И. И. Мечников, естественный исход жизни организма. Более образное определение смерти заключается в том, что она «... есть явная победа бессмыслия над смыслом, хаоса над космосом» (В. Соловьев, 1894).
Научные данные свидетельствуют о том, что у одноклеточных организмов (растений и животных) следует отличать смерть от прекращения их существования. Смертью является их гибель, тогда как прекращение существования связано с их делением. Следовательно, недолговечность одноклеточных организмов компенсируется их размножением. У многоклеточных растений и животных смерть является в полном смысле слова завершением жизни организма.
У человека вероятность смерти повышается в пубертатный период. В частности, в развитых странах вероятность смерти повышается почти экспоненциально после 28 лет.
Различают клиническую и биологическую смерть человека. Клиническая смерть выражается в потере сознания, прекращении сердцебиения и дыхания, однако большинство клеток и органов все же остаются живыми. Происходит самообновление клеток, продолжается перистальтика кишечника.
Клиническая смерть не «доходит» до биологической смерти, ибо она обратима, т. к- из состояния клинической смерти можно «возвращать» к жизни. Например, собак «возвращают» к жизни через 5—6 минут, человека — через 6—7 минут от начала клинической смерти. Биологическая смерть характеризуется тем, что она необратима. Остановка сердцебиения и дыхания сопровождается прекращением процессов самообновления, гибелью и разложением клеток. Однако гибель клеток начинается не во всех органах одновременно. Вначале гибнет кора головного мозга, затем гибнут эпителиальные клетки кишечника, легких, печени, клетки мышц, сердца.
На представлениях о клинической смерти основаны мероприятия по реанимации (оживлению) организмов, что имеет исключительно важное значение в современной медицине.
Продолжительность жизни. Сравнение данных о продолжительности жизни разных представителей флоры и фауны показывают, что среди растений и животных разные .организмы живут разное время. Например, травянистые растения (дикие и культурные) живут в течение одного сезона. Напротив, древесные растения характеризуются большей продолжительностью жизни. Например, вишня живет 100 лет, ель — 1000 лет, дуб — 2000 лет, сосна — до 3000-4000 лет.
Ряд видов членистоногих живет 40-60 лет, рыбы многих видов, например, осетровые живут 55-80 лет, лягушки — 16 лет, крокодилы — 50-60 лет, дикие свиньи — 25 лет, змеи и ящерицы — 25—30 лет, птицы некоторых видов — до 100 лет и более. Продолжительность жизни млекопитающих является меньшей. Например, мелкий рогатый скот живет — 20-25 лет, крупный рогатый скот — 30 лет и более, лошади — 30 лет, собаки — 20 лет и более, волки — 15 лет, медведи — 50 лет, слоны — 100 лет, кролики — 10 лет.
Среди млекопитающих долгожителем является человек. Еще в Библии отмечено, что Мафусаил дожил до 969 лет, а гомеровский герой Нестор прожил три человеческих века, Дандо и один из лак-мейских королей — более 500 лет.
Конечно, эти данные неточны. В действительности же многие люди доживали до 115—120 лет и более. Достоверны случаи, когда отдельные люди доживали даже до 150 лет. В то же время долгожители часто сохраняют на высоком уровне как физические, так и умственные способности. Например, Платон, Микеланджело, Тициан, И. Гёте и В. Гюго лучшие свои произведения создали после 75 лет.
Отмечено, что долгожительство характерно не только для европеоидов. Еще старые авторы отмечали, что отдельные негры жили 115-160 и более лет.
Еще в XVIII в. швейцарский физиолог А. Галлер (1708-1777) отмечал, что столетний возраст имеет семейное распространение, т. е. долговечность представляет собой наследственный признак. Современные данные не опровергают это заключение.
В случае человека различают естественную продолжительность ясизни и среднюю продолжительность жизни. Под естественной продолжительностью жизни понимают количество лет, дальше которых человек не может жить, если даже условия его существования являются самыми благоприятными. Напротив, средняя продолжительность жизни представляет собой длительность жизни индивидов определенной группы, прерываемая смертностью.
В соответствии с существующими представлениями естественная продолжительность жизни является видовым количественным признаком, подверженным контролю со стороны генотипа.
Считают, что такой контроль осуществляется в каждом периоде онтогенеза, причем первые доказательства в пользу этого заключения были получены еще в 60-е годы в экспериментах по культивированию фибробластов человека (табл. 8).
Как видно из табл. 8, выделенные из эмбрионов фибробласты способны к 50-кратному делению, после чего они погибают. Это соответствует примерно 150-летней естественной продолжительности жизни человека. Напротив, фибробласты, выделенные из тканей взрослых людей, способны к значительно меньшему числу генераций. Определенная продолжительность жизни характерна и для тканевого уровня.
Предполагают, что естественная продолжительность жизни является эволюционным приобретением вида. Что касается долгожи-тельства отдельных индивидуумов, то объяснение таких случаев обычно сводится к допущению либо наличия в генотипах долгожителей сочетаний определенных генов, либо наличия небольшого количества или полного отсутствия мутаций в их клетках.
Естественную продолжительность жизни определяют путем установления длительности периода роста человека и длительности жизни. Считают, что человек растет примерно 20 лет, но живет, как показывают долгожители, в 5-7 раз дольше. Руководствуясь этими соображениями, швейцарский физиолог Галлер еще в XVIII в. допускал, что человек может жить до 200 лет. И. И. Мечников тоже считал, что человек может жить до 150 лет, А. А. Бо-гомолец и И. И. Шмальгаузен подсчитали, что естественная продолжительность жизни человека должна составлять 120-150 лет. Однако до 100-летнего возраста доживают лишь отдельные индивиды. Поэтому фактическая средняя продолжительность жизни вопреки ее росту не совпадает с естественной продолжительностью жизни.
На повышение средней продолжительности жизни оказывает влияние ряд факторов (частота рождаемости, снижение детской смертности, эффективность борьбы с инфекциями, успехи хирургии, улучшение питания и общих условий жизни, снижение смертности в результате несчастных случаев), причем эти факторы оказываются более эффективными в случае молодого возраста членов той или иной популяции. Однако при этом естественная продолжительность жизни не увеличивается.
Таблица 8
Зависимость удвоения фибробластов при культивировании
от возраста донора
| Возраст донора фибробластов (в годах)
|
Количество удвоения
|
| Эмбрион
|
50 -10
|
| 0-20
|
30 = 10
|
| 20
|
20 = 10
|
Главнейшие причины снижения средней продолжительности жизни заключаются в детской смертности, а также смертности от голода, болезней, недостаточной медицинской помощи.
Частота смертных случаев обвально снижается в период после рождения и до достижения пубертанного возраста, а затем она повышается (рис. 100). В развитых странах частота смертности повышается почти экспоненциально после возраста около 28 лет.
Средняя продолжительность жизни у древних греков и римлян составляла примерно 30 лет. Средняя продолжительность жизни в Европе составляла в XVI в. — 21 год, в XVII в. — 26 лет, в XVIII в. — 34 года. В конце XIX в. она начала медленно возрастать. В 1988 г. в среднем по всему миру она составляла 61 год, причем в индустриально развитых странах она составляла 73, а в Африке лишь 52 года. Но известны исключения, когда продолжительность жизни растет чрезвычайно быстро, достигая очень высокого уровня, как это имело место в Швеции и Японии (рис. 101, 102).
Частоты рождаемости, смертности и время удвоения количества населения являются разными для разных стран (табл. 9).
В медицинском плане средняя продолжительность жизни — это показатель здоровья нации. СССР по числу старых людей занимал первое место в мире. Например, на 1 млн жителей приходилось 104 человека в возрасте свыше 90 лет, тогда как в Англии — 6, Франции — 7 и США — 15 человек.
Таблица 9
Некоторые популяционные характеристики народонаселения
отдельных стран в 1971 г. (по данным ООН)
| Страна
|
Количество населения (в млн)
|
Частота рождаемости (на 1000)
|
Частота смертности (на 1000)
|
Время, необходимое для удвоения количества населения (в годах)
|
| США
|
207
|
18,2
|
9,3
|
58
|
| Дания
|
5
|
14,6
|
9,8
|
88
|
| Швеция
|
8,1
|
13,5
|
10,4
|
88
|
| Колумбия
|
22,1
|
44
|
11
|
21
|
| Эквадор
|
6,3
|
45
|
11
|
21
|
| Кения
|
11,2
|
50
|
20
|
23
|
| Уганда
|
8,8
|
43
|
18
|
27
|
| Нигерия
|
56,5
|
50
|
25
|
27
|
| Индия
|
570
|
42
|
17
|
27
|
| Пакистан
|
142
|
50
|
18
|
21
|
| Индонезия
|
125
|
47
|
19
|
24
|
В связи с изменениями в продолжительности жизни в настоящее время отмечаются изменения границ трудоспособного населения по сравнению, например, с 30-ми годами нашего столетия. Во многих странах мира отмечается также разрыв между пенсионным возрастом и активностью людей, в результате чего во многих странах мира люди пенсионного возраста продолжают трудиться в той или иной форме. Особенно это распространено в нашей стране.
В 1982 г. в Вене состоялась Всемирная ассамблея по проблемам населения мира, на которой были сформулированы прогнозы по демографической проблематике до 2025 г. В соответствии с этими прогнозами предполагается, что в мире численность людей в возрасте 60 лет и старше по сравнению с 1950 г. возрастет в 5 раз, а людей старше 80 лет — в 7 раз. Другими словами, по данным этого международного форума население Земли постепенно стареет, причем скорость старения населения применительно к разным народам, странам и регионам является различной. Закономерностью является то, что чем ниже жизненный уровень населения, тем быстрее оно стареет.
Гериатрия — это одна из медицинских наук, задачей которой является разработка способов нормализации изменяющихся функций стареющего организма. Начала гериатрии уходят в далекое прошлое, ибо еще Гиппократ в древней Греции придавал большое значение умеренности в пище, приему воздушных и водных ванн. Вслед за ним многие знаменитые врачи прошлого (Гален, Абу Али ибн Сина и другие) также уделяли внимание гериатрии. В наше время проблемы гериатрии разрабатываются во многих научно-исследовательских учреждениях мира.
Однако вопреки успехам в познании биологических основ старения современная гериатрия еще не располагает методами и средствами воздействия  на угасающие с возрастом нормальные физиологические процессы. Поэтому роль гериатрии ограничивается лечением возникающих в пожилом и старческом возрасте заболеваний и исключением (при наличии возможностей) факторов риска, вызывающих преждевременное старение. на угасающие с возрастом нормальные физиологические процессы. Поэтому роль гериатрии ограничивается лечением возникающих в пожилом и старческом возрасте заболеваний и исключением (при наличии возможностей) факторов риска, вызывающих преждевременное старение.
Поскольку растения чрезвычайно разнообразны, то для них характерны специфические онтогенезы. Можно сказать, что содержание онтогенеза растений зависит от их таксономической принадлежности.
В случае одноклеточных организмов растительной природы (бактерий и других) онтогенез может быть определен жизнью клетки на протяжении времени от одного деления до другого. Деление бактериальной клетки на две дочерние клетки можно оценить в качестве завершающего этапа онтогенеза, т. е. ее смерти. Однако многие виды бактерий, например спорообразующих, могут сохраняться длительное время без размножения. Известно также, что циано-бактерии могут сохраняться жизнеспособными (без деления) в донных отложениях озер и прудов на протяжении нескольких десятков лет.
В случае многоклеточных растений, например у цветковых растений, онтогенез начинается с возникновения зародыша в семени и заканчивается смертью растения. У этих растений онтогенез состоит из ряда периодов, которые являются, по существу, возрастными. В частности, различают латентный (покоящиеся семена), дегенеративный, или виргинильный (время от прорастания семени до первого цветения), генеративный (время от первого цветения до последнего) и сепильный, или старческий (от потери способности к цветению до отмирания), периоды. В пределах этих периодов вычленяют далее ряд этапов, важнейшими из которых являются дифференциация соцветия и цветка (цветков), макро- и микроспорогенез, макро- и микрогаметогенез, оплодотворение (зи-гогенез), формирование плода и семени.
Важнейшим моментом на протяжении онтогенеза растений является морфогенез, который зависит от деления и дифференци-ровки неподвижных клеток и который обеспечивается активностью меристем, что ведет к росту растений в той или иной мере на протяжении всей жизни. Во время роста деление клеток происходит почти полностью в меристемах. Различают апикальные и латеральные меристемы. Апикальные меристемы обеспечивают рост растений в основном в длину (высоту), тогда как латеральные меристемы ответственны за толщину растений. Латеральной меристемой служит камбий, из клеток которого формируются проводящие ткани.
Не менее важным в онтогенезе является процесс органогенеза, т. е. образование и развитие корня, стебля, листьев и цветков. При этом нужно заметить, что видовая принадлежность растений определяет сроки и интенсивность заложения и развития тех или иных органов.
Например, у таких растений как ель, которая растет всю жизнь, образование репродуктивных органов и оплодотворение с последующим развитием зародыша осуществляется в течение года, тогда как у некоторых однолетних покрытосеменных растений, жизнь которых определяется одним сезоном, длительность этих процессов составляет лишь около месяца или несколько больше. Рост и деление клеток растений подвержены влиянию света, температуры, гравитации и других факторов.
Размеры, форма и расположение зародыша в семени являются разными у растений разной видовой принадлежности. То же можно сказать и о запасе питательных веществ в зародыше (жиров, углеводов, белков).
Зародыш в семени может находиться длительное время в состоянии покоя, что зависит от дегидратации семян.
Сохранение всхожести семян растений разных видов колеблется, в основном, от одного года до многих десятилетий. Например, семена тех цветов, которые широко известны под названием астр, сохраняют всхожесть один год, тогда как семена многих огородных культур — несколько лет. Известно, что найденные в египетских захоронениях семена злаковых растений сохранили жизнеспособность в течение нескольких тысячелетий.
Условия прорастания семян (освещенность, температура, влажность и др.) растений разной видовой принадлежности также существенно различны. Например, семена одних растений могут прорастать при температуре 0°С, тогда как семена других растений нуждаются в положительных температурах.
Рост и развитие растений подвержены регуляции со стороны фитогормонов (регуляторов роста растений), которые представляют собой сигнальные молекулы и которыми являются ауксины, гиббереллины, цитокинины, абсцизовая кислота и этилен. Перечисленные соединения либо синтезируются в клетках, либо транспортируются к клеткам-мишеням. Наибольший эффект этих соединений проявляется при их сочетанном действии. Например, ауксин, индолилуксусная кислота, стимулирует образование корней, но в сочетании с гиббереллином содействует росту корней в длину, а в сочетании с цитокинином — стимулирует закладку и рост боковых почек.
Предполагают, что наиболее древним является бесполое размножение, в частности, вегетативное размножение. Из последнего развилось размножение спорообразованием, несомненное преимущество которого заключается в том, что оно обеспечивает лучшие возможности для сохранения видов и особенно для расширения их ареалов (расселение).
Половое размножение является наиболее эффективным путем воспроизводства организмов, дающим возможность «перетасовки» и комбинирования генов. Предполагают, что оно развилось из бесполого, возникнув около 1 млрд лет назад, причем первые этапы в этом процессе были связаны с усложнением в развитии гамет. Примитивные гаметы характеризовались недостаточной морфологической дифференцировкой, в результате чего для многих организмов ведущей была изогамия (от греч. isos — равный, gamos — брак), когда половые клетки были подвижными изогаметами, еще не дифференцированными на мужские и женские формы. Изогамия встречается у ряда видов простейших.
В последующем получила развитие анизогамия (от греч. anisos — неравный, games — брак), характеризующаяся дифференцировкой гамет, различающихся между собой лишь по величине. Примером анизогамии является образование гамет также у ряда видов простейших. У этих организмов размеры гамет различны (макрогаметы и микрогаметы).
На более поздних этапах эволюции животных возникли резкие различия в подвижности, форме и размерах гамет, что особенно заметно в случае гамет млекопитающих. Следует добавить также, что у млекопитающих выработалась также способность продуцировать мужские гаметы в огромных количествах.
Биологическая роль полового размножения исключительно велика. Несомненно, что она имеет значительные преимущества по сравнении с вегетативным размножением и размножением споро-образованием. Еще К. А. Тимирязев (1843-1920) неоднократно обращал внимание на половое размножение как на выдающийся источник изменчивости организмов, поскольку в ходе мейоза имеет место рекомбинация генов, а при объединении гамет — образование новых сочетаний генов. Можно сказать, что в природе половое размножение является доминирующим по сравнению с другими формами размножения. У животных, размножающихся половым путем, репродуктивная способность сохраняется относительно долго. Так в случае человека способность к репродукции у женщин сохраняется в основном до 40-45 лет, а у мужчин — практически всю жизнь.
Поскольку для полового размножения требуются два родителя, то это вызывает определенные трудности во встрече партнеров. Однако в ходе эволюции у позвоночных в порядке компенсации выработался также ряд добавочных приспособлений, облегчающих перенос половых клеток самца в половые пути самки и создающих условия для развития оплодотворенной яйцеклетки. Эти приспособления в ходе эволюции развились из выделительной системы, что привело к формированию мочеполовой системы (см. раздел об эволюции половой и выделительной систем).
У многих организмов развилась независимая тенденция к прямому развитию и живорождению. Если прямое развитие отмечается уже у большинства безлегочных саламандр, у всех безногих и у значительной части бесхвостых амфибий, то живорождение у животных появляется, начиная с чешуйчатых рептилий. Появление живорождения обязано преимущественному выживанию на ранних этапах развития внутри материнского организма. Считают, что возникновение живорождения у позвоночных связано с экологическими условиями.
Например, у рептилий оно связано с адаптацией их к жизни в высоких широтах, у амфибий — с адаптацией к низким температурам, а у хрящевых рыб — с адаптацией к жизни в тропических широтах.
Диплоидное состояние сопровождается накоплением различных аллелей. Поэтому половое размножение имеет еще и то преимущество, что оно представляет организмам большую возможность изменчивости по сравнению с бесполым, а это имеет важнейшее значение в эволюции.
Явные репродуктивные преимущества имеет партеногенез, т. к. он продуцирует потомство лишь женского рода. Однако он редок. Для объяснения низкой частоты замещения партеногенезом полового размножения в естественных популяциях организмов используют две гипотезы. В соответствии с одной из гипотез (мутацион-но-аккумулятивной) пол является адаптивным приспособлением, поскольку он «очищает» геном от повторяющихся мутаций, тогда как по другой гипотезе (экологической) пол является адаптивным приспособлением в меняющихся условиях среды. Многие считают, что обе гипотезы справедливы.
У растений в связи с эволюционным развитием неподвижности в образе жизни возникла необходимость в выработке приспособлений, обеспечивающих объединение мужских и женских гамет. Эволюция водных растений привела к появлению подвижных мужских половых клеток. У семенных растений развились пыльца и пыльцевая трубка, а также семя, что способствовало исключительному распространению растений.
Половое размножение растений играет важную роль в их распространении .
1. Опишите бесполое размножение и назовите его формы.
2. Что вы понимаете под половым размножением организмов и какова его биологическая роль?
3. Опишите особенности полового процесса у одноклеточных и многоклеточных организмов.
4. Что такое гаметогенез?
б. Какова функция гамет каждого типа?
6. Какие вы знаете стадии в развитии гамет?
7. В чем заключаются сходства и различия между сперматогенезом и овогенезом?
8. Что такое мейоз и каково его биологическое значение?
9. Опишите фазы мейоза.
10. Влияет ли кроссинговер на результаты мейоза и как?
11. Опишите сущность оплодотворения.
12. В чем заключается разница между зигогенезом и партеногенезом?
13. Какова биологическая роль чередования поколений?
14. Что представляет собой половой диморфизм? Что вы понимаете под гермафродитизмом? Наблюдаются ли случаи гермафродитизма у человека и как часто?
16. Как вы представляете эволюцию способов размножения?
16. Что вы понимаете под ростом и развитием организмов? Какова связь между ростом и дифференциацией клеток?
17. В чем заключаются молекулярные основы дифференцировки клеток?
18. Сформулируйте понятие об онтогенезе и назовите периоды онтогенеза.
19. Каковы различия между прямым и непрямым развитием?
20. В чем заключается влияние оплодотворения на яйцеклетки?
21. На каком этапе реализации генетической информации осуществляется контроль действия генов?
22. Каким образом оплодотворенная яйцеклетка развивается в многоклеточную структуру?
23. Каким образом развивающиеся клетки и ткани становятся отличными одни от других в процессе развития?
24. Могут ли восстанавливаться утерянные или поврежденные части тела?
25. Какова связь в онтогенезе между старостью и продолжительностью жизни?
26. Сформулируйте и определите разницу между естественной (вероятной) и фактической продолжительностью жизни.
27. Какие вам известны теории старения организма?
28. Можно ли считать продолжительность жизни человека видовым призна-ком?
29. Какие факторы оказывают влияние на продолжительность жизни?
Гилберт С. Биология развития. М.: Мир. 1994. 235 стр.
Грин Н„ Стаут У., Тейлор Д. Биология. М.: Мир. 1996. 386 стр.
Нидон К., Летерман И., Шеффель П., Шайба Б. Растения и животные. М
.:
Мир
.
1991. 260
стр
.
Hunter R. Sex Determination, Differentiation and Intersexualiti in Placental Mammole. Cambridge University Press. 1955. 310 pp.
«...признание того,
что явление жизни невозможно
без наследственности, показывает,
что воспроизведение исторически созданных
форм жизни в ряду поколений составляет основу жизни,
обеспечивает прямую физическую связь
на протяжении всех ее этапов,
от первых организмов
до современных видов».
Н. П. ДУБИНИН,
1970
Наследственность и изменчивость — это важнейшие свойства живого, которые не только отличают живое от неживого, но и определяют совместно с размножением бесконечное продолжение жизни, ее непрерывность на всех уровнях организации живого. Теоретическое значение учения о наследственности и изменчивости для биологии невозможно переоценить. Однако данные о наследственности и изменчивости имеют также и выдающееся практическое значение. Они составляют научную основу селекции растений и животных. По образному определению Н. И. Вавилова «...селекция представляет собой эволюцию, направляемую волей человека». Данные о наследственности и изменчивости лежат в основе наших представлений о нормальной и патологической наследственности человека и животных. Наконец, генетическая инженерия, созданная в недрах молекулярной генетики, составляет основу биотехнологии, которая, по образному выражению А. А. Баева, вступила в триумвират с механической и физической технологиями, составляющими становой хребет промышленности.
Наследственность есть категория историческая. Ее ключевая роль в определении непрерывности жизни заключается в том, что она обеспечивает физическую связь между поколениями клеток или организма в виде передачи генетической информации от родителей к их потомству. Но организмы живут в условиях определенной среды. Поэтому непрерывная передача генетической информации от родителей к потомству обеспечивает единство организмов и среды.
Из всех органических молекул способностью к саморепродукции обладают только нуклеиновые кислоты. Между тем, находясь в клетках, они контролируют их структуру и свойства (активность). Поэтому уникальность жизни в генетическом смысле заключается в том, что нуклеиновые кислоты через половые клетки обеспечивают химическую связь между поколениями. Благодаря размножению, наследственности и изменчивости жизнь видов продолжается бесконечно долго как непрерывное чередование поколений с сохранением между ними химических связей.
Уникальность жизни определяется также и постоянством видов. В процессе размножения исходные организмы всегда продуцируют самих себя, т. е. «подобное рождает подобное». Потомство пары мышей всегда является мышами, точно так же, как две бактериальные клетки являются бактериями того же вида, что и их родительская клетка. Следовательно, постоянство видов определяется передачей сходства от родителей к потомству, т. е. наследованием свойств своих родителей, вследствие чего организмы всех поколений (генераций) в пределах вида характеризуются общим наследственным (генетическим) поведением.
Наследственность — это передача сходства от родителей к потомству или склонность организмов походить на своих родителей. Наследственность означает передачу анатомических, физиологических и других, свойств и особенностей от организмов одних поколений (генераций) к организмам других. Поскольку связь между поколениями обеспечивается половыми клетками, а оплодотворение представляет собой слияние ядер этих клеток и образование зиготы, то ядра половых клеток составляют физическую основу такой связи. Когда речь идет о наследственности организмов, то следует понимать, что единственным материалом, который наследуется потомством от своих родителей, является генетический материал, сосредоточенный в ядерных структурах (хромосомах) и представляющий собой гены (единицы наследственности). Следовательно, потомство наследует от своих родителей не признаки (свойства), а гены, которые контролируют эти признаки (свойства), причем показателем генетической детерминируемости признаков является наследуемость последних.
Различают наследование, которое не связано с полом, и наследование, контролируемое, ограничиваемое и сцепленное с полом. Под наследованием, не связанным с полом, понимают то наследование, которое не зависит от пола организмов-родителей или потомства. При наследовании, контролируемом полом, проявление генов отмечают у обоих полов, но по-разному. Наследование, ограничиваемое полом, заключается в том, что проявление генов происходит лишь у одного пола. Наконец, наследование, сцепленное с полом, обусловлено локализацией соответствующих генов в половых хромосомах. Кроме этих типов наследования различают также полигенное наследование, когда наследуемость признака подвержена контролю со стороны нескольких генов.
Однако организмы, которые произошли от какой-то пары родителей, не все являются совершенно одинаковыми. В одном и том же помете мышей или в одной и той же культуре бактерий (происходящей от одной бактериальной клетки) можно встретить организмы, которые по каким-то признакам будут отличаться от родителей. Иногда у потомства обнаруживаются признаки, которые были присущи лишь далеким предкам, или признаки, которые являются совершенно новыми не только для их родителей, но и для их далеких предков. Следовательно, для индивидуальных организмов характерны различия, изменчивость признаков.
Противоположным свойством наследственности является изменчивость. Она заключается в изменениях генетического материала, сопровождаемых изменениями признаков организма. Результатом изменчивости является образование новых вариантов организмов, непрерывность разнообразия жизни.
Непрерывность жизни имеет генетический характер, ибо на-следсвенность и изменчивость поддерживают стабильность свойств и способность организмов к изменчивости. Однако генетическая непрерывность жизни связана не только с наследственностью и ее изменчивостью. Она связана также со средой, в которой живут и развиваются организмы.
Все организмы живут и размножаются в среде, условия которой не безразличны для них. До тех пор, пока организм живет, его наследственность взаимодействует со средой. Внешняя вреда влияет на выражение наследственных признаков и определяет степень их проявления. Взаимодействие наследственности и среды определяет, каким организм является в данный момент и как он должен развиваться в будущем. Можно сказать, что наследственность предполагает, каким организм должен стать, но не каким он будет. То, каким организм станет в действительности, решается взаимодействием наследственности и среды.
Геном — это сумма генов или полное количество ДНК, характерное для клеток организма определенного вида.
Генотип — это сумма генов данного организма, его индивидуальная генетическая конституция, которую он получает от своих родителей. Генотип относительно стоек не протяжении всей жизни индивидуума. Для взрослого человека, независимо от его возраста, характерен тот же генотип, который был присущ ему в период внутриутробного роста и развития, в детстве, отрочестве, юности.
Фенотип — сумма всех внешних и внутренних признаков (свойств) данного организма. У всех организмов различают качественные и количественные признаки. Качественными признаками служат те, которые можно, глядя на них, сфотографировать или описать, причем степень достоверности в описании зависит от умения описывающего. Так К. Линней настолько ярко описал качественные признаки домашней собаки, что эти описания уже два столетия переходят из одного учебника в другой без изменений. Такими признаками организмов являются половые различия, форма тела, строение, масть животного, окраска цветков и плодов, форма семян, плодов и т. д. Особенно разнообразны качественные признаки у человека. Они специфичны применительно к каждому индивидууму.
Количественными признаками служат те, которые можно определить путем измерений. Например, количественными признаками у растений являются масса семян, плодов, количество, форма и размеры листьев, высота стеблей, урожайность и т. д. У домашних животных количественными признаками являются молочная и мясная продуктивность, белковое содержание мяса, количество жира в молоке коров, яйценоскость кур, масса яиц, оплата корма и т. д. В растениеводстве и животноводстве учет количественных признаков имеет очень большое значение не только в хозяйственном плане, но и в том, что их используют в селекции высокоурожайных сортов растений и высокопродуктивных пород животных, ведя отбор на хозяйственно полезные признаки. Как правило, количественные признаки и у растений и у животных контролируются не одним, а большим количеством генов, действующих в одном направлении. У человека количественными признаками являются масса тела, головного мозга, масса и размеры внутренних органов, рост, количество форменных элементов крови, степень пигментации кожи, общая интеллектуальность и т. д. Как и в случае растений и животных количественные признаки человека тоже подлежат генетическому контролю, т. е. являются полигенами.
В противоположность генотипу фенотип любого организма изменяется в процессе роста и развития на протяжении всей его жизни. В случае человека изменения фенотипа у отдельного индиви-диума можно проследить по его фотографиям, сделанным в разные периоды жизни. Можно сказать, что фенотип организма является различным в онтогенезе индивидуума, т. е. в эмбриональном периоде, после рождения, во время полового созревания и т. д.
Генотип организма определяют путем наблюдения его действия (реализации генетической информации) и влияния на фенотип в условиях определенной среды. Когда два или несколько организмов растут и развиваются в одинаковых условиях, но фенотипы их различны, то это означает, что такие организмы имеют разные генотипы. Строго говоря, фенотипы являются результатом взаимодействия различных генов (компонентов генотипа) между собой и генотипа со средой. Поэтому нельзя думать, что организм или какие-либо признаки организма зависят исключительно только от генотипа или только от среды. Два одинаковых генотипа могут развиваться в разных условиях и дать разные фенотипы. Точно так же два разных генотипа могут развиваться в условиях одинаковой среды и дать разные фенотипы. Кроме того, если организмы, развивающиеся в условиях сходной или неодинаковой среды, имеют сходные фенотипы, это еще не означает, что их генотипы одинаковы. В таких случаях часто имеет значение доминантность признаков, которая может осложнять установление генотипа.
Фенотип организма с определенным генотипом формируется не только под влиянием факторов среды, действующих в данное время, но и тех факторов, которые действовали ранее на протяжении всей жизни организма. В случае человека любой индивидуум с его физическими, анатомическими, физиологическими и психическими свойствами является продуктом роста и развития, детерминируемого определенным генотипом и осуществляющегося в условиях среды с определенной последовательностью различных факторов этой среды, включая социальные. Другими словами, каждый индивидуум представляет собой продукт его генотипа и жизненного опыта. Например, японцы, проживающие в США, превосходят по росту японцев на их родине в Японии, причем это объясняют характером диеты. Однако в обоих случаях диапазон ростовых характеристик детерминируется генетически.
Наиболее демонстративно взаимодействие наследственности и внешней среды проявляется у однояйцевых (идентичных) близнецов. Многие наблюдения свидетельствуют о том, что жизнь и воспитание идентичных близнецов в различных семьях и в условиях разной среды всегда приводили к тому, что близнецы сохраняли фенотипическое сходство, но различались между собой как личности; пример, касающийся однояйцевых близнецов, свидетельствует о том, что наследственно задатки проявляются лишь в условиях определенной среды. В частности, на развитие умственных способностей человека, которые сами по себе детерминированы генетически, может оказывать влияние и среда.
Итак, хотя фенотип нельзя свести только к генотипу или среде, различия в фенотипе могут определяться раздельными или совместными различиями генотипа или среды, наследственность и среда постоянно взаимодействуют, определяя свойства организмов. Это, однако, не означает абсолютного влияния среды на проявление всех признаков. Известны отдельные признаки, развитие которых настолько сильно ограничено генотипом, что они не подвержены модификации ни одним из известных факторов среды. Можно сказать, что эти признаки генетически очень узко детерминированы к существующему разнообразию среды. Примерами таких признаков являются группы крови и цвет глаз у человека. Одновременно есть признаки, которые зависят от внешней среды, например, уродства новорожденных в результате приема ядов или алкоголя беременными женщинами, но такие признаки не имеют прямого отношения к наследственности.
В биологии большое значение имеют вопросы, касающиеся природы изменчивости организмов и отношения изменчивости к наследственности, ибо причины различий между индивидуальными организмами не всегда одинаковы и могут быть обусловлены как факторами среды, так и факторами наследственности (генами).
Нельзя никогда найти пару организмов одного вида, которые были бы совершенно одинаковы фенотипически. В лесу, степи или на возделываемой делянке даже рядом растущие растения различаются между собой, ибо они получают разное количество света, воды, минеральных веществ. Животные также различны между собой в пределах одного вида, т. к. никогда не получают точно одинакового количества корма в разное время. Следовательно, находясь в разных условиях по отношению к питательным веществам, свету, температуре и другим внешним факторам, организмы даже с одинаковым генотипом всегда различаются между собой феноти-пически. Такие различия между сходными по генотипу организмами получили название фенотипической (модификационной) изменчивости, модификации или ненаследственной изменчивости.
Однако различия между организмами могут определяться и другими причинами. При одних и тех же условиях щенок всегда вырастает в собаку, а котенок — в кошку, ибо организмы этих видов имеют принципиально различные генетические основы.
Известно, что рост мужчин в общем является большим, чем у женщин. Однако иногда женщины выше мужчин, а у высокорослых родителей рождаются дети меньшего роста. Эти различия связаны с тем, что данный наследственный признак детерминируется многими генами, экспрессия которых может меняться. Следовательно, в случае разных генотипов индивидуальные организмы одного и того же вида также могут различаться между собой по отдельным признакам. Поэтому изменчивость, детерминируемую наследственными факторами, называют генотипической или наследственной изменчивостью. Ее возникновение связано с изменениями (мутациями) генов и хромосом, а также рекомбинациями генов. По этой причине данную изменчивость называют еще мутационной, или рекомбинационной (комбинативной) изменчивостью (в зависимости от мутаций или рекомбинаций генов). Сочетания мутантных генов с немутантными или другими мутантными генами, а также рекомбинации генов и хромосомные мутации создают генотипическое разнообразие организмов (см. § 47).
Изучая мутационную изменчивость культурных злаковых растений и их диких предковых форм, Н. И. Вавилов (1887—1943) сформулировал закон гомологичных рядов наследственности, в соответствии с которым у этих организмов мутационный процесс протекает параллельно, а возникающие мутации характеризуются сходством, образуя гомологичные ряды. По Н. И. Вавилову гомологичные ряды наследственности являются отражением сходства генотипов организмов, входящих в эти ряды. В рамках закона гомологичных рядов наследственности изменчивость организмов представлена в виде закономерного явления, присущего видам организмов. Этот закон явился также основой в подборе исходных форм для скрещиваний с последующей селекцией полезных форм организмов.
Действие закона гомологичных рядов наследственности, который сформулирован в применении к растениям, распространяется на животных и человека, наиболее яркой иллюстрацией этого заключения является моделирование многих болезней человека (наследственных и не наследственных) на животных, т. к. многие болезни одинаково встречаются как у человека, так и у животных (антропозоонозы).
Принципиальное значение имеет определение степени раздельного влияния наследственности и среды на фенотипические различия индивидуальных организмов в пределах видов. Вопреки тому, что этот вопрос уже очень давно обсуждался в генетике, оценка этих влияний и до нашего времени остается сопряженной со многими трудностями и в каждом отдельном случае нуждается в специальном рассмотрении. Тем не менее практика сельского хозяйства и экспериментальные исследования с растениями и животными свидетельствуют о том, что такое определение в применении к растениям и животным вполне возможно.
Известно, что улучшение агротехники при культивировании растений или условий содержания при разведении домашних животных, генотипы которых характеризуются не очень благоприятными возможностями, приводит лишь к некоторому повышению урожая растений или продуктивности животных, причем не воспроизводимому в потомстве этих организмов. В то же время среди культивируемых растений всегда можно найти генотипические варианты, которые дают больший урожай, а среди животных — генотипические варианты, характеризующиеся большей продуктивностью по мясу, молоку, шерсти или какому-либо другому количественному признаку. Давно замечено, что даже незначительное генотипическое улучшение дает эффект, поскольку контролирующие его гены передаются по наследству, а генотипическое улучшение воспроизводится в потомстве. Более того, генотипическое совершенствование продуктивности культурных растений и домашних животных привело к созданию огромного сортового и породного разнообразия этих организмов. Разумеется, сортовые достоинства растений зависят от качества почвы, климатических условий, количества и качества удобрений и т. д., а породные достоинства животных — от условий их содержания и кормления. Таким образом, практика сельского хозяйства свидетельствует, что фенотипические различия между организмами определяются в основном генотипом. В то же время практический опыт свидетельствует, что наибольшие результаты в растениеводстве получают сочетанием высоких сортовых достоинств растений с удобрениями и различными агрономическими приемами. Это же имеет место и в животноводстве, где наибольшая продуктивность достигается при сочетании высоких породных достоинств животных с благоприятными условиями их кормления и содержания. Именно на основе знания этих особенностей непрерывно ведется селекционная работа по созданию новых высокоурожайных культур растений и высокопродуктивных пород животных. Высокие сортовые качества растений и породные качества животных в значительной мере компенсируют другие недостатки в хозяйственной деятельности (недостаток удобрений, кормов и т. д.), но их реализация полностью возможна лишь в условиях высокой агротехники или кормления и содержания (соответственно). Например, высокие урожаи основных продовольственных культур получают не только по причине сортовых достоинств растений, но и в результате внесения значительных количеств удобрений и достаточного орошения земель.
Более точные данные о степени влияния генотипа и среды на фенотипические различия дали многочисленные старые и новые экспериментальные исследования, выполненные в условиях контролируемой среды и использования организмов в виде клонов, чистых и инбредных линий со сходными генотипами или генотипами, различающимися между собой по определенным генам.
Клоном является потомство вегетативно размножающегося индивидуального организма, например, культура бактерий, полученная в результате размножения одноклеточной бактериальной клетки, культура соматических клеток, полученная из одиночных соматических клеток животного или человека, растения, полученные из одиночных клеток исходного растения (микроклональное размножение растений), группа деревьев или кустарников, развившихся из черенков, взятых от одного растения.
Чистая линия — это потомство, полученное от индивидуального организма в результате самоопыления (в случае растений) или самооплодотворения (в случае животных). Размножение многих культивируемых растений (пшеница, овес, фасоль, горох и т. д.) происходит главным образом путем самоопыления, поэтому здесь получение чистых линий не вызывает затруднений. У животных же самооплодотворение — это довольно редкое явление, но оно все же имеет место, например у пресноводных улиток.
Как и в случае клонов, все члены линии являются генетически однородными организмами, т. к. обладают одинаковыми наследственно-константными признаками. Их генетическое однообразие более совершенно, чем у потомства, получаемого после перекрестного опыления разных организмов.
Инбредные линии — это организмы, разводимые в мире раздельнополых животных путем неоднократных скрещиваний между собой близких родственников (братьев и сестер). Генотипическое разнообразие инбредных животных становится более выраженным с каждым новым скрещиванием. Например, известен ряд инбредных линий белых мышей, крыс и морских свинок. Исследования клонов чистых и инбредных линий организмов позволили не только измерить действие факторов внешней среды, но и более точно определить влияние генотипа на фенотипические различия. Научные результаты, полученные в этой области, совпадают с данными сельскохозяйственной практики.
Многочисленные исследования взаимодействия генотипа и среды на примере организмов многих видов показали, что для реакции определенного генотипа в ответ на фактор внешней среды всегда характерен диапазон, измеряемый количеством фенотипов, продуцируемых этим генотипом. Разнообразие фенотипов, возникающих в результате взаимодействия определенного генотипа с разными факторами среды (разными средовыми условиями), генетики называют нормой реакции этого генотипа.
Многие генетически детерминированные реакции организмов на внешние факторы среды имеют адаптивный характер, что обеспечивает жизнь и размножение организмов в колеблющихся условиях среды. Среди адаптивных реакций различают физиологический гомеостаз и гомеостаз развития. Физиологический гомеостаз — это генетически детерминированная способность организмов противостоять колеблющимся условиям внешней среды У млекопитающих, в том числе у человека, типичным примером физиологического гомеостаза является константность осмотического давления в клетках и концентрация водородных ионов в крови вследствие функционирования почек и наличия в крови буферных субстанций. Гомеостаз развития — это генетически детерминированная способность организмов так изменять отдельные реакции, что функции организмов при этом в целом сохраняются. Например, выход из строя одной почки сопровождается тем, что остающаяся почка выполняет двойную нагрузку. Примером гомеостаза развития может быть также приобретение переболевшим организмом иммунитета против соответствующей инфекции.
Часто между физиологическим гомеостазом и гомеостазом развития очень трудно выявить различия, поэтому многие адаптивные реакции носят промежуточный характер. Примером такой адаптивной реакции является изменение количества эритроцитов в крови у людей в зависимости от пребывания их на той или иной высоте над уровнем моря. Количество эритроцитов у людей, живущих в разных высотных условиях, повышается по мере удаления от уровня моря. Это связано с тем, что уменьшение содержания кислорода в атмосфере вызывает интенсификацию его транспорта эритроцитами в результате увеличения количества последних. Возвращение человека из высокогорного района в район, лежащий на уровне моря, сопровождается снижением количества эритроцитов.
Норма реакции у всех организмов имеет пределы, определяя их фенотипическое разнообразие лишь в условиях среды, которая для организмов любого вида не имеет резких и необычных отклонений. Например, многие тропические растения выживают в условиях повышенных или пониженных температур, характерных для стран с жарким климатом. Однако они погибают от мороза, к которому устойчивы растения, являющиеся обитателями северных широт. В случае человека потеря генотипом в результате мутации способности детерминировать адаптивные реакции на факторы обычной для него среды сопровождается наследственным заболеванием.
Оценка различных форм взаимодействия наследственности и среды позволяет считать, что наследуется генотип, но не фенотип, т. е. наследуются гены, но не свойства и признаки. Можно далее сказать, что свойства и признаки организмов формируются в процессе развития индивидуума, причем развитие находится под контролем генов и факторов среды. Последние могут изменять проявление признаков, определяемое нормой реакции. Следовательно, каждый признак организмов обусловлен как наследственностью, так и средой.
Главным и единственным методом изучения наследственности организмов является классический генетический (гибридологический) анализ, или, как его еще называют, формальный генетический анализ. Основы этого метода были разработаны Г. Менделем. Этот метод заключается в последовательном разложении генома анализируемого организма на группы сцепленных генов, а групп сцепления — на генные локусы с дальнейшим установлением последовательности генных локусов вдоль хромосомных пар и выяснением тонкой структуры генов.
Генетический анализ в принципе подобен химическому анализу, задача которого заключается в разложении сложных химических соединений на более простые компоненты. Однако в отличие от химического анализа, например нуклеопротеидов, расщепление которых на структурные части основано на гидролизе, классический генетический анализ основывается на расщеплении (сегрегации) и рекомбинации генов в мейозе и осуществляется путем скрещиваний особей с разными признаками и учета результатов скрещиваний.
Схема генетического анализа организмов состоит из ряда последовательных этапов, а именно:
1. Идентификация генов.
2. Установление генных локусов на хромосомных парах.
3. Установление последовательности генных локусов вдоль хромосомных пар.
4. Выяснение тонкой структуры генов.
Результаты генетического анализа оформляют путем составления генетических карт.
Одним из важнейших показателей эффективности генетического анализа является его разрешающая способность, которая в общих чертах может быть аналогизирована с разрешающей способностью оптических методов исследования. Подобно тому, как разрешающая способность оптических приборов (микроскопов) ограничена волновой природой света, разрешающая способность генетического анализа ограничивается количеством исследуемого потомства, получаемого в скрещиваниях, ибо чем большим является количество потомства, тем большей является возможность обнаружения среди них редких рекомбинантов и, следовательно, установления частоты кроссинговера.
Начиная с 1910 г., в генетике в качестве экспериментальной модели (системы) широко используют плодовую мушку Drosophila melanogaster (рис. 103). Являясь эукариотом с дифференцированными тканями, этот организм очень удобен для изучения многих вопросов наследственности.
В частности, у этого организма было идентифицировано и изучено большое количество генных и хромосомных мутаций, причем хромосомные мутации из-за больших размеров в клетках слюнной железы оказались доступными для изучения с помощью обычного микроскопа.
На этом организме была показана «мощь» генетического анализа. Однако разрешающая способность генетического анализа всегда имеет ограничения, поскольку возможность получения большого количества потомства всегда ограничена до определенных пределов даже у тех видов, у которых оно составляет сотни организмов на пару, как, например, у D. melanogaster. Поэтому у организмов, размножающихся половым путем, в том числе и у плодовой мушки возможно выполнение лишь трех первых этапов генетического анализа.
 Однако изучение других генетических систем, в частности микроорганизмов, показало, что половая репродукция не является единственным путем, при котором осуществляется объединение, расщепление и рекомбинация генетических структур, происходящих от исходных (родительских) организмов. Эти процессы могут проходить и при других формах генетического обмена. У микроорганизмов (Е. coli) бактериальных вирусов (фагов) и микроскопических грибов такими формами генетического обмена являются трансформация, конъюгация и трансдукция. Общим для них в сравнении с половой репродукцией высших организмов является то, что они приводят к объединению в одной клетке родительских генов и обеспечивают их расщепление и рекомбинацию, т. е. являясь альтернативами половой репродукции, представляют собой системы рекомбинации. Поэтому генетический анализ основывается и на таких системах рекомбинации. Больше того, использование этих систем рекомбинации привело к повышению разрешающей способности генетического анализа в гигантских размерах, ибо появилась возможность оперировать с огромным количеством организмов в потомстве, а также легко осуществлять тесты комплементации, а это позволило не только создать генетические карты ряда организмов (Е. coli, В. subtilis, фаги, низшие грибы), но и изучить тонкое строение их генов. Однако изучение других генетических систем, в частности микроорганизмов, показало, что половая репродукция не является единственным путем, при котором осуществляется объединение, расщепление и рекомбинация генетических структур, происходящих от исходных (родительских) организмов. Эти процессы могут проходить и при других формах генетического обмена. У микроорганизмов (Е. coli) бактериальных вирусов (фагов) и микроскопических грибов такими формами генетического обмена являются трансформация, конъюгация и трансдукция. Общим для них в сравнении с половой репродукцией высших организмов является то, что они приводят к объединению в одной клетке родительских генов и обеспечивают их расщепление и рекомбинацию, т. е. являясь альтернативами половой репродукции, представляют собой системы рекомбинации. Поэтому генетический анализ основывается и на таких системах рекомбинации. Больше того, использование этих систем рекомбинации привело к повышению разрешающей способности генетического анализа в гигантских размерах, ибо появилась возможность оперировать с огромным количеством организмов в потомстве, а также легко осуществлять тесты комплементации, а это позволило не только создать генетические карты ряда организмов (Е. coli, В. subtilis, фаги, низшие грибы), но и изучить тонкое строение их генов.
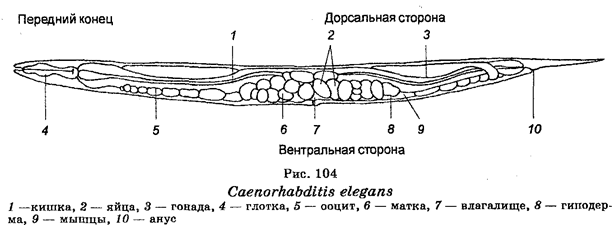
В качестве экспериментальных моделей широко используют также дрожжи. Являясь простейшими эукариотами, эти организмы обладают всеми преимуществами бактерий. Но кроме этого, они оказались доступными для изучения на них генетики митохондрий, сплайсинга РНК, гаплои-дии и диплоидии.
 Классический генетический анализ используют в генетике растений и животных, а также их культивируемых клеток. Однако по отношению к высшим организмам тех видов, которым присуще длительное время между генерациями и малое количество потомства на пару, он либо невозможен, либо очень затруднен. Из-за невозможности классического генетического анализа организмов ряда видов изучение их наследственности проводят с помощью других методов. Например, для изучения наследственности человека используют метод родословных (генеалогический анализ), цитогенетический, популяционный, близнецовый и другие современные методы (см. гл. XIII). Классический генетический анализ используют в генетике растений и животных, а также их культивируемых клеток. Однако по отношению к высшим организмам тех видов, которым присуще длительное время между генерациями и малое количество потомства на пару, он либо невозможен, либо очень затруднен. Из-за невозможности классического генетического анализа организмов ряда видов изучение их наследственности проводят с помощью других методов. Например, для изучения наследственности человека используют метод родословных (генеалогический анализ), цитогенетический, популяционный, близнецовый и другие современные методы (см. гл. XIII).
Длительное время для изучения генетического контроля развития животных организмов использовали D. melanogaster. Однако, начиная с 60-х гг., в качестве модельного объекта в генетике развития стали использовать круглого гельминта Caenorhabditis (рис. 104). Имея длину в 1 мм, эта нематода состоит примерно из 1000 клеток. Ее генетический аппарат представлен 6 парами гомологичных хромосом, на которых локализовано около 3000 генов. В гаплоидном состоянии геном состоит из 8´107
пар нуклеотидов.
Что касается растений, то для изучения генетики развития этих организмов используют травянистое растение Arabidopsis thaliana (рис. 105). Преимущества этого растения в качестве экспериментальной модели заключаются в том, что его легко культивировать в лабораторных условиях и что оно имеет очень короткий срок вегетации (всего лишь 5 недель). Кроме того, геном этого растения состоит из 7´107
нуклеотидных пар.
У всех этих организмов идентифицированы различные мутации, созданы их геномные библиотеки и секвенировано большинство генов. Секвенирование стало методом изучения тонкого строения генов у всех организмов.
Развитие молекулярной биологии привело к разработке методологии генетической инженерии, которая нашла исключительно широкое применение в животноводстве, растениеводстве, а также в изучении нормальной и патологической наследственности человека (см. раздел V).
1. Дайте определение наследственности и объясните, каким образом наследственность определяет непрерывность жизни?
2. Является ли изменчивость свойством живого и если да, то почему?
3. Какие формы изменчивости вы знаете?
4. Что важнее, наследственность или среда?
5. Дайте определение генотипа и фенотипа.
6. В чем заключается классический генетический анализ и применим ли он для изучения наследственности всех организмов? Каковы его возможности и ограничения?
7. Для чего используют в изучении наследственности и изменчивости экспериментальные модели?
8. Какие организмы используются в генетике в качестве моделей для изучения генетических закономерностей?
9. На каких уровнях изучают наследственность и изменчивость?
10. Каково значение молекулярно-генетических исследований наследственности и изменчивости?
11. Можно ли изучать тонкое строение генов, не прибегая к скрещиваниям?
Дубинин Н. П. Генетика. Кишинев: Штиинца. 1986. 534 стр.
Грин Н., Стаут У., Тейлор Д. Биология. М.: Мир. 1996. 386 стр.
Schleif R. Genetics and Molecular Biology. The Johns Hopkins University Press. 1993. 698 pp.
В соответствии с современными представлениями генетическим материалом являются нуклеиновые кислоты.
Нуклеиновые кислоты были обнаружены в ядрах клеток в 1869 г. швейцарским физиологом Фридрихом Мишером. Это открытие является настолько важным, что оно заслуживает приведения здесь цитаты из работы Ф. Мишера, в которой он описывал свои опыты, а именно: «Обрабатывая клетки гноя слабыми щелочными растворами, я получил в результате нейтрализации раствора осадок, который не растворялся ни в воде, ни в уксусной кислоте, ни в разведенной соляной кислоте, ни в обычном солевом растворе и который не мог принадлежать ни к одному из белков, известных в настоящее время». Обнаруженное вещество Ф. Мишер назвал «нуклеином». Как считают, он не мог знать, что открыл ДНК и что оказался в начале исследований ДНК. Но, определяя заслуги Ф. Мишера в качестве первооткрывателя нуклеиновых кислот, нельзя не отметить, что первое предположение о роли нуклеиновых кислот в качестве генетического материала было сформулировано в 1914 г. доцентом Петербургского университета А. Щепотьевым.
Материалом, из которого состоят гены, является дезоксирибо-нуклеиновая кислота (ДНК), а материалом, обеспечивающим декодирование генетической информации, являются РНК. У отдельных вирусов первичным генетическим материалом является рибонуклеиновая кислота (РНК). Таким образом, нуклеиновые кислоты являются хранителями (носителями) и переносчиками генетической информации.
Сложившиеся представления о том, что первичным генетическим материалом у абсолютного большинства живых существ является ДНК, основаны на ряде прямых и косвенных доказательств, среди которых исторически самым первым прямым доказательством генетической роли ДНК является установленная еще в 1944 г. способность ДНК трансформировать пневмококки из одного типа в другой. К настоящему времени трансформация установлена у микроорганизмов многих видов. В 50-е годы прямые доказательства генетической специфичности ДНК были получены также в результате изучения размножения бактериальных вирусов в бактериях и переноса ими генетической информации с помощью ДНК от одних бактериальных клеток к другим. Это явление известно под названием трансдукции. Тогда же было установлено, что перенос ДНК от одних бактериальных клеток к другим осуществляется также в процессе конъюгации бактерий. К настоящему времени трансдукция и конъюгация изучены у многих видов бактерий. В совокупности, на трансформации, трансдукции и конъюгации основывается генетический анализ микроорганизмов.
Решающее прямое доказательство генетической роли ДНК было обеспечено разработкой методов генной инженерии, создавшей возможность конструирования рекомбинантных молекул ДНК с заданными свойствами. К настоящему времени возможности генной инженерии показаны на примере клонирования многих генов самых различных организмов. Что касается косвенных доказательств, то они известны очень давно и их несколько. Для ДНК характерна специфичность локализации в клетках, поскольку она обнаруживается только в ядрах клеток (хромосомах), митохондриях (у животных) и хлоропластах (у растений). У многих микроорганизмов ДНК локализована только в ядерной области (нуклеоиде) или в цитоплазме в виде плазмид. Для организмов каждого вида характерно определенное количество ДНК на клетку (табл. 10).
Таблица 10
Количество нуклеотидов в геномах различных организмов*
| Организмы
|
Гаплоидньгй ядерный геном
|
Митохондри-альныи геном
|
| Гаплоидный набор хромосом
|
Нуклеотидные пары
|
| Нуклеотидные пары
|
| Аденовирус
|
3,0 х 105
-З.З x 105
|
| Фаг Т2
|
2,0 x 104
|
| Фаг Т5
|
1,3 x 105
|
| Е. coli
|
4,5 х 10е
|
| Дрожжи
(Saccharomyces cerevisiae)
|
18
|
1,8 х 107
|
7,4 х 104
|
| (A. tholiana)
|
7,0 х 107
|
| Дрозофила (D. melanogaster)
|
4
|
2,7 х 107
|
1,8 х 104
|
| Лягушка (Rana pipiens)
|
13
|
6,0 х 109
|
1,6 х104
|
| Мышь (Mus musculus)
|
3,0 х 109
|
1,5 х 104
|
| Крыса (Rattus norwegius)
|
21
|
6,0 х 109
|
1,5 х 104
|
| Человек (Homo sapiens)
|
23
|
5,8 х 109
|
1,5 х 104
|
*1000 пар оснований = 617 500 дальтон
Данные, которые приведены в табл. 10, показывают, что, начиная с вирусов, содержание ДНК прогрессивно повышается у бактерий, а затем и у позвоночных и, таким образом, находится в прямой связи со сложностью организации и поведения организмов. Известны, однако, исключения.
Для организмов каждого вида характерно то, что количество ДНК в соматических (диплоидных) клетках является вдвое большим, чем в половых (гаплоидных). Будучи постоянным на клетку у организмов всех видов, содержание ДНК не подвержено влиянию со стороны физиологических факторов, включая пол и старение, а также неблагоприятных воздействий, например, голодания, повышения или понижения температуры. Косвенными доказательствами генетической специфичности ДНК являются также данные о способности искусственно синтезированных аналогов азотистых оснований ДНК вызывать наследственные изменения клеток вследствие прямого включения их в ДНК клеток и данные о том, что мутагенный эффект УФ-излучения тесно связан со спектром поглощения его молекулами ДНК.
Молекулы ДНК являются линейными макромолекулами, представляющими собой длинные двойные цепи (тяжи) полимеров, составленных из мономеров, получивших название нуклеотидов (малых органических молекул) и являющихся строительными блоками ДНК. Электронномикроскопическое изображение ДНК показано на рис. 106.
У всех живых существ макромолекулы ДНК построены по одному и тому же плану. Они слагаются в основном из одних и тех же нуклеотидов, каждый из которых содержит по одной молекуле фосфорной кислоты и сахара, а также одно из четырех азотистых оснований — аденин, гуанин, цитозин или тимин. Аденин и гуанин являются пуриновыми основаниями, тогда как тимин и цито- зин — пиримидиновыми. Пурины и пиримидины называют основаниями по той причине, что в кислой среде они способны присоединять к себе ион H'1
'. Пиримидины являются производными шестичленного пиримидинового кольца, тогда как пурины представляют основания, у которых второе пятичленное кольцо слито с шестичленным кольцом.

Сахаром в ДНК является 2-дезокси-D-рибоза, отличающаяся от глюкозы тем, что в ее молекуле не 6, а 5 атомов углерода, т. е. является пятиуглеродным сахаром (пентозой). Особенностью этого сахара является также то, что он имеет атом водорода (Н), присоединенный к одному (специфическому) из атомов углерода, но не гидроксильную группу. Следовательно, этот сахар представляет собой дезоксирибозу, т. к. он является рибозой, лишенной кислорода.
Сахарофосфат соединяется с азотистым основанием посредством р-гликозидной связи. Основание прикрепляется к I положению де-зоксирибозы. Структура, образованная соединением азотистого основания и сахара, носит название нуклеозида. Таким образом, химическими группами, которые образуют ДНК, являются пуриновые и пиримидиновые азотистые основания (аденин, гуанин, тимин и цитозин), сахар (2-дезокси-D-рибоза) и фосфорная кислота.
РНК характеризуется такой же структурой, как и ДНК. Однако в отличие от ДНК в РНК сахаром является рибоза с кислородом, представляющая собой сахар с 5 атомами углерода, к одному из которых прикреплена 2'-гидроксильная группа (-ОН). Кроме того, в РНК тимин не имеет метильной группы и является урацилом, т. е. в РНК тимин заменен на урацил, также являющийся пиримидиновым основанием.
Нуклеиновые кислоты называют кислотами по той причине, что их фосфатные группы освобождают в растворах ионы водорода.
Для состава ДНК характерны закономерности, известные в качестве правил А. Чаргаффа, а именно:
1. Сумма нуклеотидов, содержащих пуриновые азотистые основания, равна сумме нуклеотидов, содержащих пиримидиновые азотистые основания

2. Содержание аденина равно содержанию тимина, а гуанина — цитозину
(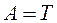 или или 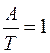 и и 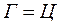 или или 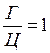 ). ).
3. 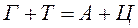 или или 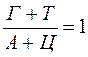
4. ДНК из разных источников может иметь различия, обусловленные в одних случаях преобладанием аденина над гуанином и тимина над питозином (А + Т > Г + Ц), в других случаях — преобладанием гуанина и цитозина над аденином и тимином (Г + Ц > А + Т).
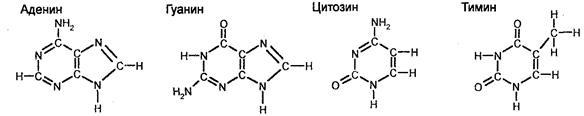
Данные о нуклеотидном составе оснований ДНК разных организмов приводятся в табл. 11.
Таблица 11
Нуклеотидный состав оснований ДНК разных организмов
| Организмы
|
Среднее значение содержания ГЦ в %
|
| Вирусы
|
28-74
|
| Бактерии
|
26-74
|
| Водоросли
|
36-68
|
| Грибы
|
36-54
|
| Высшие растения
|
36-48
|
| Простейшие
|
22-62
|
| Беспозвоночные
|
34-44
|
| Позвоночные
|
40-44
|
Для ДНК характерна структура трех видов — первичная, вторичная и третичная. Первичная структура ДНК заключается в том, что ДНК состоит из нуклеотидных цепей, у которых скелетную основу составляют чередующиеся сахарные и фосфатные группы, объединенные ковалентными 3'-, 5'-фосфодиэфирными, скелетными связями, а боковые группы представлены тем или иным основанием (одним из четырех) и присоединяются одна к другой молекулой сахара. Последовательно располагающиеся нуклеотиды ковалентно связаны фосфодиэфирными связями между сахарным остатком и фосфатом, и в результате этого объединены в полинук-леотидную цепь. Таким образом, первичная структура ДНК (как и РНК) определяется последовательностью нуклеотидов и характером их связей между сахарным остатком и фосфатом.
Представления о вторичной структуре ДНК (рис. 107) были сформулированы Д. Уотсоном и Ф. Криком еще в 1953 г. На основе данных об Х-дифракции молекул ДНК, структуре оснований и правил А. Чаргаффа эти представления сводятся к следующему:
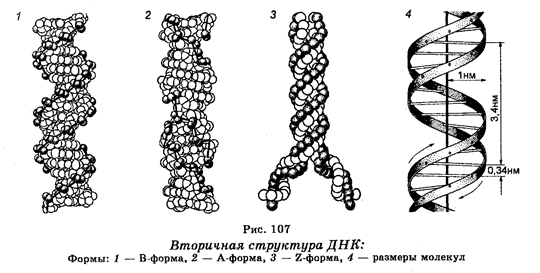
1. Молекула ДНК построена из двух скрученных направо спиралевидных полинуклеотидных цепей, причем каждый виток спирали соответствует 10 азотистым основаниям или расстоянию в 3,4 нм. Молекулы ДНК, цепи которых скручены направо, первоначально назвали В-формой.
2. Обе цепи объединены в результате закручивания одной цепи вокруг другой по общей оси. Из-за противоположной последовательности атомов в каждой цепи обе цепи инвертированы относительно одна другой, т. е. направление вдоль дуплекса есть 3' ® 5' для одной цепи и 5' ® 3' для другой.
3. Сахарофосфатные группы располагаются на внешней стороне двойной спирали, тогда как основания находятся внутри спирали под прямым углом и вдоль ее оси. Диаметр молекулы составляет 2 нм, расстояния между отдельными азотистыми основаниями в молекуле равны 0,34 нм. Таким образом, ДНК представляет собой скрученную в правостороннем направлении двойную спираль, в которой пары азотистых оснований А—Т и Г-Ц в комплементарных полинуклеотидных цепях подобны перекладинам в лестнице, а сахарофосфатные цепи являются каркасом этой лестницы.
4. Цепи в молекуле не идентичны, но комплементарны и удерживаются слабыми водородными связями между азотистыми основаниями, причем спаривание азотистых оснований для связывания цепей имеет специфический характер. Водородные связи устанавливаются не просто между азотистыми основаниями цепей, а специфически между пуриновым азотистым основанием одной цепи и пиримидиновым азотистым основанием другой. В результате этого аденин одной из цепей связывается с тимином другой цепи двумя водородными связями, тогда как гуанин одной из цепей связывается с цитозином, находящимся в другой цепи, посредством трех водородных связей.
Таблица 12
Свойства разных конформационных форм ДНК
| Свойство
|
Формы спиралей
|
| А
|
В
|
С
|
Z
|
| Направление скрученное™
|
вправо
|
вправо
|
вправо
|
влево
|
| Диаметр молекулы
|
23 А̊
|
19 А̊
|
19 А̊
|
18 А̊
|
| Количество оснований в витке
|
11
|
10
|
91/3
|
12
|
Дезоксирибозные остатки пар А-Т и Г-Ц разделены одинаковыми расстояниями. Для сахарофосфатных скелетных связей характерна полярность, поскольку фосфат связывает группу 3'-ОН одной дезоксирибозы с группой 5'-ОН другой, тогда как комплементарные цепи имеют противоположную полярность.
Двойная спираль имеет упорядоченный характер, поскольку каждая связь основание-сахар имеет одинаковое расстояние от оси спирали и перевернута на 36°. Как видно, вторичная структура отражает собой форму нуклеиновой кислоты.
Исследования рентгеновской дифракции молекул ДНК показали, что количество оснований в витках закрученной направо спирали может составлять не только 10, как у В-формы, но и 11, а то и ОУз оснований. Эти формы спиралей получили название А- и С-форм. Установлено также, что в молекулах ДНК встречаются районы, цепи в которых закручены налево. Эти районы получили название Z-форм. Различия между А-, В-, С- и Z-формами приведены в табл. 12, однако степень регулярности и конформации Z-формы еще не выяснена.
Степень суперскручивания ДНК зависит от ферментов, в частности от динамического баланса между взаимоантагонистическими ферментами ДНК — гиразой, которая ответственна за суперскручивание и ДНК — топоизомеразой I, которая устраняет суперскручивание.
Третичная структура ДНК связана с трехмерной пространственной конфигурацией молекул и зависит от внутримолекулярных условий. Однако эта структура достаточно еще не изучена.
Размеры молекул ДНК обычно устанавливают определением молекулярной массы в дальтонах и длины в количестве пар оснований. Молекулярная масса пары А-Т составляет 617 дальтон, пары Г-Ц — 618 дальтон. Молекулярная масса 1000 пар азотистых оснований (1 килобаса) составляет 617 500 дальтон или 6,175´105
/6,02×1023
г = 1,026×10-18
г = 1,026´10-6
пикограммов (пг), 1 пг ДНК = 9,75´105
килобасов = 0,975 х 106
килобасов.
Препараты ДНК, выделяемой из клеток с помощью обычных методов, имеют молекулярную массу порядка 1,0´107
. Длина витка по оси ДНК В-формы равна 34 А̊. Расстояние между парами оснований в ДНК В-формы Е. coli равно 0,34 нм.
Для характеристики строения ДНК используют также такие физические константы, как плотность ее при центрифугировании в градиенте тяжелых металлов, а также температура плавления; первая константа отражает полидисперсность препаратов ДНК, тогда как вторая — их гетерогенность. Нагревание ДНК в растворах разрывает водородные связи между основаниями в парах и разрушает вторичную структуру ДНК, т. е. вызывает плавление ДНК. В 0,1 М раствора NaCI плавление наступает при 95°С.
Плавление ДНК есть ее денатурация. Однако замечательное свойство денатурированной ДНК заключается в том, что она способна к денатурации in vitro, т. е. способна восстанавливать двухцепочечную структуру, причем ренатурация является очень точной. Две цепи денатурированной ДНК могут ренатурировать в природную двухцепочечную спиральную форму, если их последовательности комплементарны или, другими словами, если последовательности цепей позволяют формирование пар оснований, соединенных водородными связями. Ренатурацию можно оценить и в качестве гибридизации ДНК.
Между тем способность самокомплементарных последовательностей к гибридизации и формированию двухцепочечной спирали присуща на только ДНК, но и РНК. В результате этого in vitro можно конструировать двухцепочечные гибридные структуры РНК-РНК или РНК-ДНК. Способность нуклеиновых кислот к ре-натурации имеет значение в изучении специфики отдельных последовательностей, а также в таксономии.
В зависимости от локализации ДНК в клетке различают ядерные (хромосомные) и экстраядерные (экстрахромосомные) детерминанты наследственности. Кроме того, известны транспозируемые генетические элементы (инсерционные последовательности, транспо-зоны и др.).
С учетом эволюционного уровня организмов существует несколько форм организации ядерных генетических детерминантов.
Вирусный геном
. Наиболее простой формой ядерной организации генома вообще является вирусный геном, который, как уже отмечено, очень условно часто называют вирусной хромосомой. Геном самых малых РНК-овых вирусов представлен последовательностями, состоящими из нескольких тысяч нуклеотидов, что соответствует нескольким генам. Например, полная нуклеотидная последовательность РНК-содержащего бактериального вируса MS составляет 3569 нуклеотидов (три гена). Наименьшие по размерам ДНК-содержащие вирусы состоят из большего количества пар нуклеотидов. Например, полная нуклеотидная последовательность фага ФХ174 составляет 5375 пар нуклеотидов (8 генов). Геном более крупных ДНК-овых вирусов, например, фага Т2 и оспенного вируса, состоит из около 150 генов. Геном цитомегаловируса составляет 229 000 азотистых оснований.
Геном прокариот
. У микоплазм, являющихся примером, возможно, самых мелких прокариот, геном составляет всего лишь несколько сот тысяч пар оснований. Например, у Mycoplasma genitalium геном составляет 580 000 пар оснований или 470 кодирующих генетических районов. Однако лишь один генетический район участвует в кодировании белков, тогда как в контроле адгезии (прилипания) к соматическим клеткам поражаемого организма участвует 5% гено-ма. Возможно, это связано с тем, что М. genitalium существует в ассоциации с соматическими клетками человека.
ДНК в клетках Е. соИ представлена одиночной двухцепочечной кольцевой молекулой, м. м- около 2 х 109
, что составляет, примерно, 3 х 106
пар азотистых оснований. Она выполняет роль функционально активной хромосомы, получившей название нуклеоида. Последний является гаплоидной структурой. Поскольку расстояние между парами азотистых оснований в ДНК Е. coli составляет около 3,4 А, то контурная длина молекулы ДНК бактерий этого вида составляет около 0,1 см, что превышает длину содержащей ее клетки примерно в 600 раз, а диаметр лишь 20 А (диаметр одиночной клетки Е. coli равен около 0,75). Поэтому считают, что хромосома (ДНК) внутри бактерий этого вида существует в виде «свернутого генома», занимающего '/в объема клетки, т. е, в свернутой (скрученной) форме в виде около 50 петель, каждая из которых находится в сверхскрученной форме. Поскольку «свернутый геном» можно дестабилизировать обработкой РНК-азой, то считают, что в его состав входит также и РНК. Кроме того, в его составе обнаружены низкомолекулярные белки, роль которых еще не выяснена.
Хромосома Е. coli содержит около 3000-4000 генов, которые организованы на основе принципа колинеарности, означающего, что существует линейное соответствие первичной структуры гена структуре полипептидной цепи, т. е. непрерывность последовательности нуклеотидов сопровождается непрерывностью последовательности аминокислот в полипептидах.
Геном Haemophilus influenzae составляет 1,8 х 10е
пар оснований, которые организованы в 1743 кодирующих районах.
Молекулы ДНК прокариот содержат последовательности азотистых оснований, которые все являются кодирующими. В геноме прокариот каждый ген размером в несколько тысяч пар оснований повторяется лишь один раз в геноме. Исключение составляют гены для рибосомной РНК. Если прокариотическая хромосома разрывается на несколько сегментов, то каждый из них будет содержать различные последовательности.
Геном эукариотов
. Геном эукариотов по своим размерам больше генома Е. coli лишь в 2—10 раз, однако ДНК в нем значительно больше. Установлено, что помимо «работающей» ДНК, т. е. помимо эксо-нов и интронов, в хромосомах клеток эукариот, в частности в гетерохроматиновых районах хромосом содержится много ДНК, которая является излишней, т. к. не транскрибируется. Такую ДНК называют сателлитной или «эгоистической» (от англ. selfish — эгоистичный). Количество некодирующей ДНК у разных организмов является различным. Например, у дрожжей оно составляет 30% от общего количества ДНК, у насекомых — 67%, у цветковых растений (ара-бидопсис) — 69%, у нематод — 75%, у человека — 73-91%.
Примечательной особенностью некодирующей ДНК является то, что определенные последовательности оснований в ней неоднократно повторяются. Такие последовательности называют повторяющимися или просто повторами. Напротив, последовательности ДНК, которые не повторяются, т. е. существуют в одиночных копиях, называют уникальными. Их фракция составляет обычно 40—80%.
Длина повторов составляет обычно от нескольких до десятков или сотен пар оснований, причем у эукариотов повторы, как правило, локализованы в гетерохроматиновых районах хромосом, и их содержание может быть как простым, например 5'-АТАТАТ-3' в одной цепи и 3'-ТАТАТА-5' в другой, так и более сложным, например 5'-ГАААААТГА-3' в одной цепи и 3'-ЦГГГГГАЦТ-5' в другой.
Среди повторяющихся некодирующих последовательностей ДНК различают микросателлитные, минисателлитные и сателлитные последовательности .
Микросателлитные (простые) повторяющиеся последовательности представляют собой короткие повторы (3-5 пар оснований). Они найдены в геномах насекомых, позвоночных и растений. В геноме человека такие последовательности встречаются в эухромати-новых районах в количестве примерно 100 копий на геном.
Минисателлитные повторяющиеся последовательности представлены повторами, состоящими примерно на 15 пар азотистых оснований. Эти повторы обнаружены в геномах многих позвоночных и растений, включая микроскопические грибы.
Сателлитные повторяющиеся последовательности состоят из 5-10 пар азотистых оснований, а иногда даже и из 100 пар. Эти последовательности обнаружены в гетерохроматиновых районах хромосом человека ближе к центромерам или в Y-хромосоме.
Биологическое значение эгоистической ДНК остается неясным, т. к. не выяснена точно необходимость в этих сегментах ДНК и причины, поддерживающие их в составе кодирующей ДНК.
Биологическое значение повторов в «эгоистической» ДНК также не выяснено. Однако предполагают, что они вовлечены в регуляцию экспрессии и рекомбинацию генов, а также в «защиту» некоторых структурных генов, в частности генов, детерминирующих синтез гистонов, рРНК или рибосомных белков.
Для ДНК эукариотических клеток характерно также наличие палиндромов, т. е. обращенных повторов. Они встречаются в огромном количестве копий (возможно, в тысячах копий), причем их длина различна. Функции длинных палиндромов неизвестны. Что касается коротких палиндромов, то они служат сайтами узнавания ферментами-рестриктазами (см. гл. XIX).
Повторяющиеся последовательности реплицируются в геномах благодаря совместной репликации с хромосомами. Реплицируются хромосомы, реплицируются и повторяющиеся последовательности.
Большинство структурных генов представлено однокопийными последовательностями ДНК, т. е. существует в одной копии. Некоторые гены существуют в 1—3 копиях, но известны гены, которые представлены множественными копиями (см. § 58).
Поскольку у эукариотов ДНК содержится в каждой хромосоме, а каждая хромосома представлена в двух (диплоидных) или более (полиплоидных) копиях, то количество ДНК в хромосомах зависит от их плоидности. Например, гаплоидный набор хромосом половых клеток человека содержит ДНК длиной 1000 мм, причем метр этой ДНК разделяется между 23 хромосомами, для которых характерны разные размеры и формы. Следовательно, в каждой хромосоме содержится ДНК длиной от 15 до 85 мм ДНК. Напротив, диплоидный набор хромосом содержит ДНК длиной около 2000 мм.
Исходя из того, что организм человека состоит из около 1013
клеток, можно заключить, что протяженность всей ДНК человека составляет 2´1010
км (расстояние от Земли до Солнца равно 1,44´108
км).
Гетерогенность длины нуклеосомной ДНК определяется вариабельностью в длине ДНК, сцепливающей одну нуклеосому с другой. Какова роль негистоновых белков, обычно обнаруживаемых в хроматине — это вопрос, который подлежит еще выяснению.
В случае прокариотов между генами последовательность азотистых оснований ДНК и полипептидной цепью существует коли-неарность. Однако у эукариотов гены вдоль хромосом располагаются не непрерывно, поскольку они разделены другими последовательностями ДНК. Поэтому колинеарности в случае эукариотов не существует. Кроме того, в геноме эукариотов различают два типа последовательностей — транскрибируемые и транслируемые последовательности, которые определяют первичнуюструктуру белков и называются эксонами, и транскрибируемые, но не транслируемые («молчащие») последовательности, называемые интронами. Следовательно, гены содержат как эксоны, так и интроны (см. гл. XII). Можно сказать, что для генов эукариотов характерна мозаичность, которая имеет место практически на протяжении всего генома.
Длительное время считали, что ДНК содержится только в ядрах клеток, и вся наследственность понималась в качестве ядерной. Между тем с развитием молекулярно-генетических методов исследований стали обнаруживать ДНК, находящуюся за пределами ядра как у прокариотов, так и в клетках эукариотов. Эта ДНК получила название экстраядерной (экстрахромосомной) ДНК, а контролируемую такой ДНК последовательность — экстраядерной или экстрахромосомной.
Перечислим формы экстраядерных (экстрахромосомных) ДНК прокариотов и эукариотов:
1. ДНК плазмид: бактерии, низшие грибы и другие организмы.
2. ДНК органелл: митохондрии, хлоропласты, кинетопласты.
3. ДНК амплифицированных генов: гены, контролирующие синтез отдельных белков.
4. Малые полидисперсные кольцевые и линейные ДНК: экстрахромосомные копии повторяющихся (часто транспозируемых) последовательностей ДНК.
Плазмиды.
Плазмиды встречаются в цитоплазме как прокариотов, так и эукариотов, причем у бактерий они являются обычными обитателями. В частности, они идентифицированы почти у всех видов бактерий, имеющих медицинское (являющихся возбудителями болезней) или сельскохозяйственное и промышленное значение.
Плазмиды бактерий — это генетические структуры, находящиеся в цитоплазме и представляющие собой молекулы ДНК размером от 2250 до 400 000 пар азотистых оснований. Они существуют обособленно от хромосом в количестве от одной до нескольких десятков копий на одну бактериальную клетку. Различают три типа бактериальных плазмид: факторы генетического переноса, коин-тегративные и неконъюгативные плазмиды (рис. 108).
Факторы
переноса обладают лишь генами репликации и переноса. Благодаря генам репликации такие плазмиды способны к бесконечно долгому поддержанию и воспроизводству в автономном (экстрахромосомном) состоянии, а благодаря генам переноса — к передаче  от одних клеток к другим, часто преодолевая в скрещиваниях видовые и родовые барьеры. Бактерии, содержащие плазмиды этого типа, служат генетическими донорами. Они способны вступать в скрещивания с клетками, не содержащими плазмиды. от одних клеток к другим, часто преодолевая в скрещиваниях видовые и родовые барьеры. Бактерии, содержащие плазмиды этого типа, служат генетическими донорами. Они способны вступать в скрещивания с клетками, не содержащими плазмиды.
Коинтегративные
плазмиды представляют собой фактор генетического переноса, сцепленный с генами, контролирующими синтез тех или иных белков, имеющих значение для бактерий. Например, плазмиды R контролируют синтез ферментов, придающих бактериям устойчивость к антибиотикам, сульфани-ламидам и другим лекарственным веществам, плазмиды Ent — синтез энтеротоксинов, Col — колицинов, Hly — гемолизинов. Известны также плазмиды, контролирующие разрушение многих органических соединений и др. свойства. Благодаря фактору переноса эти плазмиды конъюгативны.
Неконъюгативные
плазмиды — это плазмиды, которые не передаются от одних клеток к другим, т. к. они не обладают фактором переноса. Они тоже детерминируют лекарственную устойчивость и другие свойства бактерий. Передача неконъюгативных плазмид от одних бактерий к другим обеспечивается содержащимися в клетках факторами переноса или коинтегративными плазмидами, которые мобилизуют их на перенос. Среди эукариотов плазмиды идентифицированы у низших грибов. Одна из таких плазмид у дрожжей S. cerevisiae представляет собой кольцевые молекулы ДНК размером в 6318 пар оснований, существующие в количестве 80 копий на гаплоидный геном и кодирующие белки, необходимые для собственной репликации и рекомбинации. У нейроспоры (Neurospora) плазмиды обнаружены в виде малых кольцевых молекул ДНК размером 4200-5200 пар оснований, встречающихся в количестве около 100 копий на гаплоидный геном, а у плесени Aspergilus niger — в виде кольцевых молекул ДНК размером около 13 500 пар оснований в количестве около 100 копий на клетку.
ДНК органелл.
ДНК этого класса обнаружена в случае как низших, так и высших эукариотов.
Молекулы ДНК, выделяемые из митохондрий соматических клеток животных и хлоропластов клеток растений, характеризуются небольшими размерами. Например, размеры молекул ДНК (гено-мов) митохондрий (мтДНК) разных животных (включая плоских червей, насекомых и млекопитающих), составляют 15 700—20 000, человека — 16 569 пар азотистых оснований. У простейших, например у трипаносом и парамеций, митохондриальный геном равен 22 000 и 40 000 пар оснований. Геном хлоропластов у высших растений составляет 12 000 — 200 000 пар оснований, у дрожжей — 78 000 пар оснований, у зеленых водорослей — 180 000 азотистых оснований. Во многих случаях показано, что ДНК митохондрий и хлоропластов сплошь состоит из нуклеотидных последовательностей, гомологичных последовательностям хромосомной ДНК.
Митохондриальный геном человека состоит из 13 генов, нукле-отидная последовательность которых определена и для которой характерно полное или почти полное отсутствие некодирующих участков. Эти гены кодируют собственные рибосомные РНК (12S- и 168-рРНК.) и 22 разные транспортные РНК, а также разные поли-пептиды, включая субъединичные компоненты I, II, III оксидазы цитохрома С, субъединицы 6 АТФазы, цитохрома В и девяти других белков, функции которых не известны.
Геном хлоропластов ряда высших растений состоит из 120 генов. Они кодируют 4 рибосомных РНК, 30 рибосомных белков, часть субъединиц хлоропластной РНК-полимеразы, часть белков, содержащихся в фотосистемах I и II, белковые субъединицы АТФ-синтетазы и отдельных ферментов цепи транспорта электронов, а также белковую субъединицу рибулозобисфосфаткарбоксидазы и очень многих тРНК. Хлоропластный геном очень сходен с бактериальным геномом как по организации, так и по функциям. В митохондриальном геноме человека, вероятно, отсутствуют интроны, но в ДНК хлоропластов некоторых высших растений, а также в ДНК митохондрий грибов интроны обнаружены. Считают, что хлоропластные геномы высших растений остаются без изменений примерно несколько миллионов лет. Возможно, что такая древность характерна и для митохондриальных геномов млекопитающих, включая человека.
Характер передачи мтДНК по наследству у разных организмов различен. Например, у дрожжей в результате одинакового вклада мтДНК сливающимися гаплоидными клетками в зиготу митохондриальный геном наследуется потомством от обоих родителей. Между тем показано, что у D. melanogaster и мышей мтДНК передается по материнской линии. По данным посемейного распределения ДНК в больших семьях предполагают, что мтДНК у человека также наследуется по материнской линии. Однако у морских голубых двустворчатых раковин из рода Mytilus она передается как по материнской, так и по мужской линии, причем тип передачи зависит от пола потомства. Женские митохондрий передаются матерями сыновьям и дочерям, тогда как мужские митохондрий передаются отцами сыновьям. Но у этих животных иногда встречается и передача женских митохондрий от отцов к дочерям. У большинства высших растений ДНК хлоропластов тоже наследуется по материнской линии.
ДНК, обнаруживаемая в кинетопластах трипаносом, представлена малыми (2,500 п. о.) и крупными (3700 п. о.) кольцевыми молекулами.
ДНК амплифицированных генов
. Эта ДНК встречается в форме экстрахромосомных кольцевых молекул. Например, когда эука-риотические клетки культивируют в средах с лекарственными веществами, то происходит селекция резистентных клеток с повышенным количеством копий гена, контролирующего резистен-тность. Клетки многих опухолей содержат также экстрахромосомные амплифицированные гены (наряду с хромосомными).
Малые полидисперсные кольцевые и линейные ДНК.
Молекулы ДНК этого типа (мпкДНК) имеют размеры от нескольких сот до десятков тысяч нуклеотидных пар и встречаются как в цитозоле, так и в ядре и митохондриях клеток многих организмов-эукариотов. Эти молекулы ДНК происходят или связаны с ДНК хромосом и органелл. Многие из этих молекул ДНК способны к транспозиции (см. § 45).
Транспозируемые (подвижные, мигрирующие, транслоцируемые) генетические элементы —это сегменты ДНК, способные к перемещению в пределах одного генома или с одного генома на другой.
У прокариотов Транспозируемые генетические элементы представлены сегментами ДНК двух типов — инсерционными последовательностями (IS) и транспозонами (Тп).
Инсерционные последовательности ДНК представляют собой последовательности, состоящие из 768-5000 пар азотистых оснований. Они обнаружены в плазмидах, фагах, бактериальных хромосомах, причем встречаются так часто, что многие исследователи считают их нормальными компонентами бактерий. Инсерционные последовательности (IS1, IS2, IS4, IS5, IS102 и др.) в большинстве своем многокопийны. Они перемещаются с высокой частотой. Их миграция происходит на основе генетической рекомбинации.
Транспозоны организованы значительно сложнее, нежели ин-серционные последовательности (рис. 109). В упрощенном виде можно сказать, что транспозон представляет собой сегмент ДНК, середина которого представлена геном или генами устойчивости, а фланги — инсерционными последовательностями, обеспечивающими его передвижение. Размеры транспозонов — 2000-20 500 пар азотистых оснований. Для транспозонов характерны значительные инвертированные повторы.
Транспозируемые элементы клеток-прокариот перемещаются по маршруту хромосома ® плазмида ® другая плазмида ® хромосома. Перемещение транспозонов обеспечивается специализированным репликативным процессом, который не связан с генерацией экстрахромосомных форм. В экспериментальных условиях любой транспозон можно включить практически в любую плазмиду.
Генетические элементы, сходные с транспозируемыми, существуют также в клетках эукариотов, где они представлены разными повторяющимися последовательностями ДНК. Одни из этих элементов транспозируются в результате повторного включения (реинсерции) в геном продукта реверсивной транскрипции (копии РНК). Такие элементы получили название ретроэлементов. Напротив, другие элементы транспозируются прямо через копии ДНК.
Наиболее известными ретроэлементами являются ретротранс-позоны с короткими терминальными повторами. Такими являются ретротранспозоны I или R2 у дрозофил, Line — у млекопитающих, ingi — у трипаносом. Копии этих ретротранспозонов кодируют белки, необходимые для обратной транскрипции, т. е. их транспозиция осуществляется с использованием РНК в качестве интермедиата. К этой категории ретроэлементов принадлежат также последовательности с длинными терминальными повторами, в частности последовательности copia и gypsi у дрозофил, TY-фактор дрожжей и LI — элементы у млекопитающих. У этих последовательностей повторы достигают 500 пар оснований. Наконец, ретротранспозонами многие считают также последовательности, которые, помимо инсерционной способности, обладают инфицирующими свойствами, например, отдельные ретровирусы (лейкоза птиц, лейкемии млекопитающих, иммунодефицита млекопитающих).
Элементы, которые сходны по транспозиции с транспозоном, транспозируются прямо через копии ДНК, характеризуются инвертированными терминальными повторами и открытыми рамками чтения, кодирующими фермент Транспозоны. К таким элементам принадлежат транспозоны Р и hobo у дрозофил, Ас, ДЗ и Мм — у растений кукурузы, Тс/1 — у нематод, TU — у морских ежей.
В геноме человека найдена последовательность Alu длиной порядка 300 пар оснований и повторяющаяся в 100 000-300 000 копиях на гаплоидный набор хромосом, что составляет около 5% генома человека. Alu-последовательности сходны с прямыми копиями ДНК на молекулах мРНК, ибо они содержат «отрезок» по-лидезоксиаденозина на их 3'-концах, а сходство Alu-последователь-ностей с транспозонами определяется тем, что они фланкированы прямыми повторами 7-20 пар оснований.
В геноме человека открыты тандемно расположенные повторяющиеся последовательности Hinf, составленные из субъединиц длиной 172 и 147 пар оснований, а также транспозоны .Mariner, тоже представленные повторами оснований. Транспозоны Mariner обнаружены, кроме того, в геномах дрозофилы, отдельных членистоногих, нематод и планарий.
Биологическое значение транспозонов заключается, прежде всего, в том, что они являются мутагенами (см. § 47).
Все, что известно в настоящее время о репликации ДНК, выяснено в результате многолетнего экспериментального обоснования основных положений модели структуры и репликации ДНК по Д. Уотсону и Ф. Крику.
Формулируя свою модель, Д. Уотсон и Ф. Крик предположили, что репликация ДНК происходит в несколько последовательных этапов, а именно: а) разрыв водородных связей между двумя полинуклеотидными цепями и разделение последних; б) разматывание по-линуклеотидных цепей; в) синтез вдоль каждой из полинуклеотидных цепей новой цепи с комплементарной последовательностью азотистых оснований (рис. 110), Они предположили далее, что разделение и разматывание полинуклеотидных цепей начинается с одного конца молекулы, продолжается по направлению к другому ее концу и сопровождается одновременно идущим с того же конца молекулы синтезом новых полинуклеотидных цепей. Таким образом, в репликации ДНК каждая полинуклеотидная цепь действует в качестве шаблона для вновь синтезируемой полинуклеотидной цепи, причем шаблон обеспечивает выбор определенных нуклеотидных последовательностей из всех возможных последовательностей. В результате этого каждая новая молекула ДНК состоит из одной старой цепи и одной новой (дочерней), комплементарной старой. Этот способ репликации ДНК получил название полуконсервативной репликации.
Полуконсервативный характер репликации ДНК был доказан М. Месельсоном и Ф. Сталем в 1958 г. в экспериментах, выполненных на Е. coli. Выращивая бактерии в течение первых делений в синтетической среде, содержащей в качестве источника азота 15
N («тяжелый» изотоп), а затем с среде с 14
С («легким» изотопом), они показали, что ДНК бактерий после одной генерации роста имела «гибридную» плотность (15
N/14
C), а после двух генерации по плотности наполовину была «гибридной», наполовину «легкой» (14
С), т.е. состояла из «тяжелых» и «легких» полинуклеотидных цепей.
 У прокариотов репликация ДНК начинается с 0-пункта репликации, составленного примерно 300 нуклеотидами, и продолжается в двух направлениях, образуя репликацион-ную «вилку» (рис.111). Скорость движения «вилки», т. е. скорость полимеризации составляет 500 нук-леотидов в секунду. Удвоение молекулы ДНК происходит за 40 минут. Кроме того, у прокариотов действует механизм «вращающееся кольцо», по которому репликационная вилка двигается вокруг кольца, генерируя цепи, на которых синтезируются комплементарные цепи (рис. 112). У прокариотов репликация ДНК начинается с 0-пункта репликации, составленного примерно 300 нуклеотидами, и продолжается в двух направлениях, образуя репликацион-ную «вилку» (рис.111). Скорость движения «вилки», т. е. скорость полимеризации составляет 500 нук-леотидов в секунду. Удвоение молекулы ДНК происходит за 40 минут. Кроме того, у прокариотов действует механизм «вращающееся кольцо», по которому репликационная вилка двигается вокруг кольца, генерируя цепи, на которых синтезируются комплементарные цепи (рис. 112).
Изучение ферментативного синтеза ДНК in vitro, компонентами которого являются ДНК-полимераза, дезоксирибонуклеозид 5'-трифосфаты всех четырех азотистых оснований, ионы магния и ДНК-«затравка»,
показало, что присутствие всех этих компонентов в смеси сопровождается добавлением мононуклеотидов к растущему концу цепи ДНК, причем они добавляются к 3'-гидроксильному концу «затравочной» последовательности, и цепь растет в направлении от 5'- к 3'-концу (рис. 113). Реакция катализируется ДНК-полимеразой III. После добавления в смесь ДНК-«затравки» синтез ДНК не прекращается даже тогда, когда количество вновь синтезированной ДНК достигает количества ДНК-«затравки». Если же один из компонентов в смеси отсутствует, частота полимеризации снижается во много раз. Отсутствие ДНК-«затравки» полностью исключает реакцию.
Установлено, что для репликации ДНК Е. coli in vitro необходимы белки, детерминируемые генами dna A, dna В, dna С, dna G, ДНК-гираза, а также белок, связывающийся с одиночными цепями ДНК и АТФ. Комплекс репликативных ферментов и белков получил название ДНК-репликазной системы (реплисомы).

 Изучение ферментативного синтеза ДНК in vitro показало также, что копируются обе цепи, но т. к. цепи ДНК в спирали антипараллельны, то синтез (полимеризация) одной цепи происходит в направлении от 5' к 3'-концу, тогда как другой — от 3' к 5'-концу. Синтез цепи в направлении от 5'- к 3'-концу является непрерывным, тогда как синтез в направлении от 3'- к 5'— прерывен, поскольку синтезируются короткие сегменты в направлении от 5' к 3'-концу, которые затем воссоединяются ДНК-лигазой. Короткие сегменты по 1000-2000 нуклеотидов получили название фрагментов Р. Оказаки (рис. 114). Следовательно, рост обеих цепей обеспечивается одной и той же полимеразой. Репликационная вилка асимметрична. Цепь, синтезируемую непрерывно, называют лидирующей, тогда как цепь, синтезируемую прерывно, называют «запаздывающей». «Запаздывание» второй цепи связано с тем, что синтез каждого фрагмента Оказаки осуществляется только тогда, когда в результате продвижения лидирующей цепи откроется необходимый участок цепи-шаблона. Изучение ферментативного синтеза ДНК in vitro показало также, что копируются обе цепи, но т. к. цепи ДНК в спирали антипараллельны, то синтез (полимеризация) одной цепи происходит в направлении от 5' к 3'-концу, тогда как другой — от 3' к 5'-концу. Синтез цепи в направлении от 5'- к 3'-концу является непрерывным, тогда как синтез в направлении от 3'- к 5'— прерывен, поскольку синтезируются короткие сегменты в направлении от 5' к 3'-концу, которые затем воссоединяются ДНК-лигазой. Короткие сегменты по 1000-2000 нуклеотидов получили название фрагментов Р. Оказаки (рис. 114). Следовательно, рост обеих цепей обеспечивается одной и той же полимеразой. Репликационная вилка асимметрична. Цепь, синтезируемую непрерывно, называют лидирующей, тогда как цепь, синтезируемую прерывно, называют «запаздывающей». «Запаздывание» второй цепи связано с тем, что синтез каждого фрагмента Оказаки осуществляется только тогда, когда в результате продвижения лидирующей цепи откроется необходимый участок цепи-шаблона.


У бактерий открыты ДНК-поли-меразы I, II, III. Главной является ДНК-полимераза III, которая отвечает за элонгацию цепей ДНК. Что касается данных ферментов, то ДНК-полимераза I заполняет бреши в запаздывающей цепи, тогда как функция ДНК-полимеразы II не совсем понятна.
В случае синтеза лидирующей цепи у ДНК-полимеразы имеется спаренный 3'-конец, что позволяет начать полимеризацию следующей (новой) цепи. Однако для ДНК полимеразы, синтезирующей «запаздывающую» цепь, необходима «затравка», обладающая спаренным 3'-концом (3'-гид-роксильной группой). Эту затравку в виде коротких сегментов РНК синтезирует из рибонуклеотидтрифосфа-тов ДНК-примаза на ДНК-шаблоне запаздывающей цепи. Данный процесс характерен тем, что предшествующий синтез коротких сегментов
РНК «затравливает» каждую новую инициацию синтеза ДНК. Затем включается ДНК-полимераза, полимеризуя 5'-фосфатдезокси-рибонуклеотидного остатка с 3'-гидроксильным концом цепи РНК, что приводит к нормальному синтезу цепи ДНК. В последующем «затравочная» последовательность РНК удаляется, и брешь заполняется ДНК. Таким образом, роль «затравки» в синтезе фрагментов Оказаки выполняет РНК.
Репликация ДНК эукариотов характеризуется теми же механизмами, что и у прокариотов, хотя скорость полимеризации цепей является меньшей (около 50 нуклеотидов в секунду у млекопитающих). В репликации ДНК эукариотов принимают участие те же ферменты, что и в случае прокариотов. Размеры фрагментов Оказаки здесь составляют 100-200 нуклеотидов.
Раскручивание двойной цепи ДНК происходит с участием трех разных белков, а именно: а) белки, дестабилизирующие спираль (SS В-белки). Они связываются с одноцепочечными ДНК, помогают
ДНК-геликазам раскручивать спираль и обеспечивают протяженный одноцепочечный шаблон для полимеризации; б) ДНК-гелика-зы, раскручивающие ДНК. Они прямо вовлечены в катализирова-ние раскручивания; в) ДНК-гиразы, которые катализируют формирование негативных супервитков в ДНК.
У эукариотов известно пять ДНК-полимераз (a, b, g, в и e), из которых главную роль в репликации играют полимеразы a и d.
a—Полимераза начинает синтез на ведущей (лидирующей) и запаздывающей цепях, поскольку только она обладает «затравочной» активностью. Дальнейшую элонгацию лидирующей цепи осуществляет b-фермент, а «запаздывающей» цепи —e - или d-ферменты. g-фермент, который является митохондриальным, завершает репликацию «запаздывающей» цепи, играя при этом роль, присущую в бактериях ферменту роl I.
Установлен также белок (циклин), который синтезируется в S-фазе клеточного цикла и который также необходим для репликации ДНК.
Спирализацию ДНК после репликации обеслечивают ферменты ДНК-топоизомеразы. Процесс репликации ДНК характеризуется исключительной точностью. Как отмечено выше, фрагменты Оказа-ки, продуцируемые в ДНК у эукариот, имеют длину от 100 до 20 пар нуклеотидов. Это, возможно, связано с тем, что у эукариотов синтез ДНК является более медленным (1 молекула ДНК в минуту) по сравнению с прокариотами (30 молекул ДНК в минуту).
Удвоение хромосом эукариотов является сложным процессом, поскольку включает не только репликацию гигантских молекул ДНК, но также и синтез связанных с ДНК гистонов и негистоно-вых хромосомных белков. Конечным этапом является упаковка ДНК и гистонов в нуклеосомы. Считают, что удвоение хромосом также имеет полуконсервативный характер.
Репликационное поведение хромосом основывается на трех фундаментальных свойствах, а именно: непосредственно репликация, сегрегация хромосом при репликации ДНК и делении клеток, а также репликация и предохранение концов хромосом. 0-пункты репликации существуют в хромосомах (сайты инициации репликации) также организмов-эукариотов, состоящих из определенных последовательностей азотистых оснований, причем являются множественными. Эти пункты получили название автономно реплици-рующихся последовательностей (ars-элементов). Определяя количество репликационных вилок, они удалены один от другого на расстоянии 30 000-300 000 пар азотистых оснований. В результате этого по каждой хромосоме двигается много репликационных «вилок», причем одновременно и независимо одна от другой. Инициацию репликации ДНК обеспечивают белки, связанные с 0-пун-ктом репликации, а также белки — киназы. Последние ответственны за выход ДНК из репликации. Но как действуют эти механизмы — это вопрос, который еще не получил разрешения.
За сегрегацию хромосом в дочерние клетки ответственны центромеры.
В репликации и предохранении концов хромосом имеют значение так называемые теломеры, представляющие собой повторяющиеся последовательности ДНК длиной 5—10 азотистых оснований. Их роль заключается в обеспечении доступа ДНК-полимеразы к концам цепей ДНК. Вновь образованные хромосомы содержат как старые гистоны, так и вновь синтезированные, контроль которых у млекопитающих осуществляется 20 генными блоками, каждый из которых содержит по 5 гистоновых генов.
Однако репликация эукариотической ДНК имеет и существенное отличие от репликации прокариотической ДНК. Когда ДНК эукариотов метят ^-тимидином, а затем экстрагируют из хромосом и изучают методом радиографии, то при этом наблюдают тан-демный порядок радиоактивности. Это свидетельствует о том, что одиночные молекулы ДНК содержат множественные 0-пункты репликации. Например, в ДНК клеток млекопитающих 0-пункты встречаются через каждые 40 000 - 200 000 пар оснований. Экспериментальные данные указывают на то, что репликация хромосом эукариотов происходит в двух направлениях, поскольку реплика-ционные вилки двигаются в двух направлениях из центральных 0-пунктов к репликационным терминусам (пунктам остановки репликации). Сегмент хромосомы, чья репликация находится под контролем одного 0-пункта и двух терминусов, является единицей репликации и ее называют репликоном. Размеры эукариотичес-ких репликонов зависят от вида организмов, но в общем они составляют около 10-100 нм.
Мутации (от лат. mutatio — перемена) — это изменение генов и хромосом, фенотипически проявляющиеся в изменении свойств и признаков организмов. Впервые их описал в 1901 г. голландский ученый Г. Де Фриз (1848-1935). Он же заложил основы и теории мутаций. Процесс образования мутаций во времени и пространстве называют мутагенезом. Мутации характерны для всех живых существ, включая человека. Определяя мутации и мутагенез, следует также определить и такие понятия, как дикий тип организма или гена и мутантный организм (мутант). Под диким типом понимают организмы со свойствами, находимыми в природе. Диким типом обозначают также наборы генов или отдельные гены, также являющиеся природными для организма. Образно говоря, дикий тип — это «справочный» тип, своего рода эталон, с которым сравнивают мутантные организмы и мутантные гены.
Мутантные организмы (мутанты) — это организмы, содержащие в своем геноме одну или несколько мутаций. Мутантные организмы могут отличаться от исходных (организмов дикого типа) по самым различным свойствам — морфологическим, физиологическим, биохимическим и другим. Например, у микроорганизмов мутации сопровождаются изменением формы колоний, питательных потребностей, отношения к лекарственным веществам и т. д. У высших растений мутации сопровождаются изменениями качественных и количественных признаков (рис. 115). У насекомых мутанты отличаются от исходных организмов по форме и окраске тела, крыльев, конечностей, глаз, реакции на свет, серологическим свойствам и т. д. (рис. 116). У млекопитающих мутации также ведут к изменению многих качественных и количественных признаков (рис. 117). У человека мутации приводят к различным отклонениям от нормы и сопровождаются наследственной патологией (см. гл. XIII). В целом можно сказать, что мутации являются вредными, полезными или нейтральными для организмов. Но важно помнить, что мутанты остаются организмом того же вида, что и организм дикого типа, из которого они произошли.
Мутации следует отличать от фенокопий, которые продуцируются факторами среды. Мимикрируя действие генов, они не передаются по наследству. Например, если беременных мышей экспонировать к пониженному атмосферному давлению, то некоторые из особей в пометах таких мышей будут иметь повреждения в мочеполовом тракте, не передающиеся, однако, по наследству. В случае человека фенокопиями можно считать, например, ненаследуемые повреждения скелета и слепоту, вызываемые ядами и другими вредными факторами воздействия.
Мутации возникают на всех стадиях индивидуального развития организмов и поражают гены и хромосомы как в половых клетках, причем до оплодотворения и после оплодотворения (после первого деления оплодотворенных яйцеклеток), так и в соматических, причем в любой фазе клеточного цикла. Поэтому по типу клеток, в которых возникают мутации, различают генеративные и соматические мутации (соответственно).

Генеративные мутации
происходят в генитальных и половых клетках.
Если мутация (генеративная) происходит в генитальных клетках, то мутантный ген могут получить сразу несколько гамет, что увеличит потенциальную способность наследования этой мутации несколькими особями (индивидуумами) в потомстве. Если мутация произошла в гамете, то, вероятно, лишь одна особь (индивид) в потомстве получит этот ген. На частоту мутаций в половых клетках оказывает влияние возраст организма.
Соматические мутации
встречаются в соматических клетках организмов. У животных и человека мутационные изменения будут сохраняться только в этих клетках. Но у растений из-за их способности к вегетативному размножению мутация может выйти за пределы соматических тканей. Например, знаменитый зимний сорт яблок «Делишес» берет начало от мутации в соматической клетке, которая в результате деления привела к образованию ветви, имевшей характеристики мутантного типа. Затем следовало вегетативное размножение, позволившее получить растения со свойствами этого сорта.
По типу наследования различают доминантные, полудоминантные, кодоминантные и рецессивные
мутации. Доминантные мутации характеризуются непосредственным эффектом на организм, полудоминантные мутации заключаются в том, что гетерозиготная форма по фенотипу является промежуточной между формами АА и аа, а для кодоминантных мутаций характерно то, что у гетерозигот A1
A2
проявляются признаки обоих аллелей. Рецессивные мутации не проявляются у гетерозигот.
Если доминантная мутация встречается в гаметах, ее эффекты выражаются непосредственно в потомстве. Многие мутации у человека являются доминантными. Они часты у животных и растений. Например, генеративная доминантная мутация дала начало анконской породе коротконогих овец.
Примером полудоминантной мутации может служить мутационное образование гетерозиготной формы Аа, промежуточной по фенотипу между организмами АА и аа. Это имеет место в случае биохимических признаков, когда вклад в признак обоих аллелей одинаков.
Примером кодоминантной мутации являются аллели IA
и IB
, детерминирующие группу крови IV.
В случае рецессивных мутаций их эффекты скрыты в диплоидах. Они проявляются лишь в гомозиготном состоянии. Примером являются рецессивные мутации, детерминирующие генные болезни человека.
Таким образом, главными факторами в детерминировании вероятности проявления мутантного аллеля в организме и популяции являются не только стадия репродуктивного цикла, но и до-минантность мутантного аллеля.
В зависимости от локализации в клетках различают генные (точечные) и хромосомные мутации (рис. 118).
Генные мутации заключаются в изменениях индивидуальных генов. Поэтому их еще называют точечными мутациями и классифицируют на односайтовые и многосайтовые. Односайтовая мутация затрагивает один сайт, многосайтовая — несколько сайтов генного локуса. Некоторые сайты являются «горячими точками», т. к. в них происходят концентрированные мутации, что связано с наличием в нуклеотидных последовательностях модифицированных оснований. Последние подвергаются частому дезаминированию, а это ведет к изменениям в последовательностях оснований.
Генные мутации классифицируют также на прямые и обратные (реверсивные) мутации, которые одинаково встречаются у организмов всех систематических групп.
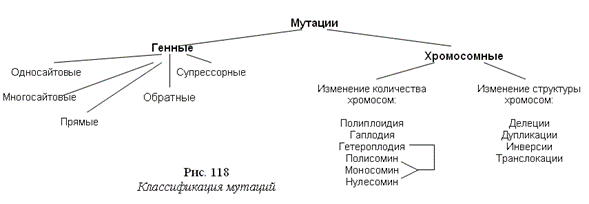
Прямые мутации — это мутации, инактивирующие гены дикого типа, т. е. мутации, которые изменяют информацию, закодированную в ДНК, прямым образом, в результате чего изменение от организма исходного (дикого) типа идет прямым образом к организму мутантного типа.
Обратные мутации представляют собой реверсии к исходным (диким) типам от мутантных. Эти реверсии бывают двух типов. Одни из реверсий обусловлены повторными мутациями аналогичного сайта или локуса с восстановлением исходного фенотипа и их называют истинными обратными мутациями. Другие реверсии представляют собой мутации в каком-то другом гене, которые изменяют выражение мутантного гена в сторону исходного типа, т. е. повреждение в мутантном гене сохраняется, но он как бы восстанавливает свою функцию, в результате чего восстанавливается фенотип. Такое восстановление (полное или частичное) фенотипа вопреки сохранению первоночального генетического повреждения (мутации) получило название супрессии, а такие обратные мутации назвали супрессор-ными (внегенными). Как правило, супрессии происходят в результате мутаций генов, кодирующих синтез тРНК и рибосом.
Большинство генов довольно устойчива по отношению к мутациям, однако известны гены, которые мутируют очень часто. Как правило, мутантный фенотип организмов является одинаковым, независимо от того, наследуется ли генная мутация от отца или матери. Тем не менее небольшое число генов у человека и мышей экспрессируется дифференциально, причем в зависимости от источника наследования мутантного гена (от отца или матери). Во всех случаях один из родительских аллелей полностью инактиви-руется, тогда как другой аллель является активным. Напоминая инактивацию Х-хромосом, это явление получило название генетического импринтинга. Следовательно, генетический импринтинг представляет собой явление неэквивалентности генных аллелей, активность которых зависит от родительского происхождения. Например, патологический синдром Прадела-Вилли у человека обязан мутации, которая встречается только в отцовском хромосомном гомологе, а синдром Ангельмана у человека возникает в результате мутации в материнском хромосомном гомологе.
Хромосомные мутации связаны с изменениями числа и структуры хромосом. Изменения в числе хромосом определяются добавлением или уменьшением всего набора хромосом, ведущим к полиплоидии или гаплоидии (соответственно), часто называемые геномными мутациями, а также добавлением или удалением одной или больше хромосом из набора, что ведет к гетероплоидии или анеуплоидии (моносомии, трисомии и др. полисемии), тогда как изменения в структуре хромосом определяются перестройками (аберрациями) их структуры.
Полиплоидия — это хромосомная мутация в виде увеличения числа полных гаплоидых наборов хромосом. Известны триплоидия (Зп), тетраплоидия (4п), пентаплоидия (5п) и т. д. Наиболее часто полиплоидия встречается у растений, поскольку для них характерны гермафродитизм и апомиксис. Почти третья часть всех видов диких цветковых растений представлена полиплоидами. Типичными полиплоидами являются виды пшеницы, у которых соматические числа 2n = 14,28 и 42, при основном гаметном числе n = 7, картофель, табак, белый клевер, люцерна и другие растения. Родственные виды, наборы хромосом которых представляют ряд возрастающего увеличения (кратного) основного числа хромосом, составляют полиплоидные ряды.
Полиплоидные растения с нечетными наборами хромосом характеризуются пониженной плодовитостью и снижением уровня количественных признаков. Напротив, для растений-полиплоидов с четными наборами хромосом характерен повышенный уровень количественных признаков.
Полиплоиды, происходящие от диплоидных организмов аналогичного вида, называют автополиплоидами. Примером автопо-липлоидов служат растения мягкой пшеницы, у которых 2п == 42 и которые являются гексаплоидами. Искусственные полиплоиды, полученные из гибридов диплоидных растений, относящихся к далеко отстоящим друг от друга видам, называют аллополипло-идами.
В природе встречаются как автополиплоиды, так и аллополип-лоиды, однако географическое распределение полиплоидов обычно отличается от распределения их диплоидиых «родственников». Например, флора о. Шпицберген содержит очень высокий процент полиплоидных рядов, тогда как в других местах количество их меньше по сравнению с диплоидами.
Полиплоидия растений имеет хозяйственную ценность (повышенные размеры плодов, большая сахаристость, лучшая сохранность и др.). По этой причине полиплоиды используют в селекционной работе для получения новых сортов сельскохозяйственных растений.
У животных полиплоидия очень редка. Она встречается у земляных червей, размножающихся партеногенетически (обнаружены полиплоидные ряды с основными числами 11, 16, 17, 18 и 19 хромосом), у некоторых членистоногих, рыб и земноводных. В частности, она описана у морских креветок Artemia salivana. Женские особи саламандр отдельных видов, которые имеют крупные эритроциты с ядрами, продуцируют триплоидные личинки с 42 хромосомами, тогда как саламандры с малыми ядерными эритроцитами продуцируют диплоидные личинки с 28 хромосомами. Все тихоокеанские лососевые рыбы являются полиплоидами.
Сирийский хомячок (Mesocricetius aurantus), у которого 2 n = 44, является аллополиплоидом, возникшим в результате естественной гибридизации между европейским хомяком (Cricetus cricetus, 2n == 22) и хомяком, принадлежащим к одному из азиатских видов (Cricetus griseus, 2n = 24).
Полиплоидия встречается также у человека в пренатальном периоде развития. В частности известно много сообщений об обнаружении триплоидии и тетраплоидии в клетках абортусов. Имеющиеся данные позволяют считать, что триплоидия встречается у 20% абортусов, а тетраплоидия — у 6% абортусов.
Редкость полиплоидии у раздельнополых животных, по-видимому, определяется тем, что она нарушает нормальные соотношения аутосом и половых хромосом в зиготах.
Кроме авто- и аллополиплоидии, в соматических клетках ряда многоклеточных организмов установлена эндополиплоидия, характеризующаяся увеличением числа хромосом в покоящемся ядре (при отсутствии митоза). От полиплоидии следует отличать псевдополиплоидию отдельных растений и насекомых, возникающую в результате однократного или многократного деления компонентов генома, когда центромеры имеют диффузный характер.
Гаплоидия — это мутация в виде уменьшения всего набора хромосом. Она найдена в основном также у растений. Известны гапло-иды свыше 800 видов растений, включая такие культуры, как пшеница и кукуруза. У животных она очень редка, а у человека совсем неизвестна.
Мутации в виде нарушений нормального количества хромосом из-за добавления или удаления одной или более хромосом в какой-то хромосомной паре называют гетероплоидией или анеуп-лоидией. Среди гетероплоидов известны полисемии, когда какая-то пара хромосом становится большей на один (трипликатом) и более гомологов, моносомии, когда какая-либо хромосомная пара теряет один гомолог, и нулесомии, когда теряется вся хромосомная пара. Эти мутации широко распространены как у животных, так и у растений. В частности трисомии и моносомии обнаружены у человека, собак и других животных, а также у многих плодовых, зерновых и овощных растений. Часто трисомии, как и моносомии, обнаруживают в клетках абортированных эмбрионов или плодов человека. Гетероплоидия сопровождается многими фенотипическими эффектами, которые, например, у человека неблагоприятны для его здоровья.
Мутации, поражающие структуру хромосом, называют хромосомными перестройками
или чаще аберрациями. Эти мутации классифицируют на внутрихромосомные и межхромосомные перестройки. Они возникают как у животных, включая человека, так и у растений.
 Внутрихромосомными перестройками являются делении (нехватки, дефишенсы), дубликации, инверсии, тогда как межхромосомные перестройки представлены транслокациями (рис. 119). Внутрихромосомные перестройки в зависимости от «разлома» плеч хромосом бывают парацентрическими («разлом» затрагивает одно хромосомное плечо) и перицентрическими («разлом» проходит по обе стороны центромеры, т. е. захватывает оба плеча). Межхромосомные перестройки обычно захватывают две или более негомологичные хромосомы. Степень изменений бывает различной и часто доходит до изменения группы сцепления. Внутрихромосомными перестройками являются делении (нехватки, дефишенсы), дубликации, инверсии, тогда как межхромосомные перестройки представлены транслокациями (рис. 119). Внутрихромосомные перестройки в зависимости от «разлома» плеч хромосом бывают парацентрическими («разлом» затрагивает одно хромосомное плечо) и перицентрическими («разлом» проходит по обе стороны центромеры, т. е. захватывает оба плеча). Межхромосомные перестройки обычно захватывают две или более негомологичные хромосомы. Степень изменений бывает различной и часто доходит до изменения группы сцепления.
Делеции
представляют собой потери сегмента хромосомы, несущего один или несколько генов. Они являются наиболее частой и опасной для человека формой генетических макроповреждений. Большие делеции заключаются в потере одного или нескольких генов или даже блоков генов. У гаплоидных организмов крупные делеции детальны. Эффект делеции у диплоидных организмов зависит от количества делегированных генов, количественных потребностей в продуктах пораженных генов, позиции генов среди функционально координированных групп и других факторов. Гомозиготные делеции для диплоидных клеток или организмов детальны.
Дупликации
(добавления) представляют собой добавления (удлинения) какого-либо сегмента хромосомы, несущего один или несколько генов, в результате того, что один и тот же сегмент хромосомы может быть повторен в процессе биосинтеза несколько раз. Этот повтор может быть малым, касаясь одиночного гена, или большим, затрагивая большое количество генов. Дупликации часто безвредны для их носителей. Предполагают, что они способствуют формированию полигенов или являются способом введения новых генов в геномы. Некоторые дуп-ликации, однако, вредны и даже детальны (см. гл. IX).
Инверсии
заключаются в поворотах на 180° сегментов, освобождающихся в результате парных разрывов в хромосомах. Если инвертированный сегмент не содержит центромеру, эту мутацию называют парацентрической инверсией, если же такой сегмент содержит центромеру, такую мутацию называют перицентрической. Инверсии оказывают влияние на мейоз, что приводит к пониженной фертильности гибридов. Описаны отдельные наследственные аномалии, вызываемые этой мутацией (см. гл. XIII).
Транслокация
— это обмен частями (сегментами) гомологич-ной и негомологичной хромосом, образованными разрывами по длине последних. Транслоцируемые сегменты могут иметь разные размеры — от небольших до значительных.
Хромосомные перестройки, как и гетероплоидия, также сопровождаются различными фенотипическими эффектами. У человека они ведут к нарушению здоровья. У растений они ведут к изменению их продуктивности.
Однако, обсуждая значение хромосомных перестроек, нельзя не заметить, что для отдельных организмов они безразличны. Например, у пионов, дурмана и других растений присутствие в гено-ме транслокаций является нормальным явлением.
В зависимости от происхождения различают спонтанные
и индуцированные
генные и хромосомные мутации, которые возникают у организмов независимо от уровня их организации.
Спонтанными
называют те мутации, которые возникают у организмов в нормальных (природных) условиях на первый взгляд без видимых причин, тогда как индуцированными называют те мутации, которые возникают в результате обработки клеток (организмов) мутагенными факторами. Главное отличие спонтанных мутаций от индуцированных заключается в том, что мутация может возникнуть в любой период индивидуального развития. Что касается случайного характера мутаций в пространстве, то это означает, что спонтанная мутация произвольно может поразить любую хромосому или ген.
Длительное время считали, что спонтанные мутации являются беспричинными, однако теперь по этому вопросу существуют другие представления, сводящиеся к тому, что спонтанные мутации не являются беспричинными, что они являются результатом естественных процессов, протекающих в клетках. Они возникают в условиях природного радиоактивного фона Земли в виде космического излучения, радиоактивных элементов на поверхности Земли, радионуклидов, инкорпорированных в клетки организмов, которые вызывают эти мутации или в результате ошибок репликации ДНК. Факторы естественного радиоактивного фона Земли вызывают изменения в последовательности оснований или повреждения оснований подобно тому, как это имеет место в случае индуцированных мутаций (см. ниже).
Ошибки репликации ДНК заключаются в том, что какой-либо нуклеотид ошибочно включается в ДНК при ее репликации и при наличии недостатка в «редактирующем» механизме, что исключает коррекцию ошибок. Ошибки в репликации ДНК, т. е. «неаккуратность» ДНК-полимеразы оказывают влияние на частоту спонтанных мутаций. Ошибки репликации ДНК могут быть также связаны с химической нестабильностью нуклеотидов, сопровождающейся изменением спаривающей способности азотистых оснований. Например, цитозин может быть дезаминирован в урацил, который затем распознается ДНК-полимеразой как тимин в течение репликации ДНК.
Определение частот спонтанных мутаций организмов разных видов проводят с помощью разных способов, один из которых связан с определением частоты таких мутаций на репликацию пары азотистых оснований в молекулах ДНК. Этот способ оказался предпочтительным в случае бактерий и других низших организмов.
Определенные к настоящему времени частоты мутаций на репликацию пары оснований и общие частоты генных мутаций у разных организмов показаны в табл. 13.
В случае человека частоты спонтанных мутаций определяют измерением прямых мутаций в пределах разных генов, которые очень чувствительны к мутациям независимо от того, являются ли условия для организмов ограничивающими или селективными.
Рассмотрим конкретный пример определения частоты спонтанных мутаций, например, среди родившихся за один год 242 257 детей 7 оказались больными ахондоплазией. Следовательно, 7 : 242 257´1 : 2 (два аллеля на зиготу) = 1,4´10-6
. Таким образом частота ахондоплазии составляет 1,4 х 10-5
. Одни гены вообще устойчивы к спонтанным мутациям, другие спонтанно мутируют чаще, третьи так часто, что их носители являются мозаиками мутантных (мутировавших) и немутантных (немутировавших) генов.
Средние частоты мутаций по многим генам у человека и домашних животных составляют примерно 1´10-9
, что значительно выше частоты мутаций микроорганизмов. Больше того, между частотами спонтанных мутаций по разным генам человека или домашних млекопитающих существуют значительные различия, достигающие 100 раз, а то и более. Подлинные причины этих различий неизвестны, хотя для их объяснения и предложено несколько гипотез. Одна из них заключается в том, что наиболее чувствительны к мутациям гены больших размеров, поскольку в них содержится много азотистых оснований и существует большая вероятность мутации отдельных из них. По другой гипотезе наиболее чувствительными к мутациям являются гены, располагающиеся в районах хромосом, являющихся «горячими» точками.
Таблица 13
Частоты спонтанных мутации разных организмов
| Организм
|
Количество пар оснований на геном
|
Частота мутаций на репликацию пары оснований
|
Общая частота мутаций
|
| Фаг Т4
|
1,8 х 106
|
1,7 х 10-3
|
3,0 х 10-3
|
| Е. coli
|
4,5 х 10е
|
2,0 х 10-10
|
0,9 х 10-3
|
| N. crassa
|
4,5 х 107
|
0,7 х 10-11
|
2,9 х 10-4
|
| D. melanogaster
|
2,0 х 108
|
7,0 х 10-10
|
0,8 х 10-3
|
Уже давно установлены гены, которые оказывают влияние на мутабельность других генов. Такие гены получили название мута-торных генов. Они содержатся в геноме организмов почти всех изученных генетически к настоящему времени видов.
Определение частот спонтанных мутаций позволяет определить вероятность мутаций в каждом новом поколении людей: 1´10-6
мутаций на ген х 5 х 10~4
генов (гаплоидный геном) = 5´10-2
мутаций на гамету (5 : 100 или 1 : 20). Далее, 1 : 20´2 гаметы на зиготу = 1 : 10 случаев, что каждая гамета несет новую мутацию. Возможно, что это очень высокая частота, но ее достоверность определяется тем, что большинство мутаций рецессивно и, следовательно, не экспрессируется у гетерозигот.
Частоту индуцированных мутаций определяют сравнением клеток или популяций организмов, обработанных и необработанных мутагеном. Если частота мутации в популяции повышается в результате обработки мутагеном в 100 раз, то считают, что лишь один мутант в популяции будет спонтанным, остальные будут индуцированными .
Индуцированными являются те мутации, которые возникают после обработки клеток (организмов) мутагенными факторами. Различают физические, химические и биологические мутагенные факторы. Большинство этих факторов либо прямо реагирует с азотистыми основаниями в молекулах ДНК, либо включается в нук-леотидные последовательности.
Среди физических мутагенов различают ионизирующую радиацию и ультрафиолетовое излучение (УФ), входящие в ту часть электромагнитного спектра, который содержит волны более короткие и с большей энергией, чем видимый свет (ниже 0,1 нм). УФ-излучение составляет значительную часть солнечного спектра. Ионизирующие излучения и УФ-излучения ответственны примерно за 10% всех встречающихся в ДНК индуцированных повреждений.
Ионизирующие излучения — это рентгеновское излучение (Х-лучи), протоны и нейтроны космических лучей, а также a-, b- и g-лучи, освобождаемые радиоактивными элементами изотопов (плутония, 23
Р, 14
С, 3
H, кобальта-90 и др.). Источником ионизирующего излучения также являются радиоактивные отходы ядерных реакторов.
Ионизирующие излучения обладают высокой энергией и могут проникать в ткани, в которых сталкиваются с атомами и вызывают освобождение электронов, оставляя позитивно заряженные свободные радикалы или ионы. В свою очередь эти ионы сталкиваются с другими молекулами, вызывая дальнейшее освобождение электронов. Поэтому вдоль трека каждого высокоэнергетического луча формируется стержень ионов, проходящий в живые ткани. По этой причине рентгеновское и другие виды излучений называют ионизирующим излучением.
Мутагенный эффект ионизирующей радиации впервые был показан в нашей стране на микроорганизмах Г. А. Надсоном и Г. С. Филипповым в 1925 г. Позднее он был показан в США на дрозофиле Г. Д. Мёллером (1890-1967), а затем и на многих других организмах. Этот эффект вызывается повышенной реактивностью атомов, присутствующих в ДНК. Ионизирующие излучения индуцируют генные мутации (транзиции, трансверсии, делеции, включения), а также хромосомные разрывы, сопровождающиеся транслокациями и другими аберрациями. В случае острого облучения погибает большинство сперматогоний, но сперматоциты выживают, в результате чего в первые 6 недель после облучения происходит снижение фертильности, за которым следует бесплодие (2—3 месяца). Должна быть обеспечена защита в течение нескольких недель до и после зачатия.
Большую опасность представляет рентгенодиагностика и рентгенотерапия брюшной полости и области таза. Поэтому зачатие в течение нескольких недель до и после облучения должно быть исключено.
Для человека удваивающая доза ионизирующего излучения по генным мутациям составляют 1 грей, по хромосомным аберрациям (транслокациям) — 0,15 грея. Характерной особенностью ионизирующего излучения является также то, что для него отсутствует пороговость в дозе, а так же то, что она обладает коммулятивным эффектом.
УФ-излучение характеризуется меньшей энергией, проникает только через поверхностные слои клеток животных и растений и не вызывает ионизации тканей. Обычно различают три вида УФ-излу-чения, а именно:
а) УФ-излучение с длиной волны 180-290 нм. Это излучение не найдено в солнечном свете, т. к. адсорбируется озоновым слоем. Оно продуцируется искусственно бактерицидными лампами, используемыми для стерилизации.
б) УФ-излучение с длиной волны 290-320 нм. Это основная фракция солнечного света. Она мутагенна.
в) УФ-излучение с длиной волны 320 нм. Оно также обладает повреждающим эффектом.
Мутагенные эффекты УФ-излучения связаны с повышенной реактивностью атомов, присутствующих в молекулах ДНК. Оно не опасно для половых клеток человека, поскольку поглощается кожей, но опасно для последней, т. к. вызывает образование в клетках кожи тиминовых димеров, мутагенный эффект которых заключается в том, что они вызывают мутации не прямо, а нарушением точности репликации.
Химическими мутагенами являются органические и неорганические кислоты, щелочи, перекиси, соли металлов, этиленамины, формальдегид, фонолы, акридиновые красители, алкилирующие соединения, аналоги пуриновых и пиримидиновых оснований и др. Считают, что для действия химических мутагенов присуща пороговость. Одни химические мутагены действуют как на реплициру-ющуюся, так и покоящуюся ДНК, тогда как другие только на реп-лицирующуюся ДНК. Примером мутагенов, вызывающих изменения и в реплицирующейся, и в покоящейся ДНК, являются алкилирующие соединения (метилметансульфонат, этилметансуль-фонат и нитрозогуанидин) и азотистая кислота. К мутагенам, действующим на реплицирующуюся ДНК, относят аналоги азотистых оснований и акридиновые красители.
Многие химические мутагены нарушают мейоз, что приводит к нерасхождению хромосом, а также вызывают разрывы хромосом и генные мутации. Например, трипофлавин действует на все стадии развития половых клеток, нитрозогуанидин — перед мейозом, а тренинон — после мейоза.
Некоторые из химических немутагенных соединений становятся мутагенами, попав в организм, как например, циклофос4эамид.
Заслуживают внимания химические вещества, используемые в качестве лекарственных соединений. Так после лечения алкилирующими соединениями необходимо избегать зачатия в первые три месяца. Известны сведения о мутагенности оральных (применяемых внутрь) химических контрацептивов, которые получили очень большое распространение в последние десятилетия, а также некоторых соединений, входящих в косметические средства и в консерванты продуктов питания.
Биологическими мутагенами являются вирусы, которые вызывают хромосомные аберрации в культивируемых клетках. Такой способностью обладает, например, вирус гриппа. Транспозируемые генетические элементы также способны вызывать генные и хромосомные мутации.
В экспериментальной работе используют разные способы получения сайтонаправленных индуцированных мутаций, т. е. мутаций, затрагивающих интересующие исследователя участки ДНК. В частности, широко используют мутагенез in vitro клонируемой ДНК, для этого последнюю обрабатывают нуклеазами (рестриктазами) или химическими мутагенами. Кроме того, известны методы мута-генеза химически синтезируемой ДНК. Наконец, возможно получение в стволовых эмбриональных клетках экспериментальных животных хромосомных аберраций генноинженерным методом.
Механизмы полиплоидии
заключаются в том, что они являются результатом извращений одного или более митотических делений клеток зародыша или результатом нерасхождения в период мейоза всего набора хромосом, ведущего к образованию диплоидных гамет. Нерасхождение хромосом у женщин имеет место в 80% случаев, а у мужчин оно наблюдается в 20% случаев, причем оно отмечается как в первом, так и во втором мейотических делениях.
Механизмы гетероплоидии
также связаны с нерасхождением хромосом. В частности установлено, что, например, у человека три-сомии обеспечиваются нерасхождением хромосом как в первом, так и во втором мейотическом делении.
Механизмы хромосомных аберраций не совсем ясны. В любом случае они связаны с разрывами хромосом, как у растений, так и у животных, что ведет к изменению последовательности хромосомных генов.
Молекулярные механизмы генных мутаций заключаются в изменениях последовательности азотистых оснований в молекулах ДНК. Эти изменения происходят в результате замен, делеций (выпадений), включений и дупликаций оснований, что ведет к изменению содержания кодонов.
Изменения, связанные с заменой оснований в молекулах ДНК, классифицируют на простые и перекрестные замены (табл. 14).
Простые замены, или транзиции, заключаются в замене одного пурина на другой пурин, и наоборот, в двухцепочечной молекуле ДНК — пары А-Т на пару Г-Ц, и наоборот. Транзиции осуществляются в процессе репликации ДНК без изменения ориентации пар пурин-пиримидин в двухцепочечной молекуле ДНК, но при этом происходят изменения в содержании кедонов.
Перекрестные замены, или трансверсии, связаны с заменой в ДНК пурина на пиримидин, и наоборот. Замещающий пиримидин спаривается с пурином, так что в двухцепочечной молекуле ДНК вместо пары пурин-пиримидин оказывается пара пиримидин-пурин. Следовательно, трансверсии приводят к новым ориентациям пар пурин—пиримидин и заключаются в замене в двухцепочечной молекуле ДНК пары А-Т на пару Ц-Г, и наоборот, пары А-Т на пару Т-А, и наоборот, пары Т-А на пару Г-Ц, и наоборот, а также пары Г-Ц на пару Ц-Г, и наоборот.
Таблица 14
Типы замен оснований в молекулах ДНК
| Исходное
основание
|
Основание, занявшее место исходного
|
Тип замены
|
| Пурин
|
Другой пурин
|
Простая замена (транзиция)
|
| Пиримидин
|
Другой пиримидин
|
То же
|
| Пурин
|
Любой пиримидин
|
Перекрестная замена (трансверсия)
|
| Пиримидин
|
Любой пурин
|
Перекрестная замена (трансверсия)
|
Спонтанные замены азотистых оснований происходят очень редко. Например, в соответствии с существующими расчетами у человека за год случается около 10—20 спонтанных замен оснований, причем одна замена может быть повторена на каждые 10 000 генов лишь 50 раз на протяжении времени в 1 млн лет. Можно полагать, что такая чрезвычайно низкая частота замен оснований в ДНК присуща как животным (млекопитающим), так и растениям. Спонтанные замены азотистых оснований возникают в ДНК в результате «ошибок», совершаемых ДНК-полимеразой и сопровождающихся неправильным спариванием оснований. Одно из объяснений этой «ошибочности» было дано Д. Уотсоном и Ф. Криком еще в 1953 г. и оно сводится к признанию в ошибочном спаривании роли тауто-мерных форм (структур, в которых протон перешел на место, противоположное обычной водородной связи) естественных оснований. Следовательно, структурные основы для мутаций в виде замен оснований обеспечивают таутомеры естественных оснований.
 Транзиции индуцируются азотистой кислотой, которая вызывает окислительное дезаминирование аденина, цитозина и гуанина, содержащих свободные аминогруппы, в гипоксантин, урацил и ксантин соответственно. Из-за того, что дезаминирование сопровождается переходом аминооснования в кетонооснование, гипоксантин, например, подобно гуанину, будет спариваться с цитози-ном, т. е. в результате дезаминирования аденина в гипоксантин пара А-Т перейдет в пару Г-Ц. В случае дезаминирования цитозина в урацил пара Г-Ц перейдет в пару А-Т. Транзиции индуцируются также алкилирующими соединениями. Например, этилметан-сульфонат алкилирует гуанин и освобождает от него ДНК без нарушения ее сахарофосфатного каркаса. Следовательно, гуанин может быть заменен любым основанием, и это ведет не только к транзициям, но и к трансверсиям. Транзиции индуцируются азотистой кислотой, которая вызывает окислительное дезаминирование аденина, цитозина и гуанина, содержащих свободные аминогруппы, в гипоксантин, урацил и ксантин соответственно. Из-за того, что дезаминирование сопровождается переходом аминооснования в кетонооснование, гипоксантин, например, подобно гуанину, будет спариваться с цитози-ном, т. е. в результате дезаминирования аденина в гипоксантин пара А-Т перейдет в пару Г-Ц. В случае дезаминирования цитозина в урацил пара Г-Ц перейдет в пару А-Т. Транзиции индуцируются также алкилирующими соединениями. Например, этилметан-сульфонат алкилирует гуанин и освобождает от него ДНК без нарушения ее сахарофосфатного каркаса. Следовательно, гуанин может быть заменен любым основанием, и это ведет не только к транзициям, но и к трансверсиям.
Транзиции часто вызываются мутагенами, действующими на ДНК только в состоянии репликации, например, 5-бромурацилом, который является аналогом тимина и способен включаться в ДНК посредством замещения тимина. Наряду с нормальной способностью 5-бромурапила спариваться с аденином иногда возникает состояние, когда он действует не как тимин, а как цитозин, что обеспечивает формирование водородных связей его не с аденином, а с гуанином. Эти «ошибки» спаривания происходят либо при включении 5-бро-мурацила в ДНК («ошибки» включения), либо при репликации ДНК после его включения («ошибки» репликации). Следовательно, время «ошибок» определяет характер транзиции. «Ошибки» спаривания, индуцируемые 5-бромурацилом, ведут к транзициям от пары Г-Ц к паре А-Т, и наоборот (от А-Т к Г-П). Подобные транзиции индуцируются также 2-аминопурином.
Замены оснований приводят к изменениям смысла кодонов, вследствие чего они приобретают способность кодировать другую аминокислоту (миссенс-мутации). Например, замена в триплете ГУА, содержащемся в гене р-гемоглобина, урацила на аденин (трансверсия) сопровождается тем, что в цепи р-гемоглобина вместо валина оказывается глутаминовая кислота. Это ведет к превращению гемоглобина в новый вариант мутантного гемоглобина (например, типа Бристоль). В результате замен оснований возникают также нонсенс-мутации, когда на измененных кодонах обрывается чтение информации гена (как правило, такими кодонами являются триплеты УАГ, УАА и УГА). Одновременно в результате замен образуются кодоны, сохраняющие исходный смысл.
Делеции и включения одного или нескольких азотистых оснований в нуклеотидных последовательностях ДНК могут быть ошибками репликации ДНК или индуцироваться акридиновыми красителями. Такие изменения называют мутациями сдвига рамки, ибо они приводят к сдвигу «рамки чтения» кода гена. Включаясь между соседними основаниями, акридин оранжевый заставляет их «раздвигаться» на расстояние в 0,6-0,8 нм.
Если акридин оранжевый присутствует в полинуклеотидной цепи-шаблоне, то результатом будет добавление основания в новую цепь в процессе репликации ДНК. Если же акридин оранжевый присутствует в клетке во время репликации ДНК, то он может включаться в новую цепь вместо основания, имитируя парное (противоположное) основание в цепи-шаблоне, и затем выйти. Это приводит к тому, что вновь реплицированной цепи будет недоставать основания, т. е. она будет реплицирована с делецией по основанию- Делеции могут затрагивать несколько оснований. Например, описаны делеции 15 оснований, которые сопровождались утратой в белке 5 аминокислот.
Дупликации (добавление) 1—2 оснований могут приводить также к мутациям со сдвигом «рамки считывания» кода. Если дупли-кация происходит внутри гена, то «рамка считывания» нарушается на большом протяжении.
Делеции и дупликации азотистых оснований представляют собой молекулярный механизм и мутации митохондриальной ДНК человека. Установлено, что из мтДНК человека могут быть делегированы сегменты длиной до 5000 пар оснований.
Особую форму молекулярных механизмов генных мутаций представляют повторы триплетов азотистых оснований. Наличие в молекулах ДНК повторов триплетов оснований сопровождается нарушениями нормального цикла репликации ДНК, с одной стороны, и аномальным синтезом белка (из-за повторов аминокислоты, кодируемой повторяющимся триплетом), с другой стороны. Например, мутации гена, контролирующего белок хантингтан, недостаток которого у человека сопровождается болезнью Хантингтона, заключаются в резком увеличении повторов триплета ЦАГ.
Мутагенные и летальные эффекты мутагенов сопровождаются структурными повреждениями, которые они вызывают в молекулах ДНК. Например, в геноме человека непрерывно происходят случайные изменения (повреждения), но сохраняются лишь отдельные из них. Причем очень редко. Так из 1000 замен азотистых оснований лишь одна приводит к мутациям. Причина заключается в том, что эти повреждения часто подвержены восстановлению. Процесс реконструкции повреждений ДНК называют восстановлением или репарацией ДНК.
Характер и механизмы исправления повреждений наиболее полно изучены в случае повреждений, индуцированных УФ-излучени-ем. Клетки реагируют на УФ-излучение тем, что в их ДНК образуются повреждения, главные из которых представляют собой фотохимические изменения в пиримидиновых основаниях, переходящие в пиримидиновые димеры, в частности в тиминовые. Последние образуются за счет ковалентного связывания соседних тиминовых оснований в одной и той же цепи молекулы посредством присоединения углерода одного тимина к углероду другого тимина. Помимо тиминовых димеров в ДНК облученных клеток происходит формирование также цитозин-тиминовых и цитозин-цитозиновых димеров, однако частота их является меньшей. Димеризация фланкирующих оснований в гене сопровождается ингибированием транскрипции и репликации ДНК. Она ведет также к мутациям. В результате этого клетка может погибнуть или подвергнуться малигнизации.
Один их механизмов восстановления повреждений ДНК действует у многих видов организмов, включая человека, и состоит в том, что экспонирование в видимом свете клеток, предварительно обработанных УФ-излучением, приводит к снижению летального эффекта в несколько раз, т. е. к фотореактивации функций облученных клеток. Это реактивирующее действие видимого света связано с расщеплением (мономеризацией) пиримидиновых димеров, причем этот процесс обеспечивается светозависимыми фотореактивирующими ферментами. Второй механизм удаления димеров пиримидиновых оснований из ДНК облученных клеток получил название темновой репарации или вырезания-восстановления. Так же, как и фотореактивация, он представляет собой ферментативный процесс, но более сложный, притом проходящий в темноте (рис. 121). Этот механизм заключается в том, что тиминовые димеры с помощью ДНК-репариру-ющих нуклеаз, осуществляющих гидролиз фосфодиэфирных скелетных связей между нуклеотидами с повреждениями (со стороны 5' от тиминового димера) и нормальной частью молекулы ДНК, подвергаются «вырезанию» из цепи ДНК, в которой после этого остаются бреши. Затем происходит «залатывание» брешей восстановительным синтезом ДНК при участии ДНК-полимеразы, связывающейся с 3'-концом поврежденной цепи ДНК, и использовании противоположной (нормальной) цепи в качестве шаблона. Удаление пиримидиновых димеров из ДНК облученных клеток путем «вырезания» и «залатывания» брешей завершается смыканием вновь реплицированного участка ДНК с соседними поврежденными участками и «замазыванием» («сшиванием») сахарофосфатных скелетных связей с помощью ДНК-лигазы. Таким образом, реализация этого механизма обеспечивается тремя репарирующими ферментами.
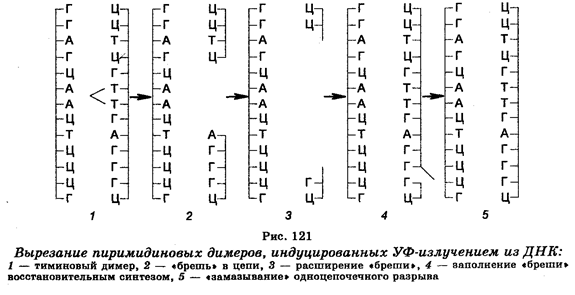
Третий механизм восстановления повреждений ДНК называют пострепликационным, или рекомбинационным восстановлением (рис. 122). Он заключается в том, что синтез ДНК в УФ-облученных клетках идет с нормальной скоростью вдоль хромосомы лишь до димера, перед которым он замедляется на несколько секунд, после чего начинается вновь, но уже на другой стороне димера. Поскольку ДНК-полимераза перескакивает через димер, то в дочерней цепи ДНК образуется брешь. Вследствие этого район, содержащий димер в одном дуплексе, будет интактным в сестринском дуплексе, т. е. в дочерних молекулах ДНК одна цепь будет содержать пиримидиновые димеры, тогда как другая будет иметь бреши, которые фактически являются вторичными повреждениями. Следовательно, район, содержащий димеры в одном дуплексе, полностью сохраняется в сестринском дуплексе. Этот процесс заканчивается рекомбинацией вдоль молекулы ДНК после ее репликации, при которой дочерняя цепь, несущая в каком-либо участке брешь, спаривается другой дочерней цепью (комплементарной), несущей брешь в другом месте. Это спаривание позволяет восстановительный синтез, который обеспечивает восстановление правильной последовательности района в каждой бреши. В качестве шаблона используется соответствующий интактный район другой дочерней цепи. Рекомбинационные события на уровне каждой бреши приводят к реконструкции интактной молекулы ДНК, способной к дальнейшей репликации. Рекомбинационное восстановление ДНК обеспечивается рядом белков-рекомбиназ.
 В ходе эволюции у клеток выработалась способность синтезировать репарирующие ферменты, которые синтезируются, когда начинают возникать повреждения ДНК. Например, у Е. coli открыта так называемая SOS-репарация, которая заключается в том, что любое повреждение ДНК, сопровождающееся нарушением ее репликации, ведет к усилению транскрипции большого количества генов (более 15), кодирующих репарирующие белки. Этот процесс сопровождается повышением выживаемости клеток. Известно также, что существуют репарирующие системы, которые активируются, если в ДНК образуются метилированные нуклеотиды. Подобные индуцированные системы репарации существуют, вероятно, и у эукариотических клеток. В ходе эволюции у клеток выработалась способность синтезировать репарирующие ферменты, которые синтезируются, когда начинают возникать повреждения ДНК. Например, у Е. coli открыта так называемая SOS-репарация, которая заключается в том, что любое повреждение ДНК, сопровождающееся нарушением ее репликации, ведет к усилению транскрипции большого количества генов (более 15), кодирующих репарирующие белки. Этот процесс сопровождается повышением выживаемости клеток. Известно также, что существуют репарирующие системы, которые активируются, если в ДНК образуются метилированные нуклеотиды. Подобные индуцированные системы репарации существуют, вероятно, и у эукариотических клеток.
У человека известен синдром «ксеродерма пигментозум», который наследуется в качестве хромосомного рецессивного признака и который характеризуется чрезвычайной чувствительностью кожи к солнечному свету, в результате чего она подвергается избыточной пигментации, а затем часто происходит и малигнизация кожных клеток, т. е. развивается кожный рак. Возникновение этого синдрома связано с дефектом способности вырезать тиминовые димеры из ДНК. Известен также синдром Блума, наследуемый в качестве рецессивного признака и заключающийся в повышенной чувствительности индивидов к солнечному свету. Этот синдром связан также с увеличением геномной нестабильности в виде повышения сестринских хроматидных обменов, хромосомных аберраций в геномах больных, повышения риска развития у них всех типов рака и, самое главное, с дефектом восстановления их ДНК. Как для синдрома «ксеродерма пигментозум», так и для синдрома Блума характерна иммунологическая недостаточность у больных.
Нормальные повреждения ДНК, индуцированные солнечным светом (УФ-компонентом), восстанавливаются «вырезанием-восстановлением».
Некоторые из потенциально летальных или вторичных повреждений, индуцируемых рентгеновским излучением, могут быть восстанавливаемыми посредством рекомбинации или какого-либо другого механизма, в котором участвуют ферменты-рекомбиназы. Предполагается также, что в отличие от повреждений ДНК, индуцированных УФ-излучением, повреждения, индуцированные рентгеновским излучением, подвержены восстановлению (через рекомбинацию) еще до первой постлучевой репликации.
Повреждения, вызываемые в ДНК химическими мутагенами, также восстанавливаются с помощью того или иного механизма. Каждый из механизмов восстановления ДНК является, по существу, системой защиты ДНК. В то же время восстановление ДНК часто сопровождается ошибками, проявляющимися в форме мутаций.
Обобщая значение репарационных механизмов в жизни клеток, можно заключить, что репарация повреждений ДНК обеспечивает поддержание стабильности генов, причем она основана на наличии двух цепей в ДНК. Именно благодаря этому повреждения в одной цепи могут репарироваться за счет информации неповрежденной цепи. Однако ДНК, вопреки тому, что она является хранителем генетической информации, все же обладает ограниченной химической стабильностью. В клетках с довольно высокой частотой встречается гидролиз, окисление и неэнзиматическое метилирование ДНК. Эти реакции взаимодействуют с восстановлением ДНК. Предполагают, что спонтанный распад ДНК, вероятно, является главным фактором в спонтанном мутагенезе, карциногенезе и наступлении старения организмов. Таким образом, ДНК представляется противоречивой структурой. С одной стороны, она очень консервативна в плане ее стабильности, с другой стороны, она очень подвержена распаду.
В историческом плане вопрос об эволюции генов является важнейшим, поскольку эволюция генов связана с истоками жизни вообще и ее совершенствованием в частности. Поскольку выявлена изначальная роль в происхождении жизни РНК, то предполагают, что начало эволюции генов датируется 3,5-4 млрд лет назад, когда сформировались первые молекулы РНК, которые каким-то образом детерминировали синтез белков, т. е. были первыми хранителями генетической информации. Однако когда выявилась необходимость в повышении эффективности синтеза белков, способность кодирования генетической информации перешла к ДНК, которая стала главным хранителем генетической информации. Что касается РНК, то она оказалась между ДНК и белком, став «переносчиком» информации. Конечно, эта гипотеза не имеет доказательств. Тем не менее многие далее считают, что появление ДНК связано с усложнением структуры клеток и, следовательно, необходимостью кодирования большого количества информации по сравнению с РНК. Другими словами, с началом участия ДНК в хранении генетической информации стал развиваться генетический код.
В последнее время большое внимание приобрела гипотеза, в соответствии с которой источником новых генов является рекомбинация эксонов, а также транспозоны, поступающие в геномы организмов.
Особый интерес в эволюционном плане представляет ДНК, которая не транскрибируется (эгоистическая ДНК). Казалось, должны быть какие-то факторы контрселекции, которые обеспечивают поддержание этой ДНК в клетках. Между тем такие факторы неизвестны. Тем не менее очень популярно предположение, что эгоистическая ДНК тоже является источником образования новых генов.
В обсуждении направления эволюции геномов известно два объяснения. Одни ученые предполагают, что увеличение геномов клеток в процессе эволюции организмов шло путем включения в ядерные структуры дополнительных копий генов, в то время как другие считают, что в эволюции шла дупликация уже образованных генов с последующей их дивергенцией. Доказательства включения генов в геномы отсутствуют, тогда как предположение о дупликации и дивергенции генов имеет существенные обоснования, причем эти обоснования исходят из данных о том, что многочисленные семейства белков кодируются наборами родственнных генов.
Установлено, например, что в кодировании яичного альбумина цыплят участвует несколько генов. Лейкоцитарный интерферон человека кодируется девятью неаллельными генами, а 8-кристаллин цыплят — двумя генами.
Белок актин у насекомых и морских ежей контролируется тоже несколькими генами, причем для каждого актина в разных сократительных клетках существует свой ген. Белки хориона контролируются тоже несколькими генами, причем объединенными в сложный кластер. Идентифицировано несколько генов, кодирующих родопсины, обеспечивающие восприятие разных цветов. Наконец, иммуноглобулины также кодируются многими генами, например, у мышей тремя генами. Известны и другие примеры. Поэтому можно сказать, что синтез лишь нескольких белков контролируется одиночными генами. Например, одним геном кодируется синтез инсулина человека и кур. В подтверждение предположения о дупликации и дивергенции генов и, следовательно, о механизмах увеличения геномов наиболее показательны данные об эволюции мультисемейства генов, кодирующих гемоглобины.
У морских червей, некоторых насекомых и рыб размеры глобиновой молекулы, переносящей кислород, составляют всего лишь 150 аминокислотных остатков. У человека каждая молекула гемоглобина состоит из двух a-полипептидных цепей и двух b-полипептидных цепей.
 Синтез гемоглобинов кодируется двумя несцепленными кластерами генов. Кластер глобиновых генов локализован на 16 хромосоме и содержит два эмбриональных, а также два почти идентичных фетальных глобиновых гена. Второй кластер, располагающийся на хромосоме II, кодирует b-глобины. Он состоит из b-глобинового гена, одного эмбрионального e-гена, двух фетальных генов, а также гена d-глобина. Синтез гемоглобинов кодируется двумя несцепленными кластерами генов. Кластер глобиновых генов локализован на 16 хромосоме и содержит два эмбриональных, а также два почти идентичных фетальных глобиновых гена. Второй кластер, располагающийся на хромосоме II, кодирует b-глобины. Он состоит из b-глобинового гена, одного эмбрионального e-гена, двух фетальных генов, а также гена d-глобина.
Исходя из данных о генетическом контроле глобинов и данных секвенирования глобиновых генов, предполагают, что самая первая дупликация гена, контролирующего синтез белка, являющегося предшественником гемоглобина, произошла 1 млрд лет назад, дав начало паре генов. Один ген из этой пары в ходе эволюции стал геном, контролирующим синтез миоглобина, другой развился в ген, контролирующий синтез гемоглобина. В последующем (около 500 млн лет назад) ген гемоглобина вновь подвергся дупликации, в результате чего возникли гены, контролирующие синтез гемоглобиновых цепей a и b. Через некоторое время дальнейшей дупликации подвергся ген b-цепи, что дало начало гену гемоглобина, синтезируемого только в эмбриональном периоде. Далее ген эмбрионального гемоглобина подвергся тоже дупликации, что дало начало уже гемоглоби-нам в и b. Еще одна дупликация гена b-цепи дала ген, кодирующий синтез e-глобина (рис. 123). Таким образом, в результате серии тандемных дупликации исходный глобиновый ген стал на одной из стадий развития кластерами генов a- и b-глобинов.
Эволюция геномов ускоряется транспозируемыми элементами, которые обладают способностью наращивать геномы. Наконец, увеличению геномов способствует генетическая рекомбинация.
Один из принципиальных вопросов связан с познанием механизмов, предупреждающих гены от дупликации в результате транскрипционной активации генов-«сосед ей». В недавнее время на примере D. melanogaster показано существование пограничных сегментов (последовательностей) ДНК, функционально изолирующих соседние гены. Длина последовательностей-изоляторов составляет около 340 пар оснований. Не исключено, что такие последовательности существуют и у других организмов.
1. На чем основаны представления о том, что генетическим материалом являются нуклеиновые кислоты? Каково значение генной инженерии в перечне доказательств генетической роли ДНК?
2. Существует ли связь между размерами генома (в количестве нукле-отидных пар) и видовой принадлежностью организмов? Приведите примеры в обоснование вашей точки зрения.
3. Что вы знаете о путях увеличения генома клеток в процессе развития организмов от низших форм к высшим?
4. Определите в сантиметрах общую длину ДНК в клетках человека.
5. ДНК стабильна при рН11, но РНК деградирует до нуклеотидов при щелочной реакции. Используйте учебники по биохимии и объясните причину этого явления.
6. Если субъединицы b и b' РНК-полимеразы составляют 0,005 доли от массы общего белка в клетках Е. coli, то сколько будет в клетке молекул РНК-полимеразы при условии, что каждая субъединица b и b' представляет цельную молекулу этого фермента?
7. Почему мочевина денатурирует РНК?
8. Состав оснований (фракция Г+Ц) двухцепочечной молекулы ДНК отражается на показателях плавучей плотности в хлориде цезия и температуры плавления (Тп), при которой половина молекул «плавится» на отдельные цепи. Было найдено, что плавучая плотность равна 1,660 + (0,098´фракция Г+Ц), фракция Г+Ц = 2,44 (Тп — 69,3), причем Tп определена в стандартном солевой растворе. Плавучая плотность ДНК крысы составляет 1,702, D. melanogaster — 1,698 и дрожжей — 1,699. Определите фракцию Г+Ц и температуру плавления для ДНК каждого вида.
9. Каково значение митохондриальной ДНК человека?
10. Что представляют собой транспозируемые генетические элементы? Как их классифицируют?
11. Что представляют собой плазмиды?
12. Что представляют собой повторяющиеся последовательности ДНК и как часто они повторяются в геноме человека?
13. В чем заключается полуконсервативный способ репликации ДНК и каково биологическое значение такого способа репликации?
14. Какова роль ферментов в репликации ДНК?
15. Есть ли разница между репликацией ДНК и репликацией хромосом?
16. Что такое нуклеосома и каковы ее размеры? Какова роль белков в упаковке ДНК в хромосомы?
17. Вычислите число нуклеотидных пар в 1 мегадальтоне двухцепочечной ДНК.
18. Как вы думаете, сколько генов имеется в одной клетке человека при условии, что длина одного гена составляет около 500 пар нуклеотидов?
19. По данным кислотного гидролиза препарат ДНК, выделенной из клеток мертворожденного плода человека, характеризовался следующим составом (в %): аденин — 25, тимин — 32, гуанин — 22, цитозин — 21. Каким образом можно объяснить этот необычный результат исследования, руководствуясь данными о структуре ДНК?
20. Какой будет длина и суммарная масса ДНК, если в ней объединить молекулы ДНК из всех клеток новорожденного ребенка, организм которого состоит из 2,6´1012
клеток?
21. Что представляют собой ферменты Оказаки и какова их роль в репликации ДНК?
22. Можете ли вы назвать экспериментальные данные, подтверждающие антипараллельную ориентацию цепей в молекуле ДНК?
23. После денатурации ДНК ренатурировали, позволив регибридиза-цию цепей до 1% их последовательностей. Затем эту ДНК подвергли обработке нуклеазой I до полного переваривания молекул, после чего ее «разогнали» путем электрофореза в агарозном геле. Каковы результаты электрофореза?
24. Каким образом можно определить генетическую локализацию IS1-последовательностей в геноме Е. coli?
25. Что собой представляют прямые и обратные мутации и каковы частоты этих мутаций по сравнению с супрессорными мутациями?
26. В чем заключаются молекулярные механизмы генных мутаций?
27. Опишите механизмы действия физических и химических мутагенов.
28. Могут ли восстанавливаться повреждения ДНК? Если да, то с помощью каких механизмов и какое значение это явление имеет в жизни организмов?
29. Можете ли вы объяснить расщепление ДНК в процессе ее восстановления с 5'-стороны от тиминового димера, но не с 3'-стороны?
Альберте В., Брей Д., Льюис Дж., Ряфф М., Роберте К., Уотсон Дж. Молекулярная биология клетки. М.: Мир. 1994. Т. 1. 815 стр.
Пехов А. П. Основы плазмидологии. М
.: РУДН. 1998. 231 стр.
Karcher S. J. Molecular Biology. Academic Press.
1995. 273 pp.
Schleif R. Genetics and Molecular Biology. The Johns Hopldns University Press.
1983. 698 pp.
Свременные представления о материальных основах наследственности берут начало от исследований Г. Менделя (1822—1884), историческая заслуга которого состоит в том, что он сформулировал теорию, объясняющую наследственность существованием и передачей факторов (единиц) наследственности от родителей к потомству через репродуктивные клетки. Г. Мендель обосновал значение генов как фундаментальных единиц жизни и установил законы передачи этих единиц от родителей к потомству. Заключения, сформулированные Г. Менделем, принято называть теперь законами наследственности.
В последующем Т. Морган (1886-1945) представил решающие доказательства в пользу того, что гены фиксированы на хромосомах.
Работы Г. Менделя, Т. Моргана и их последователей заложили основы теории гена и хромосомной теории наследственности.
Основу исследований Г. Менделя, которые проводились, когда еще не были известны хромосомы, составляют скрещивания и изучение гибридов садового гороха. Г. Мендель начал исследования, располагая 22 чистыми линиями садового гороха, которые имели хорошо выраженные альтернативные (контрастирующие) различия между собой по семи парам признаков, а именно: форма семян (круглые —шероховатые), окраска семядолей (желтые — зеленые), окраска кожуры семян (серая — белая), форма бобов (выполненные — морщинистые), окраска незрелых бобов (зеленые — желтые), расположение цветков (пазушные — верхушечные), высота растения (высокие — карликовые).
В отличие от своих предшественников, пытавшихся изучать поведение в скрещиваниях сразу нескольких признаков, Г. Мендель в скрещиваниях растения одной линии с растениями другой линии изучал поведение признаков раздельно. Когда было изучено поведение одной пары контрастирующих признаков, в дальнейших скрещиваниях наблюдению подвергалась следующая пара признаков и т. д. Закончив эту работу, Г. Мендель изучал затем поведение в скрещиваниях одновременно двух контрастирующих признаков, например, формы семян и окраска семядолей (дигиб-ридные скрещивания), а затем трех признаков — формы семян, окраска семядолей и окраски кожуры семян (тригибридные скрещивания).
Учитывая строение цветков садового гороха, Г. Мендель при скрещивании растений исходных родительских линий исключал самоопыление и допускал его лишь при скрещивании гибридов первого и второго поколений. Наконец, в поисках объяснений своих результатов Г. Мендель использовал математику. Анализируя полученные данные, он формулировал рабочие гипотезы, проверяя их правильность в последующих экспериментах. В целом работа Г. Менделя является классическим образцом методологии экспериментальной науки.
Г. Мендель скрещивал чистолинейные растения путем нанесения на пестик цветка растения одной линии пыльцы, взятой из цветка растения другой линии и наоборот. Когда в скрещивания были вовлечены растения двух линий, различающиеся по такой паре контрастирующих признаков, как круглая и шероховатая (угловатая) форма семян, т. е. одно скрещиваемое растение давало круглые семена, а другое — шероховатые (рис. 124), а затем от них были получены гибриды первого поколения (Fi), то оказалось, что все эти гибриды единообразны, т. к. они похожи на растения одной из родительских линий (давали семена круглой формы). Замечательным оказалось также и то, что все эти гибриды давали семена круглой формы независимо от принадлежности пыльцы цветкам растений одной или другой родительской линии.
Проверив в других скрещиваниях поведение остальных шести контрастирующих признаков, Г. Мендель наблюдал, что во всех случаях гибриды Fi также единообразны, ибо они были похожи на растения одной из двух скрещиваемых (родительских) чистых линий садового гороха в любом из соответствующих сочетаний. Таким образом, Г. Мендель обнаружил, что гибриды, полученные после скрещивания растений двух линий, различающихся по паре контрастирующих признаков, проявляют лишь один признак из этой пары.
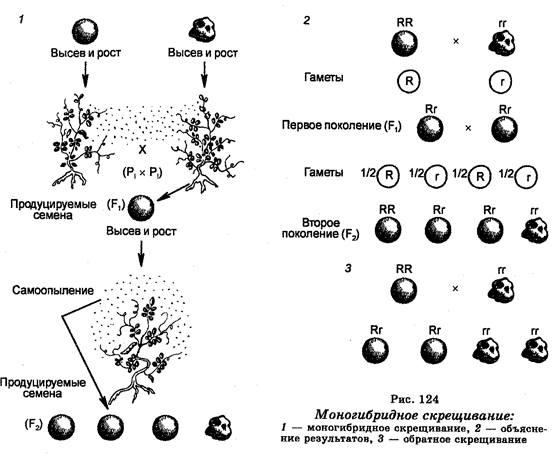
Признаки, которые проявились у гибридов первого поколения, Г. Мендель назвал доминантными. В результате всех скрещиваний доминантными признаками оказались круглая форма семян, желтая окраска семядолей, серый цвет кожуры семян, выполненность бобов, зеленая окраска незрелых бобов, наружное расположение цветков и высокий рост растений.
Продолжая эксперименты, Г. Мендель допустил самоопыление гибридов первого поколения, в результате которого цветки этих гибридов опылялись своей же пыльцой. В противоположность единообразию гибридов Г1
гибриды второго поколения (F2
) характеризовались тем, что среди них встречались как особи, у которых проявлялся доминантный признак, являющийся признаком одного из родителей, так и особи, у которых проявлялся признак другого родителя, т. е. тот признак, который находился в скрытом состоянии у гибридов F1
. Следовательно, применительно к любой паре контрастирующих признаков, по которым различались растения скрещиваемых линий, отсутствие одного из контрастирующих признаков у гибридов F1
не означало, что этот признак навсегда исчезал. Будучи лишь подавленным или скрытым у гибридов F1
, этот контрастирующий признак проявлялся затем у гибридов F2
.
Признаки, подавляемые в гибридах, Г. Мендель назвал рецессивными признаками. В числе рецессивных признаков садового гороха оказались шероховатая форма семян, зеленая окраска семядолей, белый цвет кожуры семян, морщинистая форма семян, желтая окраска незрелых бобов, верхушечное расположение цветков и карликовый рост растений.
Полное доминирование, обнаруженное Г. Менделем, в последующем было показано многими исследователями у гибридов животных (млекопитающие, рыбы и др.) и растений. В то же время было отмечено отсутствие доминантности отдельных признаков как у животных, так и у растений. Например, скрещивание красных шортгорнов с белыми сопровождается тем, что гибриды первого поколения имеют промежуточную масть (чалую). Расщепление гибридов во втором поколении дает красных, белых и чалых шортгорнов. Наконец, у многих живых существ отдельные признаки наследуются по частично доминантному типу, что бывает тогда, когда одни признаки частично доминируют над другими. Как видно, доми-нантность того или иного признака может быть выражена в различной степени и может отсутствовать полностью. Однако важно подчеркнуть, что во всех выполненных к настоящему времени скрещиваниях, независимо от степени доминантности какого-либо признака все гибриды первого поколения по этому признаку являются единообразными.
Как отмечено выше, гибриды F2
садового гороха в экспериментах Г. Менделя характеризовались разнообразием, т. к. среди них встречались особи, у которых проявлялся как доминантный, так и рецессивный признаки. Можно сказать, что гибриды F2
были двух типов: одни из них были похожи на растения одной прародительской линии, другие — на растения другой прародительской линии. Например, гибриды из тех скрещиваний, в которых Г. Мендель наблюдал пару контрастирующих признаков, касающихся формы семян, характеризовались тем, что одни из них давали семена круглой формы, а другие — шероховатой.
Однако когда Г. Мендель провел подсчет гибридных растений F2
, различающихся по форме семян, то оказалось, что из 7324 гибридов 5474 гибрида (74,74%) давали круглые семена (доминантный признак), а 1850 гибридов (25,36%) — шероховатые (рецессивный признак). Другими словами, три четверти всех гибридов F2
были похожи на растения одной прародительской линии, а одна четверть — на растения другой прародительской линии. Следова- тельно, отношение между гибридами Рд, дающими круглые семена, и такими же гибридами, дающими шероховатые семена, есть отношение между доминантными и рецессивными признаками гибридов. Аналогичные отношения между разными гибридами второго поколения были выявлены и в скрещиваниях, в которых наблюдению подлежали остальные контрастирующие признаки садового гороха (табл.15).
Таблица 16
Расщепление гибридов садового гороха второго поколения
в опытах Г. Менделя
| Признаки растений исходных линий
|
Общее количество учтенных гибридов
|
Доминантные признаки
|
Рецессивные признаки
|
| количество гибридов
|
%
|
количество гибридов
|
%
|
| форма семян
|
7324
|
5474
|
74,74
|
1850
|
25,26
|
| Окраска семядолей
Окраска кожуры семян
|
8023
929
|
6022
705
|
75,06 75,89
|
2001
224
|
24,94 24,11
|
| форма бобов
|
1181
|
882
|
74,68
|
299
|
25,32
|
| Окраска незрелых бобов
|
580
|
428
|
73,79
|
152
|
26,21
|
| Расположение цветков
|
858
|
651
|
75,87
|
207
|
24,13
|
| Высота растений
|
1064
|
787
|
73,96
|
277
|
26,04
|
| Всего:
|
19959
|
14949
|
74,90
|
5010
|
25,10
|
Как видно из табл. 15, у гибридов F2
проявлялся каждый признак из любой пары контрастирующих признаков. Другими словами, у гибридов F2
признаки подвергались расщеплению (сегрегации).
Обнаружение расщепления признаков во втором поколении гибридных растений садового гороха имело огромное значение, ибо, исходя из данных о частоте расщепления, Г. Мендель раскрыл внутреннее содержание этого явления, сформулировав гипотезу о существовании факторов (единиц) наследственности, которая затем переросла в теорию гена.
В рамках этой гипотезы Г. Мендель предположил, что контрастирующие признаки детерминируются какими-то факторами (элементами, единицами), которые передаются от родителей к потомству через половые клетки. Г. Мендель предположил также, что каждая пара контрастирующих признаков детерминируется парой факторов, которые в гибридах не смешиваются, но при образовании гамет расходятся, проходят чистыми в разные гаметы и затем оказываются в потомстве гибридов. Например, круглая форма семян, являясь доминантным признаком, детерминируется доминантным фактором, тогда как шероховатая форма семян, являющаяся рецессивным признаком, детерминируется рецессивным фактором. Каждое растение имеет пару генетических факторов на каждый контрастирующий признак, причем оба фактора присутствуют в растении одновременно.
После 1909 г. менделевские факторы наследственности по предложению В. Бэтсона (1861-1926) стали называть генами, членов генной пары — аллельными генами, или просто аллелями. Когда оба аллеля одинаковы (доминантны или рецессивны), то организм, несущий эти аллели, называют гомозиготным или гомозиготой по данному аллелю. Организм, несущий разные аллели (доминантный и рецессивный), называют гетерозиготным, или гетерозиготой.
Чтобы понять, каким образом гены передаются потомству и как они распределяются в потомстве между разными особями, необходимо уяснить сущность фенотипического и генотипического отношений, полученных Г. Менделем при изучении гибридов садового гороха. Обозначим, как это делал Г. Мендель, символом P1
исходные (родительские) растения, символами F1
и F2
— гибриды первого и второго поколений (соответственно), а символами R и г — аллельные гены (соответственно доминантный и рецессивный), детерминирующие, например, форму семян садового гороха. Поскольку растения P1
— чистолинейные, то те из них, которые дают круглые семена (являются гомозиготными и несут гены R и R), можно обозначить RR, а гаметы этих растений — R, тогда как те растения, которые дают шероховатые семена (являются тоже гомозиготными, несут гены г и г), можно обозначить гг, а гаметы этих растений — г.
Интерпретируя результаты своих опытов, Г. Мендель предположил, что при скрещивании исходных растений P1
женские гаметы R оплодотворяются мужскими гаметами г и наоборот, вследствие чего гибриды F1
будут гетерозиготными гибридами, имеющими генетическую формулу Rr и дающими семена круглой формы, т. к. аллель R подавляет аллель г. Когда гибрид F1
продуцирует гаметы, аллели Rr расщепляются, в результате чего половина гамет будет нести аллель R, половина — аллель г. Если же при самоопылении гибридов F1
женская гамета г оплодотворяется мужской гаметой R, то гибрид F2
будет иметь генетическую формулу RR, его гаметы будут гаметами R, а самоопыление таких гибридов F2
дает начало растениям RR. Если при самоопылении гибридов F1
женская гамета г будет оплодотворена мужской гаметой г, гибрид F2
будет гибридом с генетической формулой гг, его гаметы будут гаметами г, а самоопыление таких гибридов F2
дает растения гг. Если же при самоопылении гибридов F1
женская гамета г будет оплодотворена мужской гаметой г или наоборот, то образующийся гибрид F2
будет растением Rr (подобно гибриду F1
), его гаметы будут гаметами двух видов — мужские R и r и женские R и г, а самоопыление таких гибридов дает растения с генетическими формулами RR, Rr и гг.
Имея в виду соображения, приведенные выше, вернемся к гибридам F1
которые являются гетерозиготами Rr и продуцируют гаметы R и г в равных количествах. Допуская, что при оплодотворении гаметы объединяются случайно (нет преимуществ для объединения гамет R и R или г и г, или R и г) и что все формирующиеся гибридные растения жизнеспособны, можно предположить, что в результате расщепления генов формируемые гибриды будут различными как по фенотипу, так и по генотипу (см.рис. 124).
Объединение гамет гибридов F1
может давать четыре двойных сочетания аллелей, причем каждое из сочетаний должно давать начало не меньше одной четверти гибридов F2
. Оценивая генотипы гибридов F2
, можно видеть, что гомозиготные растения RR имеют лишь аллели, детерминирующие круглую форму семян, и поэтому такие растения должны давать семена круглой формы. Далее, гете-розиготные растения Rr имеют аллель для круглой формы и аллель для шероховатой формы семян, но по причине доминантности аллеля R эти растения дадут семена также круглой формы. Наконец, гомозиготные растения гг имеют аллели лишь для шероховатой формы семян, и в связи с этим такие растения дадут шероховатые семена.
Следовательно, если допущения случайного объединения гамет при оплодотворении и равной во всех случаях жизнеспособности гибридов справедливы, тогда среди гибридов F2
три четверти особей должны давать круглые семена и одна четверть — шероховатые, т. е. растения, дающие семена разной формы, среди гибридов должны встречаться в отношении 3:1, которое является феноти-пическим отношением.
Возвращаясь к данным табл. 15, можно видеть, что среди 7324 гибридов Рд, изученных Г. Менделем, 5474 гибрида давали круглые семена и 1850 — шероховатые, что составляет соответственно 74,74% и 25,26% и очень близко к отношению 3:1. Такое фенотипическое отношение среди гибридов Fg Г. Мендель обнаружил и в случае других альтернативных признаков. Говоря о мен-делевском фенотипическом отношении среди гибридов следует, однако, отметить, что оно не точно и лишь указывает на то, что можно ожидать на основе вероятности. Поэтому данное отношение будет тем точнее, чем большее количество гибридов будет исследовано.
Итак, интерпретация Г. Менделем сущности фенотипического отношения 3 : 1 заключается в том, что растения F2
, дающие круглые семена, генотипически неодинаковы, тогда как растения, дающие шероховатые семена, генотипически одинаковы (см. рис. 124). Среди гибридов F2
, дающих круглые семена, около одной трети должно быть гомозиготными растениями RR (по аллелю для круглой формы семян) и около двух третей — гетерозиготными Rr, имеющими и доминантный, и рецессивный аллели. Все растения, дающие шероховатые семена, являются рецессивными гомозиготами rr.
Чтобы проверить справедливость генной гипотезы, Г. Мендель осуществил ряд дальнейших скрещиваний. В частности, он скрещивал гибриды второго поколения, допуская их самоопыление и получая таким путем гибриды третьего поколения (F3
). Все гибриды F3
, происходящие от гибридов F3
(предположительно rr), дававшие шероховатые семена, оказались растениями, снова дающими шероховатые семена, т. е., как и следовало ожидать, растениями rr.
Что касается гибридов F3
, которые произошли от гибридов F2
, дававших круглые семена, то они оказались неодинаковыми. Самоопыление 1/3 всех этих гибридов F2
, (предположительно RR) дало начало гибридам F3
, продуцирующим семена только круглой формы, т. е. гибридам RR. Самоопыление 2/3 гибридов F2 (предположительно Rr) дало начало гибридам F3
, среди которых три четверти составляли растения, продуцировавшие круглые семена (RR и Rr) и одну четверть — продуцировавшие шероховатые семена (гг). Как и в случае гибридов F2
, здесь фенотипическое отношение также составляло соответственно 3:1.
Таким образом, гибридные растения второго поколения независимо от фенотипа генотипически оказались растениями трех типов: растения RR, дающие круглые семена и остающиеся такими в дальнейших скрещиваниях (подобно растениям исходной линии, использованной в первоначальном скрещивании), гетерозиготные растения Rr, дающие подобно гибридам первого поколения круглые семена, но расщепляющиеся в дальнейших скрещиваниях, и гомо-зиготные растения гг, дающие шероховатые семена и остающиеся такими в дальнейших скрещиваниях (подобно растениям другой исходной линии, использованной в первоначальном скрещивании), причем в отношении 1/4:1/2 :1/4 или 1:2:1 соответственно.
Отношение 1:2:1 является генотипическим и было обнаружено Г. Менделем также при анализе остальных шести признаков (цвет семян, положение цветков и т. д.). Следовательно, разница между видимым (фенотипическим) и фактическим (генотипическим) отношениями обусловлена доминантным характером аллеля R и рецессивным характером аллеля г, вследствие чего генотипы RR и Rr дают одинаковый фенотип.
Помимо скрещиваний, результаты которых приведены выше, для доказательства генной гипотезы Г. Мендель обратился также к обратным скрещиваниям, получившим позднее в литературе название анализирующих (тест-скрещиваний). Смысл этих скрещиваний заключается в том, что гетерозиготные гибриды F1
которые давали, например, круглые семена и происходили из скрещиваний между исходными растениями, дающими круглые (RR) и шероховатые (rr) семена, вновь скрещивали с исходными (родительскими) гомозиготными рецессивными растениями, продуцирующими шероховатые семена. Поскольку гаметы, проецируемые гетерозиготным (Rr) гибридом F1
, всегда являются чистыми и могут нести только или аллель R, или аллель г, причем половина гамет должна быть гаметами R, половина — гаметами г, а все гаметы, продуцируемые исходным гомозиготным рецессивным (гг) растением, должны быть только гаметами г, в случае справедливости генной гипотезы следовало ожидать, что обратное скрещивание таких растений должно привести к образованию зигот наполовину Rr и наполовину гг. Другими словами, образующееся в результате таких обратных скрещиваний потомство должно состоять наполовину из гетерозиготных организмов, проецирующих круглые семена (Rr), и наполовину из гомозиготных рецессивных организмов, продуцирующих семена шероховатой формы (гг). Осуществив обратные скрещивания и проанализировав свойства появлявшихся в этих скрещиваниях растений, Г. Мендель обнаружил, что они действительно являются наполовину гетерозиготными организмами и наполовину гомозиготными, т. е. отношение между ними составляло 1:1.
Итак, из результатов этих экспериментов следует, что каждый наследуемый признак находится под контролем пары одинаковых (аллельных) генов, из которых один аллель (один ген пары) происходит от одного родителя, тогда как другой аллель (другой ген пары) берет начало от другого родителя. Находясь в гибриде первого поколения, различные аллели не смешиваются и не влияют один на другой. Когда у зрелого организма наступает процесс образования половых клеток, то два аллеля генной пары расходятся (сегре-гируют) чистыми и оказываются в разных гаметах, продуцируемых одним и тем же гибридом. Гаметы несут лишь по одному аллелю из каждой генной пары. Появление одного аллельного гена в той или иной гамете — дело случая. В процессе последующего оплодотворения женских половых клеток мужскими и образования гибридов второго поколения гены вновь собираются в пары. Заключение Г. Менделя, в соответствии с которым члены какой-либо генной пары расщепляются и расходятся в разные гаметы, известно под названием закона расщепления (сегрегации), или первого закона наследственности (первого закона Г. Менделя), и лежит в основе теории гена.

Сейчас известно, что некоторые признаки контролируются более чем двумя аллелями, т. е. существуют множественные аллели. Например, окраска шерстного покрова у кроликов (рис. 125) контролируется четырьмя аллелями (С1
, С2
, С3
, C4
. В случае человека группы крови по антигенам А и В контролируются тремя аллелями, (см. гл. XIII). Однако их наследование также подчиняется первому закону наследственности.
Установив расщепление генов в моногибридных скрещиваниях, Г. Мендель столкнулся с необходимостью выяснить, каким образом разные пары признаков садового гороха ведут себя по отношению одна к другой, если наблюдение за их передачей от одного поколения к другому вести одновременно. Поэтому следующую часть экспериментов Г. Менделя составляли скрещивания растений садового гороха, различающихся между собой сразу по двум парам альтернативных признаков, т. е. по двум парам разных генов, контролирующих две разных пары признаков одновременно. Такие  скрещивания называют дигибридными. Здесь мы рассмотрим одно из менделевских дигибридных скрещиваний, в котором в качестве исходных были использованы гомозиготные доминантные растения, дающие семена круглой формы и желтого цвета, и гомозиготные рецессивные растения, дающие семена шероховатой формы и зеленого цвета (рис. 126). скрещивания называют дигибридными. Здесь мы рассмотрим одно из менделевских дигибридных скрещиваний, в котором в качестве исходных были использованы гомозиготные доминантные растения, дающие семена круглой формы и желтого цвета, и гомозиготные рецессивные растения, дающие семена шероховатой формы и зеленого цвета (рис. 126).
Проведенные эксперименты по наблюдению двух пар различий (круглые и шероховатые семена, желтые и зеленые семена) в дигибридном скрещивании показали, что гибриды F1
в этих скрещиваниях похожи на гибриды F1
, возникающие при моногибридных скрещиваниях. Поскольку круглая форма семян является доминантной по отношению к шероховатой, а желтая окраска семян доминантной по отношению к зеленой, гибриды F1
давали круглые семена, имеющие желтую окраску. Проведя затем скрещивание гибридов F1
друг с другом или допуская самоопыление, Г. Мендель получил гибриды F2
, от которых было исследовано 656 семян (бобов). Изучив форму и окраску семян, полученных от гибридных растений F2
, он обнаружил, что раздельно как по форме, так и по окраске семян фенотипическое расщепление (отношение количества семян с доминантными признаками к количеству семян с рецессивными признаками) проявляется в отношении 3 : 1 или ¾ : 1/4, ибо из 556 семян для 432 была характерна круглая форма (76,08%) и для 133 - шероховатая (23,92%), тогда как из этого количества семян для 416 была присуща желтая окраска (74,82%), а для 140 — зеленая (25,18%). Такой характер расщепления по форме и по окраске семян следовало ожидать, исходя из результатов моногибридных скрещиваний. Однако в данном случае вопрос заключался в другом: является ли расщепление одной пары альтернативных признаков (круглая и шероховатая форма семян) независимым от расщепления другой пары признаков (желтая и зеленая окраска семян) или эти пары признаков зависимы, тесно связаны между собой? Аналогичный вопрос можно было поставить и по-другому: всегда ли круглая форма семян сочетана с желтой окраской их, а шероховатая форма семян — с зеленой окраской или же возможно появление семян с новыми комбинациями этих признаков?
Чтобы получить ответ на этот вопрос, Г. Мендель классифицировал все 556 семян по обеим парам признаков одновременно и получил следующие данные: круглые желтые — 315 семян, круглые зеленые — 108 семян, шероховатые желтые — 101 семя, шероховатые зеленые — 32 семени. Как видно, наряду с семенами, похожими на семена, которые давали растения исходных родительских линий (круглые желтые и шероховатые зеленые), гибриды Fg давали также семена, похожие на семена исходных родительских растений только по форме или по окраске. Следовательно, среди растений, дававших круглые семена и количественно составлявших 3/4 исследованных растений (всей группы), в свою очередь 2/4 давали желтые семена и 1/4 — зеленые. Среди растений, дававших шероховатые семена и количественно составлявших 1/4 всех исследованных растений, в свою очередь 3/4 давали желтые семена 1/4 — зеленые. Анализ этих количественных отношений приводил к дальнейшему заключению:
а) 3/4 растений от 3,4 всех растений исследованной группы F2
или 9/16 растений этой группы обладают обоими доминантными признаками (круглая форма и желтая окраска);
б) 1/4 растений от 3/4 растений всей группы или 3/16 растений всей группы обладают одним доминантным (круглая форма семян) или одним рецессивным (зеленая окраска семян) признаком, т. е. признаками в новых сочетаниях (рекомбинациях);
в) 3/4 растений от 1/4 растений всей группы или 3/16 растений всей группы обладают другим новым сочетанием доминантного и рецессивного признаков (шероховатая форма и желтая окраска семян);
г) 1/4 pастений от 1/4 растений всей группы или 1/16 растений всей группы обладают обоими рецессивными признаками (шероховатая форма и зеленая окраска семян).
Возвращаясь к количественным данным по систематизации семян с учетом их формы и окраски (315, 108, 101 и 32), легко заметить, что они очень близки к выведенным выше отношениям. Поэтому, опираясь на эти результаты, Г. Мендель сделал заключение, в соответствии с которым гибриды F2
по двум парам признаков являются организмами четырех типов в отношении 9/15:3/16:3?16:1/16 или 9:3:3:1. Таким образом, результаты дигибридных скрещиваний садового гороха свидетельствовали о том, что расщепление одной пары признаков (круглая и шероховатая форма семян) совершенно не зависит от расщепления Другой пары признаков (желтая и зеленая окраска семян) и что для семян гибридов F характерны не только родительские комбинации признаков, но и рекомбинации (новые комбинации), т. е. сочетание двух пар признаков имеет независимым характер. Другими словами, перераспределение двух пар признаков при их пере
даче от одного поколения организмов к другим является независимым. Однако важно подчеркнуть, что в независимом перераспределении признаков Г. Мендель увидел независимое перераспределение факторов наследственности или, говоря современным языком, независимое перераспределение генов.
Чтобы понять независимое перераспределение генов, вновь обратимся к символике (см. рис. 126), обозначив доминантный ген, контролирующий круглую форму семян, символом R, а его аллель, контролирующий шероховатую форму семян, — символом г. Что касается доминантного гена, контролирующего желтую окраску семян, то обозначим его Y, а его аллель, контролирующий зеленую окраску семян, — у. Используя эти символы и зная, что исходные родительские растения были чистолинейными, можно полагать, что растения, дававшие круглые желтые семена, несли гены RRYY, тогда как растения, дававшие семена шероховатой формы и зеле--ного цвета, несли гены ггуу. Следовательно, растения RRYY продуцировали гаметы RY, а растения rryy — гаметы гу, объединение которых давало начало гибридным зиготам Fi с генотипом RrYy.
Как видно, гибриды F1
являются гетерозиготными растениями по двум парам генов, т. е. растениями RrYy. Если иметь в виду лишь форму семян, то гибриды F с генотипом Rr будут продуцировать гаметы Риг поровну. Если же иметь в виду одновременно форму и окраску семян, то каждая гамета, продуцируемая гибридом Fi с генотипом RrYy, будет нести не только один генный аллель для формы семян, но и один генный аллель для окраски семян. Так, например, гаметы с аллелем R или с аллелем г наполовину будут одновременно гаметами Y и наполовину одновременно гаметами у. Однако важно заметить, что сочетание аллеля, детерминирующего круглую форму семян, с аллелем, определяющим желтую окраску семян, или с аллелем, определяющим зеленую окраску семян, является делом случая. Поскольку перераспределение генов имеет независимый характер, то среди гамет R, продуцируемых Г,-гибридом RrYy, половина гамет будет обладать аллелем Y и половина — аллелем у. Учитывая то, что гаметы R сами составляют половину гамет, продуцируемых гибридом F1
, гаметы как RY, так и гу будут составлять по 1/4 от общего количества гамет гибрида F. Точно так же другая половина гамет, продуцируемая гибридом F1
, т. е. гаметы г в свою очередь наполовину будут гаметами rY и наполовину — гаметами Ry, составляя соответственно по \
I ц от общего количества гамет гибрида F1
. Другими словами, гибриды F1
продуцируют гаметы четырех типов — RY, Ry, rY и гу, причем в равных количествах. Заметим, что Ry и rY являются гаметами рекомбинантного типа.
Гаметы четырех типов, продуцируемые гибридами Гд, являются как мужскими, так и женскими, т. е. мужские гаметы будут гаметами четырех типов и женские гаметы будут гаметами четырех типов. Поскольку объединение тех или иных мужских и женских гамет при оплодотворении также является делом случая и одинаково вероятно, то скрещивания F1
´F1
дадут начало зиготам, несущим все 16 возможных комбинаций генов (рис. 126). Фенотипически организмы F2
различаются между собой, т.к. 9 из них дают круглые желтые семена, 3 — круглые зеленые, 3 — шероховатые желтые и 1 — шероховатые зеленые. Следовательно, фено-типическое отношение между организмами Fg составляет 9:3:3:1. Гибриды F2
различаются одновременно и по генотипу. Организмы, которые дают круглые и желтые семена, по генотипу можно классифицировать на четыре типа:
а) организмы с генотипом RRYY (гомозиготные по обеим парам генов и дающие потомство с таким же фенотипом и генотипом при скрещивании между собой);
б) организмы с генотипом RRYy (гомозиготные по паре генов, определяющих форму семян, но гетерозиготные по паре генов, определяющих окраску семян, вследствие чего являются чистоли-нейными в дальнейших скрещиваниях лишь по генной паре RR);
в) организмы с генотипом RrYY (гетерозиготные по паре генов, определяющих форму семян, но гомозиготные по паре генов, определяющих окраску семян, вследствие чего являются чистоли-нейными в дальнейших скрещиваниях лишь по паре генов YY);
г) организмы с генотипом RrYy (гетерозиготные по обеим парам генов и при скрещивании между собой будут давать потомство, подобное потомству Fg).
Кроме того, организмы с генотипами RRYY, RRYy, RrYY и RrYy различаются между собой и количественно, составляя отношение 1:2:2:4 (соответственно).
Приведенные выше рассуждения с использованием символов основаны на допущениях. Однако Г. Мендель проверил их экспериментально путем скрещиваний между собой всех 315 гибридов Fg, которые давали круглые желтые семена. Проанализировав потомство, полученное в этих скрещиваниях, он обнаружил организмы всех типов, которые следовало ожидать на основании допущений, приведенных выше (пп. а, б, в, г).
Для доказательства справедливости своих допущений о независимом распределении двух пар аллельных генов Г. Мендель использовал также и дигибридные анализирующие скрещивания растений Г,, дающих круглые желтые семена, с гомозиготными рецессивными по двум парам генов исходными родительскими растениями, дающими шероховатые зеленые семена. Вспомним, что гибриды Fi имеют генотип RrYy и продуцируют в равных количествах гаметы RY, rY, Ry и гу, тогда как исходные родительские гомозиготные рецессивные растения имеют генотип ггуу и продуцируют гаметы только одного типа — гу. Следовательно, в результате этих скрещиваний можно было ожидать появление организмов, которые будут давать круглые желтые семена (генотип RrYy), шероховатые желтые семена (генотип rrYy), круглые зеленые семена (генотип Rryy) и шероховатые зеленые семена (генотип ггуу), причем в отношении 1 : 1 : 1 : 1 (в равных количествах). Как показали анализирующие дигибридные скрещивания, возникавшие гибриды по своим свойствам соответствовали ожидаемым.
Независимый характер перераспределения генов был установлен Г. Менделем и в скрещиваниях растений, различающихся между собой по трем парам признаков (тригибридные скрещивания). Рассмотрим один из менделевских экспериментов, в котором скрещивали растения садового гороха, дающие желтые круглые семена и имеющие красные цветки, с растениями, дающими шероховатые зеленые семена и имеющими белые цветки. Если использовать символы R и г, Y и у для обозначения пар генов, детерминирующих форму и окраску семян, а символы С и c — для обозначения пары генов, детерминирующих окраску цветков, то тогда генотипы скрещиваемых растений двух линий можно обозначить как RRYYCC и ггуусс, соответственно, имея в виду также генотипы скрещиваемых организмов, можно было полагать, что последние продуцируют гаметы, несущие гены RYC и гены гус.
Тригибридные скрещивания дали начало гетерозиготным по всем трем парам генов тригибридам F1
с генотипом RrYyCc и вследствие доминантности фенотипически характеризующимся тем, что их семена будут круглыми желтыми, а цветки — красными. Тригибриды F1
отличались от дигибридов F1
тем, что вследствие независимого перераспределения генов они продуцировали гаметы не четырех, а восьми типов: RYC, RYc, RyC, Rye, rYC, rYc, ryC и rye, где большинство гамет (кроме RYC и гус) являлось рекомбинант-ным, т. е. несло новые (рекомбинантные) сочетания генов. Тригибриды F1
отличались от дигибридов F1
также и тем, что случайное объединение между теми или иными их гаметами (восьми типов) при оплодотворении приводило к формированию не 16, а 64 комбинаций гибридов F2
. Эти гибриды по фенотипу можно было разделить на 8 групп, состоящих из разного количества растений, для которых характерны следующие свойства:
круглые желтые семена, красные цветки — 27 растений,
круглые желтые семена, белые цветки — 9 —»—
круглые зеленые семена, красные цветки — 9 —»—
шероховатые желтые семена, красные цветки — 9 —»—
круглые зеленые семена, белые цветки — 3 —»—
шероховатые желтые семена, белые цветки — 3 —»—
шероховатые зеленые семена, красные цветки — 3 —»—
шероховатые зеленые семена, белые цветки — 1 —»—
Как мы уже видели, часть фенотипически сходных моногибридов F2
и дигибридов F2
генотипически разнообразна. Аналогичная ситуация имела место и в случае части фенотипически сходных тригибридов F2
. To, что это действительно было так, экспериментально устанавливалось путем скрещиваний тригибридов F2
между собой и изучения тригибридов F3
.
Результаты, которые можно ожидать при изучении тригибридов F2
и F3
приводятся в табл. 16.
Таблица 16
Генотипы и поведение в скрещиваниях организмов, которые можно ожидать в потомстве гибридов
F
,*
| Количество организмов
|
Класс генотипов
|
Класс фенотипов
|
Соотношение между классами фенотипов
|
Поведение а скрещиваниях при самоопылении
|
| 1
|
RRYYCC
|
27
|
Расщепления нет
|
| 2
|
RrYYCC
|
Круглые
желтые
|
Расщепление по форме семян 3 : 1
|
| 2
|
RRYyCC
|
семена,
|
Расщепление по окраске семян 3 : 1
|
| 2
|
RRYYCc
|
красные
|
Расщепление по окраске цветков 3 : 1
|
| 4
|
RrYyCc
|
цветки
|
Расщепление по форме и окраске семян 9:3:3:1
|
| 4
|
RrYYCc
|
Расщепление по форме семян и окраске цветков 9:3:3:1
|
| 4
|
RRYyCc
|
Расщепление по окраске семян и цветков 9:3:3:1
|
| 8
|
RrYyCc
|
Расщепление по форме семян, окраске семян и цветков 27 : 9 : 9 : 9 : 3 : 3 : 3:1
|
| 1
|
RRYYcc
|
Круглые желтые
|
9
|
Расщепления нет
|
| 2
|
RRYycc
|
семена,
|
Расщепление по окраске семян 3 : 1
|
| 2
|
RrYYcc
|
белые
|
Расщепление по форме семян 3:1
|
| 4
|
RrYycc
|
цветки
|
Расщепление по форме и окраске семян 9:3:3:1
|
| 1
|
RRyyCC
|
Круглые зeленые
|
9
|
Расщепления нет
|
| 2
|
RRyyCC
|
семена,
|
Расщепление по окраске цветков 3 : 1
|
| 2
|
RryyCC
|
красные
|
Расщепление по форме семян 3 : 1
|
| 4
|
RryyCc
|
цветки
|
Расщепление по форме семян и окраске цветков 9:3:3:1
|
Продолжение табл. 16
| Количество организмов
|
Класс генотипов
|
Класс фенотипов
|
Соотношение между классами фенотипов
|
Поведение в скрещиваниях при самоопылении
|
| 1
|
rrYYCC
|
Шероховатые
|
9
|
Расщепления нет
|
| 2
|
rrYyCC
|
желтые семена,
|
Расщепление по окраске семян 3 : 1
|
| 2
|
rrYYCc
|
красные
|
Расщепление по окраске цветков 3 : 1
|
| 4
|
rrYyCc
|
цветки
|
Расщепление по окраске семян и цветков 9:3:3:1
|
| 1
|
rryyCC
|
Шероховатые
|
3
|
Расщепления нет
|
| 2
|
rryyCc
|
зеленые семена, красные цветки
|
Расщепление по окраске цветков 3 : 1
|
| 1
|
rrYYcc
|
Шероховатые желтые семена, белые цветки
|
3
|
Расщепления нет
|
| 2
|
ггУусс
|
Расщепление по окраске семян 3 : 1
|
| 1
|
RRyycc
|
Круглые зеленые
|
3
|
Расщепления нет
|
| 2
|
Rryycc
|
семена, белые цветки
|
Расщепление по форме семян 3 : 1
|
| 2
|
rryycc
|
Шероховатые зеленые семена, белые цветки
|
1
|
Расщепления нет
|
* Эти гибриды были получены Г. Менделем в тригибридных скрещиваниях растений садового гороха, дающих круглые желтые семена и красные цветки с растениями, дающими шероховатые зеленые семена и белые цветки.
Заканчивая изложение экспериментов Г. Менделя, свидетельствующих о независимом характере перераспределения генов, остановимся кратко на перераспределении генов при других полигибридных скрещиваниях. Количество генных комбинаций в полигибридных скрещиваниях (например, тетрагибридных) возрастает с вовлечением в наблюдения каждой новой пары генов, ибо каждая новая пара приводит к двойному увеличению типов гамет, продуцируемых гибридами F1
, к тройному увеличению количества разных генотипов среди гибридов F2
и к увеличению в 4 раза количества возможных комбинаций при оплодотворении гамет, продуцируемых гибридами F3
.
В табл. 17 показана связь между количеством генов, вовлекаемых в скрещивание, и количеством фенотипических и генотипи-ческих групп среди гибридов.
Таблица 17
Генные комбинации в полигибридных скрещиваниях
| Количество генов, вовлекаемых в скрещивание
|
Количество фе-нотипических групп среди гибридов f2
(при полном доминировании)
|
Количество типов гамет, продуцируемых гибридами f1
|
Количество генотипически различных комбинаций
|
Количество возможных комбинаций гамет, продуцируемых гибридами f1
|
| 1
|
2
|
2
|
3
|
4
|
| 2
|
4
|
4
|
9
|
18
|
| 3
|
8
|
8
|
27
|
64
|
| 4
|
18
|
16
|
81
|
256
|
| n
|
2n
|
2n
|
3n
|
4n
|
Одна из важнейших особенностей полигибридных скрещиваний заключается в том, что увеличение количества генов, вовлекаемых в скрещивания, сопровождается снижением частоты появления среди гибридов F^ организмов исходных родительских типов и увеличением количества организмов, несущих рекомбинантные сочетания генов. Например, с вовлечением в скрещивание одного гена (одной пары аллелей) встречается по одному организму, похожему на организмы каждого исходного родительского типа среди 4 гибридов F2
, с вовлечением в скрещивание двух генов (двух пар аллелей) — среди 16 гибридов, с вовлечением в скрещивание трех генов (трех пар аллелей) — среди 64 гибридов и т. д. В тех случаях, когда количество генов (n), вовлекаемых в скрещивания, составляет десятки и даже сотни, общее количество генотипов (3n
), в том числе гомозиготных (2n
), которое может возникать в потомстве одного гетерозиготного организма, достигает огромных размеров. Благодаря независимому перераспределению генов (свободной рекомбинации генов) половой процесс создает огромное генетическое разнообразие организмов.
Подведем итоги рассмотренных в этом разделе экспериментов Г. Менделя. По данным дигибридных скрещиваний для передачи генов от одного поколения организмов к другому характерно их независимое расщепление и независимое перераспределение. Наряду с независимым расщеплением в дигибридных скрещиваниях выявляется и другая особенность. Растения F1
, возникшие в результате слияния гамет RY и гу при скрещивании исходных линий, дающих круглые желтые и шероховатые зеленые семена, в свою очередь продуцируют не только комбинации родительских гамет RY и гу, но и гаметы нового типа, несущие рекомбинантные фракции генов Ry и rY, притом в равном количестве. Что касается растений F1
, возникших после скрещивания линий, дающих шероховатые желтые и круглые зеленые семена, в результате слияния их гамет Yr и yR, то они кроме этих гамет тоже дадут гаметы, несущие рекомбинантные гены YR и уг, и тоже в равном количестве. Следовательно, образование гибридами F^ одинакового количества родительских и рекомбинантных гамет является неотъемлемой особенностью независимого расщепления и перераспределения двух пар генов.
Исследуя скрещивания, в которых исходные растения несли по три пары разных генов, Г. Мендель тоже обнаружил это явление, т. е. тригибриды F1
имея генотип RrYyCc, продуцировали гаметы восьми разных типов (RYC, RYc, RyC, Rye, rYC, rYc, ryC, rye), но в равных количествах.
Итак, генные пары двух скрещиваемых организмов расщепляются (сегрегируют) независимо одна от другой и перераспределяются, подвергаясь свободной рекомбинации между собой, причем гибридные организмы в процессе их гаметогенеза дают начало комбинациям родительских гамет и рекомбинациям гамет в равных количествах. Данное правило, установленное Г. Менделем, в настоящее время называют вторым законом наследственности (вторым законом Г. Менделя), законом независимого перераспределения генных аллелей или законом рекомбинации генов.
Цитологические основы расщепления генов (первого закона наследственности) определяются парностью хромосом, поведением хромосом диплоидных клеток при мейозе (спаривании и расхождении гомологичных хромосом) и последующим оплодотворением половых клеток, хромосомы которых несут по одному аллелю генных пар. У соматических клеток один аллель одной пары генов располагается на одном члене хромосомной пары, тогда как другой аллель представлен в другом члене хромосомной пары.
Расщепление генов происходит при гаметогенезе во время мей-отических делений. При мейозе хромосомы расходятся и проходят в разные гаметы, причем каждая гамета получает по одной хромосоме (гомологу) из пары хромосом. Независимое распределение генов также может быть объяснено поведением хромосом при мейозе (рис. 127).
Поскольку соматические клетки содержат по два набора хромосом, каждый из которых происходит от одного из родителей, то при мейозе расходятся каждая из хромосомных пар, а вместе с этим расходятся и генные пары. Важно то, что хромосомные гомологи затем перераспределяются и проходят в разные гаметы независимо один от другого. Но т. к. передвижение при редукционном делении двух отцовских или двух материнских хромосом к одному и тому же полюсу или одновременно прохождение в гаметы той или иной отцовской хромосомы вместе с какой-либо материнской хромосомой является делом случая, то гаметы несут отцовские и материнские хромосомы в самых различных смесях (сочетаниях). Следовательно, гаметы несут также разные сочетания отцовских и материнских генов. То, что аллели одной и той же хромосомной пары обязательно расходятся, определяется расположением их в одном месте (локусе) на хромосомной паре. Таким образом, закономерности расщепления и независимого перераспределения генов определяются передачей от поколений к поколениям хромосом.

Половые различия раздельнополых организмов связаны с различиями между их хромосомами, однако они по-разному проявляются у организмов разных видов.
У насекомых рода Protenor различия между хромосомами мужских и женских особей заключаются в том, что у мужских особей хромосомы представлены нечетным (меньшим) количеством (13), тогда как у женских особей — четным (14), т. е. количеством, большим на одну хромосому, которая является добавочной. Эта добавочная хромосома и определяет пол унаследовавшей ее особи в результате оплодотворения яйцеклетки мужской половой клеткой.
Однако у многих раздельнополых растений, беспозвоночных, животных, рыб, птиц и всех млекопитающих различия между хромосомами мужских и женских особей связаны не с количеством хромосом, а с их качественным составом. Например, соматические клетки мужских и женских особей D. melanogaster несут по четыре пары хромосом. Как у самцов, так и самок три пары хромосом являются одинаковыми и их называют аутосомами, но члены четвертой пары у самцов и самок неодинаковы по строению, ибо у самок оба члена этой пары являются прямыми палочковидными образованиями (Х-хромосомы), тогда как у самцов один член пары является прямым палочковидным образованием (Х-хромосома), а второй — изогнутым (Y-хромосома). Все одинаковые хромосомы у самцов и самок называют аутосомами (А), тогда как хромосомы Х и Y получили название половых хромосом. Все яйцеклетки плодовой мушки несут четыре хромосомы (ЗА+Х), из которых три являются аутосомами, четвертая — Х-хромосомой. Напротив, сперматозоиды также обладают четырьмя хромосомами, но они наполовину несут Х-хромосому (ЗА+Х) и наполовину — Y-хромосому (3A+Y). Оплодотворение любой яйцеклетки сперматозоидом, обладающим Х-хромосомой, дает начало зиготе женского типа (6А+ХХ), тогда как оплодотворение любой яйцеклетки сперматозоидом, обладающим Y-хромосомой, дает начало зиготе мужского типа (6A+XY).
У птиц самцы в соматических клетках содержат хромосомы XX, самки — хромосомы XY. Детерминирование пола у птиц сходно с этим явлением у D. melanogaster.
У человека хромосомы в соматических клетках женщин представлены 22 парами аутосом и парой половых хромосом XX (22АА+ХХ), тогда как в соматических клетках мужчин — 22 парами аутосом и парой половых хромосом XY (22AA+XY). Следовательно, каждая яйцеклетка несет 22 аутосомы и половую хромосому X, но сперматозоиды наполовину являются клетками, несущими 22 аутосомы и одну хромосому X, и наполовину клетками, несущими 22 аутосомы и одну хромосому Y. На хромосоме Y локализован ген длиной в 230 000 пар азотистых оснований, детерминирующий развитие тестисов (ZFY-секс-детерминирующий район Y). С геном ZFY связан также Y-хромосомный ген SRY, который тоже вовлечен в контроль развития тестисов и, кроме того, в контроль формирования костей. Кроме того в дополнение к гену существуют несколько аутосомных генов и один Х-сцепленный генный локус. Эти гены связаны с дефектами в развитии тестисов.
У человека и млекопитающих пол потомства также зависит от сперматозоида, оплодотворяющего яйцеклетку. Проникновение в яйцеклетку ядерного содержимого сперматозоида, несущего Х-хромосо-му, даст зиготу женского типа. В соматических клетках женских индивидов синтез мРНК идет лишь на одной хромосоме Х (в одних клетках на материнской хромосоме X, в других — на отцовской). Проникновение в яйцеклетку сперматозоида с Y-хромосомой даст зиготу мужского типа. Мужские индивиды являются гетерозиготны-ми (гетерогаметными) по полу, тогда как женские — гомозиготными (гомогаметными). Таким образом, пол генетически контролируется.
Однако у животных ряда видов пресмыкающихся пол контролируется не генотипом, а средой. Например, у многих черепах пол детерминируется температурой инкубации отложенных яиц (женские особи развиваются при повышенных температурах инкубации, мужские — при пониженных). А у аллигаторов, крокодилов и ящериц отдельных видов мужской пол детерминируется повышенными температурами. Детерминирование пола средой у позвоночных является исключением из общего правила генетического контроля пола.
Еще в 1910 г. Т. Морган (1886-1945) установил, что половые хромосомы участвуют также в контроле и ряда признаков. В его экспериментах, заключающихся в скрещиваниях белоглазых дрозофил-самок с красноглазыми дрозофилами-самцами, оказалось, что гибридное потомство состояло из красноглазых самок и белоглазых самцов. Однако среди гибридов встречались отдельные красноглазые самцы и белоглазые самки, появление которых было неожиданным, исходя из особенностей передачи хромосомы Х и нахождения на ней локуса гена, контролирующего белый цвет глаз. Анализируя этот случай, К. Бриджис (1916) предположил, что «необычные» белоглазые самцы несут по две Х-хромосомы, унаследовав их от своих «матерей», тогда как «необычные» красноглазые самцы имеют одну Х-хромосому, унаследовав ее от «отцов». Объясняя это предположение, К. Бриджис допустил, что исходные белоглазые самки продуцировали «необычные» гаметы, содержащие либо две хромосомы XX, либо ни одной хромосомы Х (наряду с гаметами, обычно содержащими по одной хромосоме X) в результате нерасхождения при мейозе их хромосом XX. Если такие «необычные» гаметы подвергались оплодотворению сперматозоидами красноглазых самцов, то это приводило к образованию зигот четырех типов: а) белоглазых самок, б) красноглазых самцов, в) особей с 3 хромосомами Х нежизнеспособных, г) особей без хромосомы Х (также нежизнеспособных). Назвав это явление первичным нерасхождение хромомсом (рис. 128). К. Бриджис затем цитологически показал, что в соматических клетках «необычных» белоглазых самок F, вместо хромосом XX присутствуют две Х и одна Y-хромосома (XXY).
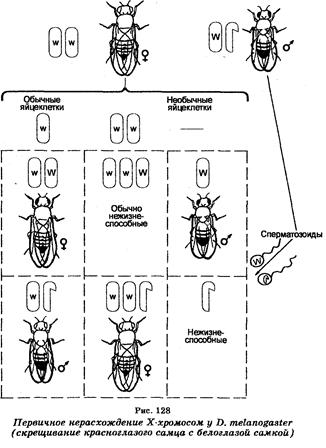
«Необычные» красноглазые самцы, не имеющие хромосомы Y, т. е. моносомики ХО, оказались в дальнейших экспериментах стерильными, тогда как белоглазые самки XXY были плодовитыми. Скрещивая последних с нормальными красноглазыми самцами и анализируя их потомство F1
, К. Бриджис открыл вторичное нерасхождение хромосом при мейозе у самок XXY (рис. 129), поскольку среди организмов F, 96% самок имели красные глаза и 4% — белые, тогда как среди самцов 96% были белоглазыми и 4% — красноглазыми. В этом случае при мейозе самки XXY продуцируют гаметы четырех типов, в которых хромосомы Х и Y распределяются неодинаково (яйцеклетки с хромосомой X, яйцеклетки, имеющие по одной хромосоме Х и Y, яйцеклетка с двумя хромосомами Х и яйцеклетки лишь с хромосомами Y).
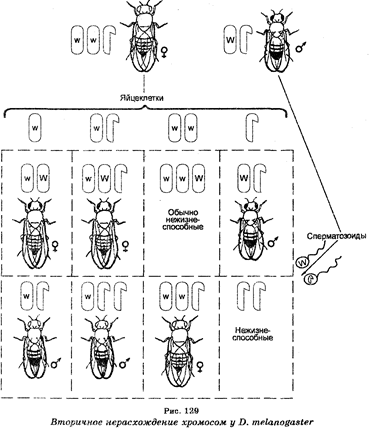
Оплодотворение яйцеклеток каждого из этих типов сперматозоидами нормального красноглазого самца приводило к образованию зигот восьми типов. Однако эти зиготы будут формироваться не с одинаковой частотой, поскольку мейоз у самок XXY связан с тем, что расхождение двух хромосом Х имеет место лишь в 92% овоцитов Объясняя вторичное нерасхождение хромосом, К. Бриджис допустил, что все белоглазые и отдельные красноглазые самки должны обладать не только парой хромосом X, но и хромосомой Y, тогда как красноглазые самцы в отличие от таких самцов, отмечаемых при первичном нерасхождении, должныиметь Y-хромосому и быть плодовитыми. Что касается отдельных белоглазых самцов, то они должны обладать одной хромосомой Х и двумя хромосомами Y. Эти допущения К. Бриджис доказал цитологически, найдя в соматических клетках дрозофилы хромосомы во всех сочетаниях, приведенных выше. Это открытие было показано на примере и других организмов (рис. 130).
 Открытие нерасхождения хромосом явилось окончательным доказательством в начале нашего века того, что гены локализованы на хромосомах. Однако теперь, когда с тех пор прошло много времени и когда к этому открытию возможен другой подход, можно сформулировать другое заключение. Фундаментальное значение открытия нерасхождения хромосом состоит в том, что оно положило начало изучению хромосомных мутаций и их механизмов. Можно добавить также, что оно означало объединение генетики и цитологии, т. е. формирование цитогенетики, оказавшейся в наше время исключительно плодотворной при изучении наследственности и наследственной патологии животных, растений и особенно человека (см. гл. XIII). Открытие нерасхождения хромосом явилось окончательным доказательством в начале нашего века того, что гены локализованы на хромосомах. Однако теперь, когда с тех пор прошло много времени и когда к этому открытию возможен другой подход, можно сформулировать другое заключение. Фундаментальное значение открытия нерасхождения хромосом состоит в том, что оно положило начало изучению хромосомных мутаций и их механизмов. Можно добавить также, что оно означало объединение генетики и цитологии, т. е. формирование цитогенетики, оказавшейся в наше время исключительно плодотворной при изучении наследственности и наследственной патологии животных, растений и особенно человека (см. гл. XIII).
Рассматривая 2-й закон наследственности, мы видели, что перераспределение генов имеет независимый характер и связано с прохождением отцовского и материнского членов каждой пары хромосом при мейозе в разные гаметы. Следовательно, в независимое перераспределение или рекомбинацию генов вовлекаются те наборы генов, которые располагаются на разных хромосомах. Такую рекомбинацию называют свободной рекомбинацией. Однако еще в 1910 г. Т. Морган установил, что гены, располагающиеся на одной хромосоме, сцеплены между собой, причем степень сцепления генов зависит от расстояния между ними. Следовательно, Т. Морган установил, что 2-му закону наследственности подчиняются лишь гены, локализованные на разных хромосомах. Однако значение этих наблюдений заключалось также и в том, что они послужили основой для формулирования в дальнейшем Т. Морганом и его сотрудниками представлений о линейном расположении генов на хромосомах. На основе этих представлений стали создавать генетические карты хромосом не только дрозофилы, но и других организмов.
Сцепление генов хорошо изучено в случаях многих видов животных и растений, показано, что оно присуще как женским, так и мужским особям. Чтобы лучше понять природу сцепления, рассмотрим его на описанном шведским генетиком Хатчисоном примере сцепления генов, детерминирующего форму и окраску семян кукурузы.
Обозначим символом С доминантный аллель, детерминирующий окраску семян, символом с — рецессивный аллель, детерминирующий отсутствие окраски семян, символом S — доминантный аллель, детерминирующий нормальную форму семян и символом s — рецессивный аллель, детерминирующий морщинистость семян (рис. 131). Если скрещивать растения CS/CS и cs/cs, то гибриды F,, обладая генотипом CS/cs, будут давать окрашенные семена нормальной формы. В соответствии со вторым законом наследственности при независимом перераспределении генов С и S гибриды F1
должны были продуцировать гаметы CS, Cs, cS и cs, причем в равных количествах, оплодотворение которых гаметами организмов cess (скрещивание гибридов F1
с исходным родителем cess) должно было дать потомство четырех фенотипических типов в отношении 1:1:1:1. Между тем скрещивание гибридов F1
с исходными родительскими организмами cess приводило к получению организмов в совершенно иных количественных отношениях, а именно:
— растения CS/cs (окрашенные семена нормальной формы) — 4032,
— растения Cs/Cs (окрашенные сморщенные семена) — 149,
— растения cS/cs (бесцветные семена нормальной формы) — 152 и
— растения cs/cs (бесцветные сморщенные семена) — 4035. Легко видеть, что среди 8368 растений превалировали организмы, дающие окрашенные нормальные по форме и бесцветные сморщенные семена (4032 + 4035 = 8067, или 96,4%), т. е. характеризующиеся комбинациями родительских признаков. Что касается остальной части растений (149 + 152 == 301, или 3,6% от 8368), которые давали окрашенные сморщенные и бесцветные нормальной формы семена, то они обладали рекомбинантными признаками. Если бы растения этих четырех типов встречались в равных количествах, то это означало бы независимое перераспределение (рекомбинацию) генных пар С-с и S-s. Между тем полученные результаты свидетельствовали о том, что перераспределение этих генных пар является зависимым, ибо комбинации родительских генов встречаются чаще, чем в 50% случаев. Другими словами, последние сцеплены между собой в 96,4% случаев.
То, что данные две пары генов имеют зависимое распределение, нашло также подтверждение в экспериментах по скрещиванию кукурузы, одни из которых дают бесцветные нормальной формы семена, а другие — окрашенные сморщенные семена. Следовательно, любые родительские комбинации двух пар генов, локализованных на одной и той же хромосоме, благодаря сцеплению оказываются вместе в одинаковом количестве гамет, продуцируемых гетерозиготами.
Рассмотренный пример сцепления двух генов является самым простым. Между тем можно предположить далее, что если ген А сцеплен с генами В и С, тогда последние также сцеплены между собой. Изучение сцепления генов у многих организмов путем скрещивания и определения независимого или зависимого (сцепленного) характера в распределении их генов позволило установить, что сцепление встречается между многими генами, а сцепленные гены составляют группы сцепления. Следовательно, геномы состоят из групп сцепленных генов или просто групп сцепления, причем количество групп сцепления обычно соответствует количеству хромосомных пар. У дрозофилы, имеющей 4 пары хромосом, установлено 4 группы сцепления, у кукурузы — 10 хромосомных пар и 10 групп сцепления, у садового гороха — 7 пар хромосом и 7 групп сцепления.
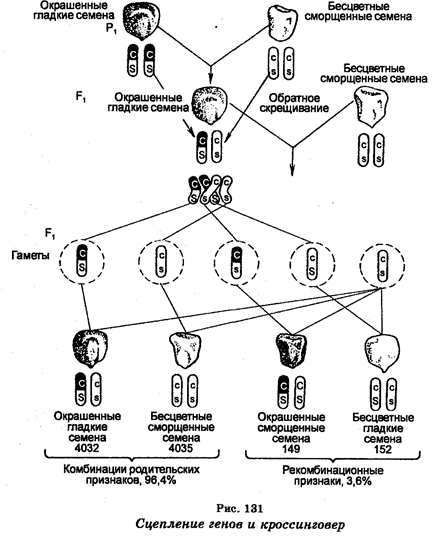
В случае животных, у которых пол детерминируется генетически, следует рассматривать гены, расположенные на Х- и Y-xpo-мосомах, в качестве самостоятельных групп сцепления.
Как мы уже отмечали, в соответствии с заключением Т. Моргана гены сцеплены тогда, когда локализованы на одной хромосомной паре. Допуская, что хромосомы остаются интактными при вступлении их в гаметы, локализованные на них гены всегда наследуются вместе. В этом случае можно говорить об их полном сцеплении. Однако, рассматривая сцепление генов у кукурузы, мы видели, что полного сцепления генов не бывает, поскольку происходит формирование гамет не только двух родительских типов, но и гамет рекомбинантных типов (вследствие рекомбинации генов). Об отсутствии полного сцепления свидетельствуют также данные, полученные при изучении других организмов, механизм этого явления заключается в том, что в процессе гаметогенеза хромосомы клеток могут подвергаться разрывам в одном или нескольких местах, а сегменты, образующиеся в результате разрыва одной хромосомы, могут смыкаться с сегментами гомологичной хромосомы при условии, что в последней тоже были разрывы, причем в аналогичных местах. Как мы видели, гены С и S у кукурузы в 97% случаев (гамет) остаются сцепленными в родительских комбинациях и примерно в 3% случаев (гамет) они не связаны между собой и находятся в рекомбинантных сочетаниях. Обмен между хромосомными сегментами гомологичных хромосом, сопровождаемый рекомбинацией сцепленных генов, получил название кроссинговера (Т. Морган), а явление, обусловленное этим механизмом, называют генетической рекомбинацией. В результате рекомбинации из двух исходных комбинаций генов создается новая комбинация. Возвращаясь к случаю рекомбинации генов у кукурузы, можно сказать, что кроссинговер произошел в сегменте хромосомы между локуса-ми (местами), занимаемыми генами С и S, вследствие двух разрывов в этих участках хромосомы у отдельных клеток.
Кроссинговер начинается с того, что гомологичные хромосомы спариваются. После этого каждый гомолог спаренных хрмосом расщепляется на две хроматиды, удерживаемые центромерой, причем между двумя хроматидами из четырех устанавливаются так называемые хиазмы (Х-образные фигуры или перекресты). В дальней  шем в этих двух хроматидах происходят разрывы, за которыми наступает воссоединение концов разорванных хроматид. Благодаря разрыву и воссоединению сегментов происходит формирование новых хроматид (рис. 132). Важно подчеркнуть, что хиазмы обусловливают перекресты лишь двух хроматид из четырех, не нарушая при этом структуры остальных двух хроматид, вследствие чего кроссинговер захватывает только две хроматиды. Благодаря этому сформированные в процессе мейоза хромосомы несут гены, располагавшиеся до мейоза на разных членах пар гомологичных хромосом. Классическая методика измерения сцепления заключается в скрещивании организмов, различающихся между собой по двум и более сцепленным генам, в получении гетерозиготных по этим генам гибридов F1
(например, АВ/ав или Ав/аВ), в обратных скрещиваниях гибридов F1
с гомозиготными по этим генам организмами (ав/ав) и в учете особенностей потомства, полученного после обратных скрещиваний. Установив количество особей с родительскими комбинациями генов и особей с рекомбинациями генов, определяют частоту рекомбинаций (в процентах к общему количеству организмов, полученных после обратных скрещиваний). Другие методы измерения сцепления основаны на результатах гибридизации соматических клеток или результатах клонирова-ния генов. шем в этих двух хроматидах происходят разрывы, за которыми наступает воссоединение концов разорванных хроматид. Благодаря разрыву и воссоединению сегментов происходит формирование новых хроматид (рис. 132). Важно подчеркнуть, что хиазмы обусловливают перекресты лишь двух хроматид из четырех, не нарушая при этом структуры остальных двух хроматид, вследствие чего кроссинговер захватывает только две хроматиды. Благодаря этому сформированные в процессе мейоза хромосомы несут гены, располагавшиеся до мейоза на разных членах пар гомологичных хромосом. Классическая методика измерения сцепления заключается в скрещивании организмов, различающихся между собой по двум и более сцепленным генам, в получении гетерозиготных по этим генам гибридов F1
(например, АВ/ав или Ав/аВ), в обратных скрещиваниях гибридов F1
с гомозиготными по этим генам организмами (ав/ав) и в учете особенностей потомства, полученного после обратных скрещиваний. Установив количество особей с родительскими комбинациями генов и особей с рекомбинациями генов, определяют частоту рекомбинаций (в процентах к общему количеству организмов, полученных после обратных скрещиваний). Другие методы измерения сцепления основаны на результатах гибридизации соматических клеток или результатах клонирова-ния генов.
На степень сцепления генов влияют различные факторы. Известно, что частота кроссинговера снижается с возрастом организмов, под воздействием ионизирующей радиации и других сильнодействующих факторов.
Кроссинговер, как уже отмечено, имеет место лишь в процессе гаметогенеза при мейозе. Однако он может происходить и в соматических клетках. Соматический кроссинговер установлен у растений и животных многих видов. У организмов, размножающихся только половым путем, результаты соматического кроссинговера не наследуются.
Изучение генетического материала на молекулярном уровне привело к выводу, что рекомбинация сцепленных генов представляет собой взаимодействие между гомологичными молекулами ДНК, конечным результатом которого является формирование структуры, построенной из частей каждого родительского гомолога. Представления о молекулярных механизмах генетической рекомбинации отражены в моделях «копирующего выбора» (copying-choice) и «разрыва-воссоединения» (breake-reunion).
Модель «копирующего выбора» была сформулирована еще в 1931 г. и в первоначальном варианте сводилась к допущению связи между репликацией и рекомбинацией генов. В последующем на основе этой модели стали считать, что рекомбинантные молекулы ДНК не содержат нуклеотидов, происходящих от ДНК родителей, они формируются заново, причем таким образом, что после спаривания гомологичных хромосом в качестве шаблона вначале используется ДНК одного родителя, затем в качестве шаблона — ДНК другого родителя. Следовательно, после репликации рекомбинантные цепи ДНК представляют собой по существу реплики определенного района одной родительской цепи и реплики определенного района другой родительской цепи ДНК.
Модель «разрыв-воссоединение» окончательно была сформулирована почти тогда же, но ее содержание определилось благодаря данным о месте и времени осуществления кроссинговера, т. е. о поведении гомологичных .хромосом при мейозе. В соответствии с этой моделью на молекулярном уровне рекомбинантная молекула ДНК строится прямым образом из сегментов родительских ДНК, причем ее построение осуществляется в результате разрыва цепей рекомбинирующих родительских молекул ДНК и последующего воссоединения образующихся сегментов двухцепочечных молекул ДНК.
Каждая из моделей имеет фактическое обоснования, но модель «копирующего выбора» не совсем согласуется с моделью строения и консервативной репликации ДНК. Напротив, модель рекомбинации «разрыв-воссоединение» согласуется с представлениями о полуконсервативной репликация ДНК. Рекомбинация молекул ДНК состоит из ряда последовательных стадий в виде разрыва, синтеза и воссоединения родительских тяжей, причем известны данные, позволяющие предполагать полярный характер рекомбинации в пределах малых районов хромосом (поляронов), равных по длине одному или нескольким генным локусам.
На основе этого предполагают, что хромосома разделена на несколько сегментов (поляронов), концы которых (любой из двух) определяют начало разделения полинуклеотидных цепей ДНК. Разделение исходных молекул, предшествуя синтезу и формированию гибридных молекул ДНК, может происходить как в одном направлении, т. е- брать начало от одного конца полярона, так и в двух направлениях (с обоих концов полярона). Больше того, предполагают даже, что рекомбинация очень сходна по своему механизму с процессом транскрипции генетической информации (образования мРНК) в смысле определения конца гена, с которого начинается разделение ДНК, и что в рекомбинации также работает ген-оператор, сходный с оператором оперона.
Анализ всех известных генотипических различий в генетической рекомбинации у организмов-эукариотов свидетельствует о том, что они обусловлены инбридингом, межвидовыми скрещиваниями, видовыми, популяционными и индивидуальными особенностями. Видовые, индивидуальные и другие особенности касаются уровня рекомбинации. Известны также специфические гены, мутации которых поражают способность к рекомбинации. Они гены у эукариот влияют на спаривание хромосом в мейозе, вследствие чего последние теряют способность к формированию пар в первой профазе мейоза, или на формирование хиазм. У бактерий известно несколько генов гес, продукты которых контролируют генетическую рекомбинацию бактерий. У фагов также обнаружены генетические системы, контролирующие их рекомбинацию. В совокупности все эти данные свидетельствуют о подверженности рекомбинаций генетическому контролю.
Генетические рекомбинации детерминируют рекомбинативную (комбинативную) изменчивость организмов.
Представления о расположении генов на хромосомах (в группах сцепления) сводятся к тому, что они располагаются в линейном порядке, причем, чем больше расстояние между генными локусами, тем большей является частота кроссинговера между ними и наоборот, линейный порядок генов характерен для групп сцепления всех организмов, включая человека, и определяет принципы построения генетических карт хромосом, которые представляют собой графическое изображение расстояний между генами в группах сцепления. Эти расстояния выражают в процентах рекомбинации, поэтому они являются генетическими. Однако их измеряют и в единицах длины ДНК. На основе представлений о линейном расположении генов строят также цитологические карты хромосом, которые позволяют представить локализацию многих генов в физических районах хромосом. В последние годы эти классические методы построения генетических и цитологических карт дополнены секвенированием генов и составлением физических карт.
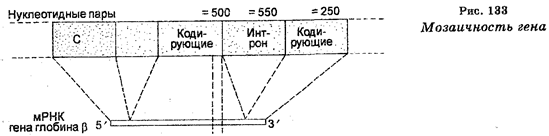
В конце 20-х гг. в нашей стране в коллективе А. С. Серебровского (1892-1948) возникла идея о дробимости гена. Тогда же Н. П. Дубинин открыл у дрозофилы явление ступенчатого алле-лизма и на основе результатов изучения этого явления сформулировал представления о сложном строении генов. Эти представле-. ния указывали на то, что линейный порядок характерен не только для расположения генов на хромосомах, но и для организации генетического материала внутри генов. В 50-е гг. в Англии Г. Понтекорво и его сотрудники установили, что у аспергилл ген состоит из многих мутационных мест (сайтов), разделяемых рекомбинацией. Тогда же С. Бензер (США) показал, что у фага Т4 функциональной единицей генетического материала является наименьший сегмент ДНК (800-1200 пар оснований), мутация которого сопровождается мутантным фенотипом. Этот сегмент был назван цистроном.
Приведенные выше данные показали, что наименьшей генетической единицей является пистрон, который, как стали считать, детерминирует синтез одного полипептида. У прокариот гены функционально активны на всем протяжении. Однако у эукариот сегменты ДНК, соответствующие индивидуальным генам, характеризуются мозаичностыо (рис. 133). Эта мозаичность определяется наличием в генах эксонов, кодирующих белки, и интронов, лишь переписываемых в мРНК, но не транслируемых. Во многих генах обнаружено по нескольку интронов (см. гл. XII).
Как отмечено выше, термин «ген» впервые для обозначения единиц наследственности был введен в литературу в 1909 г. английским генетиком У. Бэтсоном. С самого начала развития хромосомной теории наследственности представления о генах сводились к тому, что последние являются не только единицами наследственности, но и физическими единицами, располагающимися в линейном порядке на хромосомах. В рамках этих представлений в 1950 г. всеобщее признание имели следующие положения:
1. Ген является крайней единицей структуры. В соответствии с этим положением признавали, что ген нельзя разделить на субъединицы разрывом хромосом или рекомбинацией.
2. Ген является крайней единицей функции (фенотипического различия). Это положение означало, что ген детерминирует одиночную специфическую функцию в метаболизме или развитии организмов, причем любая специфическая функция не может быть «разделена» на субфункции.
На основе этих положений в то время была сформулирована гипотеза «один ген — один фермент». Между тем позднее оказалось, что белки имеют сложную полипептидную природу. В частности, известны белки, полипептидные цепи которых различны. Например, фермент Е. coli триптофансинтетаза, которая катализирует накопление индола и серина, являющихся предшественниками трип-тофана, сформирован таким образом, что одна часть молекулы имеет участок (называемый А), который соединяется с индоловым компонентом индоглицерофосфат, тогда как другая часть имеет участок (называемый В), соединяющийся с сериновым компонентом (серии + + пиридоксальфосфат). Синтез частей А и В-триптофансинтетазы детерминирован различными генами.
Данные о тонком строении генов свидетельствуют, что гипотеза «один ген — один фермент» сохраняет свое содержание и остается справедливой, но лишь в случае ферментов, представляющих собой одиночный полипептид. Данные о тонком строении генов лучше всего укладываются в постулаты «один ген — РНК одного типа, одно семейство родственых полипептидов» или «один ген — одна мРНК, один полипептид». Однако этот постулат имеет исключения, связанные с особенностями фенотипического проявления некоторых генных мутаций. В частности, недавно установлено, что мутации в тиро-зинкиназном протоонкогене человека сопровождаются одновременными изменениями четырех фенотипических функций, т. е. одновременным развитием четырех болезней (семейной мозговой ти-роидной карциномы, множественной эндокринной неоплазии типа 2А, более острой множественной эндокринной неоплазии типа 2В и болезнью Гиршенранга, характеризующейся отсутствием парасимпатической иннервации толстого кишечника). Следовательно, эти данные могут укладываться в положение «один ген — несколько признаков».
Как уже отмечено, большинство эукариотических генов являются однокопийными. Тем не менее известны гены, представленные множественными копиями. Например, гены, кодирующие белки-гистоны, у организмов ряда видов существует почти в тысяче копий, что объясняется необходимостью очень быстрого синтеза гистонов в период эмбрионального развития этих организмов. В большом количестве копий существуют гены, кодирующие синтез белка кератина (основного строительного материала перьев) у кур. Наконец, у всех исследованных позвоночных отмечено наличие значительного количества генов, кодирующих синтез четырех рибосомных РНК.
Размеры (длину) генов измеряют количеством входящих в его состав пар азотистых оснований или в единицах длины. Они весьма различны, но в среднем составляют примерно 1000—1050 пар азотистых оснований или около 357 нм (если исходить из расстояния в 0,34 нм между парами оснований в двойной спирали ДНК (0,34´1050 = 357 нм). Ген возможно охарактеризовать и в плане его массы. Поскольку молекулярная масса одной нуклеотидной пары в среднем равна 650, то масса среднего по размерам гена будет составлять 650´1050 =
= 680000. Однако известны у отдельных организмов и очень малые гены, размеры которых составляют всего лишь несколько десятков пар азотистых оснований. Например, у Е. faecalis длина отдельных генов составляет всего лишь 66 пар азотистых оснований, а у S. aureus — 78 азотистых оснований. Заметим, что средние размеры генов у Е. coil составляют, примерно, 1000 - 1100 пар азотистых оснований. Таким образом, в молекулярном плане ген представляет собой сегмент ДНК (последовательность азотистых оснований) определенной длины и массы, который контролирует синтез одного полипептида.
Однако гены кодируют не только полипептиды. Отдельные гены кодируют тРНК и рРНК. Поскольку одно азотистое основание тРНК или рРНК кодируется одним азотистым основанием ДНК, то гены, кодирующие тРНК и рРНК, характеризуются значительно меньшими размерами в сравнении с генами, кодирующими полипептиды.
В научной литературе ген часто аналогизируют с цистроном, под которым понимают наименьшую единицу наследственности. Между геном и цистроном действительно нет существенной разницы, в результате чего эти термины используют на равных правах.
Новейшие представления о тонком строении генов нашли отражение в созданных с помощью молекулярно-генетических методов физических, генетических и цитологических картах геномов растений и животных ряда видов, а также человека. Достоверность представлений о размерах и молекулярной организации генов полностью подтверждена данными генной инженерии.
1. Универсальны ли законы Г. Менделя, применимы ли они к человеку?
2. Как вы понимаете альтернативные (контрастирующие) признаки, аллель и пару аллелей?
3. Г. Мендель обнаружил, что скрещивания двух чистолинейных растений садового гороха, одно из которых давало семена круглой формы, другое — шероховатые, привели к получению гибридов, дававших семена круглой формы. Скрещивания гибридов первого поколения между собой дали начало растениям, семена которых имели круглую или шероховатую форму. Соотношение между этими растениями было 3:1.0 чем свидетельствовали эти результаты?
4. В чем заключались использованные Г. Менделем анализирующие скрещивания и какова их роль?
б. Можно ли предсказать наследование признака, контролируемого двумя аллелями?
6. Что такое наследственность, сцепленная с полом? Приведите примеры наследования пола и признаков, сцепленных с полом.
7. Что такое группы сцепления и как гены располагаются на хромосомах?
8. В 1952 г. в одном из районов Индии был пойман белый тигр (самец), которого затем скрестили с желтым тигром. Потомство от этого скрещивания приведено на рис. 134. Руководствуясь этой родословной, дайте ответы на следующие вопросы: а) Если допустить, что окраска шерсти контролируется парой аллелей, то является ли этот признак доминантным или доминантность отсутствует? б) Если окраска шерсти является примером наследования доминантности, то какая окраска (белая или желтая) является доминантным признаком? в) Определите и запишите, используя какие-либо символы, генотипы тигров Мохана, Самсона, Мохини, Малини и Кришны. г) Каково ожидаемое отношение потомства с желтым шерстным покровом к потомству с белой шерстью, полученного после скрещивания Самсона и Мохини? д) Какова вероятность рождения четырех желтых и одного белого тигрят в скрещиваниях Самсона и Мохини?
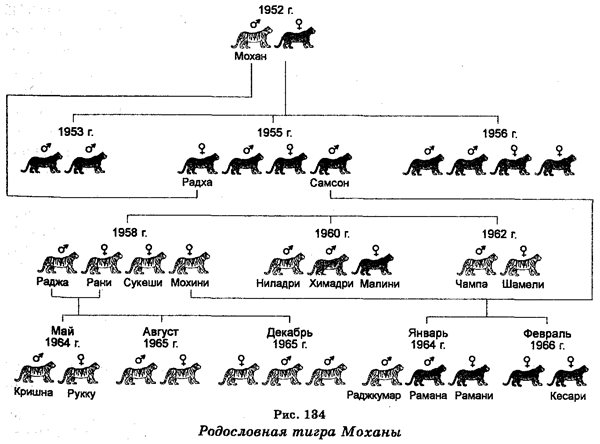
9. В семье N родились и выросли 16 детей. В табл. 18 приведены некоторые признаки, характерные для каждого из детей, когда они стали взрослыми. Используя эту таблицу, проделайте следующую работу:
а) Установите для каждого из четырех признаков тип наследования (доминантный, рецессивный, полу доминантный, отсутствие доминантности, множественность аллелей, полигенность и т. д.). б) Определите возможные генотипы и фенотипы по каждому признаку родителей N (отца и матери), в) Определите вероятность, с которой родились девочки и мальчики в этой семье.
10. Сформулируйте современную концепцию гена. Существует ли разница между геном и цистроном?
11. Что собой представляет мозаичность генов?
12. Гриб Pkytophthora castorium, который паразитирует на растениях многих видов, размножается бесполым путем с помощью подвижных зооспор, формируемых в спорангиях на мицелии, и половым путем благодаря ооспорам, которые формируются слиянием женских и мужских гамет, тоже формируемых на мицелии. Было найдено, что ооспоры, сформированные в культуре этого гриба после обработки его нитрозогуанидином, неодинаковы по своим свойствам. Одна четверть ооспор давала метионинзависимые колонии, тогда как другие три четверти ооспор напоминали родительский метиониннезависимый штамм. Анализ дальнейших генераций ооспор показал, что все метионинзависимые культуры продуцировали лишь метионинзависимые ооспоры, тогда как метиониннезависимые культуры первой генерации распадались на два класса. Около % культур характеризовались отношением около 3 : 1 метиониннезависимых и ме-тионинзависимых ооспор, тогда как отстающая треть вела себя в дальнейших скрещиваниях как фракция метиониннезависимых культур. Ооспоры в каждом случае давали начало колониям, напоминающим колонии тех культур, в которых они были сформированы независимо от того, нуждались они в метионине или нет. Каково ваше заключение о гаплоидном/ диплоидном статусе жизненного цикла грибов? В какое время развития у грибов происходит мейоз?
Таблица 18
Некоторые признаки, характерные для детей в семье
| ИМЯ
|
Способность ощущать вкус фенил-тиокарбамида
|
Группа крови по антигенам
|
. Группа крови по антигенам АВ, А, В, 0
|
Роет
(в см)
|
| Иван
|
+
|
MN
|
АВ
|
170
|
| Наталья
|
-
|
MN
|
0
|
160
|
| Елена
|
+
|
N
|
АВ
|
150
|
| Ирина
|
+
|
MN
|
А
|
170
|
| Валерий
|
+
|
N
|
В
|
180
|
| Владимир
|
+
|
М
|
А
|
170
|
| Андрей
|
-
|
MN
|
В
|
190
|
| Петр
|
-
|
N
|
0
|
180
|
| Николай
|
+
|
MN
|
о
|
180
|
| Клавдия
|
+
|
MN
|
АВ
|
160
|
| Павел
|
+
|
М
|
В
|
180
|
| Людмила
|
+
|
N
|
А
|
170
|
| Галина
|
-
|
N
|
0
|
160
|
| Виктор
|
+
|
MN
|
АВ
|
170
|
| Василий
|
+
|
MN
|
В
|
170
|
| Федор
|
+
|
М
|
А
|
165
|
13. Что вы понимаете под кроссинговером и генетической рекомбинацией? Каково происхождение кроссинговера?
14. Когда две рецессивные мутации айв теплены с рекомбинант-ной фракцией генов, будет формироваться два вида двойных гетерози-гот: цис (ав)/(++) и транс (а+)/(+в). Сформулируйте предсказание относительно фенотипических отношений между потомством, полученным самоопылением или сиб-скрещиванием по каждому из этих видов двойных гетерозигот.
Гаисинович А. Е. Зарождение генетики. М.: Наука. 1997. 195 стр
Дубинин Н. П. Генетика. Кишинев: Штииица. 1985. 533 стр.
Пехов А. П. Биология и общая генетика. М.: РУДН. 1993. 439 стр
Orel V. Gregor Mendel: The First Geneticist. Oxford University Press 1995 353 pp
Под действием генов (экспрессией, выражением генов) понимают способность их контролировать свойства или, точнее, синтез белков. Для действия генов характерен ряд особенностей, важнейшей из которых является их экспрессивность, под которой понимают степень фенотипической выраженности генов, т. е. «силу» действия генов, проявляющуюся в степени развития контролируемых ими признаков. Термин предложен Н. В. Тимофеевым-Ресовским (1900-1981). Экспрессивность генов не является постоянным свойством наследственности, ибо она очень вариабельна у растений, животных и у человека. Например, у разных людей проявляется по-разному такой признак, как способность ощущать вкус фенилтиокарбамида. Для одних людей это вещество является слишком горьким, для других его горечь кажется меньшей, что является результатом разной степени экспрессивности гена, контролирующего способность ощущать вкус этого соединения. Примером вариабельности экспрессивности генов является также экспрессивность доминантного гена, контролирующего ювенильную катаракту глаз человека. Экспрессия этого гена у разных индивидуумов варьирует от слабого помутнения хрусталика глаз до его полной непрозрачности.
С другой стороны, для действия генов у млекопитающих характерен так называемый геномный импринтинг, заключающийся в том, что два аллеля гена экспрессируются дифференциально, т. е. экспрессируется только один аллель из двух аллелей (отцовского и материнского), унаследованных от родителей. Например, у человека ген инсулинподобного фактора 2 нормально экспрессируется только из аллеля, унаследованного от отца, тогда как соседний с ним ген, кодирующий нетранслируемую РНК, экспрессируется только из аллеля, унаследованного от матери.
Важнейшей особенностью действия генов является также их пенетрантность, впервые описанная тоже Н. В. Тимофеевым-Ресовским. Под ней понимают частоту проявления того или иного гена, измеряемую частотой встречаемости признака в популяции, т. е. частотой встречаемости в популяции организмов, обладающих этим признаком. Перетрантность является статистической концепцией регулярности, с которой выражается (экспрессируется) тот или иной ген в популяции. Если какой-либо ген в популяции фенотипически выражается у индивидуумов, количество которых составляет 75% обследованных, то считают, что его пенетрантность тоже составляет 75%. Например, доминантный ген, контролирующий изменение цвета склеры глаз человека встречается у 90% людей. Следовательно, пенетрантность этого гена составляет 90%.
Экспрессивность и пенетрантность подвержены колебаниям. Причины этих колебаний не совсем ясны. Тем не менее обычно вариабельность в экспрессивности и пенетрантности генов объясняют либо модифицирующим влиянием других генов, которые получили название генов-модификаторов, либо совместным действием обоих этих факторов, а возможно и других факторов. Учет природы экспрессивности и пенетрантности генов имеет большое практическое значение в генетике человека, животных и растений. В случае человека эти явления учитывают при диагностике наследственных болезней, тогда как в животноводстве и растениеводстве они используются в селекции животных и растений.
Организмы наследуют от своих родителей не признаки и не свойства, как это долго считали ранее. Они наследуют гены, которые действуют на протяжении всей жизни организмов. В соответствии с существующими представлениями действие генов осуществляется через мРНК и приводит к образованию белков. Следовательно, мРНК являются первичными продуктами, тогда как белки являются конечными продуктами действия генов, т. е. результатом действия генов. Другими словами, гены контролируют структуру белков и ничего больше.
Поскольку материалом генов является ДНК, то в самом начале после открытия генетической роли ДНК возник вопрос и сводился к следующему: каким образом ДНК осуществляет свои функции в контроле синтеза белков? Ответ на этот вопрос заключается в том, что в ДНК содержится (закодирована) генетическая информация о синтезе белков, т. е. в ДНК содержится генетический код, под которым понимают систему записи в молекулах ДНК генетической информации о синтезе белков. Реализация генетического кода происходит в два этапа, один из которых называют транскрипцией, второй — трансляцией. Поток информации реализуется по схеме ДНК — РНК — белок. С тех пор как были открыты генетический код и механизмы его реализации (действия), эта схема получила название центральной догмы биологии.
Как отмечено выше, генетическая информация о синтезе белков содержится в молекулах ДНК и закодирована с помощью кода, получившего название генетического. Код, его структура и свойства были открыты в 60-х годах.
Структура генетического кода характеризуется тем, что он является триплетным, т. е. состоит из триплетов (троек) азотистых оснований ДНК, получивших название кодонов. Из 64 (4 х 4 х 4) возможных сочетаний нуклеотидов (кодонов) 61 является кодирующим, кодируя место аминокислоты в полипептидах. Три кодона, не кодируя места аминокислот в полипептиде, детермини- руют лишь остановку синтеза полипептида. Поэтому они названы стоп-кодонами или, иногда, терминирующими кодонами. Итак, один кодон кодирует место одной аминокислоты в полипептид-ной цепи (табл. 19).
Что касается свойств генетического кода, то их несколько. Код является неперекрывающимся, линейным, не имеющим пунктуации («запятых»), обеспечивающей свободные пространства между кодонами, и вырожденным.
Таблица 19
Генетический код
| Первое положение 5'-конец
|
Второе положение
|
Третье положение
|
| У
|
Ц
|
А
|
Г
|
| У
|
фенилаланин
|
серии
|
тирозин
|
цистеин
|
У
|
| фенилаланин
|
серии
|
тирозин
|
цистеин
|
Ц
|
| лейцин
|
серии
|
охре
|
терминатор
|
А
|
| лейцин
|
серин
|
амбер
|
триптофан
|
Г
|
| Ц
|
лейцин
|
пролин
|
гистидин
|
аргинин
|
У
|
| лейцин
|
пролин
|
гистидин
|
аргинин
|
Ц
|
| лейцин
|
пролин
|
глютамин
|
аргинин
|
А
|
| лейцин
|
пролин
|
глютамин
|
аргинин
|
Г
|
| А
|
изолейцин
|
треонин
|
аспарагин
|
серин
|
У
|
| изолейцин
|
треонин
|
аспарагин
|
серин
|
Ц
|
| изолейцин
|
треонин
|
лизин
|
аргинин
|
А
|
| метионин
|
треонин
|
лизин
|
аргинин
|
Г
|
| Г
|
валин
|
аланин
|
аспарагиновая кислота
|
глицин
|
У
|
| валин
|
аланин
|
аспарагиновая кислота
|
глицин
|
Ц
|
| валин
|
аланин
|
глютаминовая кислота
|
глицин
|
А
|
| валин
|
аланин
|
глютаминовая кислота
|
глицин
|
Г
|

Неперекрываемость генетического кода означает, что любое азотистое основание является членом только одного кодона. Ни одно азотистое основание не входит одновременно в два кодона. Например, в последовательности ААГАУАГЦА имеется три кодона ААГ, АУА, ГЦА, но не перекрывающиеся кодоны ААГ, АГА, ГАУ и т. д.
Код является линейным по той причине, что молекулы ДНК являются линейными полимерами. Кодоны в виде триплетов азотистых оснований следуют вдоль молекулы ДНК без перерывов в направлении от 5'-конп:а к 3'-концу, причем между кодонами нет свободных пространств, нет пунктуации.
Вырожденность кода определяется тем, что место в полипептиде одной и той же аминокислоты может кодироваться одновременно несколькими кодонами, но не совместно, а раздельно. Это распространяется на все аминокислоты, кроме метионина и триптофана, которым соответствуют одиночные кодоны (рис. 135).
«Мост» между геном (кодонами) и белком обеспечивается РНК. Точнее, информация, закодированная в последовательности азотистых оснований ДНК, вначале переносится от ДНК к матричной РНК (мРНК). Этот этап переноса информации носит название транскрипции и происходит у прокариотов в нуклеоиде, а у эукарио-тов — в ядре. Что же касается перевода информации с мРНК в белки, то этот этап декодирования получил название трансляции.
Транскрипция — первый этап в передаче генетической информации, сущность которого заключается в синтезе мРНК, т. е. в«перепи-сывании» генетической информации в молекулы мРНК (рис. 136). Транскрипция начинается с фиксированного пункта и заканчивается также в фиксированном пункте. Основными структурами, которые участвуют в транскрипции, являются ДНК-матрица (цепь ДНК), РНК-полимераза и хромосомные белки (гистоновые и неги-стоновые).
Молекулы мРНК составляют около 3% общей клеточной РНК. Они очень нестабильны. Период их полужизни очень краток. У прокариотов он составляет 2—10 минут, у млекопитающих и человека — около 12-16 часов, у некоторых других эукариотов — даже несколько недель. У прокариотов молекулы мРНК являются непосредственными продуктами транскрипции. Напротив, у эукариотов они являются продуктами процессинга первичных РНК-транскриптов (см. ниже).
Синтез молекул мРНК происходит в ядре клетки и очень сходен с репликацией ДНК. Отличие заключается лишь в том, что в качестве матрицы (шаблона) для копирования цепи мРНК используется лишь одна цепь ДНК. При этом копирование мРНК может начаться с любого пункта одиночной цепи ДНК, к которому прикрепляется РНК-полимераза и который называют промотором. Однако возможны случаи, когда два даже соседних гена могут транскрибироваться с разных цепей. Таким образом, для транскрипции может быть использована любая из двух цепей ДНК (рис. 137), но в любом случае одна из цепей транскрибируется одними РНК-по-лимеразами, другая — другими РНК-полимеразами, причем выбор цепи ДНК для транскрипции определяется промоторной последовательностью, которая задает направление движения РНК-полиме-разы (рис. 138).
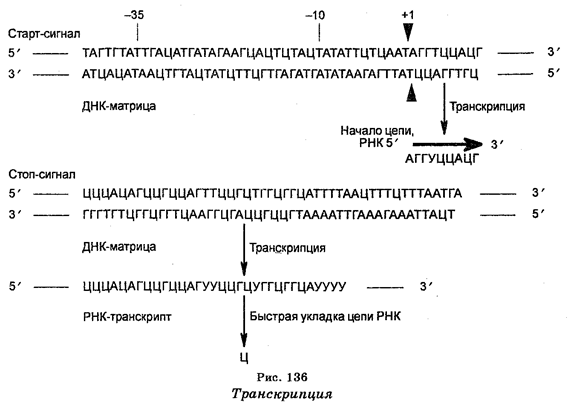
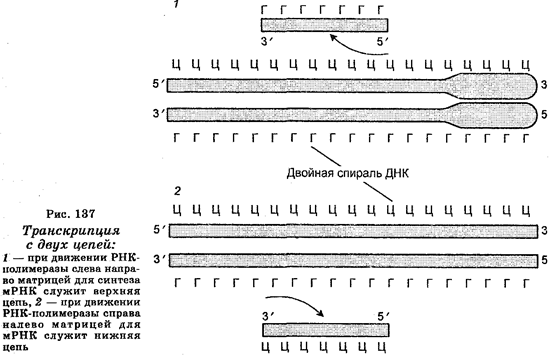
Поскольку обе цепи ДНК имеют противоположную полярность, а цепи РНК растут лишь в направлении от 5'-конца к 3'-концу, то транскрипции на каждой из цепей ДНК проходят в противоположных направлениях. Выбор цепи ДНК для транскрипции определяется содержанием последовательностей на ее промо-торных участках (местах присоединения РНК-полимеразы). Цепь, которая содержит те же последовательности, что и мРНК, называют кодирующей, а цепь, обеспечивающую синтез мРНК (на основе комплементарного спаривания) — антикодирующей. Из-за считывания кода с мРНК для его записи используют основания не А, Г, Т, Ц, а А, Г, У, Ц. Далее, мРНК не остается комплементарно связанной с ДНК-шаблоном, т. к. она освобождается от ДНК, которая затем восстанавливает свою двойную структуру. Наконец, молекулы мРНК значительно короче цепи шаблона ДНК. В одной эукаритической клетке количество молекул мРНК доходит до 10 000 и более.
Однако наряду с молекулами мРНК на ДНК образуются и другие транскрипты. В частности, транскрибируются молекулы рибо-сомной и транспортных РНК, которые также имеют важное значение в реализации генетической информации. Все эти РНК называют еще ядерными. Размеры транскриптов (транскрибируемых молекул РНК) зависят от посылаемых с цепи ДНК-шаблона сигналов начала и остановки синтеза (кодонов инициации и терминации).
Наиболее обильными РНК в клетках всех видов являются молекулы рибосомной РНК (рРНК), которые выполняют роль структурных компонентов рибосом. У эукариот синтез рРНК контролируется огромным количеством генов (сотни-тысячи копий) и происходит в ядрышке. В клетках человека гены для рРНК локализованы на 13, 14, 15, 21 и 22 парах хромосом.
В меньших количествах в клетках обнаруживаются молекулы транспортных РНК (тРНК), которые участвуют в декодировании информации (трансляции).
Все РНК транскрибируются с ДНК, которая несет множественные копии соответствующих генов. Непосредственными предшественниками в синтезе РНК являются рибонуклеозидтрифосфаты, причем здесь действует то же правило спаривания оснований за исключением того, что кодируются лишь ограниченные сегменты цепи ДНК и что тимин в ДНК заменяется на урацил в РНК. Ура-цил спаривается с аденином таким же образом, как и тимин. Цепь РНК растет в направлении от 5'- к 3'-концу с освобождением пирофосфата.
Синтез РНК обеспечивается РНК-полимеразами. У прокариот синтез мРНК, рРНК и тРНК осуществляет лишь один тип РНК-полимеразы, количество молекул которой в клетках достигает до 3000 молекул. Каждая из молекул этой РНК-полимеразы состоит из шести полипептидов, какими являются субъединицы р' и (3 (м. м. 155 ПОО и 151 000 соответственно), двух субъединиц а м. м. 36 000 и еще двух низкомолекулярных субъединиц (8 и ю). Инициация транскрипции обеспечивается субъединицей 5 — РНК-полимеразы, которая является, по существу, фактором инициации транскрипции. Как отмечено выше, связывание РНК-полимеразы с ДНК происходит на участке, называемом промотором и содержащем старт-сигнал для синтеза РНК, и контролируется белковым фактором. У Е. coli промоторы содержат последовательность ТАТААТ (бокс Прибнау), отстоящую от сайта начала транскрипции мРНК на расстояние в шесть оснований. После присоединения к промотору РНК-полимераза раскручивает в этом участке двойную спираль ДНК, обнажая цепи, каждая из которых служит затем матрицей, на которой происходит спаривание комплементарных оснований ДНК и рибонуклеозидтрифосфатов. Как только произошло спаривание двух первых мономеров РНК-полимераза продвигается дальше, обнажая дальнейшие участки цепей ДНК и добавляя последующие мономеры РНК. Удлинение цепи РНК происходит до тех пор, пока РНК-полимераза на своем пути не встретит «стоп-сигнал» и не отделится затем как от ДНК-шаблона (матрицы), так и РНК.
Напротив, в клетках эукариот (в частности, у дрожжей и человека) существуют три РНК-полимеразы (I, II, III), представляющие собой сложные молекулы, содержащие по нескольку полипептид-ных цепей. Каждая из этих РНК-полимераз, прикрепляясь к про-мотору на ДНК, обеспечивает транскрипцию разных последовательностей ДНК. РНК-полимераза I синтезирует крупные рРНК (основные молекулы РНК больших и малых субъединиц рибосом). РНК-полимераза II синтезирует все мРНК и часть малых рРНК, РНК-полимераза III синтезирует тРНК и РНК 5з-субъединиц рибосом. Количество РНК-полимераз в клетках млекопитающих различно (около 40 000 молекул РНК-полимераз I и II и около 20 000 молекул РНК-полимеразы III на клетку).
Эукариотические РНК-полимеразы также характеризуются сложным строением. РНК-полимераза II многих организмов построена из 12 различных полипептидов, три из которых гомологич-ны субъединицам р', р и а РНК-полимеразы Е. coli. РНК-полимеразы I и III обладают 5 субъединицами, сходными с субъединицами РНК-полимеразы II. РНК-полимераза II инициирует транскрипцию, причем для этого требуется белок ДНК-геликаза, детерминируемая у дрожжей геном RA25, а у человека — геном XRB. Большинство эукариотических промоторов содержат последовательность ТАТА, локализованную на расстоянии от 30 до 120 оснований от сайта транскрипционного сайта. У эукариотов для связывания РНК-полимеразы с промотором необходимы специальные белки, выполняющие функцию факторов инициации транскрипции (TF I, TF II, и TF III для РНК-полимераз I, II и III соответственно).
Транскрипция у эукариот является более сложным процессом по сравнению с транскрипцией у прокариотов. Длина последовательностей РНК (транскриптов), синтезируемых той или иной РНК-полимеразой, доходит до 50 000 нуклеотидов и более, причем за одну секунду они удлиняются на 30 азотистых оснований. Однако будучи точными копиями транскрибируемых генов, формируемые первичные транскрипты являются гетерогенными, т. к. не на всем протяжении способны к трансляции. По этой причине транскрипты называют гетерогенной ядерной РНК (гяРНК) или про-мРНК.
Для того чтобы про-мРНК стала «зрелой» мРНК, она еще в ядре вовлекается в процессинг, который заключается в том, что из про-мРНК с помощью ферментов «вырезаются» нетранслируемые участки (интроны), после чего транслируемые участки (эксоны) воссоединяются (рис. 139). Воссоединение называют сплайсингом (от англ. splice — сращивать). В результате процессинга образуются непрерывные последовательности зрелой мРНК, которые по своим размерам значительно меньше молекул про-мРНК, т. е. являются более короткими (табл. 20). Молекулы гяРНК содержит обычно более 50 000 нуклеотидов, тогда как после сплайсинга мРНК содержит всего лишь 500-3000 нуклеотидов.
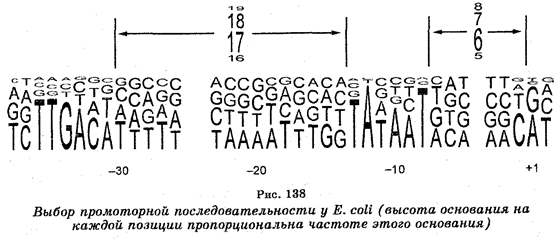
Подсчитано, что гяРНК составляет половину всей клеточной РНК, тогда как на долю зрелой мРНК приходится лишь 3% клеточной РНК.
Размеры интронов составляют обычно 80—1000 азотистых оснований.
 Биохимические механизмы сплайсинга определяются участием в этом процессе малых гетерогенных ядерных рибонуклеопроте-иновых частиц (sm RNP). Таких частиц выявлено несколько (V^, Ug, U^, Ug и другие), каждая из которых содержит по 90-150 нуклеотидов и по 10 разных белков. Все эти структуры контролируются в интерфазном ядре, причем их концентрация контролируется одной из киназ. Биохимические механизмы сплайсинга определяются участием в этом процессе малых гетерогенных ядерных рибонуклеопроте-иновых частиц (sm RNP). Таких частиц выявлено несколько (V^, Ug, U^, Ug и другие), каждая из которых содержит по 90-150 нуклеотидов и по 10 разных белков. Все эти структуры контролируются в интерфазном ядре, причем их концентрация контролируется одной из киназ.
Один и тот же транскрипт РНК может подвергаться разному сплайсированию, в результате чего сплайсированные участки зрелой мРНК могут кодировать разные белки, что свидетельствует в пользу эволюционного значения этой реакции.
Помимо модификации ядерной про-мРНК путем «вырезания» и сплайсинга ее сегментов нередко имеет место так называемое «редактирование» РНК, которое заключается в конверсии одного основания в другое в мРНК. Например, в клетках печени синтезируемый белок аполидопротеин имеет молекулярную массу порядка 512 000 дальтон, а в клетках кишечника лишь 242 000. Это является результатом конверсии цитозина в урацил (в клетках кишечника), что ведет к образованию стоп-кодона, а, следовательно, и к синтезу более короткого белка. Наконец, возможна модификация мРНК и путем посттранскрипционного добавления к ее 3'-концу от 30 до 500 нуклеотидов полиадениловой кислоты на расстоянии 15 нуклеотидов от последовательности ААУААА. По этой причине транскрипций заканчивается вдали от полиА-сигнала, а процессинг удаляет экстрануклеотиды перед полиА-добавлением (полиаденозином).
Синтезированная зрелая мРНК является первичным продуктом действия генов. В случае Е. coil после формирования она переходит затем из ядра в цитоплазму и на рибосомы, на которых спаривается с рРНК рибосомной субъединицы 30 S. Последовательность мРНК, которая связывается с рРНК рибосомной субъединицы 16 S, получила название последовательность Шайно-Дальгарно. Здесь мРНК служит матрицей для формирования полипептидных цепей на рибосомах. Считают, что в клетках имеется по 2000-3000 молекул мРНК, находящихся на разных уровнях синтеза и распада. В частности, установлены рибозимы с полинуклеотидкиназной активностью, способные катализировать АТФ-зависимое фосфорили-рование (см. гл. X).
Молекулы рРНК и тРНК также являются продуктами процессинга.
Открытие интронов поставило вопрос об их происхождении. В объяснении их происхождения используют две гипотезы. В соответствии с одной гипотезой интроны были представлены уже в предковых генах, в соответствии с другой интроны были включены в гены, которые оригинально были непрерывными.
Таблица 20
Результаты процессинга первичных генных транскриптов
| Ген
|
Длина гена
|
Длина мРНК
|
Количество интронов
|
| Р-глобин
|
1,5
|
0,6
|
2
|
| Инсулин
|
1,7
|
0,4
|
2
|
| Альбумин
|
25
|
2,1
|
14
|
| Каталаза
|
34
|
1,6
|
12
|
| Протеинкиназа С
|
11
|
1,4
|
7
|
Наряду с описанной схемой транскрипции у некоторых РНК-овых вирусов известна так называемая обратная транскрипция, при которой матрицей для синтеза ДЕК является РНК и которая осуществляется ферментом, получившим название обратной транскрип-тазы (ревертазы). Здесь реализация генетической информации идет по схеме РНК — ДНК — белок. Как свидетельствуют исследования, обратная транскриптаза найдена как у прокариотов, так и эукариотов. Считают, что ревертаза имеет очень древнее происхождение и существовала еще до разделения организмов на прокарио-ты и эукариоты.
Трансляция
является важной составной частью общего метаболизма клетки и ее сущность заключается в переводе генетической информации с мРНК, являющейся первичным продуктом действия генов, в аминокислотную последовательность белков. Трансляция происходит в цитоплазме на рибосомах и является центральным процессом в синтезе белков, в котором помимо рибосом участвуют мРНК, 3-5 молекул рРНК, 40-60 молекул разных тРНК, аминокислоты, около 20 ферментов (аминоацил-тРНК синтетаз), активирующих аминокислоты, растворимые белки, вовлекаемые в инициацию, элонгацию и терминацию полипептидной цепи.
Рибосомы состоят наполовину из белка и наполовину из рРНК (по 3-5 молекул на каждую рибосому). Размеры рибосом выражают в единицах скорости седиментации при центрифугировании (S). У прокариот размеры рибосом составляют 70 S, у эукариот — 80 S. Рибосомы построены из пары субъединиц (большой и малой), которые диссоциируют по завершению трансляции мРНК. У Е. coli большая субъединица (50 S) содержит две молекулы рРНК (5 S и 23 S) и 30 полипептидов, тогда как малая субъединица (30 S) содержит одну молекулу рРНК (16 S) и 19 полипептидов. У эукариот большая субъединица содержит три разных молекулы рРНК (58, 5,8 S и 20 S), тогда как малая субъединица — одну молекулу рРНК (18 S).
Транспортные (адапторные, растворимые) РНК являются малыми (5 S) молекулами длиной в 75—80 нуклеотидов. Их необходимость в трансляции определяется тем, что в отличие от ферментов, узнающих субстрат прямым образом, кодоны мРНК не способны прямо узнавать аминокислоты. Для этого должны существовать специальные адаптеры, узнающие и кодон и аминокислоту. Функцией таких адаптеров и обладает тРНК. Нуклеотиды тРНК построены из остатка фосфорной кислоты, углеводной части (рибозы) и основания. Главными нуклеотидами тРНК являются адениловый, гуаниловый, цитидиловый и уридиловый нуклеотиды. Вместе с тем одна из особенностей структуры тРНК заключается в том, что все они содержат по нескольку необычных, так называемых минорных нуклеотидов, причем последние являются химическими модификациями аденилового, гуанилового, питидилового и уридило-вого нуклеотидов (в основном в виде метилированных пуринов или нуклеотидов, обладающих метилированной рибозой). Некоторые из этих минорных нуклеотидов находятся в одном и том же районе у разных тРНК.
В молекуле тРНК вопреки ее одноцепочечной структуре имеет место комплементарное спаривание оснований, а это ведет к определенной конформации тРНК, заключающейся в том, что четыре ее сегмента формируют свернутую структуру (вторичную), которая имеет форму клеверного листа (рис. 140). В свою очередь эта структура подвергается дальнейшему свертыванию, превращаясь в так называемую многоскладчатую L-образную форму. Важнейшая особенность тРНК заключается в том, что по обоим концам L-образной фигуры сохраняются неспаренные нуклеотиды. Нуклеотиды одного конца фигуры формируют антикодон, а нуклеотиды другого конца (3'-конца) образуют последовательность (ЦЦА), обеспечивающую ковалентную связь с присоединяемой свободной аминокислотой.
Все тРНК характеризуются специфической последовательностью нуклеотидов. Их антикодоны комплементарны кодонам мРНК. Антикодоны располагаются в центре тРНК. Известно 55 антико-донов. Каждая тРНК способна присоединять и переносить только одну аминокислоту, но на каждую аминокислоту имеется 1-4 молекул тРНК.
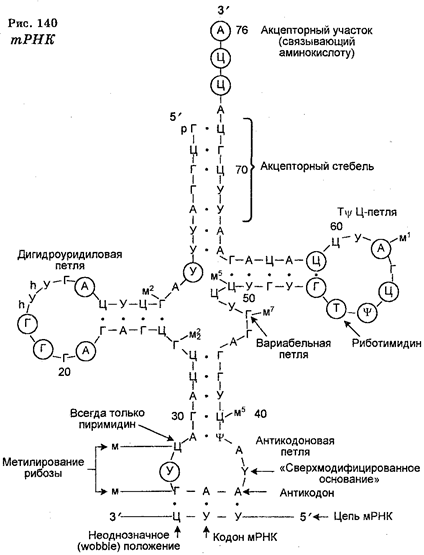
Первый этап трансляции происходит в цитоплазме и заключается в комбинировании каждой аминокислоты с АТФ (в образовании аденилированной аминокислоты) и специфическим ферментом аминоацил-тРНК-синтетазой (рис. 141). В результате этого устанавливается связь между фосфатом АМФ и карбоксильной группой аминокислоты (-Р-О-С-), которая приводит к образованию комплексов, состоящих из аминокислоты, АМФ и специфического фермента. Пирофосфаты в процессе образования этих комплексов удаляются. Следует заметить, что для каждой аминокислоты существует своя синтетаза, т. е. в клетках имеется 20 разных синтетаз.
Второй этап трансляции осуществляется также в цитоплазме. Поскольку аминоацил-тРНК-синтетазы специфически распознают аминокислоты и их тРНК, то второй этап состоит во взаимодействии образованных комплексов аминокислота — АМФ — специфический фермент (аминоацил-тРНК-синтетаза) со специфическими тРНК (один комплекс — одна тРНК). Поскольку на одном из концов имеется последовательность (конечное основание — аденин, а два предыдущих — цитозин и цитозин), то связывание одной аминокислоты со специфической тРНК происходит путем установления связи между рибозой конечного нуклеотида (адениловой кислотой) и карбоксильной группой аминокислоты (-С—О-С-). Вследствие этого взаимодействия происходит формирование так называемых аминоацил-тРНК, представляющих собой комплексы аминокислоты со специфической тРНК, и освобождение в процессе образования этих комплексов АМФ и фермента (аминоацил-тРНК-синтетазы). Следовательно, аминоацил-тРНК являются прямыми предшественниками полипептидного синтеза на рибосомах.
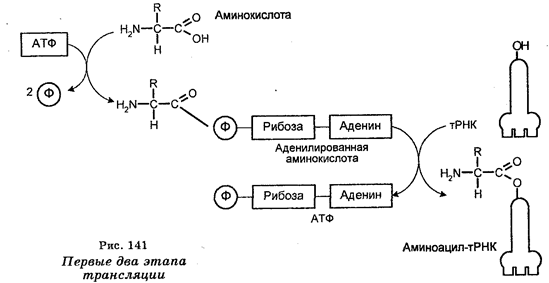
Осуществление этих двух этапов приводит к активации аминокислот. Одни синтетазы активируют 2'-гидроксил конечного основания тРНК, тогда как другие активируют 3'-гидроксил, а некоторые активируют и 2'-и 3'-гидроксилы. Однако эти различия не имеют значения, поскольку после освобождения аминоацильная группа на тРНК мигрирует взад и вперед.
Третий этап трансляции осуществляется на рибосомах и заключается в декодировании мРНК. В нем участвуют как мРНК, так и различные аминоацил-тРНК. Как отмечено выше, мРНК, отошедшая от ДНК в ядре и прошедшая через ядерную мембрану в цитоплазму, прикрепляется к РНК-последовательности меньшей (30 S) субъединицы рибосомы. Выше отмечено также, что последовательность мРНК, которая связывается с последовательностью рРНК рибосомной субъединицы 30 S, получила название рибосомосвязывающего сайта или последовательности Шайно-Дальгарно. Между тем каждая рибосома имеет два сайта, связывающих тРНК. Сайт А или аминоацил-тРНК-связывающий участок (акцепторный сайт), связывает приходящую аминоацил-тРНК, которая несет аминокислоту, предназначенную для добавления в растущую полипептидную цепь рядом с ранее добавленной аминокислотой. Сайт Р, или пептидил-тРНК-связывающий сайт (донорный сайт), связывает пептидил-тРНК, к которой прикреплен растущий полипептид. Специфичность связывания аминоацил-тРНК в этих сайтах обеспечивается кодонами мРНК, которые составляют часть сайтов А и Р. Это связывание происходит благодаря водородным связям, устанавливаемым между определенными основаниями (антикодоном) каждой аминоацил-тРНК и соответствующими основаниями (кодоном) мРНК. Первое и второе основания кодона всегда спариваются с третьим и вторым (соответственно) основаниями антикодона, тогда как третье основание кодона, если оно является урацилом, спаривается с гуанином или гипоксантином антикодона, если же оно является аденином — то с гипоксантином антикодона, но если гуанином — то с урацилом антикодона. Как уже отмечено, в обеспечении взаимодействия мРНК с тРНК участвует рРНК 16 S.
После связывания с мРНК аминоацил-тРНК помещают (включают) аминокислоты вдоль молекулы мРНК в последовательности,  соответствующей последовательности триплетов азотистых оснований в мРНК (рис. 142). Наращивание полипептидной цепи обеспечивается тем, что при синтезе белка рибосомы (полисомы) движутся вдоль цепи мРНК, т. е. рибосомы осуществляют считывание мРНК от одного конца до другого (рис. 143). Эффективность работы рибосом чрезвычайно велика. Например, у бактерий одна рибо-сома за 1 секунду присоединяет к полипептидной цепи свыше 20 аминокислот. Одновременно происходит формирование пептидных связей, обеспечиваемое несколькими ферментами-трансферазами, один из которых катализирует одновременно связывание аминоа-цил-тРНК с рибосомной, происходящее в присутствии ГТФ как кофактора. Каждая пептидная связь образуется ковалентным связыванием атома углерода карбоксильной группы первой аминокислоты с аминогруппой второй аминокислоты. При этом в процессе связывания происходит открепление тРНК первой аминокислоты от углерода карбоксильной группы своей аминокислоты. Каждая вновь добавляемая аминокислота встает на место, следующее за аминокислотой, добавленное ранее. Как видно, полипеп-тидная цепь наращивается с карбоксильного конца, а аминокислоты добавляются последовательно. Трансляция осуществляется в направлении от 5'- к 3'-концу полипептидного тяжа. соответствующей последовательности триплетов азотистых оснований в мРНК (рис. 142). Наращивание полипептидной цепи обеспечивается тем, что при синтезе белка рибосомы (полисомы) движутся вдоль цепи мРНК, т. е. рибосомы осуществляют считывание мРНК от одного конца до другого (рис. 143). Эффективность работы рибосом чрезвычайно велика. Например, у бактерий одна рибо-сома за 1 секунду присоединяет к полипептидной цепи свыше 20 аминокислот. Одновременно происходит формирование пептидных связей, обеспечиваемое несколькими ферментами-трансферазами, один из которых катализирует одновременно связывание аминоа-цил-тРНК с рибосомной, происходящее в присутствии ГТФ как кофактора. Каждая пептидная связь образуется ковалентным связыванием атома углерода карбоксильной группы первой аминокислоты с аминогруппой второй аминокислоты. При этом в процессе связывания происходит открепление тРНК первой аминокислоты от углерода карбоксильной группы своей аминокислоты. Каждая вновь добавляемая аминокислота встает на место, следующее за аминокислотой, добавленное ранее. Как видно, полипеп-тидная цепь наращивается с карбоксильного конца, а аминокислоты добавляются последовательно. Трансляция осуществляется в направлении от 5'- к 3'-концу полипептидного тяжа.
тРНК характеризуются исключительно высокой специфичностью, что проявляется в их антикодоновых последовательностях, соответствующих кодонам, доступности для распознания нужной аминоацил-тРНК-синтетазой и в точности связывания с сайтами А и Р на рибосомах.
Инициация, элонгация и терминация полипептидного синтеза находятся под генетическим контролем.
Наряду с кодонами, детерминирующими последовательность аминокислот, существуют кодоны, определяющие начало и конец чтения иРНК. В синтезе белка существенная роль принадлежит N-концевой аминокислоте формилметионину и его тРНК. N-формилметионил-тРНК (ОНС×NH×CH×(CH2×CH2× SCH3
)CO×О×тРНК) образуется в результате формилирования а-ами-ногруппы метионина NH2×СН(СН2×СН2×SCH3
)×СО×ОН в метионил-тРНК. Поскольку формулирование характерно только для метионина и катализируется ферментом трансформилазой, то считают, что формилметионин-тРНК является инициатором синтеза полипепти-да. Это означает, что все полипептиды в процессе синтеза начинаются с метионина. N-формилметионин является N-концевой аминокислотой всех белков.
Инициация полипептидной цепи у кишечной палочки начинается с образования комплекса между мРНК, формилметионин-тРНК и рибосомной единицей 30 8, которое обеспечивается факторами (белками) инициации IF1, IF2 и IF3, а также ГТФ. Этот комплекс вступает в комбинацию с 50 S-рибосомной единицей, в результате чего формилметионин-тРНК становится связанной с пептидиловым сайтом. Энергия для этого обеспечивается гидролизом одной молекулы ГТФ. Кодоны АУГ, ГУА и ГУГ на 5'-конце или рядом с ним направляют включение N-формилметионина в качестве N-концевой аминокислоты белка. Можно сказать, что эти кодоны являются специфическими инициаторами белкового синтеза. Наиболее активным является кодон АУГ.
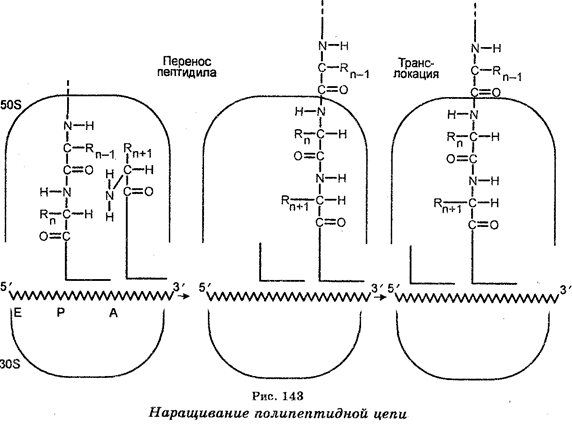
Элонгация (удлинение) полипептидной цепи обеспечивается белковыми факторами элонгации ef-TS и EF-Tu, а также гидролизом одной молекулы ГТФ, а движение молекулы мРНК с одного сайта рибосомы на другой обеспечивается фактором элонгации EF-G и гидролизом одной молекулы ГТФ. Каждый раз мРНК движется на три нуклеотида. У бактерий частота элонгации составляет 16 аминокислот в секунду. Это означает, что рибосомы двигаются вдоль мРНК со скоростью 48 нуклеотидов в секунду.
Терминация (окончание) синтеза детерминируется стоп-кодо-нами УАГ, УАА и У ГА. Когда один из этих кодонов подойдет к А-сайту рибосомы, то полипептид, тРНК в Р-сайте и мРНК освобождаются, а рибосомные субъединицы диссоциируют. Окончание синтеза белка связано с активностью белковых факторов освобождения — RF-1 и RF-2. Диссоциировав, рибосомные субъединицы начинают трансляцию другой молекулы мРНК. Большинство мРНК симультанно транслируется несколькими рибосомами (полисомами). Например, цепь гемоглобина из 150 аминокислот синтезируется на пентарибосомном комплексе. У прокариот синтез и трансляция мРНК происходят в направлении от 5'-конца к 3'-концу. Далее, у них нет ядерной мембраны. Поэтому трансляция мРНК начинается еще до завершения ее синтеза. Напротив, у эукариот транскрипция и трансляция разделены во времени, поскольку требуется время для перехода мРНК из ядра через ядерную мембрану в цитоплазму.
Синтез белков является исключительно точным механизмом. Обобщенные результаты исследований частоты ошибок в белковом синтезе показывают, что одна ошибка, т. е- одно включение «неправильной» аминокислоты, происходит лишь на каждые 10 000 включенных аминокислот. Точность механизма белкового синтеза обеспечивается точностью связывания аминокислот со своими тРНК и точностью спаривания кодонов мРНК с антикодонами тРНК.
Помимо генетического кода, который содержится в ядерной ДНК, существует генетический код, содержащийся в ДНК мито-хондрий животных и человека, а также в ДНК хлоропластов растений. В митохондриях и хлоропластах помимо ДНК существуют и другие структуры, которые в совокупности с ДНК образуют самостоятельный аппарат синтеза белков. Размеры митохондриальных рибосом очень варьируют. В частности, размеры митохондриальных рибосом человека составляют 60 S.
Для митохондриального генетического кода характерны те же структуры и свойства и те же механизмы транскрипции и трансляции, что и в случае ядерного генетического кода. Однако известны и отличия. В митохондриальной ДНК все нуклеотиды входят в состав кодонов, кодирующих либо белки, либо рРНК и тРНК. Для трансляции используется только 22 тРНК (в отличие от 31 тРНК в ядерном коде и 30 тРНК в хлоропластном коде), причем отдельные молекулы тРНК могут узнавать любое основание, находящееся в кодоне в третьем положении. Митохондриальная ДНК человека и других млекопитающих содержит 64 кодона, из которых 4 являются стоп-кодонами.
Известно содержание антикодонов всех 22 тРНК (табл. 21). Каждый антикодон в случае митохондриального генетического кода способен спариваться с несколькими кодонами мРНК. Например, антикодон УАГ спаривается с кодонами ЦУУ, ЦУЦ, ЦУА и ЦУТ, кодирующими лейцин. 22 антикодона тРНК спариваются с 60 кодонами иРНК. Установлено, что митохондриальные тРНК подвержены «редактированию» (модификации транспорта тРНК) путем полиаденилирования, в результате чего создаются антикодоны терминации.
Таблица 21
Антикодоны в митохондриальном генетическом
коде млекопитающих*
| ГАА
|
Фенилаланин
|
ГУА
|
тирозин
|
| УАА
|
Лейцин
|
ГУГ
|
гистидин
|
| УАГ
|
Лейцин
|
УУГ
|
глютамин
|
| ГАУ
|
Изолейцин
|
ГУУ
|
аспарагин
|
| ЦАУ
|
Метионин
|
УУУ
|
лизин
|
| УАЦ
|
Валин
|
ГУЦ
|
аспарагиновая кислота
|
| УГА
|
Серии
|
УУЦ
|
глютаминовая кислота
|
| УГГ
|
Пролин
|
ГЦА
|
цистеин
|
| УГУ
|
Треонин
|
УЦА
|
триптофан
|
| УГЦ
|
Аланин
|
УЦГ
|
аргинин
|
| ГЦУ
|
серии
|
| УЦЦ
|
глицин
|
* Антикодон ГАА спаривается с кодоном УУУ и УУЦ, антикодон УАА — с кодонами УУА и УУГ, антикодон УАГ — с кодонами ЦУУ, ЦУЦ, ЦУА и ЦУГ и т. д., за исключением того, что антикодон ЦАУ спаривается с кодонами АУА и АУГ
Генетический код ДНК и белоксинтезирующий аппарат хлоропластов несколько отличны от кода и белоксинтезирующего аппарата митохондрий.
Прежде всего хлоропластный код кодирует намного больше белков по сравнению с митохондриальным кодом. Рибосомы хлоро-палстов сходны с рибосомами кишечной палочки, а синтез полипептидной цепи начинается с N-формилметионина (как и у бактерий).
Генетический код ядерной ДНК универсален, т. к. он одинаков у всех живых существ, т. е. у всех живых существ используются одинаковые наборы кодонов. Признание универсального характера генетического кода является выдающимся современным доказательством единства происхождения органических форм (см. главы XIV, XV и XVI).
С тех пор как были определены основные черты структуры генетического кода, стали формулировать также гипотезы относительно его эволюции, причем к настоящему времени известно несколько таких гипотез. В соответствии с одной гипотезой первоначальный код (в примитивной клетке) состоял из очень большого количества двусмысленных кодонов, что исключало правильную трансляцию генетической информации. Поэтому в процессе эволюции организмов развитие генетического кода шло по линии сокращения ошибок в трансляции, что привело к коду в его современном виде. Напротив, по другой гипотезе код возник в результате сведения до минимума летальных эффектов мутации в процессе эволюции, причем селективное давление вело к устранению бессмысленных кодонов и к ограничению частоты мутаций в кодо-нах, изменения которых не сопровождались изменениями в последовательности аминокислот, либо сопровождались заменами лишь одной аминокислоты на другую, но функционально связанную. Развившись в процессе эволюции, код однажды стал «замороженным», т. е. таким, каким мы видим его сейчас.
В соответствии с третьей гипотезой предполагают, что ранний архетиповой код был дуплетным, состоя из 16 кодонов-дуплетов. Каждый из 15 дуплетов кодировал каждую из 15 аминокислот, из которых, как предполагают, состояли белки примитивной клетки, тогда как оставшийся свободным 60-й дуплет обеспечивал свободное пространство («брешь») между генами. В связи с установлением каталитической способности РНК и высокой концентрации РНК в рибосомах предполагают, что в примитивных клетках молекулы тРНК сами катализировали свое связывание с аминокислотами, а роль рибосом выполняли первые рРНК. Триплетный код возник тогда, когда в процессе эволюции образовались остальные пять аминокислот, причем его возникновение связано с добавлением третьего основания в каждый кодон.
Предполагают, что современный генетический код является результатом длительной эволюции примитивного кода, кодировавшего лишь несколько аминокислот, притом только несколькими триплетами, составленными из азотистых оснований двух типов.
В последующем эволюция кода заключалась в уменьшении количества бессмысленных триплетов и увеличении количества смысловых. Это привело к тому, что большинство триплетов стало «читаться». Завершающая стадия в эволюции кода была связана с увеличением количества аминокислот, подверженных «опознанию» соответствующими нуклеотидами (триплетами), а также с синтезом клетками соответствующих тРНК и активирующих ферментов. Когда количество и структура белков стали такими, что уже ни одна новая аминокислота не могла улучшить селективные преимущества организмов, код «заморозился» в его современном виде.
Что касается митохондриального кода, то его считают более примитивным по сравнению с ядерным. Предполагают, что, например, антикодон УАА в современном митохондриальном коде мог быть также и антикодоном архетипового кода для кодонов, в которых первые два основания являются У, а третье могло быть У, Ц, А или Г. Но можно предполагать, что митохондриальный код возник в результате упрощения бактериального кода, если признать происхождение митохондрий от бактерий. Оценивая особенности белкового синтеза, контролируемого митохондриальным генетическим кодом в сравнении о хлоропластным, остается неясным, почему хло-ропластный генетический код кодирует намного больше белков по сравнению с митохондриальным генетическим кодом.
Как видно, современные взгляды на происхождение и эволюцию генетического кода весьма противоречивы, ибо пока нет еще экспериментальных данных, которые можно было бы использовать для достаточного обоснования той или иной гипотезы.
Что понимают под генетическим контролем экспрессии или регуляции действия генов? Это понятие означает, что экспрессия гена или набора генов может избирательно увеличиваться или уменьшаться (индуцироваться или репрессироваться) селективно. Регулирующее действие осуществляют белки, которые могут вмешиваться в транскрипцию. На экспрессию оказывает влияние изменение уровня АТФ, но это соединение не является результатом.
Сведения о регуляторных механизмах экспрессии генов по большей части получены в результате изучения образцов контроля активности генов, распространяемых на последовательность реакции в биосинтезе микроорганизмами белков, на гены фага лямбда, 5 S-гены Xenopus, гены, обеспечивающие скрещивание дрожжей, и гены, вовлеченные в контроль развития эукариотов. Сравнение механизмов, контролирующих действие генов у разных организмов, показывают чрезвычайное разнообразие в этих механизмах. В этом убеждает рассмотрение наиболее изученных систем. В применении к бактериям известно два механизма, один из которых контролирует активность ферментов, тогда как второй — синтез ферментов (синтез специфических белков). Сущность контроля (регуляции) активности ферментов иллюстрируется примером биосинтеза изолейцина, ранним предшественником которого является треонин и превращение которого в изолейцин осуществляется в результате пяти последовательных реакций с участием ферментов. Если к культуре бактерий, обладающих самостоятельной способностью синтезировать аминокислоты, в том числе изолейцин, прибавить изолейцин, то это приводит к прекращению клетками синтеза данной аминокислоты. Ростовые потребности клеток в это время обеспечиваются лишь экзогенным изолейцином. Механизм этого явления заключается в ингибировании (подавлении) активности фермента, катализирующего превращение треонина в последующий предшественник изолейцина. Синтез восстанавливается лишь тогда, когда экзогенный изолейцин истощается в среде.
Уникальность этого явления связана с тем, что ингибитор (конечный продукт) и нормальный субстрат имеют различную структуру и не конкурируют за один и тот же сайт связывания на ферменте. Можно сказать, что фермент несет два сайта связывания, один из которых специфичен для субстрата, другой — для ингибитора. Нормально субстрат прикрепляется к активному сайту фермента. Однако если к этому специфическому сайту прикрепляется ингибитор, то наступает структурное превращение (транзиция) в ферменте, вследствие чего нормальный субстрат больше не прикрепляется, что блокирует активность фермента, катализирующего конец биосинтеза либо одну из его стадий. Это явление получило название аллостерической транзиции (рис. 144).
В основе аллостерического взаимодействия лежит любое измерение в активности фермента, вызываемое избирательным связыванием на втором сайте фермента, причем этот сайт не перекрывает сайта на ферменте для связывания субстрата. Фермент, по существу, становится химическим трансдуктором, позволяющим взаимодействие между двумя молекулами — ингибитором и субстратом, которое другим способом исключено. Определенные ферменты чувствительны к активированию при соединении их с эф- фекторной молекулой, отличной от каталитического субстрата. Кроме того, определенные ферменты чувствительны к активированию одним метаболитом и подавлению другим. Поскольку возможны мутации, которые могут поражать один ингибиторный сайт, не затрагивая другого, фенотипически они проявляются в резистентности клеток к ингибированию конечным продуктом и в выработке ими больших количеств конечного продукта. Таким образом, аллостерическая транзиция обеспечивает исключительно гибкую систему регуляции активности ферментов.
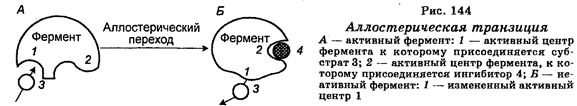
Синтез ферментов регулируется с помощью индукции и репрессии ферментов, заключающихся в стимуляции или подавлении синтеза специфических ферментов как ответной реакции на добавление в среду компонента, повышающего концентрацию эффектора в клетке.
Примером индукции ферментов является случай с ферментами бактерий, обеспечивающих утилизацию лактозы. Бактерии приобретают способность сбраживать лактозу после некоторого культивирования в присутствии этого углевода. Это определяется синтезом ими b-галактозидазы, которая расщепляет лактозу на глюкозу и галактозу, а также b-галактозидпермеазы и b-галактозидтранс-цетилазы, обеспечивающих проникновение субстрата в клетку и ацетилирование некоторых токсических галактозидов в направлении их детоксификации (соответственно). Следовательно, лактоза индуцирует синтез ферментов, причем этот синтез является координированным .
Примером репрессии ферментов может служить синтез трипто-фана, который образуется из антраниловой кислоты с участием ант-ранилатсинтетазы. Если бактерии посеять в среду, в которой в качестве источника азота и углерода содержатся NH4
Cl и глюкоза, соответственно, то они очень хорошо растут и самостоятельно синтезируют триптофан (как и другие аминокислоты). Но если в среду добавить экзогенный триптофан, то бактерии перестают синтезировать эту аминокислоту. Важно заметить, что добавление триптофана останавливает синтез всех ферментов, участвующих в его биосинтезе. Таким образом, конечный продукт подавляет весь биосинтез.
Опираясь на данные об индукции и репрессии белков, французские ученые Ф. Жакоб и Ж. Моно (1961) сформулировали модель генетического контроля синтеза белков, компонентами которой являются гены структуры, регуляции и операторные гены, а также цитоплазматический репрессор. По этой модели молекулярная структура белков определяется генами структуры, первичным продуктом которых является мРНК. Синтез мРНК может быть начат лишь на определенном пункте цепи ДНК (операторе), от которого может зависеть и транскрипция нескольких сцепленных структурных генов. Группа генов, транскрипционная активность которых координируется одиночным оператором, представляет собой опе-рон, являющийся единицей первичной транскрипции и единицей координированной экспрессии генов. Таким образом, бактериальные опероны транскрибируются в полицистронные мРНК. Существуют также гены-регуляторы. Под контролем того или иного гена-регулятора продуцируется цитоплазматический фактор-репрессор, который обладает реверсивной способностью связываться со специфическим оператором. Благодаря этому связыванию комбинация репрессора и оператора блокирует начало транскрипции всего оперона (формирование мРНК), контролируемой оператором и, следовательно, предотвращает синтез белков, управляемый структурными генами, принадлежащими оперону.
Репрессор обладает свойством специфически связываться (реагировать) с малыми молекулами (эффекторами). В случае индуцированных ферментных систем репрессор связывается с оператором и блокирует транскрипцию олерона. Присутствие эффектора (индуктора) связывает (инактивирует) репрессор, и это приводит к тому, что происходит транскрипция и трансляция генов оперона. Другими словами, репрессор, соединенный с эффектором, теряет родство к оператору и не связывается с ним, а это сопровождается активацией оперона. В случае репрессибельных ферментов репрессор сам по себе является неактивным, т. е. не имеет родства к оператору и не блокирует транскрипцию оперона. Он активируется лишь в результате комбинации (соединения) с конечным продуктом в биосинтезе, в результате чего блокирует транскрипцию оперона. Следовательно, транскрипция оперона происходит в отсутствие эффектора (конечного продукта), тогда как присутствие эффектора сопровождается ингибированием оперона.
Регуляторные механизмы в случае индуцибельных и репрессибельных систем имеют негативный характер, т. к. подавляют синтез специфических белков. Регуляторный механизм оперирует на генетическом уровне, контролируя частоту синтеза мРНК. Опероны классифицируют на катаболизирующие (индуцибельные) и синтезирующие (репрессибельные).
Примером катаболизирующих оперонов является лактозный оперон (рис. 145), в состав которого входят структурные гены z, у и a, кодирующие b-галактозидазу, b-пермеазу и b-трансацетилазу, соответственно, ген-регулятор lac 1, кодирующий синтез репрессоpa (белок мм. 37 200, состоящий из четырех идентичных единиц, содержащих по 377 аминокислотных остатков), промоторы P2
и Р1
, к которым присоединяется РНК-полимераза, и ген-оператор (О), управляющий функционированием структурных генов.

Механизм регуляции лактозного оперона заключается в том, что в отсутствие индуктора (лактозы) репрессор активен, т. е. продукт гена lac 1 связан с оператором O1
в соединении с двумя другими операторами (О2
и О3
) и это блокирует транскрипцию полигенной мРНК, т. е. предупреждает движение РНК-полимеразы с промотора. Операторы О2
и О3
, которых часто называют псевдооператорами, повышают связывание репрессора с оператором О1
. Промотор P1
частично перекрывается вторым промотором P2
, который в нормальных условиях не имеет существенного значения. РНК-полимераза может подходить к обоим промоторам, но основным все же является промотор Р1
, поскольку движение РНК-полимеразы к промотору P2
ингибируется белком CRP, который при этом стимулирует транскрипцию с промотора P1
. При наличии в среде индуктора репрессор теряет активность, т. е. связывается с индуктором, вследствие чего оператор становится свободным (деблокируется), и в результате движения РНК-полимеразы происходит транскрипция мРНК. При наличии в среде лактозы синтез ферментов увеличивается в 1000 раз. Инициация транскрипций требует наличия циклического АМФ и белка сгр. Следовательно, лактозный оперон контролируется негативно путем контроля частоты синтеза мРНК (рис. 146).
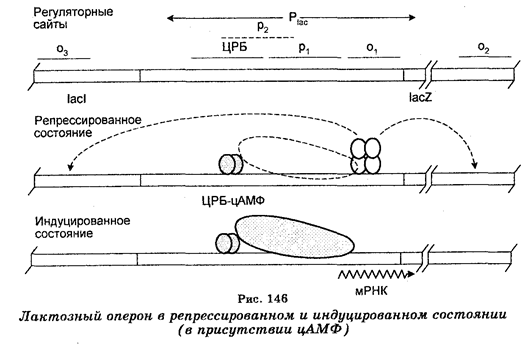
Однако возможна и позитивная регуляция лактозного оперона. Если вслед за индуктором в среду, где выращены бактерии,прибавить глюкозу, то наступает катаболитная репрессия (3-галак-тозидазы, сопровождающаяся снижением уровня цАМФ. Если же в культуру прибавить экзогенный цАМФ, то катаболитическая репрессия снимается и лактозный оперон будет функционировать нормально. Таким образом, цАМФ является позитивным регулятором (регуляция происходит в присутствии индуктора).
Примером биосинтезирующего оперона является триптофановый оперон, который состоит из 5 генов trpE, D, С, В и А, кодирующих ферменты, участвующие в биосинтезе триптофана (рис. 147). Первая химическая реакция, характерная для синтеза триптофана (как и для тирозина и фенилаланина), заключается в конденсации (накоплении) эритрозы-4-Ф и фосфоенолпирувата, необходимых для образования З-дезокси-арабино-гептулозоната-7-Ф (ДДГФ), катализируемого ДАГФ-синтетазой. Образующийся продукт (хоризмат) с помощью ферментов превращается в триптофан. Экспрессия этого оперона находится под негативным контролем репрессора, являющегося продуктом гена trp R. Репрессия конечным продуктом (трип-тофаном) заключается в том, что неактивный в свободном состоянии репрессор не мешает переходу РНК-полимеразы к промотору, т. е. не мешает транскрипции мРНК. Репрессор становится активным в результате связывания с конечным продуктом (триптофаном). Когда комплекс репрессор + триптофан связывает оператор, то это предупреждает преход РНК-полимеразы к промоторам, т. е. подавляет синтез мРНК. Более тонкий механизм заключается в том, что продукт гена trp R (репрессор) ингибирует синтез одной из ДАГФ-синтетаз, в частности, синтетазу АгоН, путем ингибирования ее мРНК, а следовательно, ингибирует и мРНК всего гистидинового оперона.
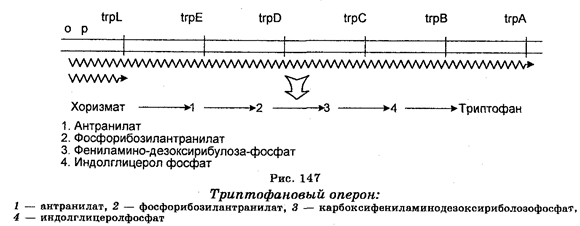
То, что существуют различные пути регуляции генов, подтверждается данными о регуляции оперона ara BAD, который ответственен за метаболизм арабинозы, встречающейся обычно в стенках клеток растений, а у микроорганизмов — являющейся одним из источников углерода и энергии. Арабиноза попадает в организм животных или человека с растительной пищей, где она метаболи-зируется микроорганизмами. Однако прежде она должна поступить в бактерии. Это поступление арабинозы в бактериальные клетки, т. е. транспорт арабинозы обеспечивается двумя независимыми системами, представляющими собой продукты генов ara Е и ara FG (рис. 148).
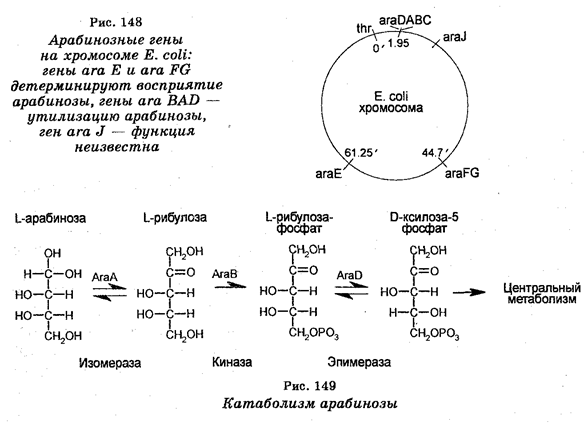
Система ara Е обладает низким родством к арабинозе, поэтому она используется лишь при очень высоких концентрациях этого углевода, тогда как система ara FG, обладающая высоким родством к арабинозе, используется при очень низких концентрациях углевода (порядка 10-7
М).
Гены, кодирующие ферменты, которые необходимы для катаболизма арабинозы, показаны на рис. 149. Катаболизм арабинозы начинается с того, что она вначале конвертируется синтезируемая под контролем гена ara А арабинозной изомеразой в L-рибулозу. Затем рибулоза фосфорилируется рибулозокиназой, являющейся продуктом гена ara В в L-рибулозу-б-фосфат. В свою очередь рибулозо-фосфат конвертируется в П-ксилозу-б-фосфат с помощью рибулозо-фосфатэпимеразы, являющейся продуктом гена ara D. Важно отметить, что гены транспортной системы, а также дополнительный арабинозоиндуцируемый ген и гены метаболизма арабинозы локализованы в разных районах хромосомной карты Е. coil. Все эти наборы генов регулируются позитивно арабинозой.
Репрессия метаболических и транспортных генов в ага-опероне осуществляется продуктом (белком) гена ara С. Этот белок является позитивным регулятором. В то же время белок гена ага С может и репрессировать экспрессию этих генов. Таким образом, этот белок существует в двух состояниях — индуцирующем и репрессирующем, индуцируя или репрессируя инициацию транскрипции (начало синтеза мРНК) арабинозного оперона. Индуцирующим свойством белок ага С обладает в присутствии арабинозы, позитивно действуя на один сайт, репрессирующим — в отсутствие этого углевода, действуя негативно на другой сайт (рис. 150).
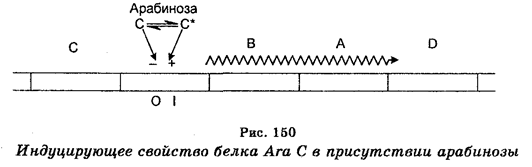
Индукция арабинозного оперона происходит в присутствии цАМФ и белка CRP, являющегося рецептором для цАМФ. Главная роль этого белка заключается в позволении индукции арабинозного оперона только в отсутствие глюкозы. Это получило название катаболитной репрессии.
Опероноподобная организация генетического контроля действия генов показана у фага лямбда, вируса иммунодефицита человека и многих других микроорганизмов.
Считают, что в геноме эукариотических клеток содержится 10 000-50 000 генов-регуляторов. Регулирующие белки, синтезируемые под контролем этих генов, связываются со специфическими последовательностями ДНК вблизи регулируемых генов.
Для некоторых организмов — эукариотов (например, дрожжей) также характерна опероновая организация функционально связанных генов.
Рассмотрение вопроса о регуляции действия генов у организмов на более высокой ступени эволюционной лестницы показывает, что здесь отмечается еще большее усложнение и разнообразие в контроле. Изучение генов, контролирующих синтез 5S-PHK у южноафриканской жабы Xenopus показало, что в овоцитах гены, кодирующие 5S-PHK, экспрессируются лишь в период развития овоци-тов, тогда как ядерные гены соматических клеток, кодирующие синтез 5S-PHK, экспрессируются как в овоцитах, так и в клетках эмбрионов. Установлено также, что синтез 5S-PHK стимулируется белковыми факторами TFA, TFB и TFC. Два последних фактора используются также в транскрипции других генов.
В случае большинства эукариотов считают, что у них нет опе-роноподобной организации генетического контроля действия генов, т. е. эукариотические гены регулируются индивидуально, транс-крибируясь в моноцистронные мРНК, либо эукариотические гены регулируются частично полицистронно, частично моноцистронно. В пользу второго предположения свидетельствуют данные об обнаружении у нематоды С. elegans генов, организованных в опероны. У организма этого вида процессинг полицистронной мРНК в моно-цистронную мРНК происходит до трансляции.
Считают, что генетический контроль синтеза белков у прокари-отов и у эукариотов осуществляется как на уровне транскрипции (частота синтеза мРНК, процессинг мРНК, транспорт мРНК из ядра, стабильность мРНК), так и на уровне трансляции (частота трансляций, регуляции синтеза белковых факторов, ответственных за инициацию, элонгацию и терминацию полипептидной цепи). Считают также, что у эукариотов в генетической регуляции имеет значение структура хроматина, которая блокирует доступ специфических активирующих белков (активаторов) к промоторам. В клетках эукариотов установлен специфический комплекс, который обеспечивает АТФ-зависимое разрушение нуклеосом, позволяя при этом активаторам связываться с нуклеосомным стержнем, что ведет к транскрипции. С другой стороны, установлено, что синтез некоторых ферментов интенсифицируется под влиянием стероид-ных гормонов. Контроль может осуществляться и после трансляции (контроль протеолиза).
1. Что вы понимаете под действием генов и каковы продукты действия генов? Назовите первичные и вторичные продукты действия генов.
2. Дайте определение экспрессивности и пенетрантности генов. Приведите примеры этих свойств генов.
3. Что закодировано в молекулах ДНК?
4. Каковы структура и свойства генетического кода?
б. Что позволяет считать генетический код универсальным?
6. Что вы знаете о разных типах РНК и о тРНК?
7. В состав бактериальной рибосомы диаметром 23 нм входит 35% белка. Сколько белковых молекул м. м. 17 300 может содержаться в ри-босоме, если предположить, что на долю белков приходится 36% объема рибосомы?
8. Что вы знаете о митохондриальном генетическом коде?
9. В чем заключаются различия между ядерным и митохондриаль-ным генетическими кодами?
10. Каково происхождение генетического кода?
11. Один из сегментов молекулы мРНК имеет последовательность УГААГЦГГАУУЦГЦ. В результате мутации шестой слева Ц оказался замененным на У. Как это скажется на полипептиде?
12. Какова разница между индукцией и репрессией ферментов?
13. В чем заключается концепция оперона и каково ее значение в понимании механизмов действия генов?
14. На каких уровнях реализации генетической информации осуществляется генетический контроль экспрессии генов?
15. Чем определяются трудности в изучении генетической регуляции действия генов у эукариот?
16. Сколько типов РНК участвует в биосинтезе белков и что известно о нуклеотидном составе, химических и физических свойствах РНК каждого типа?
17. В научной литературе имеются данные о том, что 7-метил-гуано-зин-б'-монофосфат ингибирует синтез белков в бесклеточной системе, происходящей из ретикулоцитов. Известно также, что на 5'-конце многих молекул мРНК эукариотических клеток имеется 7-метилгуанозин. Известно также, что если удалить эту группу из мРНК (химическим путем) вируса везикулярного стоматита, то это не предотвращает трансляцию в бесклеточной ретикулоцитной системе. Можете ли вы объяснить значение этих данных?
18. Какое влияние оказывает на регуляцию триптофанового оперона увеличение длины РНК-полимеразы?
Дубинин Н. П. Генетика. Кишинев: Штиинца. 1985. 533 стр.
Пехов А. П. Биология и общая генетика. М.: РУДН, 1993. 439 стр.
Schleif R. Genetics and Molecular Biology. The Johns Hopkins Univesity Press. 1993. 698 pp.
Наследственность человека, как и наследственность других живых существ, является менделевской наследственностью, ибо признаки человека генетически детерминированы и их передача от поколения к поколению происходит на основе законов наследственности, открытых и обоснованных Г. Менделем.
Применимость к человеку классического генетического анализа как основного метода изучения наследственности и изменчивости исключена из-за невозможности экспериментальных скрещиваний, длительности времени достижения половой зрелости и малого количества потомства на пару (семью). Поэтому для изучения нормальной и патологической наследственности используют другие методы.
1.
Генеалогический метод (метод родословных).
Часто этот метод называют клинико-генеалогическим. Генеалогия — это учение о родословных. Поэтому смысл данного метода заключается в изучении наследственности человека путем учета и анализа распределения наследственных признаков в семьях, т. е. в изучении наследственности человека по родословным. Метод сводится к изучению родословных связей и передачи признаков среди близких и дальних родственников, прямых и непрямых.
Исследование того или иного признака в семье начинают с того члена семьи, который представляет интерес (исходный пациент, или пробанд). Потомки одних и тех же родителей, происходящие из разных зигот (братья и сестры), получили название сибсов. Родословные составляют путем учета возможно большего количества родственников, используя для обозначения поколений, мужчин, женщин, браков, типов зиготности и т. д., различные символы, перечень которых приводится на рис. 151.
С помощью этого метода возможно установление наследственного характера признака, типа и частоты наследования того или иного признака, сцепленности признака с полом, а также определение зависимости или независимости распределения признаков. Анализируя родословные, можно обнаружить различия между близким сцеплением и аллелизмом. На рис. 152 приводится в качестве примера родословная с доминантным наследованием, а на рис. 153 — родословная, демонстрирующая независимое распределение неаллельных генов. Метод характеризуется относительно большой разрешающей способностью. Однако он имеет недостаток, связанный с трудностями сбора сведений о проявлении того или иного признака у родственников пробанда, поскольку люди плохо знают свои родословные.
С 1892 г. в судебной практике используют метод отпечатков пальцев (дерматоглифику). Иногда к этому методу прибегают в анализе родословных, но он не имеет самостоятельного значения.
2.
Цитологический метод
. Этот метод заключается в цитологическом анализе кариотипа человека в норме и патологии. С его помощью исследуют нарушения хромосом, изменяющие количество и структуру.
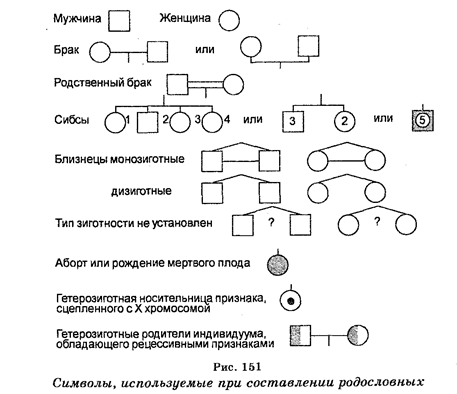
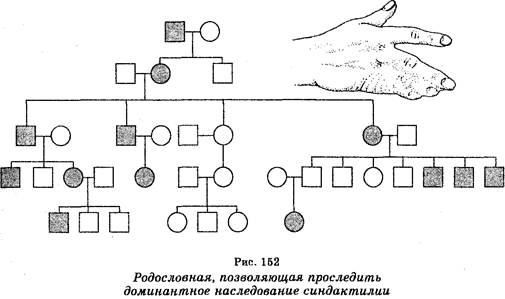
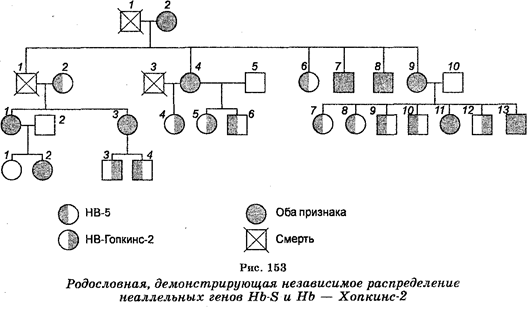
Цитологический метод основывается на данных о количестве, размерах и структуре хромосом. В соответствии с денверовской классификацией (1960) хромосомы обозначают номерами, увеличивающимися по мере уменьшения размеров хромосом. Так первая пара представлена самыми крупными хромосомами, вторая — меньшими, третья — еще меньшими и т. д.
В соответствии с рекомендациями IV Международного конгресса по генетике чечеловека в Париже (1971) при описании добавочных хромосом их числи помещают после общего числа хромосом и половых хромосом со знаком «+» или «-» перед номером вовлеченной ауто-сомы. Например, запись (формула) 47, ХХ+21 означает кариотип женщины с трисомией по 21 паре. Напротив, кариотип мужчины с экотрахромосомой Х обозначают как 47, XXY. Знак «плюс» или «минус» помещают, сопровождая хромосомный символ, чтобы указать удлинение или укорочение хромосомного плеча. Буква q символизирует длинное плечо, ар — короткое. Например, запись 46, XY,1 q+ указывает на увеличение длины длинного плеча хромосомы № 1. Кариотип: 47, XY,+14p+ символизирует мужчину с 47 хромосомами, включая дополнительную хромосому в паре (№ 14) с повышением в длине ее короткого плеча. Сокращениями def (дефишенс), dup (дупликация), г (кольцо, возникающее после воссоединения двух разрывов в хромосоме), inv (инверсия) и t (транслокация) обозначают аберрации хромосом. Номера хромосомы или хромосом помещают после сокращений в скобках. Например, запись 46, XX, г(18) означает кариотип женщины с 46 хромосомами, включая г-хромо-сому № 18. Формула 46, X, inv (Xq) есть кариотип женщины с 46 хромосомами, включая одну нормальную Х хромосому и изохромосому (с двумя генетически идентичными плечами) для длинного плеча хромосомы X. Банды помечают числами в порядке удаления цент-ромеры вдоль короткого плеча (р) и длинного плеча (q) хромосомы.
Главная ценность цитогенетического метода заключается в том, что он позволяет установить связь между нарушениями кариотипа и изменениями фенотипа, т. е. связь между нарушениями в определенной хромосомной паре и определенным наследственным дефектом. Это в свою очередь помогает найти принадлежность гена к определенной группе сцепления. Основное преимущество этого метода заключается в его простоте. Однако этот подход имеет существенные ограничения. Прежде всего с его помощью могут быть исследованы только крупные нарушения в структуре хромосом, видимые с помощью светового микроскопа. Следовательно, это ограничивает количество анализируемых генетических детерминантов. Далее, этот подход может обеспечить изучение генотипов лишь на уровне групп сцепления.
3.
Популяционный метод.
Этот метод основан на законе Харди-Вайнберга и заключается в изучении распространения генов в популяциях человека. В условиях свободного скрещивания частота, с которой возможна встреча двух аллелей в диплоидном организме, равна произведению частот каждого аллеля. Если относительную частоту доминантного аллеля А в двухаллельной системе обозначить р, относительную частоту рецессивного аллеля а обозначить q и если р + q == 1, то при свободном скрещивании частота трех генотипов составляет следующие значения: АА = р2
, Аа = 2pq и аа = q2
. Следовательно, зная о равновесии по Харди-Вайнбергу, можно определить влияние названных выше факторов на относительные частоты этих трех генотипов в поколениях. Как видно, данный метод позволяет изучать не только географическое распространение и частоту тех или иных генов, но и влияние на эти показатели разных факторов.
4.
Близнецовый метод
. Этот метод заключается в изучении генетических закономерностей, присущих однояйцевым (монозигот-ным) и разнояйцевым (дизиготным) близнецам. Обычно сопоставляют монозиготных партнеров с дизиготными, а результаты анализа близнецовой выборки сравнивают с результатами анализа общей популяции. Метод позволяет выяснять наследственную предрасположенность в проявлении ряда признаков и заболеваний, устанавливать коэффициент наследуемости и степень влияния факторов внешней среды на проявление признаков. Успех в использовании этого метода чаще связан с изучением тех признаков, которые не подвержены резкому влиянию со стороны внешних факторов, например, группы крови, пигментации глаз и др. Недостаток метода связан с неполнотой сведений о пренатальном и постнатальном развитии близнецов.
5.
Перенос генов
. Под этим названием различают группу методов, позволяющих переносить гены от одних клеток к другим.
Гибридизация соматических клеток — это метод, основанный на том, что соматические клетки животных способны к гибридизации, при которой образуются гибриды клеток, в ядрах которых содержится набор хромосом обеих сходных клеточных линий, т. е. гибриды являются полиплоидами. В процессе роста гибриды могут терять отдельные хромосомы. Для гибридов, полученных из скрещиваний соматических клеток человека с соматическими клетками млекопитающих, характерно то, что преимущественно теряются человеческие хромосомы. Следовательно, наблюдение одновременной потери той или иной хромосомы и признака указывают на локализацию гена, контролирующего признак в данной хромосоме. В исходных скрещиваниях можно использовать также клетки человека с частично удаленными из них хромосомами. Метод имеет ограничения, определяемые невозможностью экспрессии чужеродных генов в гибридах.
Перенос хромосом — это метод, позволяющий выделить хромосомы и трансформировать ими клетки.
Перенос ДНК — это метод трансформации клеток очищенной ДНК. Метод позволяет переносить одновременно около 50 генов.
6.
Молекулярно-генетические методы
. Эти методы связаны с выделением ДНК, рестрикционным картированием, клонированием сегментов длиной до 50 000 пар оснований и секвенированием отдельных генов.
Кроме того ряд молекулярно-генетических методов направлен на разделение, идентификацию и измерение генных продуктов (белков). Подсчитано, что в клетках человека синтезируется около 30 000 разных белков. Поэтому ставится задача создать каталог белков и построить карту белков человека.
7.
Моделирование наследственных болезней
. Этот метод основан на законе Н. И. Вавилова о сходных рядах наследственности и заключается в моделировании наследственных болезней на животных, у которых встречаются отдельные из этих болезней, например, гемофилии на собаках. Кроме того, используют «сконструированные» линии лабораторных животных, обладающих теми или иными мутантными генами. Например, для изучения болезни Леша-Найяна используют белых мышей, полученных введением в их эмбрионы культивируемых клеток с дефектом по гипоксантин-фосфорибозилтрансферазе.
На основе результатов изучения наследственности человека создают генетические, молекулярные и белковые карты.
У человека ДНК локализована в хромосомах и митохондриях. Количество ДНК в соматических клетках составляет несколько миллиардов нуклеотидных пар.
Кариотип человека показан на рис. 154. Нормальное диплоид-ное число хромосом у человека равно 23, из которых 22 пары хромосом являются аутосомами, а 23-я пара у мужчин представлена половыми хромосомами XY (рис. 154). У женщин — XX (рис. 155). Как уже отмечено, в зависимости от положения центромеры различают телоцентрические, акроцентрические, субметацентрические и метацентрические типы хромосом (см. гл. VI). У человека тело-центрических хромосом нет. Тем не менее на этой основе обычно хромосомы человека распределяют по группам, что зависит от положения центромеры, определяющей длину отрезков (плеч) по обеим сторонам центромеры.
У человека установлено 24 группы сцепления, из которых 22 соответствуют аутосомам, а две — хромосомам Х и Y. Количество генных локусов в геноме составляет примерно 100 000, из которых значительная часть идентифицирована и картирована. Основное количество идентифицированных генов локализовано на аутосомах, меньшее — на Х- и Y-хромосомах. В частности, наиболее «заселенными» генами являются хромосомные пары 1,6,7,11,16и Х-хромо-сома, длина которой составляет около 100 000 000 пар оснований.
 Гены определяют физическое и психическое здоровье человека. Поэтому человек наследует все признаки, которые характерны для него как для живого существа. Любой индивид наследует от своих Гены определяют физическое и психическое здоровье человека. Поэтому человек наследует все признаки, которые характерны для него как для живого существа. Любой индивид наследует от своих
родителей телосложение, рост, вес, форму головы, овал лица, особенности скелета, минеральную плотность костей, размеры отложений жира в подкожной клетчатке, строение и цвет кожи и волос, восприимчивость или устойчивость к тем или иным болезням, походку, манеру держаться, способность к тем или иным наукам или музыке, группу крови, способность клеток к определенной биохимической активности (рис. 156) и т. д. Следовательно, каждый признак, каждая структура, каждая функция человеческого организма детерминирована генами. Так же, как и в случае других живых существ, у человека одни гены начинают действовать еще на ранних этапах онтогенеза, Другие — на поздних.
Для человека характерно исключительное генетическое разнообразие, ибо он является самым гетерозиготным организмом. Простое менделев-ское наследование у человека определяется тем, что тот или иной признак контролируется лишь парой аллельных генов, причем известны все типы наследования (табл. 22).
Наиболее изученным примером менделевской наследственности доминантного типа у человека является наследование способности ощущать вкус фенилтиокарбамида. По многолетним наблюдениям в США, например, 70% лиц из разных исследованных групп способны ощущать вкус этого вещества, тогда как остальные этой способностью не обладают. Среди чешского населения количество лиц, ощущающих вкус фенилтиокарбамида, составляет 67% от общего количества обследованных. Отдельные индивиды способны ощущать вкус этого вещества даже при наличии его в ничтожных количествах (19 х 100-8
молекул/мл).
Исследование родителей и детей во многих семьях показало, что способность ощущать вкус фенилтиокарбамида детерминируется доминантным аллелем Т, тогда как неспособность — рецессивным аллелем t. Следовательно, гетерозиготы (Tt) и доминантные гомозиготы (ТТ) по этой паре генов ощущают вкус вещества, причем в одинаковой степени. Если оба родителя не ощущают вкус вещества, то у них никогда не рождаются дети, «чувствующие» это вещество. «Чувствующие» дети появляются лишь в браках, в которых оба родителя или один из родителей ощущает вкус вещества. В семьях, в которых оба или один из родителей является доминантным гомозиготным организмом (ТТ х ТТ, ТТ х Tt, ТТ х tt), все дети будут ощущать вкус фенилтиокарбамида. Что касается браков между гетерозиготами (TtxTt), а также между гетерозиготами и рецессивными гомозиготами (Tt х tt), то среди детей в этих браках будут встречаться как ощущающие, так и не ощущающие вкус фенилтиокарбамида, причем в первом случае с классическим менделевским отношением 3:1, во втором — 1:1.
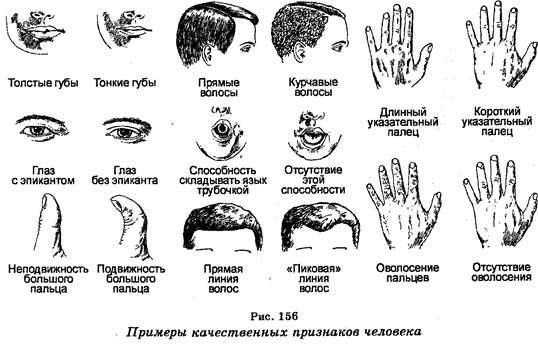
Таблица 22
Наследование некоторых признаков человека
| Признаки
|
Типы наследования
|
| Доминантный
|
Рецессивный
|
| Форма черепа
|
Короткий (брахицефалия)
|
Длинный (долихоцефалия)
|
| Размер глаз
|
Большие
|
Маленькие
|
| Цвет глаз
|
Карие
|
Голубые
|
| Разрез глаз
|
Прямой
|
Косой
|
| Тип глаз
|
Монголоидный
|
Европеоидный
|
| Острота зрения
|
Близорукость
|
Нормальная
|
| Размер носа
|
Крупный
|
Средний или маленький
|
| Форма носа
|
Острый и выступающий вперед
|
Нормальный
|
| —"—
|
Узкий («сухой»)
|
Широкий
|
| —"—
|
Выпуклый с горбинкой («орлиный»)
|
Прямой
|
| —"—
|
Прямой
|
Вогнутый
|
| Размеры хрящевых крыльев носа
|
Покрывает перегородку полностью или почти полностью
|
Оставляет перегородку открытой
|
| Ширина ноздрей
|
Широкие
|
Узкие
|
| Высота переносицы
|
Высокая и узкая
|
Низкая и широкая
|
| Ширина ушей
|
Широкие
|
Узкие
|
| Длина ушей
|
Длинные
|
Короткие
|
Продолжение табл. 22
| Признаки
|
Типы наследования
|
| Доминантный
|
Рецессивный
|
| Характер мочки уха
|
Висящая свободно
|
Срощенная
|
| Наличие острой верхушки уха (дарвиновский бугорок)
|
Присутствует
|
Отсутствует
|
| Длина подбородка
|
Длинный
|
Короткий
|
| Ширина подбородка
|
Широкий
|
Узкий и острый
|
| Форма подбородка
|
Прямой
|
Отступающий назад
|
| Щель между зубами-резцами
|
Имеется
|
Отсутствует
|
| Строение волос
|
С мелкими завитками
|
Вьющиеся, волнистые или прямые
|
| —"—
|
Вьющиеся
|
Волнистые или прямые
|
| —"—
|
Волнистые
|
Прямые
|
| —"—
|
Жесткие прямые («ежик»)
|
Мягкие прямые
|
| Цвет волос
|
Рыжие
|
Светло-русые
|
| —"—
|
Каштановые
|
Светло-русые
|
| Поседение волос
|
До 25 лет
|
После 40 лет
|
| Облысение
|
У мужчин
|
У женщин
|
| Белая прядь волос надо лбом
|
Имеется
|
Отсутствует
|
| Овал лица
|
Круглое
|
Продолговатое
|
| Характер нижней губы
|
Толстая отвисающая
|
Нормальная
|
| Толщина кожи
|
Толстая
|
Тонкая
|
| Цвет кожи
|
Смуглая
|
Белая
|
| Наличие веснушек
|
Имеются
|
Отсутствуют
|
| Рост
|
Нормальный
|
Пропорциональная кар-ликовость
|
| —"—
|
Диспропорциональная карликовость
|
Нормальный
|
| Преобладание руки
|
Праворукость
|
Леворукость
|
| Кисть руки
|
С 6, 7 пальцами (полидактилия)
|
С 5 пальцами
|
| Строение ногтей
|
Тонкие и плоские
|
Нормальные
|
| Цвет ногтей
|
Голубовато-белые
|
Обычные
|
| Узоры на коже пальцев
|
Эллиптические
|
Циркулярные
|
| Тип голоса у мужчин
|
Бас
|
Тенор
|
| Тип голоса у женщин
|
Сопрано
|
Альт
|
| Абсолютный музыкальный слух
|
Имеется
|
Отсутствует
|
Наряду с доминантным и рецессивным типами наследования нормальных признаков у человека встречается кодоминантное наследование, которое заключается в том, что у гетерозиготных индивидуумов формирование признака детерминируют оба аллеля, т. е. проявляются эффекты обоих аллелей. Типичным примером такого наследования является наследование групп крови. У человека четыре группы крови, определяемые на основании антигенных свойств эритроцитов по так называемой системе АВО. Ими являются группы крови А, В, О и АВ или II, III, I и IV группы соответственно. Данные о распространении групп крови среди разного населения приведены в табл. 23.
Дифференциация групп крови основывается на наличии в эритроцитах людей антигенов А и В (табл. 24). Эритроциты людей, относящиеся к первой (нулевой) группе крови, не имеют антигенов А и В, но в сыворотке крови таких людей есть антитела, агглютини-рующие (склеивающие) эритроциты людей, обладающие антигенами А, В или АВ. Эритроциты людей, относящихся ко второй группе крови (группе А), имеют антиген А, а в сыворотке крови таких людей есть антитела, агглютинирующие эритроциты с антигенов В.
Таблица 23
Распространение групп крови системы АВО среди людей
различных национальностей
| Национальность
|
Количество людей с группой крови (в % )
|
| 0(1)
|
А (II)
|
В (III)
|
АВ (IV)
|
| Австралийцы
|
57
|
38,5
|
3
|
1,5
|
| Австрийцы
|
42
|
40
|
10
|
8
|
| Англичане
|
46,4
|
43,4
|
7,2
|
3
|
| Греки
|
38,5
|
41,6
|
16,2
|
4
|
| Индейцы (Сев. Америка)
|
23,5
|
76,5
|
0
|
0
|
| Итальянцы
|
47,2
|
38
|
11
|
3,8
|
| Казахи
|
33,7
|
26,8
|
30,9
|
8,6
|
| Киргизы
|
37,6
|
26,5
|
28,5
|
7,4
|
| Немцы
|
40
|
43
|
12
|
5
|
| Французы
|
43,2
|
42,6
|
11,2
|
3
|
| Японцы
|
27
|
41
|
18
|
14
|
Таблица 24
Реакции между сывороткой и эритроцитами от лиц,
относящихся к разным группам крови
| Группа крови
|
Сыворотка агглготипирует эритроциты групп
|
Эритроциты агглютинируются сывороткой групп
|
| 0
|
(первая)
|
А, В, АВ
|
Никакой
|
| А
|
(вторая)
|
В,АВ
|
0, В
|
| В
|
(третья)
|
А,АВ
|
0, А
|
| АВ
|
(четвертая)
|
Никакой
|
0, А, В
|
Таблица 25
Наследование групп крови системы АВО у человека
| Группа крови родителей
|
Возможные группы крови детей
|
| мать
|
отец
|
| 0
|
А
|
0, А
|
| 0
|
В
|
0, В
|
| 0
|
АВ
|
А, В
|
| А
|
А
|
0, В
|
| А
|
В
|
0, А, В, АВ
|
| А
|
АВ
|
А, В,АВ
|
| В
|
В
|
0, В
|
| В
|
АВ
|
А, В, АВ
|
| АВ
|
АВ
|
А, В, АВ
|
Эритроциты людей, относящихся к третьей группе крови (группе В), имеют антиген В, а в сыворотке крови таких людей есть антитела, агглютинирующие эритроциты с антигеном А. Наконец, эритроциты людей, относящихся к четвертой группе крови (группе АВ), имеют как антиген А, так и антиген В, но в сыворотке крови таких людей нет антител ни против антигена А, ни против антигена В. Индивиды А, В и АВ экспрессируют гликозилтрансферазные активности, конвертируя Н-антиген в А- или В-антигены, тогда как 0(Н)-индивиды такой активностью не обладают: гены для А и В различаются заменами одиночных оснований, изменяющих 4 аминокислотных остатка. Ген О не продуцирует функциональных трансфераз.
Изучения наследования групп крови детьми родителей с известной группой крови (табл. 25) показало, что антигенные свойства эритроцитов контролируются геном, имеющим три аллеля — lА
, IВ
и 1О
, из которых аллель IA
контролирует продукцию антигена А, а аллель IB
— антигена В. У гетерозиготных организмов lA
IB
каждый аллель независим и под контролем каждого из них осуществляется синтез антигенов (кодоминантность), но оба аллеля доминируют над аллелем FS вследствие чего эритроциты людей с первой группой крови (гомозигот по 1O
) не имеют антигенов А и В. Группа крови А (II) бывает лишь у гомозигот IA
IA
или гетерозигот IA
I0
, тогда как группа крови В (III) — у гомозигот IB
IB
или гетерозигот IB
I0
. При кодоминантности (IA
IB
) отмечается группа крови АВ (IV).
Антигены АВО найдены не только в эритроцитах, но и в эпителиальных клетках. Поэтому систему АВО следует называть эпитокровяной системой. Она является изоантигенной системой, состоящей из трех углеводных антигенов А, В и Н.
Помимо групповых антигенов системы АВО, эритроциты человека обладают также антигенами, на основе которых возможна дифференциация других групп крови. Среди них хорошо изучены, например, антигены М и N, детерминируемые парой аллельных генов ММ
и МN
(соответственно), которые доминантны. Вследствие этого индивидуумы, гетерозиготные по обеим аллелям (МM
МN
), обладают эритроцитами, имеющими как антиген М, так и антиген N, и принадлежат к группе MN. Эритроциты с М-антигеном будут у людей, гомозиготных по аллелю МN
, т. е. MN
MN
(группа крови N).
Многие нормальные признаки наследуются сцепленно с полом. В большинстве случаев формирование того или иного признака зависит от действия многих пар генов (полигенных систем) и от взаимодействия их со средой.
Давно замечено, что в популяциях человека индивиды различаются между собой не только физически, но и по поведению. Попытки понять основы таких различий у человека особенно в применении к его интеллектуальным возможностям часто приводили к противоречиям, в результате чего одни свойства рассматривали в качестве наследственных, другие — приобретенных. Поэтому в прошлом причины этих различий были объяснены в положении «природа и воспитание» (nature a. rupture).
Между тем изучение признаков человека показало, что в формировании любого из них имеют значение и наследственность и среда, но что могут быть вариации, заключающиеся в том, что одни признаки зависят больше от наследственности, другие — от среды. Например, группа крови зависит полностью от наследственности, но размеры тела и углеводный метаболизм прямо и существенно определяются как генетическими факторами, так и средой. Способность использовать язык зависит от наследственности (от генетически детерминируемых анатомических особенностей гортани и полости рта), но и, несомненно, от опыта, т. е. от того, что индивид изучает на протяжении жизни в определенной среде.
Особенно большое значение положение «природа и воспитание» имеет в оценке генетических основ поведения человека.
Установлено, что среда, в которой развивается любой животный организм, очень сильно влияет на некоторые особенности поведения его во взрослом состоянии. Например, женские особи макак-резус, отделяемые после рождения от их матерей и лишаемые возможности в раннем возрасте общаться с другими обезьянами, недостаточно активны затем в половом поведении, в социальных играх.
Известно, что особую важность в определении поведения имеет . нервная система. Несомненно также значение гормональной системы, ибо у высших животных, включая человека, гормоны принимают участие не только в репродукции, но и в детерминировании индивидуального темперамента, реакций на стресс. Животные, половое поведение которых является «осознанным», к гормонам чувствительны только в определенное время (сезон), в другое время гормональный стимул не вызывает у них никакого ответа. Напротив, у человека способность к репродукции не зависит от сезона, поэтому гормональное влияние в половом поведении является меньшим. Как нервная, так и гормональная системы в конечном итоге находятся под генетическим контролем (природы). Поэтому поведенческий потенциал человека, вероятно, детерминируется генетически, но этот потенциал может быть реализован только в условиях определенной среды (воспитания).
В контексте познания основ поведения человека часто прибегают к рассмотрению биологических основ преступности. В конце прошлого века итальянский психиатр Чезаре Ломброзо (1835-1909) сформулировал биологическую теорию преступности, суть которой заключается в том, что основу преступности составляют биологические факторы, т. е. преступники являются «прирожденными» преступниками. Эта теория, в которой биологическим фактором преступности позднее стали признавать наследственность, оказалась очень живучей до нашего времени. Точно так же известны утверждения о генетических основах проституции и других пороков людей. Между тем еще никто не идентифицировал ни гены преступности, ни гены проституции. Современные представления основаны на том, что наследуются не гены преступности или проституции, а реакции, т. е. способность индивидов реагировать тем или иным образом в тех или иных условиях жизни.
Вопреки достижениям биологических наук серьезную проблему составляют помехи методов измерения (оценки) интеллекта человека. Еще на заре развития генетики Ф. Гальтон и К. Пирсон в Англии предложили для оценки интеллекта использовать коэффициент корреляции интеллекта у детей и их родителей, а также у детей одной семьи (сибсов). Отстаивая эффективность генеалогического подхода, Ф. Гальтон даже считал, подобно многим людям своего поколения, что женщины по интеллекту стоят ниже мужчин. Однако этот метод не нашел широкого применения.
В 1905 г. французы А. Бине и Т. Симон предложили метод измерения умственных способностей в виде определения так называемого коэффициента интеллектуальности (IQ-тест; от английского intelligence — интеллект, quotinent — коэффициент). В последующем американцы Л. Терман и М. Меррил модифицировали этот метод, определяя интеллектуальные способности детей в возрасте 2-14 лет и оценивая их по таким параметрам, как восприятие, память, внимание, зрение и др. Результаты определялись в баллах, причем средняя одаренность (средний умственный возраст) определялась 100 баллами, дебильность — 70—50 баллами, идиотизм — от 0 до 20 баллов. Талант определялся на основе 125 баллов.
Современная методика определения уровня умственных способностей детей основана на определении разных способностей (уровень общих знаний, сообразительность, словарный запас, географические знания, выполнение арифметических заданий и т. д.) и на суммарной оценке этих способностей.
Определение умственных способностей с помощью IQ-теста вызывает на протяжении многих лет противоречивое отношение к этому тесту со стороны многих ученых, причем эта противоречивость основана на преувеличении либо биологических, либо социальных факторов развития человека, т. е. на признании или непризнании наследуемости коэффициента интеллектуальности.
Между тем многочисленные данные свидетельствуют о том, что коэффициент интеллектуальности характеризуется значительной наследуемостью. В то же время установлено, что вариабельные результаты IQ-теста, часто имеющие место, зависят от влияния нескольких генов и что многие генетические эффекты осуществляются в очень широком диапазоне через среду. Таким образом, вопреки многим недостаткам, IQ-тест может использоваться для определения того направления в поведении людей, которое связано с их способностью к абстрактному мышлению. Однако его использование по-прежнему осложняется нерешенностью многих вопросов. В частности, в литературе нет удовлетворяющего всех точного определения интеллекта, нет согласия в том, что означает, когда говорят один индивид «умнее» другого, нет одинакового понимания важности различий в интеллекте или в амбициях, альтруизме, жестокости и успехах в достижении людьми поставленных целей. По-прежнему остаются невыясненными вопросы о различиях по IQ-тесту людей, принадлежащих к разным социальным группам. Но самое главное состоит в том, что вопреки наследуемости IQ все же не совсем ясно, какова степень влияния на показатели IQ-тестов «природы» и «воспитания» раздельно.
Изучение генетических основ поведения еще отстает от изучения других разделов биологии человека. Главная причина заключается в том, что у человека невозможно прямо связать генетику поведения с менделевскими отношениями, равно как и трудно определить степень наследуемости основных признаков. Кроме того существуют сложности в изучении поведенческих реакции на молекулярном уровне.
В рассмотрении нормальной наследственности человека выдающееся значение имеет осознание его генетической индивидуальности.
Все организмы, возникающие в результате перекрестного оплодотворения гамет, представляют собой полигибридные гетерозигот-ные виды по многим парам генов. Человек не является исключением из этого правила и также имеет полигибридную гетерозиготную природу, что с учетом влияния среды находит отражение в его фенотипических свойствах.
Считают, что человек является самым гетерозиготным видом. В соответствии с законами расщепления и независимого перераспределения генов, открытыми Г. Менделем, одиночная половая клетка человека содержит лишь половину (гаплоидное число) хромосом и генов, представленных в соматических клетках организма в диплоидном наборе. Если, например, какой-либо индивидуум является гетерозиготным организмом по трем парам генов Аа, Вв и Сс, то его половые клетки несут лишь половину таких генов (А, В, С или а, в, с). Следовательно, потомству такого индивида будет передана также половина генов родительского организма, например, генов А, В и С. Между тем разные половые клетки одного и того же родителя несут разные наборы хромосом, а оплодотворение той или иной клетки является делом случая. Поэтому от одних и тех же родителей дети наследуют разные наборы генов как от одного, так и от другого родителя, вследствие чего братья и сестры всегда имеют разные генотипы, а следовательно и разные фенотипы. Одинаковые генотипы и фенотипы, как уже отмечено выше, присущи лишь монозиготным близнецам.
Учитывая соображения, приведенные выше, а также то, что один эйякулят человека содержит около 200-500 млн сперматозоидов, можно допустить, что индивид, гетерозиготный, например, по 30 парам генов, способен дать 280
разных типов гамет. Поскольку в оплодотворении участвует по одной гамете от каждого родителя, то, исходя из способности каждого родителя давать 230
разных типов гамет, можно представить, что в потомстве одной брачной пары количество возможных генотипов зигот, возникающих после объединения разных половых клеток, будет составлять величину порядка 260
.
Как видно, расщепление и независимое перераспределение генов (рекомбинация) является причиной гигантского генетического разнообразия (полиморфизма) людей. В природе нет двух индивидуумов с одинаковым генотипом. Каждый человек несет специфический набор генов, вследствие чего каждый человек генетически индивидуален и неповторим. Исключение составляют лишь однояй-цовые близнецы, для которых характерны одинаковые генотипы.
Генетическая индивидуальность людей определяется на уровне ДНК. Как установлено, в мире нет двух индивидуумов (кроме монозиготных близнецов), у которых были бы сходными минисателлитные повторяющиеся последовательности азотистых оснований в их ДНК. Каждый индивидуум имеет свой уникальный ДНК-профиль, сконструированный в ходе индивидуального развития из определенного количества азотистых оснований, располагающихся в молекулах ДНК в определенной последовательности, т. е. каждый индивидуум является уникальным и неповторимым по его ДНК. Этот подход и его методические основы реализованы в методологии, получившей название ДНК-дактилоскопии, смысл которой заключается в исключительно точной идентификации личности по ДНК (по результатам секвенирования ДНК), что в современной практике нашло широкое применение в определении отцовства, в установлении родственных отношений между людьми, а также в точном установлении принадлежности волос, крови или спермы в практике судебно-медицинских экспертиз. Однако генетическое разнообразие людей может уменьшаться под влиянием разных факторов. Как известно, на протяжении более 3000 лет у евреев традиционна передача обязанностей священников от отца к сыну. Между тем хромосомой, которая определяет мужской пол, является хромосома Y. Новейшие исследования молекулярной организации хромосомы-Y у раввинов и их прихожан в Европе, Северной Америке и Израиле показали, что Y-хромосомы лишь 1,5% раввинов содержали мобильные последовательности азотистых оснований ДНК, тогда как эти последовательности обнаруживались в Y-xpoмосомах 18,4% прихожан. Эти данные свидетельствуют о сужении генетического разнообразия у лиц определенной профессиональной принадлежности. Интересно, что выявленные особенности оказались независимыми от этнического происхождения исследованного еврейского населения.
Еще в большей мере генетическое разнообразие уменьшается в результате родственных браков (браков между двоюродными братьями и сестрами), которые издавна очень широко распространены среди населения многих этнических групп и частота которых довольно высока в отдельных странах. Например, в странах Азии и Африки частота родственных браков в среднем составляет 1 : 20, тогда как в Англии она равна всего лишь 1 : 200. С генетической точки зрения родственные браки в отдаленной перспективе создают благоприятный эффект, т. к. устраняют неблагоприятные генные аллели из генного пула тех или иных популяций людей. Однако ближайший эффект родственных браков представляется неблагоприятным, т. к. они ведут к повышенной возможности появления наследственных болезней уже в следующем поколении. Следовательно, с генетической точки зрения родственные браки таят в себе возможность повышения частоты неблагоприятных генов в популяциях людей и, следовательно, опасны для здоровья.
На очень большом материале, собранном в Японии, установлено, что дети двоюродных братьев и сестер имеют возможность в одном из 16 случаев стать гомозиготными по локусам, унаследованным от их дедов и контролирующим наследственную глухоту и мышечную дистрофию. Установлено также, что смерть детей в браках двоюродных братьев и сестер на 4,4% выше, чем в неродственных браках. Подобным образом генетически индивидуальны организмы разных видов животных и растений (кроме клонов бактерий и соматических клеток животных и растений, а также чистых линий растений и инбредных линий животных).
Интерес к наследственной патологии человека восходит ко временам Гиппократа (460—370 вв. до н. э.), который, вероятно, первым привлек внимание к повторению в семьях таких аномальных признаков, как косоглазие и облысение. Он наблюдал также, что эпилепсия и некоторые болезни глаз, вызывающие слепоту у людей преклонного возраста, встречались лишь в определенных семьях. Интерес к генетике человека в XVIII в. привлек П. Маупертус, собиравшего родословные семей, в которых встречались полидактилия и альбинизм. Анализируя родословные, он предсказывал возможность будущих рождений с этими аномалиями.
Во второй половине прошлого века Ф. Гальтон (1822—1911) применил к изучению наследственности человека статистику. Он привлек также внимание к социальному аспекту наследственной патологии.
В 1909 г. А. Гаррод описал ряд болезней в качестве «врожденных ошибок метаболизма», показав, что в основе наследственных болезней лежит отношение «ген — метаболическая реакция». Больше того, он даже предположил, что некоторые из таких болезней, как, например, алкаптонурия, контролируются рецессивными генами.
Наследственная патология приобрела очень большое значение. Например, смертность детей в раннем возрасте в результате врожденных пороков развития по грубым определениям в 1900 г. составляла 5 на 1000 новорожденных. Такой она является и сейчас. Однако в 1900 г. общая смертность младенцев составляла 150 на 1000 новорожденных, тогда как в 1960 г. она составила 20 на 1000. Оставаясь константной, смертность от врожденных пороков сейчас ответственна за 25% всех смертностей младенцев (в 1900 году она была ответственна за 4%).
Особое значение наследственная патология приобрела в наше время в связи с химическим и радиационным загрязнением среды обитания. Представления о судьбе зачатий дает рис. 157. В соответствии с расчетами от 1/4 до 1/2 всех известных болезней представляют болезни генетической природы. Наследственные болезни стоят на третьем месте после болезней сердечно-сосудистой системы и рака. Считают, что около 3% новорожденных страдают от того или иного генетического дефекта. Одна из каждых гамет человека несет ошибочную информацию, введенную новой мутацией, большая часть которых «оплачивается» ранними абортами. Описано свыше 3000 разных наследственных аномалий, являющихся проявлением мутаций, которые в совокупности представляют собой генетический груз современного человечества.
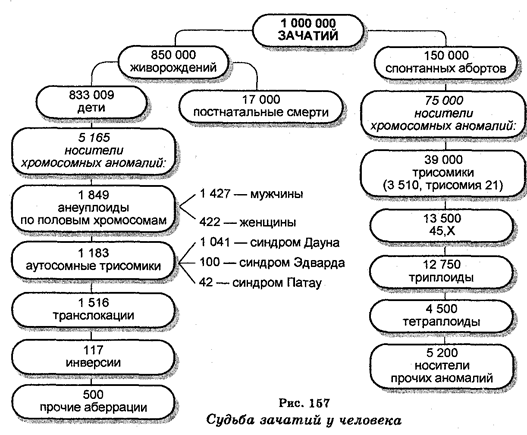
Различают сегрегационный и мутационный груз. Сегрегационный груз — это часть генетического груза, унаследованная людьми современных поколений от людей, принадлежавших к поколениям, жившим на протяжении многих предыдущих веков. Возможно, этот груз пришел к предыдущим и современным поколениям от предков, живших на разных этапах антропогенеза. Можно сказать, что сегрегационный груз представлен «старыми» мутациями. Мутационный груз — это часть генетического груза, которая обусловлена «новыми», т. е. «свежими» мутациями генов и хромосом, возникающими заново в каждом новом поколении.
Итак, наследственная патология обусловлена мутациями хромосомных генов и мутациями хромосом. По этой причине болезни, основу которых составляют мутации генов, условно называют генными болезнями, а те болезни, основу которых составляют мутации хромосом, называют хромосомными болезнями.
Поскольку выяснено, что в митохондриях содержится свой генетический код, изменения которого сопровождаются патологией человека, то выделяют небольшую группу болезней, условно называемых митохондриальными болезнями.
Продолжая эту классификацию наследственных болезней, можно добавить, что еще различают и так называемые мультифактори-альные болезни, в этиологии (причинах) которых имеет место наследственное предрасположение, с одной стороны, и влияние факторов среды, с другой.
Наследственная патология, обусловленная мутациями хромосомных генов
. К настоящему времени известно около 1500 или даже более разных наследственных болезней, являющихся результатом мутаций хромосомных генов. Эти болезни встречаются с разной частотой, но в общем их частота составляет около 1 : 10 000 и даже менее. В пересчете на население, состоящее из нескольких миллионов человек, количество наследственно больных этими болезнями составляет внушительные цифры. Но самое главное заключается в том, что эти болезни характеризуются значительной тяжестью и почти всегда нарушениями не только физического, но и психического здоровья. Подавляющее большинство их не поддается лечению, либо лечение некоторых из них возможно, но оно не является радикальным (см. § 69).
В зависимости от типа наследования, различают аутосомно-доминантные и аутосомно-рецессивные генные болезни, а также болезни, детерминируемые генами, локусы которых находятся на половых хромосомах. Важнейшей особенностью генных болезней является их гетерогенность, заключающаяся в том, что разные мутации могут сопровождаться сходным фенотипическим проявлении болезни. Это явление затрудняет клиническую диагностику многих генных болезней.
Аутосомно-доминантный тип наследования характерен для ряда болезней (табл. 26). Такие болезни встречаются в каждом поколении у гетерозиготных носителей, причем среди сибсов в соотношении 1:1. Далее, для этих болезней характерна как полная, так и не всегда полная пенетрантность генов. Не всегда полной является и экспрессивность генов; причем различия затрагивают не только разные семьи, но и членов одной семьи. Например, нейрофиброматоз в одних семьях имеет генерализованный характер, в других проявляется в виде отдельных кожных поражений. Часть аутосомно-доминантных болезней может проявляться лишь через некоторое время после рождения. Например, хорея Хангингтона проявляется примерно между 30—40 годами жизни больных. Наконец, для аутосом-но-доминантных болезней характерно протекание с повышенной тяжестью у гомозиготных доминантных индивидуумов.
Ахондроплазия является результатом миссенс-мутации, сопровождаемой заменой глицина на аргинин в белке, ответственном за пролиферацию хрящей в суставах длинных костей. Например, карлики фертильны, но гомозиготная ахондроплазия является летальной в неонатальный период, поражая 25% потомства гетерозиготных родителей. Частота ахондроплазии 1 : 15 000.
Установлено, что аутосомно-доминантная болезнь Альцгеймера, характеризующаяся дегенеративным поражением центральной нервной системы, является результатом миссенс-мутации гена на хромосоме 21 и мутацией генов, располагающихся на хромосомах 19 и 14. Следовательно, эта болезнь является полигенной, причем изменения в разных генных локусах выражаются в разной тяжести болезни.
Таблица 26
Аутосомно-доминантные болезни
| Болезни
|
Проявление
|
| Централопатическая эпилепсия
|
Первичные приступы возникают в возрасте между 4-16 годами, после чего реже и обычно исчезают к 40 годам
|
| Глаукома
|
Слепота, вызываемая повышенным давлением жидкости в глазу и дегенерацией нервных клеток (вызывается также аутосомными рецессивными генами)
|
| Хорея Хантингтона
|
Непроизвольные движения лица и конечностей, позже наступают нарушения психики. Симптомы начинают проявляться примерно между 30-40 годами
|
| Мышечная дистрофия
|
Аномалия функций мышц (вызывается также аутосом-ным рецессивным и Х-сцепленным генами)
|
| Полипоз кишечника
|
Формирование множественных полипов, обычно ведущих к раку
|
| Пигментирующий ретинит
|
Воспаление сетчатки глаза с повышенной пигментацией, ведущее к слепоте
|
| Врахидактилия
|
Укорочение концевых костных фаланг (короткопалость)
|
| Ахондроплаэия
|
Карликовость, укорочение конечностей
|
Таблица 27
Аутосомно-рецессивные болезни
| Болезяи
|
Проявление
|
| Серповидно-клеточная анемия
|
Развитие хронической гипоксии и анемии с расстройствами кровообращения и тромбозами
|
| Цистический фиброз
|
Нарушения функций поджелудочной и других желез, образование толстого слоя слизи, ведущей к пневмонии и эмфиземе. Смерть наступает обычно в детском возрасте. Представляет собой наиболее частую генетическую аномалию детей (1 на 3700 рождений)
|
| Галактоземия
|
Пониженные уровни галактозо-1-фосфатуридилтранс-феразы ведут к увеличению печени, катаракте и нарушениям психики
|
| Гидроцефалия
|
Избыточное накопление жидкости в черепной коробке, вызывающее физические и психические нарушения
|
| Глухота врожденная
|
Около половины случаев детской глухоты вызывается этим аллелем
|
| Болезнь Нимена-Пика
|
Накопление липидов в нервных клетках, вызывающее нарушение психики, увеличение печени, замедленный рост. Смерть наступает обычно в первые три года жизни
|
| Пернициозная анемия
|
Нарушения в метаболизме витамина Вдг, что ведет к симптомам, связанным с уменьшением в крови количества эритроцитов
|
| Болезнь Тея-Са-са
|
Прогрессивно развивающиеся паралитические явления, нарушения психики, слепота. Смерть наступает в первые три года жизни. В 27—53% случаев встречается в браках двоюродных сестер и братьев
|
| Фенилкетонурия
|
Нарушения в тонусе мышц, уменьшение пигментации кожи, волос, радужной оболочки глаз, микроцефалия, умственная отсталость
|
Аутосомно-рецессивные болезни (табл. 27), в общем, встречаются чаще, чем аутосомно-доминантные, и проявляются лишь у гомозиготных носителей мутантных аллелей, рождающихся в семьях, где оба родителя являются гетерозиготными или один является гомозиготой, второй — гетерозиготой. Конечно, больные рождаются и в семьях, где оба родителя являются гомозиготными носителями мутантных аллелей (больными).
Для распространения болезней, наследуемых по этому типу, характерна неравномерность. Например, такие болезни, как болезни Тея-Сакса, Нимена-Пика и мышечная деформирующая дистония очень часты среди восточно-европейских евреев (ашкенази), достигая частот, превышающих частоты этих болезней среди населения других национальных групп. Например, болезнь Тея-Сакса, причиной которой является мутация гена, контролирующего лизосом-ную гексозоаминидазу А, среди евреев-ашкенази в Австрии составляет 11%, в Чехии и Словакии — 9%, в Венгрии — 7%, в Румынии — 4%, в Польше, бывшем СССР — 3%. Однако среди людей этой группы очень редка фенилкетонурия. Акаталазия, наследственный дисхроматоз и некоторые другие болезни с высокой частотой встречаются среди японцев. Кровные браки способствуют появлению гомозиготных рецессивных носителей очень редких му-тантных аллелей, и в этом состоит большая генетическая опасность кровных браков.
Еще в начале века было показано, что эритроциты многих индивидуумов агглютинируются сывороткой кроликов, иммунизированных кровью обезьян-резус. Эритроцитарный антиген, который ответственен за эту реакцию, получил название резус-фактора, а ген, который детерминирует это свойство, был назван Rr (или Rh rh). Следовательно, индивидуумы, эритроциты которых содержат Rh-фактор, являются резус-положительными (Rh+
). У резус-положительных индивидуумов (Rh+
) отец также является Rh+
(генотип RR), но мать Rh-
(генотип rr). Когда резус-положительный плод находится в матке резус-негативной женщины (рис. 158), то он вызывает образование в крови матери Rh-антител, которые реагируют с эритроцитами плода. Эта реакция сопровождается развитием анемии плода и его абортом или смертью после рождения.
Такая аутосомно-рецессив ная аномалия, как альбинизм, который обусловлен рецессивным геном, вследствие чего альбинизм является рецессивным по отношению к нормальной пигментации, также встречается в разных странах с разной частотой. В Европе и США один альбинос приходится на 20 000 жителей, тогда как в Южной Америке и Африке — значительно чаще. ная аномалия, как альбинизм, который обусловлен рецессивным геном, вследствие чего альбинизм является рецессивным по отношению к нормальной пигментации, также встречается в разных странах с разной частотой. В Европе и США один альбинос приходится на 20 000 жителей, тогда как в Южной Америке и Африке — значительно чаще.
Анализ родословных многих семей, в которых обнаружены альбиносы, свидетельствует, что дети-альбиносы обычно рождаются от родителей-неальбиносов наряду с детьми-неальбиносами, причем отношение между детьми-неальбиносами и детьми-альбиносами является менделевским (3 : 1). Решающие доказательства того, что альбинизм — это наследственный признак, получены при изучении идентичных близнецов. Как правило, если один из них альбинос, другой также является альбиносом, тогда как из двух неидентичных близнецов лишь один может быть альбиносом. Неальбиносы являются либо гомозиготными доминантными организмами (АА), или гетерозиготными организмами (Аа). Альбиносы являются гомозиготными рецессивными организмами (аа). Брак между альбиносом и неальбиносом не дает детей-альбиносов, т.к. рецессивный аллель а редок и большинство людей не являются носителями такого аллеля. В браке между альбиносом и неальбиносом дети-альбиносы могут быть только тогда, когда неальбинос является гетерозиготным организмом, причем среди детей соотношение между альбиносами и неальбиносами будет составлять 1 : 1. В браках между альбиносами рождающиеся дети всегда альбиносы.
Среди наследственных болезней, передающихся в качестве ауто-сомно-рецессивных признаков и приуроченных к отдельным географическим районам, интерес представляет серповидно-клеточная анемия. Известно, что нормальные эритроциты обладают гемоглобином А, каждая молекула которого состоит из двух поли-пептидных цепей к и двух цепей Р. Каждый й-полипептид представлен специфическими последовательностями, состоящими из 141 аминокислоты, а каждый а-полипептид — специфическими последовательностями, состоящими из 149 аминокислот. Напротив, у больных серповидно-клеточной анемией были найдены эритроциты серповидной формы, которые обладают гемоглобином S, являющимся результатом мутации (замены пары А-Т на пару Т-А) гена НbА, контролирующего синтез гемоглобина А, к аллелю Нbb
S
, контролирующему синтез гемоглобина S. Данная мутация сопровождается тем, что в эритроцитах больных одна половина гемоглобина оказывается гемоглобином А, другая половина — гемоглобином S. Химические различия между гемоглобинами А и S заключаются в том, что b-полипептидные цепи в их глобинах (белковых частях) различаются одной аминокислотной заменой: в b-цепи гемоглобина А шестой аминокислотой является глутаминовая кислота, а в b-цепи гемоглобина S — валин. Генотип индивидов, гомозиготных по S-аллелю, будет Hbb
S
/ Hbb
S
. Удлиненные серповидные эритроциты не способны обеспечивать транспорт кислорода к тканям, поэтому во многих случаях болезнь заканчивается смертельно еще в детском возрасте. В результате мутаций гена HbA
в виде замены в нем пар азотистых оснований происходит образование и других вариантов гемоглобина (помимо гемоглобина S), которых известно более 100 и которые обнаружены в равных популяциях людей. Эти варианты различаются между собой элек-трофоретически, а образование некоторых из них также сопровождается неблагоприятными фенотипическими эффектами разной тяжести.
Свыше 200 генов локализовано на половых хромосомах. Перечень некоторых наследственных болезней, которые контролируются генами, локализованными на Х- или Y-хромосомах, приведен в табл. 28. На хромосоме Х идентифицировано свыше 70 генов, контролирующих гемофилию, мышечную дистрофию, цветовую слепоту (дальтонизм), ювенильную глаукому, оптическую атрофию (дегенерацию зрительного нерва) и др. Большинство из этих болезней наследуются по рецессивному типу. Доминантный тип наследования болезней, которые детерминируются генами, сцепленными о Х-хромосомой, очень редок.
Таблица 28
Наследственные болезни, контролируемые генами,
локализованными на Х- или Y-хромосоме
| Болезни
|
Проявление
|
| Гемофилия
|
Недостаток фактора крови VIII, сопровождающийся несвертыванием крови, в результате чего гемофилия даже при маленьком повреждении может стать смертельной
|
| Гипофосфате-мия
|
Потеря организмом фосфора и недостаток и кальции, что сопровождается размягчением и мальформацией костей
|
| Мукополиса-харидоз П
|
Нарушения в метаболизме белково-углеводных соединений, отставание в росте, умственная отсталость, глухота
|
| Мышечная дистрофия
|
Нарушения структуры и функции мышц, часто начинающиеся в 20-80-летнем возрасте
|
| Ночная слепота
|
Неспособность видеть в темноте
|
| Синдром Рей-фенстейна
|
Форма мужского псевдогермафродитизма, характеризующаяся недостаточным развитием тестисов и повышенным развитием грудных желез
|
| Тестякулярная феминизация
|
Нарушения в развитии мужчин, в результате чего они имеют внешние признаки женщины, включая влагалище, но не имеют матки. Гонады, локализованные в брюшной полости, являются дегенеративными тестисами
|
| Y-сцепленная волосатость ушей
|
Длинные волосы, растущие с внутреннего края ушей
|
| Синдактилия
|
Перепончатообразное сращение второго и третьего пальцев на ноге
|
Наиболее известной из генных Х-сцепленных болезней является гемофилия (несвертываемость крови на воздухе), детерминируемая рецессивным геном и характеризующаяся довольно высокой смертностью, особенно в детском возрасте. Гемофилией болеют почти только мужчины, причем те, которые являются сыновьями женщин-носителей рецессивного гена гемофилии. Если мужчины-гемофилики вступают в брак со здоровыми женщинами, то их сыновья будут здоровы и не будут нести ген гемофилии, т. к. наследуют хромосому Y, свободную от этого гена. Что касается дочерей мужчин-гемофиликов, то все они будут внешне здоровыми, но будут гетерозиготными по гену гемофилии, т. е. носителями этого гена. Более того, сыновья последних в 50% случаев также унаследуют ген гемофилии (равным образом гетерозиготными окажутся и 50% дочерей матери-гемофилика). Поскольку у мальчиков нет второй Х-хромосомы, то унаследованный ими рецессивный мутантный ген гемофилии проявляет свое действие, и они страдают гемофилией. Поскольку у девочек две хромосомы X, то вследствие того, что на второй хромосоме Х локализован доминантный (нормальный) ген, унаследованный рецессивный ген не проявляет своего действия, и девочки не болеют гемофилией. Таким образом, в рассмотренном случае 50% мальчиков будут поражены гемофилией и 50% девочек окажутся гетерозиготными носителями гена гемофилии.
Гемофилией могут страдать и женщины, но лишь тогда, когда они рождаются от родителей, один из которых является гемофиликом (отец), другой — гетерозиготным носителем (мать). Однако это встречается очень редко.
Передача рецессивного гена, детерминирующего гемофилию от гетерозиготных носительниц к их дочерям, внучкам и т. д., которые становятся гетерозиготными носителями и сыновья которых в 50% случаев болеют гемофилией, хорошо прослеживается при ознакомлении с генеалогией некоторых царствовавших семей в Европе, которая восходит к английской королеве Виктории, бывшей гетерозиготной по полулетальному гену гемофилии в Х-хромосоме (рис. 159). От гемофилии умерли три правнука королевы Виктории, испанские инфанты Альфонсо, Гонзалес и Хуан, являвшиеся сыновьями Альфонса XIII и Виктории-Евгении Баттенбергской. Гемофиликом был также сын последнего русского царя Николая II, который унаследовал ген гемофилии от своей матери, бывшей царицы Александры Федоровны (Алисы). Последняя унаследовала этот ген через мать от своей бабки королевы Виктории.
Другим примером наследования генов, сцепленных с Х-хромосомой, является наследование дальтонизма, который, например в США, встречается у 8% мужчин и у 0,5% женщин. Наследование дальтонизма похоже на наследование гемофилии, т. к. рецессивный ген локализован на хромосоме X. Отец передает Х-хромосому ко всем дочерям, но ни к одному из сыновей, а мать передает одну из ее двух хромосом Х ко всем детям. В связи с этим все сыновья матери-дальтоника тоже являются дальтониками, причем независимо от состояния зрения отца. Однако если отец имеет нормальное зрение, то нормальное зрение будут иметь все дочери от этого брака, хотя они и будут являться гетерозиготными носителями. В браке последних с мужчинами, зрение которых нормально, будут рождаться девочки с нормальным зрением, а мальчики — наполовину дальтоники и наполовину с нормальным зрением. Девочка-дальтоник может родиться лишь в браке мужчины-дальтоника с женщиной-дальтоником или гетерозиготной носительницей.
В литературе описан случай, когда мужчина был одновременно гемофиликом и дальтоником, что является доказательством локализации этих генов на одной и той же хромосоме X.
Синдром Леша-Найяна — это заболевание почек мальчиков, сопровождающееся умственной отсталостью и связанное с недостаточностью гипоксантин-фосфорибозилтрансферазы, необходимой для синтеза ДНК. Наследуется через рецессивный ген, сцепленный с Х-хромосомой. Следовательно, мать передает дефектный ген мальчикам через Х-хромосому. Предполагают, что 1/2 мальчиков, родившихся от матери-носителя, будут наследовать дефектный ген.
Мышечная дистрофия ювенильного возраста также зависит от рецессивного гена, сцепленного с Х-хромосомой. Эта болезнь проявляется в мышечном истощении, которое начинается в раннем подростковом возрасте и заканчивается параличами конечностей в среднем и позднем подростковом периоде. Умирают обычно, не достигнув 21 года. Болезнь наследуется по тому же типу, что и синдром Леша-Найяна.
Примерами наследственных аномалий, контролируемых генами, локализованными на Y-хромосоме, являются синдактилия (перепончатообразное сращение 2-го и 3-го пальцев на ноге) и гипертрихиоз (волосатость) края ушной раковины. Поскольку Y-хромосома встречается только у мужчин, эти гены передаются к потомству только по мужской линии.
Характерной особенностью ряда генных болезней является то, что их географическая приуроченность приходится на старые популяции людей, что объясняют так называемым компенсационным преимуществом. Классическим примером такой компенсации является преимущество гетерозиготных носителей гена серповидно-клеточной анемии относительно малярии. Гомозиготы по этому гену погибают, но гетерозиготы более устойчивы к малярии, чем нормальные. При болезни Тея—Сакса у евреев-ашкенази компенсационное преимущество обеспечивается большей устойчивостью их к туберкулезу по сравнению с другим населением в том же самом регионе. Поэтому смертность их от болезни Тея—Сакса является «платой» гомозигот за выживание гетерозигот.
Характерной особенностью генных болезней является также и то, что многие из них, помимо специфических клинических черт, сопровождаются также умственной отсталостью больных. В частности, умственная отсталость установлена в случае около 60 но-зоологических форм аутосомно-доминантных и аутосомно-ре-цессивных аномалий (включая фенилкетонурию). По данным американских авторов в США около 1/3 населения, возможно, имеет один или несколько аутосомных рецессивных генов, которые в гомозиготном состоянии могут сопровождаться умственной отсталостью индивидуумов. Считают также, что 23% всех случаев умственной отсталости контролируется генами, локализованными на половых хромосомах и детерминирующими разные наследственные болезни.
Исследования молекулярных механизмов в этнологии генных болезней позволило установить существование таких болезней, возникновение которых связано с нарушениями регуляции действия генов. Например, синдром Рубенштейма-Тайби, который характеризуется нарушениями в строении концевых фаланг конечностей и умственной отсталостью больных, является результатом мутации гена, детерминирующего синтез белка, действующего в качестве коактиватора ц-АМФ-регулируемой экспрессии этого гена.
Наследственная патология, обусловленная мутациями хромосом. Наследственные болезни, связанные с мутациями хромосом, имеют широкое распространение. Считают, что хромосомные болезни живорожденных составляют 35—40% от всех наследственных болезней.
Для всех или почти всех хромосомных болезней характерной особенностью является то, что они начинают проявляться еще во внутриутробном периоде развития, оказываясь летальными. Установлено, что гибель зародышей происходит уже на стадиях зиготы и бластулы (в первые две недели развития). Хромосомные мутации ответственны примерно за 50% спонтанных абортов и 6% мертво-рождений. Считают, что те или иные хромосомные дефекты имеются у 0,6-0,7% всех новорожденных. Смертность от хромосомных болезней во внутриутробном периоде составляет примерно 1 : 1000.
Различают хромосомные наследственные болезни, детерминируемые мутациями количества хромосом, и хромосомные наследственные болезни, детерминируемые мутациями структуры хромосом.
Таблица 29
Некоторые наследственные болезни, детерминируемые
хромосомными мутациями
| Хромосомная формула
|
Клинический синдром
|
Частота при рождениях
|
Главные фенотипические характеристики
|
| 47, XX +21 или 47, XY +21
|
Болезнь Дауна
|
1 : 700
(у европейцев)
|
Монголоидность, открытый рот с большим языком, умственная отсталость. 1/6 часть больных умирает в первый год после рождения
|
| 47, XX +13
|
Трисомия 13 (синдром Патау)
|
1 : 20 000
|
Глухота, аномалии сердца, полидактилия, умственная дефективность
|
| 47, XX +18 или 47, XY +18
|
Трисомия .18 (синдром Эдварда)
|
1 : 8000
|
Множественные врожденные пороки многх органов. Умственная отсталость. 90% больных умирают в первые 6 месяцев. Известно, когда один ребенок дожил до 5 лет
|
| 45, X
|
Синдром Шерешевского-Тернера
|
1 : 3000
|
Женщины с недостаточным половым развтием, короткой фигурой, нарушениями сердечно-сосудистой системы
|
| 47, XXY
|
Синдром Клайнфелтера
|
1 :500
|
Мужчины с недоразвитыми тести-сами, но развитой грудью, с женским голосом, длинными конечностями
|
| 47,XXX
|
Синдром Трипло Х
|
1 : 700
|
Больные женщины внешне не отличаются от нормальных, но их фолликулы недостаточно развиты. Менструации нерегулярные
|
| 48,ХХХХ 48,XXXY 48,XXYY 49,XXXXY
50, XXXXXY
|
Различные полисомии
|
Аномалии скелета, умственная отсталость и другие симптомы
|
Мутационные нарушения плоидности хромосом человека в сторону гаплоидии неизвестны. Что касается полиплоидии, то она описана в виде триплоидии и тетраплоидии при исследовании спонтанно абортированных эмбрионов и плодов, а также мертворожденных. По некоторым данным 1% всех зачатий человека представлен триплоидными зиготами. Аборты триплоидов, как правило, происходят к концу третьего месяца беременности.
Наиболее часты болезни, являющиеся результатом анеуплои-дии (гетероплоидии) аутосом и половых хромосом. Клинически значимые аномалии составляют половину всех хромосомных аномалий новорожденных. В табл. 29 представлены данные о некоторых из таких наследственных аномалий.
Наиболее частой формой анеуплоидии являются полисомии по тем или иным хромосомам, частота которых составляет 5,7´10-4
на гамету/генерацию. В частности, аутосомные полисомии в виде трисомии, характеризуются тем, что одна из пар в хромосомном наборе имеет добавочную хромосому, т. е. представлена тремя го-мологичными хромосомами (2n + I) или, как говорят, представлена не дупликатом, а трипликатом. Например, болезнь Дауна, характеризующаяся серьезными нарушениями здоровья, включая нарушения психической деятельности и иммунологическую недостаточность, обусловлена трисомией по 21-й паре хромосом. Минимальный размер трисомного участка при этой трисомии составляет 300 000-400 000 пар оснований. У шимпанзе синдром Дауна является результатом трисомии по 22-й паре, которая гомологич-на 21-й хромосомной паре человека.
Аутосомные трисомии бывают и в случае других хромосом. Например, трисомия по 13-й паре обуславливает синдром Патау, по 18-й паре — синдром Эдварда. Известны также трисомии по 22-й, 8-й и др. парам хромосом. Описаны также случаи даже аутосом-ных тетрасомий и пентасомий, но они детальны и обнаруживаются только при исследовании абортусов. Что касается аутосомных мо-носомий, то они тоже детальны и тоже обнаруживаются в клетках абортусов.
Более частой формой гетероплоидии является анеуплоидия по половым хромосомам, частота которой у людей в разных странах в среднем составляет 9,3´10-4
на гамету/генерацию. Различают полисомии по половым хромосомам и Х-моносомии. Наиболее известными примерами наследственных заболеваний человека, в основе которых лежит гетероплоидия по половым хромосомам, являются синдромы Клайнфелтера и Трилло X, которыми страдают женщины, а также синдром 47, XYY, который характеризуется агрессивностью, умственной отсталостью и антиобщественным поведением больных мужчин.
Причинами трисомии считают нерасхождение хромосом при га-метогенезе (мейозе) у одного из родителей. В частности, трисомии в 90% случаев возникает в результате нерасхождения хромосом при овогенезе.
Такая гетероплоидия, как ХХХ-26/28, также является результатом нерасхождения хромосом при овогенезе и влияния возраста матери, тогда как анеуплоидия XXY-56/112 является результатом нерасхождения хромосом при сперматогенезе, причем возраст обоих родителей не имеет значения.
Примером наследственного заболевания, в основе которого лежит Х-моносомия, является синдром Шерешевского-Тернера. Х-мо-носомия служит, вероятно, главной причиной полового недоразвития женщин.
Частота многих хромосомных аномалий зависит от возраста матерей, что хорошо показано в случае синдрома Дауна (табл. 30).
Как правило, мутации количества хромосом происходят в гаметах одного из родителей. Поэтому, все клетки организма, в зачатии которого принимала участие одна из мутантных гамет, будут содержать аномальный хромосомный набор. Однако иногда количественные хромосомные мутации могут случаться в процессе первых делений зиготы, образованной нормальными гаметами. Из такой зиготы разовьется организм, часть клеток которого будет иметь нормальный диплоидный набор, другая же часть — аномальный. Это явление называют хромосомным мозаицизмом, а индивидов, обладающих мозаицизмом — хромосомными мозаиками. Мозаицизм более част по половым хромосомам. Такие мозаики имеют генотип Х/ХХ, X/XY, XX/XY, XXY/XX.
Таблица 30
Возраст матерей и частота синдрома Дауна
| Возраст матерей
|
Частота синдрома
|
| до 20 лет
|
1 : 2000
|
| до 30 лет
|
1 : 1000
|
| до 30-34 лет
|
1,6 : 1000
|
| до 35-39 лет
|
4 :1000
|
| до 40—44 лет
|
14 : 1000
|
| до 45 и выше
|
25 : 1000
|
Для хромосомных болезней, причиной которых являются мутации количества хромосом, также как и для генных, характерна в качестве «сопутствующего» симптома умственная отсталость. Она часта в случае аутосомных синдромов. Что касается половых хромосом, то около 1% умственно отсталых женщин имеют одну или более лишнюю хромосому X.
Хромосомные болезни, детерминируемые мутациями структуры хромосом, более редки, а их клиническое проявление менее выражено или неспецифично по сравнению с аномалиями, вызываемыми мутациями количества хромосом. Мутации структуры хромосом выявлены во всех парах аутосом. Часто многие из этих мутаций называют частичными трисомиями и моносомиями. Например, деления по хромосомной паре 5 вызывает синдром «кошачий крик», связанный с анатомическими изменениями лицевого скелета. Синдром «дупликация-делеция» в хромосоме 3° заключается в спонтанных абортах. Описаны, однако, два рождения, при которых ребенок не способен сидеть, не может есть твердую пищу, имеет очень короткий нос. Предполагают, что при синдроме Дауна в некоторых случаях происходит транслокация сегмента хромосомы 21 на другую хромосому. По данным японских авторов 14 из 321 больного болезнью Дауна характеризовались транслокацией части хромосомы на одну из нескольких других хромосом.
Таблица 31
Частота встречаемости отдельных болезней среди родственников
| Болезнь
|
Частота (%)
|
Конкордантность у близнецов (%)
|
| среди родственников
|
в популяции
|
монозиготные
|
дизиготвые
|
| Ишемическая болезнь сердца
|
30-60
|
19
|
67
|
43
|
| Язвенная болезнь
|
8
|
0,6
|
50
|
14
|
| Шизофрения
|
14
|
1
|
67
|
18
|
| Ревматизм
|
10
|
2
|
37
|
7
|
Наследственная патология, обусловленная митохондриальными генами. Известно небольшое количество наследственных болезней, получивших название митохондриальных из-за того, что в их основе лежат изменения в ДНК митохондрий, т. е. митохондриальных генов.
Одной, наиболее известной из таких болезней, является наследственная оптическая нейропатия Лебера, связанная с поражением мышц глаза и проявляющаяся в виде внезапной потери зрения в период между юношеским и взрослым возрастом из-за инактива-ции зрительного нерва.
Описаны также хроническая прогрессирующая офтальмоплегия, синдромы Кернса-Сэйра и Пирсона, митохондриальная энцефаломиопатия и молочный ацидоз.
Известны данные, в соответствии с которыми перестройки в митохондриальной ДНК фенотипически проявляются в развитии диабета Diabetus melitus.
Болезни с наследственным предрасположением. Болезни, в патогенезе которых играет роль наследственность и проявление которых зависит от действия факторов внешней среды, называют болезнями с наследственным предрасположением. Такими болезнями являются атеросклероз, гипертоническая болезнь, ишемическая болезни сердца, ревматизм, язвенная болезнь, дерматиты, некоторые формы диабета, шизофрения и другие. Доводами в пользу наследственной предрасположенности служит существенно большое накопление повторных случаев болезни среди родственников больных по сравнению с популяционной частотой данного заболевания, а также значительное повышение показателей конкордантно-сти в парах монозиготных близнецов по сравнению с. такими показателями в парах дизиготных близнецов (табл. 31).
Генетическая природа болезней с наследственным предрасположением неодинакова. Различают моногенные и полигенные болезни с наследственным предрасположением.
Моногенные болезни с наследственным предрасположением — это болезни, при которых предрасположенность определяется одним геном во взаимодействии с точно известным фактором среды. Полигенные болезни с наследственным предрасположением — это те болезни, при которых предрасположенность определяется многими генами и также во взаимодействии со многими факторами среды. Такие болезни часто называют мультифакториальными болезнями.
Изучение мультифакториальных болезней затруднено по многим причинам, одна из которых заключается в поисках генетических маркеров предрасположения. Например, в случае атеросклероза повышенный уровень холестерина контролируется несколькими генами, один из которых'ответственен за повышение уровня холестерина (семейная гиперхолистеринемия), второй — за повышение в сыворотке концентрации триглицеридов (семейная глицеридемия), третий — за повышение уровня обоих липидов (комбинированная гиперлипидемия). Поскольку холестерин и триглицериды имеют значение в развитии раннего атеросклероза коронарных сосудов, то считают, что эти гены в сочетании с фактором среды создают механизм предрасположения к атеросклерозу и развитию инфаркта миокарда.
Болезни с наследственной предрасположенностью характеризуются разной степенью выраженности, что является отражением разных уровней накопления факторов предрасположения и комбинации их с разными по степени и направлению воздействия факторами среды (стресс, климатические условия, инфекции и др.).
Различают пренатальную и постнатальную диагностику. В пре-натальной диагностике используют ряд методов, среди которых наиболее эффективным является так называемый амниоцентоз (рис. 160). Суть этого метода заключается в получении от женщин околоплодной жидкости и исследовании ее непосредственно путем микроскопии содержащихся в ней клеток или после культивирования последних.
С помощью метода амниоцентоза возможно определение пола и кариотипа плода. Пренатальную диагностику обычно используют в семьях, где риск рождения наследственно больного ребенка составляет около 3-5%.
Постнатальная диагностика наследственных болезней основывается на результатах генетического и клинического обследования пациентов. Генетическое обследование пациентов основывается в первую очередь на результатах генеалогического анализа. В зависимости от показаний привлекают также цитогенетические, биохимические, иммунологические и другие методы.
Большое значение имеют методы массовой («просеивающей») диагностики с целью проверки населения на возможность скрытых форм наследственных аномалий. «Просеивающую» диагностику применяют к новорожденным с целью выявления галактозе-мии, муковисцидоза и других наследственных болезней и к определенным группам населения с целью выявления гетерозигот-ного носительства болезни Тея-Сакса, серповидноклеточной анемии и талассемии.
Во многих случаях диагностика наследственных болезней является успешной. Однако в случае гетерогенных наследственных аномалий (разных аномалий, но сходных по проявлению) она очень затруднена. Особенно сложной является диагностика болезней, характеризующихся умственной отсталостью, т. к. она осложнена разнообразием факторов среды.
Наследственные болезни пока не поддаются радикальному лечению, ибо пока не разработаны методы исправления генотипа. Но многие наследственные болезни можно лечить, используя симптоматическое лечение или влияя на развитие болезни. В частности, при некоторых наследственных болезнях прибегают к диетотерапии.
Например, патогенез такого наследственного заболевания, как галактоземия, связан с накоплением в клетках галактозы из-за отсутствия фермента р-0-галакто-1-фосфатуридилтрансферазы, вследствие чего развиваются изменения в печени и головном мозге, и, наконец, ослабление умственной деятельности, то лечение болезни обычно проводят исключением материнского молока и назначением диеты, не содержащей галактозы.
Другим примером диетотерапии является лечение фенилкето-нурии, которая может быть обнаружена простым исследованием мочи новорожденных. Новорожденному с этой болезнью на весь период детства назначают диету с пониженным содержанием фе-нилаланина, что предупреждает умственную отсталость и другие симптомы фенилкетонурии. Когда «вылеченные» становятся матерями, они вновь должны вернуться к диете с пониженным содержанием фенилаланина и таким образом обеспечить подходящую среду для плода. Их потомство (девочки) тоже может нуждаться в низкой по фенилаланину диете в раннем детстве, чтобы предупредить симптомы болезни.
С целью лечения часто прибегают к введению в организм недостающего фактора. Например, при первициозной анемии, гемофилии и антигемофильной глобулиновой недостаточности прибегают к периодическим инъекциям специфического (недостающего в организме) белка, что временно улучшает состояние больных.
Для лечения ряда болезней используют переливание крови или удаление из организма токсических веществ с помощью лекарственных средств. Лекарственную терапию часто используют также с целью индукции недостающих ферментов. Например, отдельные наследственные болезни лечат фенобарбиталами, которые стимулируют индукцию недостающих ферментов. При гомо-литической болезни новорожденных (Ш^-дети) матерям вводят глобулин Rh, который разрушает Rh-позитивные клетки плодового происхождения и предупреждает образование Rh-антител, которые могут повредить другие плоды в последующих беременностях.
Важное место в лечении наследственных болезней занимает хирургическое лечение. Например, часто прибегают к удалению толстой кишки при полипозе, селезенки — при сфероцитозе, почек — при цистинозе. Многие аномалии (незаращение верхней губы, врожденные пороки сердца, полисиндактилия, болезнь Нимена-Пика и др.) исправляют хирургической коррекцией или с помощью трансплантаций. Однако важно подчеркнуть, что при лечении генетический дефект у больных сохраняется. Вступая в детородный возраст, наследственные больные передают неблагоприятные гены своему потомству.
В широком плане в рамках разных методов лечения наследственных болезней можно рассматривать и евфенические мероприятия, под которыми понимают компенсацию естественных недостатков человека, в фенотипе, т.е. улучшение здоровья человека через фенотип. Часто евфенические мероприятия называют лечением адаптивной средой. Они известны давно и к ним относят пре-натальную и постнатальную заботу о потомстве, иммунизации, переливания крови, трансплантации органов, пластическую хирургию, обагащение диет белками, витаминами и микроэлементами, исключение из диет отдельных углеводов, физическую культуру, лекарственную терапию при тех наследственных болезнях, проявление которых зависит от факторов среды. Однако евфенические мероприятия, как и симптоматическое и патогенетическое лечение, тоже не приводят к радикальным результатам, ибо благодаря евфенике нельзя кардинально преодолеть наследственные дефекты. Кроме того эффекты, достигнутые в результате евфенических мероприятий, не передаются по наследству и не сопровождаются уменьшением количества неблагоприятно действующих генов в популяциях человека.
Этиологическое лечение наследственных болезней, которое должно приводить к кардинальному исправлению наследственных аномалий, еще не разработано. Однако в связи с достижениями физико-химической биологии сформулированы и разрабатываются программы изменения в желаемом направлении фрагментов генетического материала, детерминирующих наследственные аномалии человека.
Профилактика наследственных болезней сводится к профилактике болезней, унаследованных от предыдущих поколений и к профилактике болезней, вновь возникающих в результате мутаций в зародышевых клетках родителей.
Профилактика болезней, унаследованных от предыдущих поколений (сегрегационный груз), осуществляется, главным образом, с помощью медико-генетической консультации. Основные задачи медико-генетического консультирования сводятся к тому, чтобы преодолеть степень риска рождения ребенка с наследственной патологией в данной семье и помочь родителям принять правильное решение, а также насторожить на риск рождения наследственно отягощенного ребенка и облегчить возможность постановки раннего диагноза и принятия лечебных мер. В глобальном плане медико-генетическая консультация должна понижать частоту неблагоприятных генов в популяциях человека.
Результативность медико-генетической консультации зависит от точности знания родословной и точности определения скрытого носительства патологических генов, диагностики гетерозиготного состояния. Медико-генетическая консультация связана с исключительной морально-этической ответственностью консультирующего генетика или врача в познании тайн при анализе родословных и в определении степени риска. От медико-генетической консультации, как врачебного заключения, следует отличать консультацию как
учреждение. В нашей стране первая медико-генетическая консультация была организована еще в конце 20-х гг. (С. Н. Давиденков). В настоящее время медико-генетические консультации существуют во многих странах мира.
Профилактика наследственных болезней, вновь возникающих в результате мутации (мутационный груз), сводится к предупреждению загрязнения среды обитания человека факторами, которые могут действовать как мутагены. В условиях научно-технической революции серьезное значение приобретает слежение за генетическими процессами в популяциях человека (см. гл. XVIII).
1. Применимы ли законы Г. Менделя к человеку? Если да, то покажите, на чем основана их применимость.
2. Чем отличаются методы изучения наследственности человека от основного метода генетики — классического генетического анализа?
3. Каким образом можно установить принадлежность группы сцепления к той или иной паре хромосом человека?
4. Какова ценность метода клонирования генов в генетике человека?
5. Назовите нормальные признаки человека, детерминируемые ауто-сомными, доминантными и рецессивными генами.
6. Приведите примеры наследственности человека, сцепленной с полом.
7. Что вы знаете о генетической индивидуальности людей?
8. Что такое генетический груз и чем он определяется?
9. Как классифицируют наследственные болезни?
10. Назовите генные наследственные болезни, наследуемые как ауто-сомные доминантные и рецессивные признаки, а также признаки, сцепленные с полом.
11. Что вы знаете о хромосомных болезнях, их частоте, особенностях распространения и этиологии?
12. Что вы знаете о болезнях с наследственным предрасположением и об их отличиях от наследственных болезней?
13. Аномалия «заячья губа» встречается с частотой 0,1% . Конкордант-ность по этому признаку между идентичными близнецами составляет 50%, между сибсами — 3,5%, между двоюродными братьями и сестрами — 0,7%, а между троюродными братьями и сестрами — 0,3%. На степень конкордантности не влияет пол. Можете ли вы на основе этих данных определить тип наследования «заячьей губы»?
14. В эритроцитах человека найдено три электрофоретически различных формы кислой фосфатазы А, В и С. При исследовании 178 англичан оказалось, что их эритроциты содержат разные фосфатазы, а именно:
фосфатаза А — 17 человек, фосфатазы В + С — 9,
фосфатаза В — 61 человек, фосфатазы А + С — 5,
фосфатаза С—О, фосфатазы А, В и С — О,
фосфатазы А + В — 86 человек.
Объясните эти результаты. Почему ни у кого не найдена фосфатаза С?
16. Какими могут быть заключения в следующих трех случаях семейной ситуации?
а) Дядя женщины (брат ее матери) болел гемофилией. Женщина забеременела и желает знать, каков риск рождения у нее ребенка-гемофилика. Как могла бы измениться ситуация, если бы дядей-гемофиликом был брат ее отца?
б) Женщина имела двоюродного брата, который умер от фиброцис-тита. Имеется ли риск рождения у этой женщины больного ребенка?
в) Старшая сестра мужчины страдала хореей Хантингтона. Мужчине 40 лет и он хочет знать о риске для себя и детей, которых он желает иметь. Что вы скажете в ответ на его вопросы?
16. Какова, по вашему мнению, результативность лечения наследственных болезней? Какое социальное значение имеет невозможность лечения некоторых наследственных болезней?
17. В чем состоит профилактика наследственных болезней? Каковы возможности и ограничения генетической консультации? Каким образом можно привлечь внимание к генетической консультации индивидов детородного возраста?
18. Что вы знаете о научных направлениях в разработке способов радикального лечения наследственных болезней?
19. Что такое генетический мониторинг и каково его значение в профилактике наследственных болезней, в охране среды обитания человека?
Бочков Н. П., Захаров А. Ф., Иванов В. И. Медицинская генетика. М.: Медицина.
1984. 366 стр. Дубинин Н. П., Карпец И. И., Кудрявцев В. Н. Генетика, поведение, ответственность.
М.: Изд. политической литературы. 1982. 304 стр. Цехов А. П. Биология и общая генетика. М.: Изд. РУДН. 1993. 439 стр.
«...твердо помнить должно,
что видимые телесные на Земле вещи
и весь мир не в таком состоянии
были с начала от создания,
как ныне находим,
но великие происходили
в нем перемены...»
М. В. ЛОМОНОСОВ
(1763)
Масса Земли составляет около 4´1018
тонн, а возраст — около 4,5-5 млрд лет. Считают, что жизнь возникла на Земле примерно 3,5-3,8 млрд лет назад.
Она оказала существенное влияние на атмосферу, которая изменялась от окисляющей к неокисляющей.
Огромное разнообразие живых форм, населяющих сейчас Землю, является результатом длительного процесса эволюции, под которой понимают развитие организмов во времени или процесс исторического преобразования на Земле, результатом которого является многообразие современного живого мира. Термин «эволюция» (от лат. evolutio — развертываю) был введен в науку в 1762 г. швейцарским натуралистом Ш. Бонна (1720-1793).
Вначале эволюция шла очень медленно. Первыми и единственными живыми обитателями Земли в течение 3 млрд лет были микроорганизмы. Многоклеточные появились после четырех пятых времени начала существования Земли. Эволюция человека заняла несколько последних миллионов лет. Центральным моментом эволюции является филогенез (от греч. phyle — племя, genesis — развитие), — процесс возникновения и развития вида, т. е. эволюцию вида.
Представления о развитии жизни отражены в теории эволюции, которая основывается на данных об общих закономерностях и движущих силах развития живой природы. Она представляет собой синтез достижений дарвинизма, биологии, генетики, морфологии, физиологии, экологии, биогеоценологии и других наук. В наше время теория эволюции, основу которой составляет дарвинизм, — это наука об общих законах развития органической природы, методологическая основа всех специальных биологических дисциплин.
В этом разделе мы рассмотрим теорию эволюции. Будут приведены также данные о происхождении жизни, о микроэволюции и видообразовании, а также о ходе, главных направлениях и доказательствах эволюции. В самостоятельных главах мы излагаем сведения об эволюции систем органов животных и о происхождении человека.
Эволюция протекает на всех уровнях организации живой материи и на каждом уровне характеризуется новообразованием структур и появлением новых функций. Объединение структур и функций одного уровня сопровождается переходом живых систем на более высокий эволюционный уровень.
Проблемы происхождения и эволюции жизни на Земле принадлежали и принадлежат к числу величайших проблем естествознания. Эти проблемы привлекали к себе внимание человеческого ума с самых незапамятных времен. Они являлись предметом интереса всех философских и религиозных систем. Однако в разные эпохи и на разных ступенях развития человеческой культуры проблемы происхождения и эволюции жизни решались по-разному.
В основе современной теории эволюции лежит теория Ч. Дарвина. Но эволюционизм существовал и до Ч. Дарвина. Поэтому, чтобы лучше понимать современную теорию эволюции, важно знать о взглядах на мир до Ч. Дарвина, о том, как развивались идеи эволюционизма.
Самыми древними взглядами на природу были мистические, по которым жизнь связывали с силами природы. Но уже у самых истоков культуры в древней Греции на смену мистическим истолкованиям природы приходят начала других представлений. В тот период возникла и стала развиваться доктрина абиогенеза и спонтанного самозарождения, в соответствии с которой признавалось, что живые организмы возникают спонтанно из неживого материала. Тогда же появились и эволюционные идеи. Например, Эмпедокл (490—430 гг. до н. э.) считал, что первые живые существа возникли из четырех элементов мировой материи (огонь, воздух, вода и земля) и что для природы характерно закономерное развитие, выживание тех организмов, которые наиболее гармонично (целесообразно) устроены. Эти мысли были очень важными для дальнейшего распространения идеи о естественном происхождении живых существ.
Демокрит (460-370 гг. до н. э.) считал, что мир состоит из множества мельчайших частиц, которые находятся в движении, и что жизнь является не результатом творения, а результатом действия механических сил самой природы, приводящих к самозарождению. По Демокриту самозарождение живых существ происходит из ила и воды в результате сочетания атомов при их механическом движении, когда мельчайшие частицы влажной земли встречаются и соединяются с атомами огня. Самозарождение представлялось случайным процессом.
Предполагая, что черви, клещи и другие организмы возникают из росы, ила, навоза, волос, пота, мяса, моллюски из влажной земли, а рыбы из морской тины и т. д., Платон (427-347 гг. до н. э.) утверждал, что живые существа образуются в результате соединения пассивной материи с активным началом (формой), представляющим собой душу, которая затем движет организмом.
Аристотель (384-322 гг. до н. э.) утверждал, что растения и животные возникают из неживого материала. В частности, он утверждал, что некоторые животные возникают из разложившегося мяса. Признавая реальность материального мира и постоянство его движения, сравнивая организмы между собой, Аристотель пришел к заключению о «лестнице природы», отражающей последовательность организмов, начинающуюся с неорганических тел и продолжающуюся через растения к губкам и асцидиям, а затем к свободно живущим морским организмам. Однако, признавая развитие, Аристотель не допускал мысли о развитии низших организмов к высшим.
Взгляды Аристотеля оказали влияние на века, ибо последующие греческие и римские философские школы полностью разделили идею самопроизвольного зарождения, которая все больше и больше наполнялась мистическим содержанием. Описания различных случаев самозарождения даны Цицероном, Овидием, позднее Сенекой, Пли-нием, Плутархом и Апулеем. Идея изменяемости прослеживается во взглядах древних философов Индии, Китая, Месопотамии, Египта. Раннее христианство обосновывало доктрину абиогенеза примерами из Библии. Подчеркивалось, что самозарождение действует от сотворения мира до наших дней.
В течение средних веков (V-XV вв.) вера в самопроизвольное самозарождение была господствующей среди ученых того времени, ибо философская мысль тогда могла существовать лишь в качестве богословской мысли. Поэтому сочинения средневековых ученых содержат многочисленные описания самозарождения насекомых, червей, рыб. Тогда часто считали, что даже львы возникли из камней пустыни. Знаменитый врач средневековья Парацельс (1498—1541) приводил рецепт «изготовления» гомункулуса (человека) путем помещения спермы человека в тыкву. Как известно, Мефистофель из трагедии Гёте «Фауст» называл себя повелителем крыс, мышей, мух, лягушек, клопов и вшей, чем И. Гёте подчеркивал чрезвычайные возможности самозарождения.
Средневековье не внесло новых идей в представления о развитии органического мира. Напротив, в тот период царило креационисти-ческое представление о возникновении живого в результате акта творения, о постоянстве и неизменности существующих живых форм. Вершиной креационизма было создание лестницы тел природы: бог — ангел — человек — животные, растения, мицеллы.
Гарвей (1578—1667) допускал, что черви, насекомые и другие животные могли зарождаться в результате гниения, но при действии особых сил. Ф. Бэкон (1561—1626) считал, что мухи, муравьи и лягушки могут самопроизвольно возникать при гниении, однако к вопросу подходил материалистически, отрицая непреодолимую грань между неорганическим и органическим. Р. Декарт (1596—1650) также признавал самопроизвольное зарождение, но отрицал участие в нем духовного начала. По Р. Декарту самозарождение — это естественный процесс, наступающий при определенных (непонятных) условиях.
Оценивая взгляды выдающихся деятелей прошлого, можно сказать, что доктрина самозарождения не подвергалась сомнению вплоть до середины XVII в. Метафизичность воззрений в XVII—XVIII вв. особенно проявлялась в представлениях о неизменности видов и органической целесообразности, которые считались результатом мудрости творца и жизненной силы.
Однако вопреки господству метафизических представлений в XVI-XVII вв. все же происходит ломка догматического мышления средневековья, обостряется борьба против духовной диктатуры церкви, возникает и углубляется процесс познания, который привел в XVIII в. к существенной аргументации против теории абиогенеза и к возбуждению интереса к эволюционизму.
Осуществив в 1665 г. ряд экспериментов с мясом и мухами, Ф. Реди (1626—1697) пришел к заключению, что личинки, возникающие в гниющем мясе, являются личинками насекомых, и что такие личинки никогда не возникнут, если мясо поместить в закрытый контейнер, недоступный для насекомых, т. е. для откладывания ими яиц. Этими экспериментами Ф. Реди опроверг доктрину самозарождения высших организмов из неживого материала. Однако в материалах и рассуждениях Ф. Реди не исключалась мысль о спонтанном самозарождении микроорганизмов и гельминтов в кишечнике человека и животных. Следовательно, сама идея самозарождения еще продолжала существовать.
В 1765 г. Л. Спаланцани (1729-1799) во многих опытах показал, что развитие микробов в растительных и мясных настоях исключается кипячением последних. Он выявил также значение времени кипячения и герметичности сосудов. Его заключение сводилось к тому, что если герметичные сосуды с настоями кипятить достаточное время и исключить проникновение в них воздуха, то в таких настоях микроорганизмы никогда не возникнут. Однако Л. Спа-ланцани не сумел убедить своих современников в невозможности самозарождения микроорганизмов. Идею самозарождения жизни продолжали защищать многие выдающиеся философы и естествоиспытатели того времени (И. Кант, Г. Гегель, X. Гей-Люссак и др.).
В 1861-1862 гг. Л. Пастер представил развернутые доказательства невозможности самозарождения в настоях и растворах органических веществ. Экспериментально он доказал, что источником загрязнений всех растворов являются бактерии, находящиеся в воздухе. Исследования Л. Пастера произвели огромное впечатление на современников. Англичанин Д. Тиндаль (1820-1893) нашел, что некоторые формы микробов очень резистентны, выдерживая нагревание до 5 часов. Поэтому он разработал метод дробной стерилизации, называемый сейчас тиндализацией.
Опровержение доктрины абиогенеза сопровождалось формированием представлений о вечности жизни. В самом деле, если самозарождение жизни невозможно, рассуждали многие философы и ученые, то тогда жизнь вечна, автономна, рассеяна во Вселенной. Но как она попала на Землю? Чтобы ответить на этот вопрос, шведский ученый Аррениус (1859-1927) в начале нашего века (1912) сформулировал гипотезу панспермии, в соответствии с которой жизнь существует во вселенной и переносится в простейших формах с одного небесного тела на другое, включая Землю, под давлением световых лучей. Сторонники этой гипотезы считали, что перенос жизни на Землю возможен и с помощью метеоритов. Однако гипотеза панспермии вызывала возражения в том плане, что в космическом пространстве действуют факторы, которые губительны для микроорганизмов и что эти факторы исключают циркуляцию микроорганизмов за пределами Земной атмосферы. Становилось все более ясным, что жизнь уникальна, что истоки жизни следует искать на Земле.
Еще в XVII-XVIII вв. внимание натуралистов и философов привлекал ряд вопросов, обсуждение которых расшатывало догмы о неизменяемости органического мира. Один из таких вопросов касается изменяемости видов. Тогда для многих мысль о возможном изменении организмов под влиянием климата, пищи, почвы, упражнений и неупражнений органов, гибридизации и других факторов казалась само собой разумеющейся. Например, Ф. Бэкон (1561-1626), Д. Рей (1628-1705), Ж. Бюффон (1707-1788), И. И. Лепехин (1740-1802), Н. Я. Озерецковский (1750-1827) и др. допускали изменение организмов под влиянием климата и почвы, а Ш. Бонна признавал, что паразитические черви произошли от свободноживу-щих червей. Даже К. Линней не исключал изменяемости видов под влиянием почвы и климата, скрещиваний организмов разных видов между собой. Мысль об изменяемости видов поддерживали Эразм Дарвин (1731-1802), К. Ф. Вольф (1733-1794), французские философы-материалисты Д. Дидро (1713-1784), Ж.-О. Ламеттри (1709-1751), П. Гольбах (1723-1789). Допущение изменяемости видов часто сопровождалось верой в наследование организмами приобретенных свойств. Однако мысль об изменяемости видов еще была далека от идеи исторической преемственности видов, развития органического мира от низшего к высшему.
Не меньшее значение в то время имел вопрос о «естественном родстве» организмов. Речь шла о группировке организмов на основе их естественного родства, о допущении, что отдельные организмы могли произойти от общих родоначальников. Например, Ж. Бюффон считал, что могли быть «общие родоначальники» для нескольких семейств, в частности для млекопитающих, им допускалось 38 общих родоначальников. В России мысль о происхождении организмов ряда видов от общих родоначальников развивал П. С. Пал-лас (1741-1811).
Далее, привлекал внимание вопрос о факторе времени в изменении организмов. В частности, значение фактора времени для существования Земли и формирования на Земле органических форм признавали И. Кант (1724-1804), Д. Дидро, Ж. Бюффон, М. В. Ломоносов (1711-1765), А. Н. Радищев (1749-1802), А. А. Каверзнев (1748-?). И. Кант определял возраст Земли в несколько миллионов лет, а М. В. Ломоносов писал, что время, которое было необходимо для создания организмов, является большим церковного исчисления. Признание фактора времени имело несомненное значение для исторического понимания развития организмов. Однако представления о времени в тот период сводились лишь к мысли о неединовременнос-ти появления организмов разных видов, но не к признанию развития организмов во времени.
Важное значение тогда имел вопрос о последовательности природных тел. Значительный вклад в формирование идеи последовательности природных тел принадлежит Ш. Боннэ и Г. Лейбницу. В России эту идею поддерживал А. Н. Радищев. Не имея достаточных знаний об организмах, Ш. Боннэ, Г. Лейбниц и другие натуралисты того времени возродили аристотелевскую «лестницу природы» . Расположив на ней организмы по ступеням (на главной ступени оказался человек), они создали «лестницу существ», в которой имелись непрерывные переходы от Земли и камней к Богу. Ступеней в лестнице было столько, сколько есть животных. Отражая мысль о единстве и связи живых форм, об усложнении организмов, «лестница существ» в целом явилась порождением метафизического мышления, ибо ее ступени отражали простое соседство, но не результат исторического развития.
Существенное внимание в те времена привлекал вопрос о «прототипе» и единстве плана строения организмов. Допуская существование исходного существа, многие признавали единый план строения организмов. Дискуссии по этому вопросу имели важное значение для последующих представлений об общности происхождения.
Для многих большой интерес привлекал вопрос о трансформации организмов. Например, французский натуралист Б. де Маис (1696-1738) считал, что в море живут вечные семена жизни, которые дают начало морским живым формам, трансформирующимся затем в земные организмы. Отмечая позитивную роль трансформизма в эволюционизме, следует все же отметить, что он был механическим и исключал мысль о развитии, об историзме.
Наконец, центром внимания в то время был вопрос о возникновении органической целесообразности. Многие философы и натуралисты признавали, что целесообразность не изначальна, что она возникла естественным путем в результате браковки дисгармонических организмов. Обсуждение этого вопроса продвигало эволюционизм, но не достигало существенного результата, ибо появление одной формы рассматривалось независимо от появления другой.
Итак, к концу XVIII в. появились идеи, противоречащие представлениям о неизменяемости видов, но они не сложились в систему взглядов, а метафизичность мышления мешала полностью отвергнуть религию и взглянуть на природу по-новому. Первым, кто специально обратился к изучению проблем эволюции, был французский ученый Ж.-Б. Ламарк (1744—1829). Созданное им учение явилось завершением предыдущих поисков многих натуралистов и философов, пытавшихся осмыслить возникновение и развитие органического мира.
Ж.-Б. Ламарк был деистом, т. к. считал, что первопричиной материи и движения является творец, но дальнейшее развитие происходит благодаря естественным причинам. По Ламарку творец осуществил лишь первый акт, создавая самые простые формы, которые затем развивались, дав начало всему многообразию на основе естественных законов. Ламарк был также антивиталистом. Считая, что живое возникает из неживого, он рассматривал самозарождение в качестве естественного закономерного процесса, являющегося начальным пунктом эволюции. Признавая развитие от простого к сложному и опираясь на «лестницу существ», Ламарк пришел к заключению о градации, в которой он увидел отражение истории жизни, развитие одних форм из других. Ламарк считал, что развитие от простейших форм до самых сложных составляет главное содержание истории всего органического мира, включая и историю человека. Однако, доказывая эволюцию видов, Ламарк полагал, что они текучи и между ними нет границ, т. е. фактически он отрицал существование видов.
Главными причинами развития живой природы по Ламарку является врожденное стремление организмов к усложнению через совершенствование. По Ламарку, эволюция идет на основе внутреннего стремления к прогрессу, а положения об упражнениях и неупражнениях органов и о передаче по наследству приобретенных под влиянием среды признаков являются законами. Как думал Ламарк, факторы среды влияют на растения и простые организмы прямо, «вылепливая» из них, как из глины, нужные формы, т. е. изменения среды приводят к изменению видов. На животных факторы среды влияют косвенно.
Изменения среды приводят к изменению потребностей животных, изменение потребностей ведет к изменению привычек, а изменение привычек сопровождается использованием или неиспользованием тех или иных органов. В обоснование этих взглядов Ламарк приводил многие примеры. Например, форма тела змей, как он считал, является результатом привычки этих животных ползать по земле, а длинная шея жирафа обязана необходимости доставать плоды на деревьях.
Использование (упражнение) органа сопровождается его дальнейшим развитием, тогда как неиспользование органа — деградацией. Изменения, индуцированные внешними условиями (обстоятельствами), наследуются потомством, накапливаются и ведут к переходу одного вида в другой.
Исторические заслуги Ламарка заключаются в том, что ему удалось показать развитие от простого к сложному и обратить внимание на неразрывную связь организма со средой. Однако обосновать эволюционное учение Ламарку все же не удалось, ибо ему не удалось выяснить подлинные механизмы эволюции. Как отмечал К. А. Тимирязев (1843—1920), Ламарку не удалось объяснить важнейший вопрос, касающийся целесообразности организмов. Учение Ламарка содержало элементы натурфилософии и идеализма, поэтому ему не удалось убедить современников в том, что эволюция действительно имеет место в природе.
Творцом первой подлинно научной теории эволюции стал великий английский ученый Чарлз Роберт Дарвин (1809-1882). Главным трудом Ч. Дарвина является книга «Происхождение видов путем естественного отбора или сохранение благоприятствуемых пород в борьбе за жизнь» (1859), существенным дополнением к которой служат его книги «Изменение домашних животных и культурных растений» (1869) и «Происхождение человека и половой отбор» (1871).
Эволюционное учение Дарвина состоит из трех разделов, а именно: совокупность доводов в пользу того, что историческое развитие организмов действительно имеет место; положение о движущих силах эволюции; представления о путях эволюционных преобразований.
Доводы в пользу того, что эволюция действительно имеет место, Ч. Дарвин черпал из разных наук. Наиболее убедительные доказательства были взяты им из палеонтологии. Например, обнаружение в древнейших слоях ископаемых остатков организмов, сильно отличающихся от современных, и постепенное увеличение сходства ископаемых остатков организмов из позднейших слоев для Ч. Дарвина было летописью эволюции. Далее, Ч. Дарвин использовал данные эмбриологии того времени, которые свидетельствовали о единстве происхождения организмов, а также данные о закономерностях распределения организмов на суше и в воде и явной зависимости организации животных и растений от условий обитания (на материках и островах), которые свидетельствовали в пользу эволюции и разных направлений эволюции на материках и островах. Наконец, он широко использовал достижения сельскохозяйственной практики.
Движущими силами эволюции Ч. Дарвин назвал наследственность, изменчивость и естественный отбор. Он считал, что наследственность и изменчивость позволяют фиксировать изменения и трансформировать их в поколениях.
Изменчивость вызывает разнообразие, а наследственность передает эти изменения потомству. Следовательно, для всех живых существ в результате изменений и скрещиваний характерна наследственная гетерогенность.
Уделяя внимание изменчивости и стремясь объяснить причины закономерных преобразований живых существ в процессе эволюции, Ч. Дарвин пришел вначале к заключению, что в искусственных условиях породы животных и сорта растений созданы искусственным отбором.
Что же касается естественных условий, то в поисках ответов на вопросы об источниках целесообразной организации и жизнедеятельности живых существ в природе, о механизмах возникновения новых форм, сохраняющихся в данных условиях существования и дающих начало новым разновидностям, затем новым видам и более крупным систематическим подразделениям Ч. Дарвин сформулировал представления о борьбе за существование и естественном отборе.
Поскольку все организмы очень склонны к интенсивному размножению, то в пределах каждого вида производится потомства больше, чем выживает. Излишнее потомство гибнет в результате борьбы за существование, формы которой очень разнообразны. По Ч. Дарвину борьба за существование происходит как между видами, так и внутри видов, причем внутривидовая борьба является более ожесточенной по сравнению с межвидовой, ибо особи обитают в одной местности, нуждаются в одинаковой пище, подвергаются одинаковым опасностям и т. д. В результате борьбы за существование выживают наиболее приспособленные, т. е. те организмы, которые обладают каким-либо признаком, обеспечивающим приспособление. Следовательно, неизбежным следствием борьбы за существование является естественный отбор. «Сохранение благоприятных индивидуальных различий и изменений и уничтожение вредных я назвал естественным отбором или переживанием наиболее приспособленных» (Ч. Дарвин). Ч. Дарвин считал, что естественный отбор является главным механизмом эволюции.
Основу представлений Ч. Дарвина о путях эволюционных преобразований составляла мысль о дивергенции (расхождении) признаков. По Ч. Дарвину в связи с изменяющимися условиями жизни и благодаря естественному отбору выживали те формы, которые наиболее полно отклонялись от исходной, притом в разных направлениях. Это отклонение происходило на основе дивергенции (расхождения) признаков и вело к уменьшению конкуренции, т. к. благодаря дивергенции организмы получали возможность использовать разнородные условия существования. Ч. Дарвин считал, что в результате дивергенции образуются разновидности, которые являются зачатком вида, т. е. дивергенция создает новые виды. По Ч. Дарвину изменения организмов происходят в силу естественной закономерности, но в то же время они случайны по отношению к результатам естественного отбора. Являясь результатом отбора, органическая целесообразность носит относительный характер. Этим Ч. Дарвин преодолел метафизическое противопоставление случайности необходимости, что до него никому не удавалось.
Заслуги Ч. Дарвина перед естествознанием имеют непреходящее значение. Он всесторонне обосновал исторический метод в применении к природе, создал теорию эволюции и изгнал из науки креационизм.
Учение Дарвина касалось эволюции крупных групп организмов, включая все таксоны, в большие отрезки времени, измеряемые геологическими масштабами и на обширных территориях. Следовательно, классический дарвинизм является учением о макроэволюции. Однако оно оказалось недостаточно разработанным в генетическом плане. Дарвин имел в виду эволюцию особей, но ведь особи живут, как сейчас известно, в популяциях. Поэтому через 8 лет после выхода «Происхождения видов» англичанин Ф. Дженкин поставил такой вопрос: «Если отбор оставляет в живых те особи, которые лишь незначительно отличаются от других, то уже при последующем скрещивании наступает „поглощение" новых признаков, т. к. партнер по скрещиванию, вероятнее всего, не имеет этого нового свойства — произойдет растворение признаков в потомстве». Дарвин называв этот вопрос кошмаром Джен-кина, т. к. никогда не мог дать удовлетворяющий ответ.
Дарвинизму длительное время не хватало генетической основы, но генетика очень быстро сама подошла к эволюции. Используя методологию классического генетического анализа, генетики стали анализировать роль отдельных факторов эволюции путем вычленения элементарных единиц и протекающих в них процессов.
Первый шаг на пути объединения дарвинизма и генетики заключался в законе Дж. Харди-В. Вайнберга, которые в 1908 г. показали, что в популяции при свободном скрещивании, отсутствии мутаций данного гена и отсутствии отбора по данному признаку соотношение генотипов АА, Аа и аа остается постоянным. Это позволило им сформулировать закон, содержание которого сводится к тому, что частоты генов в бесконечно большой панмиксической популяции без давления каких-либо внешних факторов стабилизируются уже после одной смены поколений. Однако таких популяций, как известно, в природе не существует. Поэтому значение закона заключается в том, что накопленные наследственные изменения в генофонде популяций бесследно не исчезают, т. е. частоты генов постоянны.
Исходя из закона Харди—Вайнберга и учитывая влияние отбора и возникновение новых мутаций, в 1926 г. С. С. Четвериков (1880-1959) показал, что в результате спонтанного мутационного процесса во всех популяциях создается генетическая гетерогенность. Другими словами, в популяциях всегда есть мутации, в большинстве случаев в малых количествах. Он показал также, что популяция насыщена мутациями, как губка, и что мутации служат основой (материалом) эволюционного процесса, идущего под действием естественного отбора.
Позднее Н. П. Дубинин и Д. Н. Ромашов (1932) показали, что когда популяции малы, в них происходят явления, получившие название генетико-автоматических процессов (по Н. П. Дубинину, 1931) или дрейфа генов (по С. Райту, 1932). В результате генетико-автоматических процессов изменяются частоты встречаемости генов, устраняются гетерозиготы и появляются гомозиготы. Изолированная популяция становится доминантной гомозиготной или рецессивной гомозиготной. Если дрейфует мутантный летальный ген, это ведет к вымиранию организмов. Таким образом, структура популяции зависит не только от появления новых мутаций, но и от простого изменения частоты встречаемости данного гена. Эти и другие генетические исследования связали эволюционную теорию с генетикой.
В современной теории эволюции значительное место занимают вопросы происхождения жизни на Земле, ибо в рамках теории эволюции происхождение жизни рассматривают в качестве самого первого (изначального) процесса образования живых форм (необиогенеза) из неорганических веществ, т. е. в виде химической эволюции. В наше время абиогенезу противопоставлена концепция постепенного возникновения и развития живого материала в течение длительного периода времени. Эта концепция получила название ар-хебиоза или археогенеза.
Общепризнанной теорией происхождения жизни является теория, впервые предложенная в 1924 г. А. И. Опариным (1894— 1980) в его книге «Происхождение жизни». В дальнейшем эта теория подвергалась неоднократным уточнениям со стороны ее автора. Большой вклад в ее развитие внесли и многие другие ученые. В соответствии с теорией А. И. Опарина жизнь является результатом исторического односторонне направленного развития в виде постепенного усложнения органических субъединиц и развития их в сложные системы, обладающие свойствами живого (рис. 161).
Предполагают, что Земля сформировалась 4,5-5 млрд лет назад из облаков космической пыли. Она увеличивалась по массе в результате притяжения пылевых частиц благодаря действию гравитационных сил. Чтобы понять, каким образом возникли самые первые живые формы, необходимо вспомнить, какой была древняя атмосфера. В наше время земная атмосфера состоит почти на 80% из азота, а остальные 20% представлены главным образом кислородом, двуокисью углерода и некоторыми другими газами (в очень малом количестве). Однако древняя атмосфера была составлена из водяных паров, водорода, аммиака и метана. Водяные пары образовались из воды и концентрировались в атмосфере по той причине, что Земля еще была очень горячей. По мере остывания Земли, водяные пары конденсировались, образовывая облака, выпадали с дождями на Землю, а затем вода вновь превращалась в пар, который поднимался в атмосферу. Процессы испарения воды и конденсации водяных паров продолжаются и в наше время.
В соответствии с теорией А. И. Опарина жизнь возникла в несколько стадий. На первой стадии в древней атмосфере сформировались простейшие углеводороды. Предполагают, что необиогенез начался 3—3,5 млрд лет назад.
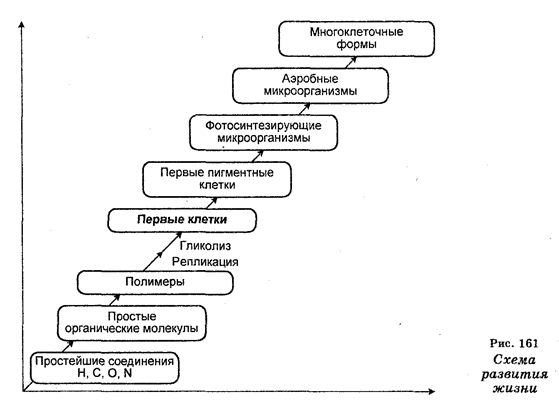
На второй стадии происходило освобождение углеводородов в атмосферу Земли, где они реагировали с водяными парами, аммиаком и другими газами. Эти реакции интенсифицировались коротковолновым УФ-излучением и электрическими разрядами и происходили в среде, содержащей очень мало кислорода, но, как отмечено выше, много водорода, метана, аммиака и паров воды. Под влиянием Солнца вода разлагалась на кислород и водород. Водород улетучивался, тогда как кислород окислял аммиак до молекулярного азота, а углеводороды до спиртов, альдегидов, кетонов и органических кислот. Эти соединения с дождями выпадали из влажной холодной атмосферы в моря и океаны, где они накапливались, а благодаря абиогенной полимеризации и конденсации становились по строению близкими к тем химическим соединениям, которые входят в состав живых организмов. Так возникали полимерные соединения, подобные белкам, нуклеиновым кислотам и АТФ. Вода морей и океанов становилась как бы «первичным бульоном».
На третьей стадии в «первичном бульоне» происходило образование так называемых коацерватных (от лат. acervatus — скрученный, нагроможденный) капель, которые, достигнув определенной величины, приобретали свойство открытых систем, реагирующих со средой. А. И. Опарин считал, что в процессе эволюции органических субстанций коацервация оказалась мощным средством концентрации соединений высокой молекулярной массы, в частности белковоподобных субстанций, растворенных в океане. В дальнейшем шло совершенствование и селекция коацерватов, которые становились устойчивыми системами, обособленными от среды и способными к реакциям (рис. 162).
На четвертой стадии у коацерватов совершенствовался метаболизм, синтезировались и упорядочивались мембраны, в результате чего установилась корреляция между нуклеиновыми кислотами и белками, что означало появление первичных организмов.
Никто не знает, какими были первые живые формы, но предполагают, что ими были самореплицирующиеся (автокаталитические) молекулы. Что касается первичных организмов, то ими, вероятно, были гетеротрофы. Из-за отсутствия кислорода в примитивной земной атмосфере и гидросфере метаболизм первичных организмов был анаэробным. Как предполагают, они питались за счет органических веществ, растворенных в первичном океане. Однако запасы абиогенно возникших органических субстанций постепенно истощались, что давало преимущество тем организмам, которые обладали способностью поглощать свет и создавать органические вещества из неорганических соединений углерода. В результате отбора в условиях повышенной концентрации СО^ в среде возник биологический путь синтеза органического вещества — фотосинтез. Опираясь на время возникновения цианобактерий, предполагают, что это произошло около 3,5´109
лет назад. Вслед за фотосинтезом наступило расхождение организмов на растения и животных.
Прогрессивное развитие первичных организмов и естественный отбор обеспечили затем возникновение автотрофов, использующих солнечную (световую) энергию для синтеза органических соединений из неорганических. В свою очередь это способствовало дальнейшему непрерывному синтезу органических веществ и появлению свободного газообразного кислорода в атмосфере, что полностью изменило жизненные процессы на Земле.
Близкие взгляды развивал Н. Г. Холодный (1882-1953), который тоже считал, что вначале образовались углеводороды, а затем из них в результате окисления образовались органические кислоты, спирты и т. д. В отличие о.т А. И. Опарина, Н. Г. Холодный считал, что жизнь возникла не в мировом океане, а в мелководьях после появления суши, что способствовало более интенсивной концентрации органических веществ и образованию коацерватов. Превращение пробиотических структур (пробионтов) в примитивные организмы произошло с окончательным высыханием мелководий.

Близкие взгляды развивал также английский ученый Д. Бернал (1901—1971), который полагал, что неорганическими предшественниками были углекислота, неорганические ортофосфаты, аммиак, сероводород и что первичный синтез органических соединений заключался в образовании очень простых молекул (состоящих из трех-шести атомов) из воды, метана и аммиака. В последующем в результате полимеризации возникли более сложные структуры, связанные с клеточной организацией. По Д. Берналу возникновение жизни шло по схеме атом — молекула — мономер — полимер — организм. Но в отличие от А. И. Опарина, он считал, что конденсация органических молекул происходила не путем образования коацерватов, а путем адсорбции первых полимеризованных структур на минеральных частицах, причем это происходило не в океане, а в илистых пластах под водой, в попеременно сухих и влажных пластах земли.
Теория А. И. Опарина нашла международное признание, ибо она имеет ряд доказательств, среди которых важнейшими являются результаты экспериментов, выполненные американским ученым С. Миллером еще в 1953 году. Пропуская электрический разряд через нагретую смесь, состоящюю из метана, водорода, аммиака и водяных паров, и, таким образом, моделирующую древнюю атмосферу, а затем охлаждая водяные пары (имитируя осадки), С. Миллер получил абиогенным путем аминокислоты (глицин и аланин), альдегиды, молочную, уксусную и другие кислоты (рис. 163). В исследованиях других ученых, которые вместо электрического разряда использовали ионизирующее излучение, УФ-излучение и тепло, было показано абиогенное образование других аминокислот, азотистых оснований, полисахаридов.
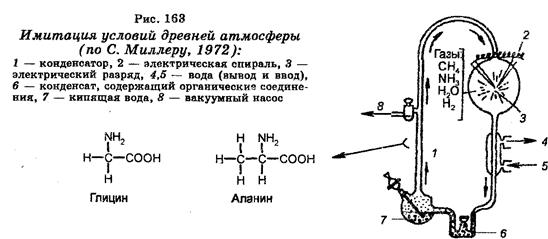
В дальнейшем американский ученый С. Фоке (1977) установил, что при повышенных температурах и удалении свободной воды из смесей аминокислот имеет место поликонденсация аминокислот с образованием структур типа протеиноидов (белковопо-добных структур) молекулярной массой 4000-10 000 дальтон. В этих структурах последовательность аминокислот была произвольной без соблюдения какого-либо порядка. Тем не менее эксперименты С. Фокса явились существенным основанием к предположению роли высыхания в образовании простейших полипептидов. По данным других исследователей под действием УФ-излучения или ионизирующего излучения в водных растворах нуклеотидов также может происходить их поликонденсация с образованием связей 3'—5'.
Имея ряд доказательств, теория А. И. Опарина в наше время оказалась фундаментальной основой в формировании дальнейших представлений о происхождении жизни. С ней не согласны лишь «научные» креационисты, которые считают, что Земля возникла 10 000 лет назад и имеет сверхъестественное происхождение. Они считают также, что 10 000 лет назад появились растения и животные в современном виде и что их происхождение является тоже сверхъестественным. Однако, как и всякая другая научная теория, теория А. И. Опарина непрерывно подвергается совершенствованию. В частности, в последнее годы стали появляться новые научные данные, на основе которых сложилось представление о том, что жизнь возникла не в океане и не в мелководье, как полагали А. И. Опарин и Н. Г. Холодный, а в геотермальных источниках в раннем периоде (в архее) истории Земли и что первым полимером были молекулы РНК. Понимание РНК в качестве первого биологического полимера (по времени возникновения) означает совершенно новое представление, а именно: жизнь началась с РНК. Предполагают, что местами абиогенной поликонденсации азотистых оснований РНК (формирования цепей РНК) могли служить такие минералы, как цеолиты, характеризующиеся трехмерной сетчатой структурой. Эти минералы могли быть катализаторами (катализирующими свойствами обладала их поверхность), сходными с белками-ферментами, появившимися значительно позднее. Поэтому, располагаясь в трещинах вулканических пород вблизи геотермальных источников, цеолиты «плавали» в горячих растворах (40°С) с высоким содержанием метана, аммиака, фосфатов и рН порядка 8,0, катализируя при этом сборку олигонуклеотидов в РНК. В последующем молекулы РНК развились в самореплицирующиеся структуры, которые затем стали изолированными от цеолитов. Наконец, они объединились с липидами, что способствовало подготовке их к жизни в океане, где завершалось образование первичных жизненных форм.
Взгляды на то, что жизнь начиналась с РНК, согласуются с открытием в 1982 г. энзиматической активности ряда молекул РНК, получивших название рибозимов и сложившимися представлениями о «мире РНК». Трехмерная структура рибозимов имеет вид «головки молотка». Они могут кодировать информацию в наследуемой форме и катализировать химические реакции в отсутствие белков. В частности, известны рибозимы, которые действуют как олигонуклеотидные лигазы, полинуклеотидные киназы и изомера-зы, а также обладают способностью катализировать реакции алки-лирования. Исходя из данных, в соответствии с которыми молекулы РНК обладают как информативными свойствами, так и способностью катализировать довольно широкий набор реакций, стали предполагать, что они обеспечивали метаболизм еще до появления белкового синтеза, собирая себя самостоятельно в нуклео-тидном бульоне. В пользу этого предположения свидетельствуют данные об образовании цитозина в результате реакция цианоацет-альдегида в концентрированном растворе мочевины (условия испаряющихся лагун или высыхающих побережий на ранних этапах развития Земли).
Первичные молекулы РНК развивались затем в самореплици-рующиеся структуры благодаря мутациям и рекомбинациям. На следующей стадии РНК-молекулы стали синтезировать белки, вначале посредством развития адапторных молекул РНК, которые могли связывать активированные аминокислоты, затем посредством расположения их соответственно РНК-шаблону, используя молекулы РНК рибосом. Впоследствии появилась ДНК, информация в которой копировалась РНК посредством реверсивной (обратной) транскрипции, после чего РНК приобрела промежуточную роль.
Предполагают, что мир РНК связывает «пребиотический мир» с ДНК. Известны экспериментальные данные, свидетельствующие о том, что олигонуклеотиды РНК обеспечивают синтез комплементарных цепей так называемой пептидной нуклеиновой кислоты (ПНК), в которой вместо фосфодиэфирных скелетных связей скелетом является амид.
В пользу представлений о пребиотической стадии эволюции жизни на Земле служат данные и о том, что одноцепочечная ДНК способна лигировать олигодезоксинуклеотиды в присутствии некоторых ионов. Следовательно, ДНК тоже обладает каталитической активностью, действуя как металлофермент (ДНК-зим). Следовательно, и РНК и ДНК обладают двойной способностью: катализировать реакции и хранить генетическую информацию.
Итак, в соответствии с моделью «мир РНК» в развитии жизни было время, когда все биологические реакции катализировались РНК. Если данное предположение справедливо, тогда оно ведет к решению давнего парадокса «что раньше, курица или яйцо», возникшего на основе его современного парадокса «что раньше, функциональные белки или нуклеиновые кислоты с их информативными свойствами».
Между тем возникает важнейший вопрос, а именно: если появление РНК действительно имело место на ранних стадиях эволюции на Земле, то существовала ли она на самом начальном этапе возникновения жизни, т. е- была ли она в самом «начале»? Вопрос этот очень труден для объяснений. Здесь возможны пока лишь самые общие допущения. Поскольку экспериментальный синтез рибонуклеотидов и неэнзиматическая репликация РНК в предполагаемых пребиотических условиях могла быть маловероятной, то легче допустить, что жизнь началась не с РНК, а с более простых репликационных систем, которые позднее (но на ранних этапах), были заменены РНК, ставшей играть ведущую роль в возникновении жизни. Такой системой мог быть, например, полимер, состоящий из белковоподобного скелета с азотистыми основаниями. Этот полимер мог выполнять роль шаблона в реакциях сборки более сложных соединений. Итак, возникновение жизни можно представить в ряде последовательных стадий, а именно: пребиоти-ческие химические процессы -» пре-РНК-мир (первые саморепли-цирующиеся молекулы) -> РНК-мир (биохимические реакции, катализируемые генетически кодируемыми рибозами) —> ДНК/белок-мир (биохимические реакции, катализируемые ДНК-кодируе-мыми белковыми ферментами) -> первые живые одноклеточные организмы. В соответствии с результатами молекулярно-генети-ческих исследований общий предок живых форм появился 2 млрд лет назад. Напротив, палеонтологические находки древнейших цианобактерий свидетельствуют о том, что такой предок существовал уже 3,5 млрд лет назад, а по результатам обнаружения углеродистых включений в скальных апатитах предполагают, что жизнь существовала 3,8 млрд лет назад.
Поскольку эволюция представляет собой процесс, протекающий во времени, то узловым вопросом является определение этого времени. Главным путем определения эволюционного времени, т. е. определения геологической эпохи, к которой относятся палеонтологические находки. Поэтому представления о ходе эволюции, т. е. об основных этапах развития жизни, можно получить, ознакомившись с так называемой шкалой геологического времени, которая сконструирована посредством изучения ископаемых остатков организмов и использования радиоактивных изотопов (метода радиодатирования), а также использования данных геологии, биогеографии, систематики и других наук. С помощью этой шкалы можно проследить все изменения земной фауны и флоры во времени (табл. 32).
Историю Земли подразделяют на несколько очень крупных последовательных промежутков времени, называемых эрами, которые в свою очередь подразделяют на периоды, а периоды — на эпохи. Иногда эти подразделения продолжают, подразделяя эпохи на века. Эрами являются катархей, архей, протерозой, палеозой (возраст рыб и амфибий), мезозой (возраст рептилий) и кайнозой (возраст млекопитающих).
Конец одной эпохи и начало другой сопровождались существенным преобразованием лика Земли, менялось соотношение между размерами суши и морей, происходили горообразовательные процессы, менялся климат. Изменения лика Земли сопровождались сменой флор и фаун, изменениями в структуре биогеоценозов.
Предполагают, что жизнь возникла на границе катархея и архея, который начался около 3,5 млрд лет назад и длительность которого составляет около 2 млрд лет. Эта эра характеризовалась широкой вулканической деятельностью. Обнаружение в архейских породах графита и чистого углерода предположительно указывает на существование в этой эре очень простых живых организмов растительной и животной природы, в частности архебактерий и цианобактерий.
В протерозое происходило отложение большого количества осадков, неоднократно формировалось ледниковое покрытие Земли, активной была вулканическая деятельность. Обнаружение ископаемых остатков медуз, грибов, водорослей, плеченогих, членистоногих (ракоскорпионов) и других организмов, живших в ту эру, свидетельствует о том, что уже тогда существовали микроорганизмы, многие типы водных простейших и беспозвоночных. Предполагают, что первыми микроорганизмами были анаэробы. Земных форм жизни еще не было, но уже тогда произошли переходы от неядерных к ядерным формам и от одноклеточных к многоклеточным, что представляет собой крупнейший скачок в развитии жизни, в становлении ее в качестве геологического фактора.
В палеозойской эре возникли представители основных групп растений и почти всех типов и классов животных (кроме птиц и млекопитающих). Можно сказать, что эта эра знаменовала важнейший этап в развитии жизни, связанный с завоеванием растениями и животными суши.
В частности, в первом периоде эры (кембрии) жизнь по-прежнему существовала лишь в воде. В морях были широко распространены одноклеточные и многоклеточные водоросли, примитивные ракообразные и паукообразные формы, например, мечехвосты, морское дно было заселено губками, кораллами и иглокожими, брюхоногими и двухстворчатыми моллюсками. Очень распространены были плеченогие (сидячие животные с двухстворчатой раковиной) и трилобиты (примитивные членистоногие). В то время уже существовали все виды морских животных, кроме хордовых. Материки оставались безжизненными.
В ордовикский период продолжалось начавшееся в кембрии погружение материков в воду. Теплел климат. Начался выход растений на сушу. В пресных водах обитали гигантские скорпионы. В воде появились первые позвоночные. В частности, в пресных водах обитали щитковые, представлявшие собой примитивных бесчелюстных рыб, не имевших парных плавников.
В течение силура продолжался выход растений на сушу. Первыми растениями были травянистые псилофиты (плауны, хвощи) и древесные формы ликопсидов, сходные с папоротниками. Существовали все ветви беспозвоночных. Начался выход на сушу животных (паукообразных). Появились животные, дышащие воздухом.
В девонский период этой эры поднималась суша, сокращалась площадь морей. Климат стал более континентальным. На суше впервые появляются примитивные голосеменные растения (семенные папоротники), появились первые леса гигантских папоротников, хвощей и плаунов, появились первые настоящие листья. В пресных водах появились настоящие акулы. Предки костных рыб дали начало двоякодышащим, кистеперым и лучеперым рыбам. Луче-перые дали начало в более поздние периоды костистым рыбам, а кистеперые стали предками наземных позвоночных.
В течение каменноугольного периода (карбона) этой эры происходит небольшое поднятие суши и потепление климата. Заболоченные леса состояли из обычных папоротников, семенных папоротников (появились первые настоящие семена) и широколиственных зеленых растений, давших начало залежам каменного угля. В изобилии были земноводные, широко распространились пресмыкающиеся. Появились первые насекомые (предки стрекоз и тараканов).
В течение пермского периода этой эры происходили разные изменения рельефа Земли и климата. Поднятие суши сопровождалось похолоданием климата. Вымирают папоротникообразные, шире стали распространяться голосеменные растения. Типичными растительными формами в тот период были цикадофиты и хвойные. На Земле в изобилии обитали пресмыкающиеся (поликозавры и терапсиды). Вымирает более 80% видов морских беспозвоночных, но в морях увеличилось количество костных рыб и головоногих моллюсков. Значительного развития достигают пресмыкающиеся (растительноядные и хищные), среди которых большое место занимали динозавры, принадлежащие к типам птицетазовых (травоядных) и ящеротазовых (хищных).
В мезозойской эре продолжалось поднятие суши, иссушался климат. В эту эру происходило возникновение и вымирание разных пресмыкающихся. Мезозой часто называют эпохой рептилий.
В триасовом периоде этой эры достигают расцвета семенные папоротники, цикадовые и хвойные. В морях в еще большей мере повысилось количество костных рыб и головоногих моллюсков.
В юрском периоде этой эры появляются покрытосеменные растения (цветковые), древнейшие птицы, расширяется многообразие морских пресмыкающихся (ихтиозавров, плезиозавров). Уже существует четыре отряда млекопитающих, все представители которых были яйцекладущими животными.
В меловом периоде этой эры были распространены покрытосеменные растения (магнолии, пальмы, клены, дубы и др.). Появились первые цветки. Отмечено появление современных костных рыб и исчезновение морских пресмыкающихся. Вымирают динозавры. Крупные пресмыкающиеся (крокодилы, черепахи, гаттерии) сохраняются лишь в экваториальном поясе. Впервые появляются змеи, птицы. Распространяются млекопитающие, способные рождать живых детенышей (сумчатые).
В кайнозойской эре произошло интенсивное горообразование, похолодание климата. Эта эра характеризуется временем цветковых растений, насекомых, птиц, млекопитающих и появлением человека. В этой эре возникли морские организмы всех' современных групп, на Земле появились травянистые покрытосеменные растения, птицы, современные млекопитающие, включая обезьян.
В третичном периоде этой эры различают эпохи палеоцен, эоцен, олигоцен, миоцен и плиоцен. В палеоцене уже существовали примитивные плацентарные млекопитающие. В палеоцене и эоцене возникли первые плотоядные хищники и древнейшие копытные. В эоцене и олигоцене на смену первым хищникам пришли формы, давшие начало современным кошкам, собакам, медведям и ласкам, а также тюленям и моржам. Что касается копытных (лошадей, верблюдов и слонов), то их эволюция шла в сторону увеличения размеров тела и изменения количества пальцев.
Таблица 32
Геохронологическое подразделение истории Земли
| Эра
|
Период
|
Эпоха
|
Продолжи-
тельность
(в млн. лет)
|
Время от начала периода до наших дней (в млн. лет)
|
Преобладающие формы жизни
|
Климат
|
Геологические условия
|
| Кайнозой (новая)
|
Четвертичный
|
Совре-
менная
|
0,02
|
1
|
Травянистые растения, млекопитающие. Эпоха человека
|
теплый
|
Конец последнего ледникового периода
|
| Поейсто-цен
|
1
|
1,7
|
Вымирание многих видов растений и крупных млекопитающих.
Появление человека
|
Холодный
|
Повторные оледенения (четыре ледниковых периода)
|
| Третичный
|
Плиоцен
|
12
|
13
|
Цветковые растения. Млекопитающие (появления лонов, лошадей, верблюдов, сходных с современными)
|
Прохладный
|
Поднятие отдельных гор. Остепенение суши. Вулканическая активность
|
| Миоцен
|
13
|
25
|
Появление первых человекообразных обезьян
|
Прохладный
|
Образование гор. Вулканическая активность
|
| Олигоцен
|
11
|
36
|
Леса. Однодольные цветковые растения. Предшественники многих современных родов млекопитающих (включая человекообразных обезьян и людей)
|
теплый
|
Материки низменные
|
| Эоцен
|
22
|
58
|
Пацентарные млекопитающие
|
теплый
|
Горы размыты. Внутриконтинентальных морей нет
|
| Палеоцен
|
5
|
63
|
Примитивные млекопитающие. Угасание царства динозавров
|
| Мезозой
|
Мел
|
72
|
136
|
Покрытосесенные однодольные и двудолные растения. Первые цветки. Динозавры. Примитивные млекопитающие
|
Образуются горы, отложения мела. Сланцев
|
| Юра
|
46
|
181
|
Цикадофиты и хвойные. Динозавры. Древнейшие птицы, насекомоядные, сумчатые
|
Материки довольно возвышены. Мелководные моря
|
| Триас
|
49
|
230
|
Голосеменные, динозавры
|
Теплый, усиливается засушливость
|
Материки приподняты над уровнем моря
|
Продолжение табл.32.
| Эра
|
Период
|
Эпоха
|
Продолжи-
тельность
(в млн. лет)
|
Время от начала периода до наших дней (в млн. лет)
|
Преобладающие формы жизни
|
Климат
|
Геологические условия
|
| Герцинское горообразование
|
| Палеозой (древний)
|
Пермь
|
50
|
280
|
Голосеменные, насекомые, пресмыкающиеся. Появление предков динозавров
|
Аридный
|
Материки приподняты, периодическое оледенение
|
| Карбон (каменноугольный)
|
65
|
345
|
Папоротникообразные, плауны, голосеменные, первые настоящие семена. Земноводные
|
Теплый, влажный
|
Болота, поднятие суши, материки низменнные
|
| Девон
|
60
|
405
|
Первые леса (папоротники). Первые настоящие листья. Рыбы.
|
Аридный
|
Небольшие внутриконтинентальные моря. Оледенение
|
| Силур
|
20
|
425
|
Водоросли, хвощи, плауны. Морские паукообразные (скорпионы). Начался выход животных на сушу
|
Засушливый
|
Обширные внутриконтинентальные моря. Поднятие суши
|
| Ордовик
|
75
|
500
|
Морские водоросли. Наутилоидные. Трилобиты, моллюски
|
Теплый, включая Арктику
|
Заметное погружение суши
|
| Кембрий
|
100
|
600
|
Морские водоросли, «взрыв» морских многоклеточныех. Трилобиты и плеченогие, губки и иглокожие
|
Умеренный
|
Материки низменные
|
| Второе великое горообразование
|
| Протерозой
|
1000
|
1600
|
Примитивные водоросли и водные грибы. Морские простейшие. В конце – моллюски, черви и др. Морские беспозвоночные
|
Интенсивное осадкообразование. Вулканическая активность, обширная эрозхия и многократные обледенения
|
| Первое великое горообразование
|
| Архей
|
2000
|
3600
|
Ископаемые не обнаружены (обнаружено органическое вещество в скальных породах)
|
Значительная вулканическая активность. Слабое осадкообразование. Обширная эрозия
|
Копытные в ходе эволюции разошлись на группу с четным числом пальцев (коровы, овцы, верблюды, олени, жирафы, свиньи и гиппопотамы) и группу с нечетным числом пальцев (лошади, зебры, тапиры, носороги). В эоцене существовали китообразные формы, давшие начало китам и дельфинам. В олигоцене широко были распространены также предковые формы человекообразных обезьян и человека.
В миоцене и плиоцене древесная растительность вытесняется травянистыми формами. Наступает расцвет покрытосеменных растений, членистоногих и млекопитающих. Конец третичного периода датируется появлением всех современных семейств животных и растений.
Следующий период (четвертичный) этой эры, длящийся последние 1,5-1,7 млн лет, состоит из плейстоценовой и современной эпох. Для плейстоцена характерны четыре последовательных оледенения. Растения и животные, которые были распространены в плейстоцене, сходны с современными растениями и животными. Однако в плейстоцене вымерли многие растения (особенно лесные) и млекопитающие (мамонты, саблезубые тигры). Появились многие травянистые формы растений, но, самое главное, появился человек. Современная эпоха началась около 11 000 лет назад, когда отступил последний ледник.
Хотя в палеонтологической летописи существуют многие пробелы, тем не менее она полностью раскрывает происхождение многих существующих в настоящее время видов животных и растений. Например, очень хорошо прослежено происхождение лошадей, слонов, парнокопытных, хищников.
Эволюцию на уровне крупных таксонов называют макроэволюцией. Основополагающие представления о макроэволюции в целом сводятся к тому, что все современные живые существа являются измененными потомками одной или нескольких форм, существовавших ранее. Более молодые по происхождению формы животных и растений произошли от предков, менее отличавшихся друг от друга, а те, в свою очередь, развились от общего единого источника (рис. 164).
Считают, что первыми возникли архебактерии. Примерно 3,5 млрд лет назад. Затем возникли истинные бактерии, а от них ответвились вирусы, цианобактерии, красные водоросли и зеленые жгутиковые. От последних ответвились водоросли (от которых позднее произошли, в свою очередь, высшие растения) и бесцветные ризофлагелляты, из которых развились диатомовые водоросли, плесневые грибы, губки и простейшие. Часть простейших стала паразитической.
Вторым крупнейшим этапом эволюции была трансформация одноклеточных организмов в многоклеточные. Считают, что первые эукариоты появились 1,5 млрд лет назад. Их появление есть крупнейший эволюционный переход, ибо в основе появления эукарио-тов лежит формирование митоза и развитие клеточных структур. Из цилиарных простейших (цилиофор) развились многоклеточные (синтициальные) плоские черви. Последние разветвились на пять линий, одна из которых повела к иглокожим и хордовым. Остальные линии повели к большинству других типов животного мира.
Первыми позвоночными были остракодермы, которые развились из беспозвоночных, а из остракодерм развились миноги и панцирные акулы. Хрящевые рыбы (акулы и скатовые) и костные рыбы развились из панцирных акул, а из костных рыб развились кроссоптериги, которые являлись примитивными рыбами и которые затем разошлись на две эволюционные ветви, одна из которых привела к дипловертебронам (ранним земноводным), называемым стегоцефалами. Дипловертеброны были предшественниками современных амфибий и пресмыкающихся. Первые пресмыкающиеся (котилозавры) разветвились на ветви, давшие начало динозаврам и морским пресмыкающимся, которые затем вымерли, а также черепахам, крокодилам, ящерицам, змеям, птицам и млекопитающим.
Эволюционная линия, ведущая к человеку, начинается от ту-пай и ведет к ранним приматам, разветвляясь затем на обезьян Нового света, обезьян Старого света, понгид и гоминид. От понгид (настоящих обезьян) произошли гориллы, орангутанги и шимпанзе, а от гоминид произошел непосредственный предшественник современного человека (см. гл. XVI).
Главные направления эволюции сформулированы А. Н. Север-цовым (1866-1936), который впервые четко определил различия между прогрессивным и регрессивным развитием, обосновал понятия о морфофизиологическом и биологическом прогрессе, а также о морфофизиологическом и биологической регрессе.
Морфофизиологический прогресс — это изменения структуры и общей жизнедеятельности на пути эволюции от простых форм к сложным, от одноклеточных организмов к многоклеточным. Напротив, Морфофизиологический регресс — это упрощение организации и жизнедеятельности, выражающееся в редукции органов (например, хорды у оболочников) на пути эволюции.
Следствием морофофизиологического прогресса является биологический прогресс. Он характерен для организмов, которые характеризуются возрастанием приспособленности к окружающей среде, что ведет к увеличению их численности и расширению их ареалов. Однако биологический прогресс может быть результатом не только морфофизиологического прогресса. Иногда от является и результатом морфофизиологического регресса
В конечном итоге биологический прогресс ведет к процветанию вида.
В современную эпоху процветающими являются покрытосеменные растения, насекомые, костные рыбы, птицы, млекопитающие.
Значение биологического прогресса заключается в том, что он играет важную роль в эволюции и обеспечивает главные направления эволюции в виде ароморфозов, идиоадаптаций и общей дегенерации.
Ароморфозы — это крупные изменения организмов, которые поднимают на новый более высокий уровень их морфофизиологическую организацию и жизнедеятельность. Кроме того, они открывают организмам путь к проникновению в новую среду обитания. Ароморфозы возникают скачками. Примерами ароморфозов А. Н. Северцов называл развитие легких и четырехкамерного сердца, превращение парных плавников рыб в парные конечности земноводных. По А. Н. Се-верцову, ароморфозы обозначили возникновение организмов новых классов и типов организмов, завоевание организмами новых просторов. У млекопитающих ароморфозами были: появление живорождения, шерстного покрова, постоянной температуры тела и др.
Идиоадаптация — это изменения, которые не приводят к повышению уровня организации организмов, но сопровождаются приданием им большей приспособленности к существующим условиям жизни, больших возможностей прогрессивного развития. Примером идиоадаптаций являются приспособления насекомых к жизни в разных средах (в воде, почве, воздухе), к разным типам питания. Другим примером служит разнообразие форм у насекомоядных (землеройка, крот, еж, выхухоль и др.), что позволило занять им разные экологические ниши.
Общая дегенерация — это изменения, которые заключаются в снижении уровня организации живых существ и в снижении активности отдельных или ряда органов. Примерами общей дегенерации А. Н. Северцов называл биологию саккулины из группы паразитических ракообразных (крабов), а также биологию оболочников. Выживаемость этих организмов обеспечивается чрезвычайным разнообразием их личиночных форм.
Следствием морфофизиологического регресса является биологический регресс. Биологический регресс — это снижение приспособленности организмов к среде, сокращение их численности и ареалов. Конечным итогом биологического регресса является вымирание организмов.
Учение А. Н. Северцова о главных направлениях эволюционного процесса было развито дальше в трудах И. И. Шмальгаузена, который рассматривал организм как целое в историческом и индивидуальной развитии. Среди главных направлений эволюционного процесса И. И. Шмальгаузен большое внимание уделял различным формам ароморфозов.
Доказательства эволюции получены в разных науках. Классические доказательства эволюции получены, прежде всего, в палеонтологии в результате изучения ископаемых организмов, живших в прошлые эпохи. Предполагают, что в ходе эволюции вымерло около 200 000 видов животных. В более глубоких слоях Земли обнаруживаются остатки более древних форм жизни, тогда как в поверхностных слоях находят остатки более поздних форм. Можно сказать, что история жизни на Земле написана на языке ископаемых остатков. Палеонтологический материал дает также основания судить о темпах и направлениях эволюции.
Доказательства эволюции получены в биогеографии, которая является наукой о распространении растений и животных. В биогеографии различают шесть биогеографических областей. Каждая из этих областей характеризуется специфическими обитателями (растениями и животными), называемыми эндемиками, под которыми понимают организмы видов, родов и таксонов, ограниченных в своем распространении определенными территориями.
В Палеоарктической области (Европа, африканский север от Са-хары, часть Азии к северу от Гималаев, Азорские острова и острова Зеленого мыса) эндемичными, если говорить о животных, являются кроты, олени, быки, овцы, козы, скворцы и сороки.
В Неоарктической области (Гренландия и Северная Америка) эндемичными являются горные козлы, луговые собачки, опоссумы, скунсы, еноты, сойки и американские грифы. Кроме того здесь встречаются формы, андемичные для палеоарктической области.
В Неотропической области (Южная и центральная Америка, юг Мексики и острова Вест-Индии) обитают альпаки, ламы, цепко-хвостые обезьяны, тапиры, ленивцы, вампиры, муравьеды и многие виды птиц, не встречающиеся в других частях земного Шара.
В Эфиопской области (Африка к югу от Сахары, Мадагаскар) обитают шимпанзе, гориллы, зебры, носороги, трубкозубы, бегемоты, жирафы, многие виды птиц, пресмыкающихся и рыб, не обнаруживаемые в других областях.
В Восточной области (Индия, Цейлон, Индокитай, юг Китая, Малайский полуостров и отдельные острова Малайского архипелага) обитают орангутанги, черные пантеры, индийские слоны, гиббоны и долгопяты. В Австралийской области (Австралия, Новая Зеландия, Новая Гвинея и др. острова Малайского архипелага) эндемичными являются утконосы, кенгуру, вомбаты, коала и другие сумчатые животные. Эндемичными являются бескрылые птицы эму и казуар, а также птица-лира и какаду.
Географические закономерности, характерные для фауны, присущи и флоре этих биогеографических областей.
Одно из основных положений биогеографии заключается в том, что каждый вид растений и животных возникал только однажды и только в одном месте (центре происхождения), откуда он расселялся до тех пор, пока не встречал какую-нибудь преграду, например, географическую, климатическую, пищевую и т. д. Географические ареалы близких видов, как правило, не совпадают, но они и не очень отдалены один от другого.
Особенности географического распространения животных и растений являются отражением специфики эволюции каждого вида. Например, сходство фауны Неоарктической и Неотропической областей обусловлено относительно недавним геологическим разделением Северной Америки и Евразии в результате образования Берингова пролива. Наличие в Австралии однопроходных и сумчатых, которых нет ни в одной из других областей, является результатом того, что этот континент давно (в мезозое) был изолирован от других частей суши. Примитивным млекопитающим — аборигенам Австралии не довелось конкурировать с более приспособленными плацентарными млекопитающими, поэтому они дали начало формам, легко приспособившимся к местным условиям.
Далее, сейчас аллигаторы встречаются только в реках юго-востока США и в р. Янцзы (КНР), тогда как магнолии произрастают только в восточной части США, в КНР и Японии. В начале кайнозоя Северная Америка соединялась с Азией (на месте теперешнего Берингова пролива). При тогдашнем теплом климате аллигаторы и магнолии на этой территории были распространены повсеместно. В дальнейшем из-за поднятия Скалистых гор и похолодания (оледенения) аллигаторы и магнолии погибли повсеместно, кроме юго-восточной части США и восточной части КНР, где не было оледенения. Поскольку виды магнолий и аллигаторов были разделены в отдаленных частях земного шара на протяжении многих миллионов лет и эволюционировали обособленно, то в современную эпоху, оставаясь близкородственными видами, они несколько различаются между собой.
Еще Ч. Дарвин отмечал различия островных и материковых флор и фаун. Например, на Галапагосских островах, возраст которых составляет всего лишь 3—5 млн лет и которые никогда не были связаны с материком, отсутствуют лягушки и жабы, ибо ни они, ни их яйца не смогли бы выжить в морской воде, с которой неизбежно пришлось бы столкнуться мигрирующим на этот остров животным. На этих островах нет также наземных млекопитающих, но на одном из этих островов встречаются гигантские ящерицы и черепахи, которые очень близки некоторым формам, обитающим на побережье Эквадора. Это указывает на эволюцию растений и животных после заселения этих островов.
В систематике главное доказательство эволюции заключается в том, что все живые существа можно расположить в иерархическую систему таксономических единиц — виды, роды, семейства, отряды, классы и типы. Это означает, что все организмы связаны между собой филогенетически в результате существующих между ними эволюционных взаимоотношений. Принадлежность организмов к тем или иным систематическим группам свидетельствует о том, что большинство промежуточных форм, существовавших в прошлом, вымерло. Если бы виды всех существовавших в прошлом организмов жили до настоящего времени, то классифицировать живой мир на таксономические группы было бы невозможно. Будучи генетически различными, виды представляют собой независимо эволюционирующие и репродуктивно изолированные единицы. Поскольку можно предполагать, что у генетически сходных видов общий предок существовал в менее отдаленном прошлом по сравнению с генетически различными видами, то степень генетических различий является, по существу, мерой, на основе которой сейчас совершенствуют филогенетическое древо,
Степень генетических различий между видами определяют либо прямо путем изменений последовательностей нуклеотидов в генах, либо косвенно путем изменений последовательностей нуклеотидов в рРНК, или последовательностей аминокислот в белках. Результаты сравнения последовательностей ДНК разных организмов позволяют определить количество пар нуклеотидов, в которых в ходе эволюции имели место замены азотистых оснований (табл. 33), тогда как сравнение белков от разных организмов позволяет определить различия в аминокислотных последовательностях, т. е. судить о близости организмов (рис. 165) и о связи последовательностей со скоростью эволюции (табл. 34, рис. 166). На основе данных о филогении отдельных белков строят филогенетическое древо, которое, как показано для цитохрома С, совпадает с филогенетическим древом, построенным по ископаемым останкам. На основе реконструкции филогении и определения степени генетических различий по аминокислотным последовательностям ряда белков считают, что гены, кодирующие эти белки у животных, происходят от общего предка.
Таблица 33
Количества нуклеотидных замен в генах, кодирующих
цитохромы С у разных организмов
| Организм
|
1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
14
|
16
|
16
|
17
|
18
|
19
|
20
|
| 1. Человек
|
—
|
1
|
13
|
17
|
16
|
13
|
12
|
12
|
17
|
16
|
18
|
18
|
19
|
20
|
31
|
33
|
36
|
63
|
56
|
66
|
| 2. Макак -резус
|
—
|
12
|
16
|
IS
|
12
|
11
|
13
|
16
|
15
|
17
|
17
|
18
|
21
|
32
|
32
|
35
|
62
|
57
|
65
|
| 3. Собака
|
—
|
10
|
8
|
4
|
6
|
7
|
12
|
12
|
14
|
14
|
13
|
30
|
29
|
24
|
28
|
64
|
61
|
66
|
| 4. Лошадь
|
—
|
1
|
5
|
11
|
11
|
16
|
16
|
16
|
17
|
16
|
32
|
27
|
24
|
33
|
64
|
60
|
68
|
| 5. Осел
|
—
|
4
|
10
|
12
|
15
|
15
|
15
|
16
|
15
|
31
|
26
|
25
|
32
|
64
|
59
|
67
|
| 6. Свинья
|
—
|
6
|
7
|
13
|
13
|
13
|
14
|
13
|
30
|
25
|
26
|
31
|
64
|
59
|
67
|
| 7. Кролик
|
—
|
7
|
10
|
8
|
11
|
11
|
11
|
25
|
26
|
23
|
29
|
62
|
59
|
67
|
| 8. Кенгуру
|
-
|
14
|
14
|
15
|
13
|
14
|
30
|
27
|
26
|
31
|
66
|
58
|
68
|
| 9. Утка
|
—
|
3
|
3
|
3
|
7
|
24
|
26
|
26
|
29
|
81
|
62
|
66
|
| 10. Голубь
|
—
|
4
|
4
|
8
|
24
|
27
|
26
|
30
|
89
|
62
|
66
|
| 11. Курица
|
—
|
2
|
8
|
28
|
26
|
28
|
31
|
61
|
62
|
66
|
| 12. Пингвин
|
—
|
8
|
28
|
27
|
28
|
30
|
62
|
61
|
65
|
| 13. Черепаха
|
—
|
30
|
27
|
30
|
33
|
65
|
64
|
67
|
| 14.Гремучая змея
|
—
|
38
|
40
|
41
|
61
|
61
|
69
|
| 15. Тунец
|
—
|
34
|
41
|
72
|
66
|
69
|
| 16.Муха комнатная
|
—
|
16
|
88
|
63
|
65
|
С целью определения степени сходства белков используют также иммунологическое и электрофоретическое сравнение белков. Степень иммунологического сходства белков выражают в иммуно-логическом расстоянии (табл. 35), которое приближенно можно связать с различиями по аминокислотным последовательностям. Электрофоретические исследования позволяют выявить электрофоретическое сходство белков и на основе этих данных также определить генетические расстояния между видами.
Таблица 34
Связь между аминокислотными различиями в и-и р-глобинах со скоростью эволюционной дивергенции
| Животные
|
a-цепь
|
b-цепь
|
a+b (среднее)
|
Миллионы лет с начала дивергенции
|
| Плацентарные в сравнении между собой
|
16,1
|
16,7
|
16,4
|
100
|
| Кенгуру в сравнении с плацентарными млекопитающими
|
21,7
|
26,9
|
24,3
|
160
|
| Куры в сравнении с млекопитающими (сумчатыми и плацентарными)
|
29,6
|
31,7
|
30,6
|
215
|
| Змеи в сравнении с теплокровными
|
39,2
|
290
|
| Земноводные в сравнении с наземными животными
|
46,7
|
48,9
|
47,8
|
380
|
| Костные рыбы в сравнении с четвероногими (рептилиями, амфибиями и млекопитающими)
|
49,3
|
49,6
|
49,5
|
400
|
Таблица 35
Иммунологические различия между приматами (по альбуминам)
| Вид приматов
|
Антисыворотка в альбуминам
|
| Человека
|
Шимпанзе
|
Гиббона
|
| Человек
|
о
|
3,7
|
14,4
|
| Шимпанзе
|
5,7
|
0
|
14,6
|
| Горилла
|
3,7
|
6,8
|
11,7
|
| Орангутан
|
8,6
|
9,3
|
11,7
|
| Гиббон
|
10,7
|
9,7
|
0
|
Эволюция подтверждается данными сравнительной морфологии, эмбриологии, физиологии, биохимии и генетики.
Результаты сравнительного изучения строения животных в пределах систематических групп показывают, что оно имеет общий план (рис. 167). Особенно это проявляется в случае гомологичных органов, которые характеризуются общим строением, сходством эмбрионального развития, иннервации, кровоснабжения и отношения к другим органам. Например, гомологичными являются передний ласт тюленя, крыло летучей мыши, передняя нога собаки и рука человека (рис. 168) и другие. Эти органы состоят из почти одинакового количества костей и мышц, их сосуды и нервы имеют сходную топографию. Гомологичные органы с несомненностью указывают на общность происхождения животных, у которых есть эти органы.
Помимо гомологичных органов морфологическим свидетельством в пользу эволюции являются данные о рудиментарных органах, являющихся «остатками» развитых органов, имевшихся в прошлом у предковой формы. Например, у человека имеется свыше 100 разных рудиментов, типичными примерами которых являются червеобразный отросток слепой кишки (аппендикс), копчик (слившиеся хвостовые позвонки), волосяной покров туловища и конечностей, третье веко. Червеобразный отросток слепой кишки является результатом перехода человека на диету с большим содержанием мяса и меньшим количеством клетчатки. Слепой вырост в таких условиях уменьшился и стал рудиментом. Напротив, у травоядных животных он представляет собой активно функционирующий орган. Кости задних конечностей китов в связи с переходом их к водному образу жизни стали рудиментами, располагающимися в толще брюшных мышц.

На эмбриологические доказательства эволюции обращал внимание еще Ч. Дарвин. В 1866 г. Э. Геккель сформулировал биогенетический закон, в соответствии с которым онтогенез есть повторение филогенеза (краткое и быстрое резюме филогенеза по терминологии Э. Геккеля), т. е. зародыши в процессе развития как бы сокращенно повторяют эволюционный путь своих предков. Как оказалось позднее, Э. Геккель был прав в принципе, но не в деталях, поскольку зародыши высших животных сходны лишь с зародышами низших животных, но не со взрослыми особями низших форм, как это считал Э. Геккель.
Эмбриологические данные в пользу эволюции сводятся к тому, что на ранних стадиях развития между зародышами млекопитающих, рыб, земноводных и пресмыкающихся существует много сходных признаков. В то же время эти организмы обладают рядом собственных признаков, присущих только им и обеспечивающих приспособленность к развитию в утробе материнского организма, но не под скорлупой яйца.
 Данные эмбриологии свидетельствуют о том, что после гаструляции развитие зародышей проходит в одном из двух направлений. С одной стороны, бластопор превращается либо в анальное отверстие, либо занимает положение около этого отверстия, как это имеет место в случае иглокожих и хордовых. С другой стороны, бластопор развивается в ротовое отверстие, либо занимает положение около этого отверстия, как это имеет место в случае кольчатых червей, моллюсков, членистоногих и др. При развитии как в одном направлении, так и другом между эктодермой и энтодермой развивается мезодерма, которая у иглокожих и позвоночных закладывается частично (карманами или выпячиваниями первичной кишки), а у кольчатых червей и других организмов закладывается из специальных клеток, дифференцирующихся на ранних этапах развития. Данные эмбриологии свидетельствуют о том, что после гаструляции развитие зародышей проходит в одном из двух направлений. С одной стороны, бластопор превращается либо в анальное отверстие, либо занимает положение около этого отверстия, как это имеет место в случае иглокожих и хордовых. С другой стороны, бластопор развивается в ротовое отверстие, либо занимает положение около этого отверстия, как это имеет место в случае кольчатых червей, моллюсков, членистоногих и др. При развитии как в одном направлении, так и другом между эктодермой и энтодермой развивается мезодерма, которая у иглокожих и позвоночных закладывается частично (карманами или выпячиваниями первичной кишки), а у кольчатых червей и других организмов закладывается из специальных клеток, дифференцирующихся на ранних этапах развития.
Вслед за появлением мезодермы у всех хордовых развивается дорзальный полый нервный тяж, а также нотохорд (основа скелета тела) и жаберные щели (отверстия в глотке). На ранних стадиях развития у зародышей, например человека, как и у зародышей рыб, имеются жаберные щели, дуги аорты, пересекающие жаберные перегородки, сердце с одним предсердием и одним желудочком, примитивная почка (пронефрос) и хвост с мышцами. Таким образом, на ранних стадиях развития зародыши человека напоминают зародыши рыб.
На более поздних стадиях развития зародыши человека приобретают сходство с зародышами пресмыкающихся, в частности, зарастают жаберные щели, сливаются кости, из которых состоят позвонки, исчезает старая почка (пронефрос) и образуется новая почка (мезонефрос), разделяется предсердие на правую и левую части. Еще позднее у зародышей человека развивается четырехкамерное сердце и метанефрос (совершенно новая почка), исчезает нотохорд. На седьмом месяце внутриутробного развития плод человека покрыт волосами и имеет «обезьянье соотношение» в размерах тела и конечностей.
Явление рекапитуляции признаков у человека объясняют действием сохранившихся «рыбьих», «земноводных» и «пресмыкающихся» генов, существовавших у предковых форм, вслед за которыми действуют гены млекопитающих, затем приматов. Следовательно, на начальных стадиях развития человека действуют гены далеких предков и проявляются признаки, сходные с признаками далеких предков, на поздних же стадиях действуют гены, характерные для млекопитающих, в частности приматов, в результате чего проявляются признаки млекопитающих и приматов. Закономерность здесь такова, что у зародышей человека раньше проявляются признаки, присущие типам и классам, а позднее признаки, присущие родам и видам.
Например, у человека и свиньи процессы развития дивергиру-ют раньше, чем у человека и человекообразных обезьян, поскольку предки человека разошлись с предками свиней раньше, чем произошло расхождение человека и человекообразных обезьян, эволюционировавших от одного предка.
Основные физиологические доказательства эволюции заключаются в сходстве физиологических процессов — дыхания, пищеварения, кровообращения, выделения, реакций на раздражения у организмов многих систематических групп.
Данные биохимии в пользу эволюции очень значительны. Во-первых, химический состав всех живых организмов является сходным. В элементарном составе организмов преобладают три элемента — С, О и Н, в атомарном составе 50-60% приходится на С (в земной коре он составляет 0,35%), что указывает на естественное родство всего органического мира. Далее, у всех организмов белки построены из 20 аминокислот. Между белками животных и растений имеется сходство. Например, между гемоглобином позвоночных, гемоцианином беспозвоночных и хлорофиллом растений существует принципиальное сходство, что указывает на единство происхождения всех организмов. Кроме того, имеется иммуноло-гическое сходство между белками крови разных животных. Например, ближайшими родственниками человека являются человекообразные обезьяны, затем обезьяны Старого света, цепкохвостые обезьяны Нового света и, наконец, долгопяты. Кошки, собаки и медведи образуют одну родственную группу, тогда как коровы, овцы, козы олени и антилопы — другую. Тюлени и морские львы стоят ближе к хищникам.
Отмечается сходство ферментов и биосинтетических путей у различных организмов. У всех животных и растений в качестве универсального звена в энергетических реакциях используется АТФ. Налицо также сходство продуктов выделения. У человека и других приматов конечным продуктом обмена пуринов является мочевая кислота, у других млекопитающих — аллантоин, у земноводных и у большинства беспозвоночных — мочевина, а у некоторых беспозвоночных — аммиак. Эволюция позвоночных сопровождалась постепенной потерей способности синтезировать ферменты, необходимые для постепенного распада мочевой кислоты.
Основное доказательство эволюции, представляемое генетикой, заключается в установлении универсального характера ядерного генетического кода. Универсальностью обладают также митохон-дриальный и хлоропластный генетические коды. Можно сказать, что ядерный и экстраядерный генетические коды — это важнейшее доказательство дарвиновской идеи единства происхождения всех форм жизни, т. е. происхождения всех живых форм из одного корня.
Наконец, в пользу эволюции свидетельствует практика сельского хозяйства, на что неоднократно указывал сам Ч. Дарвин, приводя множественные примеры происхождения культурных растений и домашних животных от диких предков. Например, все сорта белокочанной, листовой, цветной и брюссельской капусты, а также кольраби и брокколи происходят от одного вида дикой капусты, а кукуруза берет начало от травянистого растения (теосинта), произрастающего в Андах и в Мексике. Можно сказать, что среди культивируемых сельскохозяйственных растений нет ни одного вида, который был бы в «диком» состоянии. Это же заключение относится и к домашним животным. Все существующие ныне породы крупных и мелких домашних животных также происходят от диких предков.
Эволюция на уровне вида (на внутривидовом уровне) получила название микроэволюции. В рамках учения о микроэволюции различают элементарную эволюционную единицу, элементарное эволюционное явление, элементарный эволюционный материал и элементарные факторы эволюции.
Элементарной единицей эволюции является популяция. Популяция (от фр. population) — это совокупность особей одного вида, длительно (в течение многих поколений) населяющих определенную территорию, свободно скрещивающихся между собой и отделенных от других популяций этого же вида давлением изоляции. В составе вида может быть одна или несколько (много) популяций.
Для популяций характерны такие показатели, как ареал, численность особей, размеры по числу особей в пространстве, возраст, половой состав, генетическая гетерогенность (см. гл. XVII).
Популяция является элементарной эволюционной единицей по той причине, что изменения отдельных особей не приводят к эволюционным изменениям. Эволюционируют не особи, а группы особей, составляющие популяцию. Что же касается особей, то они являются объектом действия естественного отбора (см. ниже). В эволюционном процессе популяция является неделимой единицей, характеризующейся экологическим, морфофизиологическим и генетическим единством. В существовании элементарных эволюционных единиц (популяций) проявляется такое важнейшее свойство жизни, как дискретность.
Совокупность всех генов во всех их аллельных формах в гаметах организмов, составляющих популяцию, представляет собой пул генов, тогда как совокупность всех генотипов популяций называют их генофондом. В случае индивидуального диплоидного организма частота какого-либо аллеля может составлять 100%, 50% или 0%, но в популяции организмов частота этого аллеля является функцией количества (%) индивидуумов в популяции, которые имеют этот ген. Поэтому в популяции организмов частота аллеля может составлять от 0 до 100%. Стойким является тот генетический пул, в котором частоты генов, независимо от их функций, являются постоянными во всех следующих одно за другим поколениях организмов.
В популяционной генетике различают разницу между частотой генов (аллелей) и частотой генотипов. Допустим, какая-либо воображаемая популяция состоит из 100 индивидуумов, из которых 20 гомозиготны по аллелю а, 60 гомозиготны по аллелю А, а 20 гетерозиготны. Следовательно, в этой популяции количество людей с генотипом АА составит 60%, с генотипом Аа — 20% и с генотипом аа — тоже 20%. Из этих частот возможно вычисление общего числа генов и частот генов в популяции (табл. 36).
Представления о поведении в популяциях генных аллелей отражены в названном выше законе Дж. Харди и В. Вайнберга, в соответствии с которым в постоянной популяции при отсутствии свободного скрещивания и давления других факторов (отбор, мутации и др.) частота доминантных и рецессивных аллелей была, есть и будет константной. Этот закон предсказывает, что частоты генов порядка 0,7 для А и 0,3 для гена а будут поддерживаться и в будущих поколениях, если популяция является устойчивой. Частоты генотипов будут изменяться, если в популяции начнутся случайные скрещивания, но до тех пор, пока не достигнут равновесия. Если частоты аллелей у исходных организмов были одинаковы у обоих полов, то при случайных скрещиваниях равновесие частот генотипов по определенному локусу достигается уже за одно поколение. В свою очередь равновесие частот генотипов остается неизменным до тех пор, пока популяция пребывает в устойчивом состоянии. Если в популяции представлены лишь два аллеля (А и а) с частотами р и q, то частоты всех трех возможных генотипов (АА, Аа и аа) можно описать следующим уравнением:
(p+q)2
- р2
+ 2pq +q2
.
Напротив, если в популяции представлены три аллеля (А1
A2
и A3
), частоты которых есть р, q и г, частоты возможных генотипов (A1
A1
, А2
А2
, А3
А3
, Al
A2
, A1
A3
и A2
A3
) тоже можно описать следующей формулой: (p+q+r)2
= p2+q2+r2
+2pq+2pr+2qr.
Важно подчеркнуть, что рассматриваемый закон применим лишь для бесконечно больших популяций, причем его применимость демонстрируется многими примерами. Например, в случае альбинизма в популяции присутствуют генотип аа (альбиносы) наряду с нормальными генотипами АА, и Аа. Известно также, что частоты альбиносов во многих популяциях человека составляют 1 альбинос на 10 000. В соответствии с законом Д. Харди-В. Вайнберга частота гомозигот аа составляет q2
, т. е. q2
= 0,0001.
Таблица 36
Частоты генов в популяции
| Генотип
|
Количество индивидов (в % )
|
Количество аллелей каждого вида в генотипе
|
Общее число генов (в %)
|
Частота генов
|
| А/А
|
60
|
120 А/20А
|
140/200 =70
|
А =0,7
|
| А/а
|
20
|
20а
|
60/200=30
|
а = 0,3
|
| а/а
|
20/100
|
40а/200
|
100
|
0,1
|
Следовательно, q = Ö0,0001 = 0,01, а частота нормального аллеля будет равна 0,99. Таким образом, частоты нормальных генотипов (нормально пигментированных индивидуумов) будут равны: р2
= 0,992
= 0,98 для генотипа АА, тогда как для генотипа Аа 2pq = 2 х 0,99 х 0,01 = 0,02.
Как было установлено в 1926 г. С. С. Четвериковым, природные популяции насыщены рецессивными мутациями, обеспечивающими скрытый резерв (материал) для действия естественного отбора. По закону Д. Харди—В. Вайнберга в гетерозиготном состоянии присутствуют редкие аллели.
Элементарным эволюционным явлением служит мутация. В результате мутаций популяции становятся генетически гетерогенными, т. е. представляют собой смесь разных генотипов. Если условия существования популяции постоянны, ее генетический состав в среднем будет неизменным (как следует из закона Д. Харди-В. Вайнберга). Однако, если имеется давление со стороны каких-либо внешних факторов, причем в одном направлении, то тогда происходит векториальное изменение генотипического состава популяции, т. е. возникает элементарное эволюционное явление. Как показал С. С. Четвериков, популяции подобно губке насыщены рецессивными мутациями, но фенотипически являются однородными. Однако помимо генных мутаций в популяциях существуют хромосомные мутации, создавая хромосомный полиморфизм.
Мутации случайны во времени и пространстве применительно к отдельным организмам. Однако мутационные изменения организмов в популяции не являются направляющими для эволюционного процесса. Направления изменений организмов определяются естественным отбором.
Накопление в генеративных клетках мутаций и их отбор обеспечивают не только микроэволюцию, но и макроэволюцию. Таким образом, случайность в виде мутаций создает беспорядок, тогда как закономерность в виде естественного отбора создает упорядоченность.
Современные данные о транспозируемых генетических элементах и о подвижности геномов свидетельстуют о том, что многие гены могут изменять свой статус и частоту в течение одной или нескольких генераций без дифференциальной репродукции целых генотипов, т. е. неменделевским путем. Например, многократное повторение коротких последовательностей нуклеотидов приводит к образованию новых генов. Так ген коллагена цыплят содержит несколько десятков эксонов, являющихся повторами одной и той же последовательности, состоящей из 9 нуклеотидов.
Помимо мутаций эволюционным явлением служит также рекомбинация генов, но ее вклад в микроэволюцию является меньшим.
Элементарным эволюционным материалом служат любые ге-нотипически различные особи и группы особей, т. е. мутантные организмы или особи-носители мутаций, различающиеся между собой генетически. С эволюционной точки зрения важнейшей характеристикой мутантных организмов как элементарного эволюционного материала является частота появления их в природных популяциях. Наличие организмов-носителей мутаций создает различия между близкими природными группами особей (популяциями, подвидами, видами).
Элементарными факторами эволюции служат естественный отбор, мутационный процесс, популяционные волны, изоляция, дрейф генов, миграция, с действия которых начинается эволюция в популяциях. Естественный отбор является важнейшим направляющим фактором эволюции, поскольку его основная функция заключается в устранении из популяций организмов с неудачными комбинациями генов и сохранение генотипов, которые не нарушают процесса приспособительного формообразования. Действие естественного отбора проявляется в пределах популяции, но объектами приложения естественного отбора являются отдельные мутантные особи, которые являются элементарным материалом, на котором работает естественный отбор.
Специфика действия естественного отбора определяется тем, что организмы, которым благоприятствует отбор, характеризуются большей эффективностыо размножения и, следовательно, большей приспособленностью. Таким образом, приспособленность организмов является отражением их эффективности размножения. Скорость, с которой снижается частота организмов — обладателей тех или иных генотипов, называют коэффициентом отбора. Отбор действует как против, так и в пользу какого-либо аллеля (организма-носителя этого аллеля). Поэтому результатом действия естественного отбора является либо элиминация того или иного аллеля, либо появление полиморфизма, заключающегося в том, что в популяции будут присутствовать организмы-носители двух или более аллелей одного и того же гена. Отбор может действовать в нескольких случаях, в частности, против рецессивного или доминантного аллеля, в пользу или против гетерозигот.
Естественный отбор является результатом борьбы за существование, представляющейся обычно в двух формах — внутривидовой и межвидовой. Внутривидовая борьба за существование является наиболее упорной, т. к. организмы одного и того же вида конкурируют за сходные условия существования (за свет, воду, места охоты, самку и т. д.). Межвидовая борьба приобретает остроту у видов, обитающих в сходных экологических условиях.
Различают три основные формы естественного отбора: движущий, или обновляющий, стабилизирующий и дизруптивный.
Представления о движущем (обновляющем) отборе были сформулированы Ч. Дарвином. Этот отбор обеспечивает преобразование старых и выработку новых приспособлений организмов, что ведет к смене нормы реакции. По существу он заключается в адаптациогенезе и взаимодействии популяций со средой. В этом проявляется его творческая роль, а вслед за его действием и творческий характер эволюции.
Один из известных примеров движущего отбора, описанного еще Ч. Дарвином и действующего против рецессивных гомозигот, связан с индустриальным меланизмом бабочки — березовой пяденицы (Biston betularia). До середины прошлого века бабочки этого вида на Британских островах имели светло-серую окраску и были гомозиготными по рецессивному аллелю, контролирующему окраску тела. Однако после того, как в промышленных районах Англии стволы деревьев стали чернеть от копоти, выбрасываемой из заводских труб, начала появляться разновидность этих бабочек, окрашенных в темный цвет, т. е. гомозиготных доминантных и гетеро-зиготных. Бабочки темного цвета почти полностью вытеснили бабочек светлой разновидности, ибо последние оказались более доступными для питающихся ими птиц. Следовательно, отбор «подхватил» бабочек с темной окраской тела. Другим примером отбора против рецессивных гомозигот был отбор при фенилкетонурии человека в то время, когда патогенез этой болезни был неясен и когда еще не прибегали к диетотерапии. Рецессивные гомозиготные организмы (дети) без соответствующей диеты оказывались неприспособленными к существованию и погибали еще до достижения половой зрелости.
Отбор в пользу гетерозигот часто называют сверхдоминированием, поскольку гетерозиготы превосходят по выживанию гомозигот. Обычно он завершается созданием устойчивого полиморфного равновесия в популяции. Примером такого отбора является отбор при серповидноклеточной анемии, которая, как отмечено в гл. XIII, возникает в результате того, что в эритроцитах индивидов, гомозиготных по аллелю Нbs
, синтезируется аномальный гемоглобин. Нормальный гемоглобин синтезируется в эритроцитах индивидов с аллелем Нba
. Индивидуумы с генотипом Hbs
Hba
по причине невозможности приспособления к среде из-за аномального гемоглобина в большинстве случаев умирают еще до достижения половой зрелости.
Между тем в тех районах тропической Африки и Среднего Востока, где распространена малярия, вызываемая Р. falciparum, частота аллеля является довольно большой (до 30%). Причина заключается в том, что гетерозиготы Нba
Hbs
более устойчивы к тропической малярии по сравнению с нормальными гомозиготами. Следовательно, в малярийных районах гетерозиготы обладают селективными преимуществами по сравнению с гомозиготами обоих типов, для которых характерна значительно большая смертность от анемии (Hbs
Hbs
) или от малярии (Hba
Hbs
). Гетерозиготы имеют предпочтение к выживанию и размножению, поддерживая в популяциях высокую частоту аллеля Нbs
. Таким образом, в данном случае отбор благоприятствует гетерозиготам.
Очень известными современными примерами движущего отбора является отбор резистентных микроорганизмов (контрселекция на резистентность к антибиотикам), а также отбор резистентных форм насекомых, крыс и других животных на резистентность к используемым в борьбе с ними химическим соединениям.
Понятие о стабилизирующем отборе было сформулировано в 1946 г. И. И. Шмальгаузеном (1884-1963). Если исторически сложившаяся определенная приспособительная форма («адаптивная норма») сохраняет свою полезность в жизни организмов, то стабилизирующий отбор автоматически отсекает все отклонения от «нормы» и последняя как бы берется под охрану этого отбора. Стабилизирующий отбор закрепляет норму реакции, закрепляет уровень, достигнутый организмами в процессе эволюции. Стабилизирующий отбор действует обычно в условиях среды, долго остающейся однотипной. Примером стабилизирующего отбора является гибель потомства в пометах млекопитающих, размеры которых (пометов) выше среднего значения.
Дизруптивный естественный отбор (от англ. disrupt — разрывать) — это такой отбор, когда ни одна из групп генотипов в популяции не имеет преимуществ из-за одновременного изменения условий среды. В этом случае у одних организмов отбор идет по одному признаку, у других — по другому, в результате чего популяция как бы разрывается на группы особей, каждая из которых затем эволюционирует самостоятельно. Этот отбор действует против средних промежуточных форм организмов и бывает как индивидуальным, так и групповым. Наиболее част дизруптивный отбор в мире растений.
Диалектическая взаимосвязь между разными формами естественного отбора является отражением чрезвычайной противоречивости эволюции как процесса, но в любом случае творческая и направляющая роль принадлежит только движущему (дарвиновскому) отбору. Стабилизирующий и дизруптивный отборы играют второстепенную роль.
Определенное значение в эволюции имеет половой отбор, направленный на успех в размножении организмов, но этот отбор также имеет подчиненное значение.
Мутационный процесс в качестве элементарного фактора эволюции важен тем, что он является поставщиком элементарного эволюционного материала (мутантных организмов), поддерживая генетическую гетерогенность природных популяций. Однако важно заметить, что, выполняя эту роль, мутационный процесс в качестве фактора эволюции также не направляет ход эволюционных изменений. Эту функцию осуществляет естественный отбор.
Популяционные волны или волны жизни (колебания численности особей в популяциях в сторону от средней численности) являются элементарными факторами эволюции по той причине, что они тоже служат поставщиками элементарного эволюционного материала для естественного отбора (Четвериков С. С.).
Примерами их служат колебания численности вредителей полей и огородов в разные годы. Эволюционное значение волн жизни состоит в том, что они резко изменяют (повышают) концентрацию в популяциях редко встречающихся аллелей (генотипов), в результате чего под отбор попадают организмы, обладающие редкими генами, которые обычно ускользают от действия отбора.
Дрейф генов, или генетико-автоматический процесс, является следствием популяционных волн и, как отмечено выше, заключается в том, что в малых популяциях имеет место либо потеря какого-то аллеля, либо резкое повышение его концентрации. Примером дрейфа генов является утрата многих аллелей у баптистов, переселившихся около 250 лет назад из Германии в США. Но дрейф генов также не определяет направления эволюции.
Изоляция в качестве элементарного фактора обеспечивает барьеры, исключающие репродукцию. Различают географическую, экологическую и генетическую изоляцию, каждая из которых ведет в конечном итоге к репродуктивной изоляции на межвидовом уровне. При этом установлены презиготические и постзиготические механизмы, предотвращающие скрещивания.
К презиготическим изолирующим механизмам относят географическую и экологическую изоляции, которые исключает встречи потенциальных брачных партнеров. Известна также поведенческая изоляция, когда потенциальные брачные партнеры встречаются, но из-за недостаточного полового влечения эти встречи заканчиваются отрицательно, и сезонная изоляция, когда партнеры встречаются, но не спариваются, ибо спаривание у них в нормальных условиях обычно происходит в разное время. Наконец, известна гаметическая изоляция, при которой после совокупления животных не происходит осеменения, т. к. сперматозоиды утрачивают жизнеспособность в женских половых путях.
Постзиготические изолирующие механизмы также различны. Например, после встречи партнеров яйцеклетки могут оплодотвориться, но зиготы погибают. Если зиготы развиваются, то гибриды все же не достигают половой зрелости.
Далее могут формироваться гибриды, но их жизнеспособность оказывается невысокой, либо они могут быть стерильными (не способными продуцировать гаметы, обладающие нормальными свойствами).
Важнейшей особенностью изоляции как элементарного фактора эволюции является ее длительность. Нарушая скрещивания и закрепляя различия в генотипах, изоляция как бы расчленяет популяцию на генотипически различные группы организмов, полностью исключая среди них обмен генами, усиливая дивергенцию признаков. Изоляция является, по существу, усилителем начальных стадий генотипической дифференциации, развертывающейся в популяциях. Она подставляет разделенные части популяций под действие естественного отбора. Однако изоляция тоже не направляет эволюцию. Это делает только естественный отбор.
Миграция аллелей (мутантных организмов) ведет к изменению в популяциях как частот аллелей, так и генотипов. Например, негритянское население США унаследовало около 30% генов от белых предков. Миграция способствует объединению генных пулов популяций.
Видообразование, т. е. появление нового вида — это центральный и важнейший завершающий этап эволюции. Вид — это совокупность особей, которые имеют сходное строение и характеризуются сходными функциями, в природе скрещиваются только между собой, приспособлены к жизни в определенных условиях, имеют характерный ареал распространения и общее происхождение. Будучи реальной биологической категорией, виды состоят из популяций, причем особям, образующим вид, присуща сформировавшаяся в ходе эволюции единая генетическая программа.
Для отличия одних видов от других используют ряд критериев. Одним из важнейших критериев вида является морфологический критерий, под которым понимают строение организмов, принадлежащих к тому или иному виду. В случае отдаленных видов морфологический критерий весьма точен. Однако в случае близких видов этот критерий не является совершенным.
Генетический критерий вида заключается в количестве хромосом, присущих данному виду, а также в характере последовательности азотистых оснований в ДНК и аминокислотных остатков в полипептидах (белках). Этот критерий характеризуется чрезвычайной чувствительностью и позволяет различать даже очень близкие виды.
Эколого-географический критерий определяется ареалом и экологической нишей организмов, входящих в данный вид. Другими словами, каждому виду присущ собственный ареал и собственная среда обитания.
Наконец, критерием вида является и репродуктивная изоляция, когда организмы одного вида не способны к скрещиваниям с организмами другого вида, либо они обладают этой способностью, но их потомство все же не является плодовитым. Поскольку каждый из названных критериев имеет ограничения, для определения видовой принадлежности организмов эти критерии используют в совокупности.
Для видов характерны популяционные различия. Вид может состоять как из одной, так и нескольких популяций. Кроме того, вид может занимать как небольшую территорию, так и гигантские пространства.
Эволюция представляет собой процесс в двух измерениях. Один из них является анагенезом, под которым понимают эволюцию организмов в одном направлении, тогда как другой является кладоге-незом, представляющим собой эволюцию в разных направлениях. Примером анагенеза является, видимо, эволюция Н. erectus в Н. habilis, тогда как для кладогенеза характерно расщепление одной эволюционной линии на две или больше. Поэтому главное содержание кладогенетической эволюции заключается в видообразовании путем разделения одного вида на два и более.
Образование новых видов происходит постепенно (географическое видообразование) и измеряется длительными отрезками времени либо мгновенно (квантами) на протяжении короткого периода времени.
Постепенное (географическое) или, как его называют аллопат-рическое (от лат. allos — разный, patria — родина) видообразование начинается с дивергенции признаков и осуществляется в два этапа. На первом этапе такого видообразования происходит географическое разделение популяций реками, морями, горами, пустынями, различными особенностями ландшафта (рис. 169). Популяции приспосабливаются к существующим условиям благодаря естественному отбору. Между разными популяциями прерывается обмен генами. Это углубляет генетическую дифференциацию, в которой некоторую роль еще может играть дрейф генов, особенно, когда популяции имеют небольшие размеры, вплоть до нескольких особей. Возникновение генетических различий сопровождается появлением репродуктивных изолирующих механизмов. В результате действия последних происходит накопление в каждой из популяций своего набора генов и достижение популяциями того состояния, которое уже может быть определено в качестве подвида.
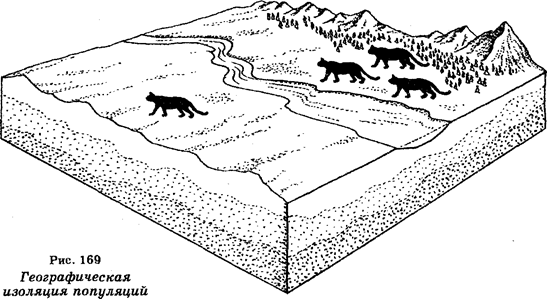
На втором этапе постепенного видообразования происходит завершение репродуктивной изоляции. При этом генетические различия достигают такого уровня, когда гибриды еще будут появляться, но будут полностью нежизнеспособными. Каждый подвид продолжает эволюционировать независимо. Наконец наступает превращение подвидов в настоящие виды. Аллопатрическое видообразование происходит на разных территориях.
Помимо аллопатрического видообразования различают симпатрическое (от лат. sym — одинаковый, patria — родина) видообразование. Симпатрическое видообразование происходит на одной территории. Примером такого видообразования является возникновение эндемичных видов млекопитающих и рыб в озере Байкал.
Возникновение нового вида завершается лишь тогда, когда организмы в течение определенного периода изоляции приобретают признаки, которые способствуют или обеспечивают репродуктивную изоляцию даже после разрушения изолирующих преград. Таким образом, в генетическом смысле сущность видообразования заключается в возникновении двух хорошо интегрированных генных комплексов из одного родительского генного комплекса.
Постепенное видообразование применительно к организмам разных систематических групп происходит с разной скоростью. Например, птицы эволюционировали не очень быстро, тогда как гоминиды эволюционировали очень быстро.
Новые виды образуются не из самых высокоразвитых видов, а из более простых. Однако эволюция не всегда идет от простого к сложному. Напротив, имеет место и эволюционный «регресс». Например, большинство паразитов человека и животных произошло от свободноживущих организмов, для которых характерна более сложная организация. В частности, бескрылые насекомые произошли от крылатых, а киты — от четвероногих млекопитающих.
Мгновенное (квантовое) или скачкообразное (сальтационное) видообразование наблюдается в результате мутации и сокращения сроков действия репродуктивных изолирующих механизмов. Примером такого видообразования является возникновение в течение одного поколения полиплоидов растений, которые обладают характеристиками новых видов. Видообразование через полиплоидию известно и у животных, хотя у последних этот механизм видообразования очень редок. Полиплоиды считают самостоятельными видами не только по причине резкого отличия их от исходных видов, но и по той причине, что они репродуктивно изолированы от исходных видов.
Важнейшей особенностью эволюции является склонность организмов эволюционировать в определенном направлении, т. е. часто эволюции присуща определенная заданность или предначертан-ность. Эта особенность была выявлена еще в XIX в. выдающимся русским эволюционистом В. О. Ковалевским (1842-1883) на примере эволюции конечностей копытных животных. В. О. Кавалевс-кий установил, что эволюция конечностей у предков лошадей шла в заданном (преднамеренном) направлении от трехпалых конечностей предков к однопалым конечностям, которые характерны для современных лошадей, поскольку их копыта представляют одиночные пальцы. Другие ученые отметили эволюционную предна-чертанность в случае эволюции хобота и бивней у предков современных слонов.
Эволюция видов всегда необратима, и это есть одна из важнейших закономерностей эволюции. Необратимость означает, что исчезнувшие виды никогда не возвращаются, т. е. вновь возникающие виды никогда не бывают сходными с видами, существовавшими прежде. Это положение иллюстрируется следующим примером. Известно, что в ходе эволюции вымерло 200 000 разных видов животных. Но среди появившихся затем и существующих сейчас видов нет ни одного аналогичного какому-либо вымершему виду. С другой стороны, появление нового вида есть процесс однократный, т. е. одинаковые виды никогда не возникают дважды. Следовательно, любой вид уникален и неповторим.
Существенное место в теории эволюции занимает вопрос об устойчивости существующих видов. Говоря об устойчивости видов, следует признать, что она является исторической реальностью. В видах удерживаются все те приобретения, которые достигнуты в процессе эволюции.
В пользу признания устойчивости видов можно привести два довода. Первый довод сводится к тому, что виды (с учетом всех известных критериев вида) существуют очень длительное время, измеряемое миллионами лет, без видимых изменений. Что касается второго довода, то он сводится к признанию факта, что если бы виды не обладали устойчивостью, то закрепление результатов эволюции просто было бы невозможным. Следовательно, невозможной была бы и сама эволюция.
Однако устойчивость видов не является абсолютной. В процессе жизни виды подвергаются изменчивости. В природных условиях генетическая изменчивость очень широко распространена (табл. 37). Особенно это заметно на примерах индивидуальной изменчивости. Хотя порой и трудно различить в морфологической изменчивости удельный вклад генотипической и фенотипической изменчивости, тем не менее у растений можно обнаружить изменения, например, по цвету семян и цветков, по характеру роста и другим признакам.
У животных морфологическая изменчивость затрагивает, например, окраску и форму раковин у моллюсков, форму крыльев бабочек и других членистоногих, окраску оперения птиц, шерсти млекопитающих и т. д. Еще большая изменчивость отмечается у человека, причем по многим признакам (черты лица, цвет и форма волос, пигментация кожи и др.).
Изменчивость создает возможности для гибкого приспособления организмов к меняющимся условиям среды. Можно сказать, что посредством изменчивости виды могут «маневрировать», не теряя основных характеристик, в процессе своей жизни.
Таблица 37
Генотипическая изменчивость в природных популяциях
животных и растении
| Организм
|
Число
видов
|
Среднее число локусов у вида
|
Средняя по-лииорфиость
|
Средняя гетерозиготность
|
| Плодовые мушки
|
28
|
24
|
0,529
|
0,150
|
| Осы
|
6
|
15
|
0,243
|
0,062
|
| Другие насекомые
|
4
|
18
|
0,531
|
0,151
|
| Морские беспозвоночные
|
14
|
23
|
0,439
|
0,124
|
| Наземные улитки
|
5
|
18
|
0,437
|
0,150
|
| Рыбы
|
14
|
21
|
0,306
|
0,078
|
| Земноводные
|
11
|
22
|
0,336
|
0,082
|
| Пресмыкающиеся
|
9
|
21
|
0,231
|
0,047
|
| Птицы
|
4
|
19
|
0,145
|
0,042
|
| Млекопитающие
|
30
|
28
|
0,206
|
0,051
|
| Самоопыляющиеся
|
12
|
15
|
0,231
|
0,033
|
| Перекрестноопыляющиеся
|
5
|
17
|
0,344
|
0,078
|
| Среднее по группам
|
| Беспозвоночные
|
57
|
22
|
0,469
|
0,134
|
| Позвоночные
|
68
|
24
|
0,247
|
0,060
|
| Растения
|
17
|
16
|
0,264
|
0,046
|
Как отмечено в начале этого раздела, эволюция протекает на всех уровнях организации живой материи, причем каждый эволюционный уровень является результатом низшего и служит эволюционным материалом для высшего уровня. Ресурсы естественного отбора также имеются на всех уровнях. Организмы каждого нового эволюционного уровня обладают потенциальной возможностью для более быстрого развития по сравнению с предыдущим. Например, млекопитающие имеют больше возможностей для дальнейшей эволюции, чем одноклеточные животные.
Обобщение результатов молекулярно-генетических исследований эволюции привело японского ученого М. Кимуру (1968) и других исследователей к формулированию гипотезы, по которой большинство изменений, наблюдаемых в популяциях, обусловлено мутациями, которые не оказывают влияния на приспособление организмов к окружающей среде и которые заключаются в случайных заменах азотистых оснований в молекулах ДНК. В соответствии с гипотезой нейтральности молекулярной эволюции скорости замен оснований в ДНК (аминокислот в белках) могут быть постоянными, т. к. преобладающее число таких замен селективно нейтрально.
При одинаковой способности альтернативных аллелей придавать организмам одинаковую приспособленность к условиям среды, изменение частот встречаемости этих аллелей в поколениях организмов может происходить лишь в результате дрейфа генов. Следовательно, скорость замены аллелей в популяциях и скорость происходящих отсюда замен аминокислот в белках будет постоянной.
По гипотезе нейтральности молекулярной эволюции большинство мутаций любых генов вредно для организмов и мутанты элиминируются с помощью естественного отбора. Однако могут возникнуть мутации, которые не меняют ни морфологических, ни функциональных признаков, в результате чего организмы-носители таких мутаций не подвержены естественному отбору.
Следовательно, эволюция на молекулярном уровне в основном заключается в замене одних нейтральных генных аллелей другими, но такими же нейтральными аллелями.
Сторонники гипотезы нейтральности эволюции на молекулярном уровне считают, что представления о нейтральности эволюции на молекулярном уровне не противоречат дарвинизму.
Сторонники гипотезы нейтральности эволюции также считают, что эволюция ДНК и белков представляет собой молекулярные «часы» эволюции.
Многие эволюционисты придают большое значение генам-регуляторам. Предполагают, что мутации регуляторных генов очень важны для адаптивной эволюции, однако данные в пользу этого предположения еще незначительны.
С самого начала теория эволюции оказалась ареной острой борьбы идей, в которой дарвинизму с самого начала стали противопоставлять неоламаркизм или подменять его социал-дарвинизмом.
В неоламаркизме различают психоламаркизм и механоламар-кизм. Психоламаркисты в качестве главного фактора эволюции называли психику. По их мысли эволюцию определяет выработка привычек к новым действиям, постепенное превращение сознательных актов в автоматизм. Психоламаркисты отрицали естественный отбор и считали, что наследственность — это память, которая запечатлевает все воздействия внешней среды, воспроизводящиеся в последующих поколениях в адекватной форме. Механоламаркис-ты полагали, что в основе эволюции лежит переход материи от недифференцированного состояния в дифференцированное. Находясь в состоянии взаимодействия, организм и среда стремятся к равновесию, которое часто нарушается под воздействием факторов среды. Равновесие восстанавливается прямым уравновешиванием в результате непосредственного приспособления и косвенным уравновешиванием через естественный отбор. Все неоламаркисты считали органическую целесообразность свойством организмов, данным изначально.
В XIX в. возник и перешел в XX в. социал-дарвинизм, основателем которого был Э. Геккель, много сделавший для защиты и пропаганды дарвинизма. Однако в трактовке эволюции Э. Геккель и другие социал-дарвинисты отступали к Ламарку, признавая на равных условиях отбор и прямое приспособление на основе унаследования полезных признаков. В то же время они стремились использовать биологические закономерности для объяснения закономерностей развития общества. Главным двигателем общественного развития они считали борьбу за существование и естественный отбор. На основе естественного отбора они пытались обосновать возникновение классов и классового общества, объяснить низкий жизненный уровень народов колоний (теперь развивающихся стран) биологическими причинами. В новейшее время социал-дарвинизм слился с евгеникой, в частности, с той ее частью, в рамках которой предлагается перестроить геном человека. Однако большинство биологов отрицает социал-дарвинизм, так же как и евгенические программы перестройки генома человека.
С самого начала своего возникновения дарвинизм встретил неприязнь католической церкви, которая на длительный период времени превратилась в отрицание этой церковью эволюции. Больше того, эволюция отрицалась креационизмом, а сейчас отрицается «научным» креационизмом. Однако под напором фактических данных католическая церковь в последнее время вынуждена была изменить свое отношение к теории эволюции. В октябре 1996 г. в обращении к Папской академии наук папа римский Иоанн Павел II заявил, что сейчас в пользу теории эволюции так много данных, что их нельзя больше- игнорировать. В частности, папа отметил, что сейчас экспериментальные науки описывают и измеряют с возра-стающейся прецизионностью многочисленные проявления жизни, объясняя их в эволюционной шкале времени, и что теория эволюции есть «...более, чем только теория». Этим заявлением завершается длительный спор католической церкви со сторонниками эволюции в пользу последней. Поскольку «научный» креационизм имел поддержку со стороны католической церкви, то заявление ее главы значительно ослабило положение «научных» креационистов.
Признание эволюции есть одна из выдающихся основ в формировании научного мировоззрения людей, в формировании представлений о научной картине мира.
1. В чем заключается эволюционизм?
2. На чем основываются современные доводы в пользу эволюции?
3. Почему современную теорию эволюции называют синтетической и в чем состоит ее отличие от классического дарвинизма?
4. В чем заключается сущность современного понимания происхождения жизни? Почему современная теория происхождения жизни является материалистической?
5. Что является главным в учении Ч. Дарвина?
6. Какова роль случайности в эволюции?
7. Назовите главные направления в эволюции, приведите примеры.
8. Дайте определение макро- и микроэволюции. Каково соотношение между макро- и микроэволюцией?
9. Назовите элементарные единицы, явления, материал и факторы эволюции.
10. Дайте определение популяции. Почему элементарной единицей эволюции является популяция?
11. Что представляет собой естественный отбор? Какие формы естественного отбора вы знаете? Какова роль движущего отбора?
12. Что вы знаете об отборе в пользу гетерозигот? Расскажите о серпо-видноклеточной анемии человека и объясните, почему в малярийных районах гетерозиготы Hba
Hbs
обладают селективными преимуществами?
13. Дайте определение вида.
14. Дайте определения анагенеза и кладогенеза. Есть ли разница между этими процессами?
15. Что вы знаете о скорости эволюции? Как вы понимаете механизм видообразования?
16. Обратима ли эволюция? Почему виды устойчивы, какое это имеет значение для эволюции?
17. Происходит ли образование новых видов в современную эпоху?
18. Можно ли считать новыми видами организмы, создаваемые генной и клеточной инженерией?
19. Что вы знаете о гипотезе нейтральности молекулярной эволюции?
20. Какие вы знаете антидарвиновские концепции эволюции?
21. В чем заключается общенаучное значение теории эволюции?
22. Какова роль теории эволюции в биологии и в практической деятельности человека, связанной с растениями и животными?
23. Различия в последовательностях азотистых оснований ДНК человека и зеленой мартышки равны 9,6%, человека и обезьяны-капуцина — 16,8%, а зеленой мартышки и капуцина — 16,6%. Можете ли вы реконструировать филогению этих трех видов и указать вероятный процент замен азотистых оснований в ходе эволюции?
Аиала Ф. Введение в популяционную и эволюционную генетику. М.: Мир. 1984. 227 стр.
Дубинин Н. П. Генетика. Кишинев: Штиинца. 1985. 533 стр.
Майр Э. Популяции, виды и эволюция. М.: Мир. 1974. 460 стр.
Опарин А. И. Жизнь, ее природа, происхождение и развитие. М.: Наука. 1968.
Руттен М. Происхождение жизни. М.: Мир. 1973. 411 стр.
Северцов А. Н. Главные направления эволюционного процесса. М.: Виомедгиз. 1934. 150 стр.
Тимофеев-Ресовский Н. В., Воронцов Н. Н., Яблоков А. В. Краткий очерк теории эволюции. М.: Наука. 1969. 407 стр.
Яблоков А. В.. Юсуфов А. Г. Эволюционное учение. М.: Высшая школа. 1989. 335 стр.
Czifto G. Without Miracles Universal Selection Theory and the Second Darwinian Revolution. NIT Press. 1957. 385 pp.
Rosemweig M. L. Species Diversity in Space and Time. Cambridge University Press. 1995. 436 pp.
Эволюция человека, или антропогенез (от греч. anthropos — человек, genesis — развитие) — это исторический процесс эволюционного становления человека. Науку, изучающую происхождение человека, называют антропологией, возникновение которой датируют рубежом XVIII-XIX вв.
Эволюция человека качественно отличается от эволюции организмов других видов, ибо в ней действовали не только биологические, но и социальные факторы. Сложность проблем антропогенеза углубляется тем, что человек сам по себе двулик. Одним лицом он обращен к животному миру, из которого вышел и с которым остается связанным анатомически и физиологически, а вторым — в мир создаваемых коллективным трудом научных и технических достижений, культуры и т. д. Человек, с одной стороны, является биологическим существом, с другой — социальным.
Еще в начале XX в. И. И. Мечников отмечал, что «...теория происхождения человека может быть причислена к числу наиболее прочных научных теорий». Будучи связанным тесными узами Кровного родства с животным миром, человек имеет свою биологическую предысторию. Однако человек имеет качественные отличия от животных. Эти отличия возникли под влиянием социальных факторов в процессе исторического развития человеческого общества.
Наследственная конституция человека сложилась в результате постепенного и длительного процесса эволюции. У непосредственных предшественников человека в процессе эволюции на базе генетической программы непрерывно возникали противоречия между их морфофизиологической организацией и формирующимися элементарными способами «инстинктивно-трудовой» деятельности. Разрешение этого противоречия путем естественного отбора вначале привело к изменениям передних конечностей, затем к развитию коры больших полушарий головного мозга, и наконец, к появлению сознания. Можно сказать, что это был первый, но решающий акт в завершении специализации генов на гены структуры и гены-регуляторы, причем сознание обеспечило не только становление, но и дальнейшее развитие человека.
В последующем темпы биологического развития человека стали снижаться. Поскольку появление сознания обеспечивало новые формы и возможности приспособления к среде, это вело к уклонениям от действия естественного отбора, в результате чего биологическое развитие уступило место социальному развитию и совершенствованию. Как заметил в свое время К. А. Тимирязев, биологическая эволюция человека осталась за порогом его истории. Таким образом, современный человек является продуктом его генов и среды. Выйдя из природы, человек остался тесно связанным с ней, со всем живым миром.
В научном плане учение об антропогенезе тесно связано с учением о расогенезе. Вопреки научному пониманию расогенеза извращение научных представлений о происхождении рас ведет к расизму, в основе которого лежит реакционная идея о высших и низших расах.
Удивительно, но история взглядов на антропогенез гораздо древнее самой антропологии, ибо еще ученые античного мира обсуждали вопросы о месте человека в природе и о появлении человека. Например, Анаксимандр (610-546 гг. до н. э.) и Аристотель (384-322 гг. до н. э.), определяя место человека в природе, признавали его предками животных. Разделяя животных на «кровяных» и бескровных, Аристотель отнес человека к группе «кровяных» животных, а между человеком и животными в «кровяной» группе поместил обезьян. То, что человек близок к животным, признавал К. Гален (около 130-200), который формулировал заключения об анатомия человека на основе результатов вскрытия низших обезьян.
К. Линней (1707-1778) по сравнению со своими предшественниками пошел намного дальше, выделив отряд приматов, отнеся к нему полуобезьян, обезьян и род людей с одним видом — Человек разумный (Homo sapiens) и подчеркивая сходство человека и обезьян. Однако К. Линнею не удалось правильно очертить границы этого отряда, ибо он относил к нему одновременно то ленивца, то летучих мышей. Не все современники К. Линнея признавали его систему, в частности принадлежность человека к отряду приматов. Были предложены также другие варианты системы, в которых ранг человека был значительно завышен, поскольку за человеком признавалось отдельное царство природы. Этим, по существу, отрывали человека от животных.
Идею родства человека и обезьян поддерживали в XVIII в. И. Кант (1724-1804), Д. Дидро (1713-1784), Ж. Ламеттри (1709-1751), А. А. Каверзнев (1748-?), А. Н. Радищев (1749-1802), Ж. Ламарк (1744-1829). Больше того, Ж. Ламеттри даже считал, что таксономическое объединение человека и животных не унижает человека, что способность ощущения и мышления есть у всех животных и зависит от степени их организации.
Вопреки правильному решению вопроса о родстве человека с животными, вопрос о том, как произошел человек, в трудах древних философов остался открытым. Считают, что первая гипотеза антропогенеза была сформулирована, вероятно, Ж.-Б. Ламарком. Полагая, что человек имел обезьяноподобных предков, Ж.-Б. Ламарк впервые назвал очередность эволюционных достижений в превращении обезьяноподобного предка в человека, причем важнейшее значение он придал переходу древесных четвероногих к двуногой локомоции и к жизни на земле. Ламарк описал изменения скелета и мускулатуры предков человека в связи с переходом к прямохождению. Но, переоценив роль среды, он, как и в случае других организмов, все же неправильно представлял движущие силы эволюции человека.
Н. Г. Чернышевский (1828-1889) считал, что антропогенез является одним из проявлений исторического развития природы. А. Уоллес (1823-1913) высказал предположение, что в эволюции человека большое значение имели формы, передвигавшиеся на двух конечностях, и что вслед за прямохождением происходило увеличение мозга. Он предположил, что история возникновения человека была очень длительной во времени. Несомненно, что эти и другие сходные высказывания были лишь значительным шагом вперед в понимании вопроса о появлении человека, однако они не были исчерпывающими и не привели к образованию научной теории происхождения человека. Подлинно научная теория происхождения человека стала формулироваться тогда, когда появилось эволюционное учение Ч. Дарвина, ставшее базой для этой теории.
Отмечая, как рассматривали происхождение человека в прошлом, нельзя не отметить, что существовали и продолжают существовать и другие взгляды. Противники материалистической философии всегда приписывают человеку сверхъестественное происхождение. Отрывая сознание от тела и признавая его первичным, они признавали и признают независимость в создании тела и души, отделимость души от тела после смерти. Взгляды сторонников сверъхестественного происхождения человека давно слились с библейской концепцией, в рамках которой утверждается внезапное сотворение человека на седьмой день Творения, состоявшегося 10 000 лет назад. В наше время эту концепцию защищают сторонники «научного» креационизма.
В противоположность значительному распространению «научного» креационизма в последней четверти XX в. под давлением научных фактов Папа Римский Иоанн Павел II в 1986 г. вынужден был признать происхождение тела человека в рамках теории эволюции, но не души человека. В октябре 1996 г. он повторил свои высказывания об эволюции человека. Учитывая авторитет главы римско-католической церкви, можно заключить, что его высказывания означают конец антиэволюционным взглядам на природу человека.
В основе современных представлений о происхождении человека лежит концепция, в соответствии с которой человек вышел из мира животных, причем первые научные доказательства в пользу этой концепции были представлены Ч. Дарвином в его труде «Происхождение человека и половой отбор» (1871). В последующем по мере развития анатомии и эмбриологии эти доказательства пополнялись новыми данными, которые указывали на анатомическое сходство и сходство эмбрионального развития человека и животных. В настоящее время в пользу концепции животного происхождения человека служит ряд доводов, наиболее важными из которых являются следующие:
1.
Для человека характерны все черты, присущие типу Хордовые, в частности:
а) билатеральная (двусторонняя) симметрия в строении тела,
б) наличие в зародышевом развитии хорды и жаберных щелей в полости глотки,
в) расположение нервной системы в форме дорсальной трубки.
2.
Для человека характерны все черты подтипа Позвоночные (Черепные), а именно:
а) наличие внутреннего осевого скелета, основой которого является развитый позвоночный столб, с передним концом которого сочленена черепная коробка, а также наличие двух пар конечностей,
б) центральная нервная система имеет вид трубки, переходящей в головной мозг, который состоит из 5 отделов,
в) сердце развивается на брюшной стороне тела.
3.
Для человека характерны все черты класса Млекопитающие, а именно:
а) живорождение и вскармливание молоком, наличие молочных желез, волосяного покрова,
б) теплокровность и обилие потовых желез для обеспечения терморегуляции,
в) разделение полости тела диафрагмой на брюшной и грудной отделы,
г) наличие 4-камерного сердца, левой дуги аорты, отсутствие в зрелых эритроцитах ядер,
д) дыхательная система представлена легкими, трахеей, бронхами, альвеолами,
е) наличие всех костей, характерных для млекопитающих. У человека нет ни одной лишней кости, которая бы отсутствовала у млекопитающих. В скелете имеется 7 шейных позвонков, 2 мыщелка затылочной кости и 3 слуховых косточки, характерные для млекопитающих,
ж) наличие молочных и постоянных зубов трех групп,
з) проявление атавистических признаков, наличие рудиментарных органов (мышцы, приводящие в движение ушную раковину, отросток слепой кишки, третье веко глаза и другие).
4.
Для человека характерны все черты подкласса Плацентарные, а именно:
а) наличие плаценты,
б) вынашивание плода внутри тела матери и питание его через плаценту.
5.
Для человека характерны все черты отряда Приматы, а именно:
а) наличие одной пары грудных молочных желез,
б) концы пальцев (концевые фаланги) имеют ногти, а ладони покрыты узорами,
в) противопоставление большого пальца передней конечности остальным, что обеспечивает брахиацию (использование конечностей для хватательных движений),
г) наличие менструального периода и беременности длительностью в 9 месяцев,
д) антигены системы АВО человека и человекообразных обезьян сходны. Группы крови А (II) и В (III) обнаружены у всех человекообразных обезьян, группа О (I) лишь у шимпанзе. По существу, кровь шимпанзе и гориллы можно переливать человеку,
е) наличие сходства в количестве и строении хромосом. Для человека характерны 23 пары хромосом, для человекообразных обезьян — 24 пары, из которых 13 пар по своему строению одинаковы в обоих случаях,
ж) наличие значительной гомологии ДНК человека с ДНК обезьян. Например, гомология ДНК человека и шимпанзе составляет 91—92%, человека и гиббона — 76%, а человека и макаки-резус — всего лишь 66%,
з) одинаковая чувствительность человека и человекообразных обезьян к возбудителям одних и тех же болезней и сходство клинического проявления последних, и) сходство между генами, контролирующими синтез белков у приматов. Это сходство устанавливают на основе структуры белков, контролируемых этими генами. Например, в клетках всех организмов, дыхание которых связано с использованием кислорода, синтезируется ци-тохром С (белок-переносчик электронов в митохондри-ях). Этот белок представляет собой сложную молекулу, состоящую примерно из 100 аминокислот. Как показывает табл. 38, степень сходства структуры цитохрома С коррелирует с анатомическим, физиологическим и филогенетическим сходством приматов. Человек отличается от других приматов всего лишь одной аминокислотной заменой.
В случае а-гемоглобина человека и гориллы имеется лишь одно различие в последовательности аминокислот, тогда как человека и лошади — 18 различий, человека и карпа — 71 различие. Между человеком и шимпанзе имеется исключительное сходство по строению белков (различие по 44 функциональным белкам не превышает 1%).
Место человека в системе животного мира определяется тем, что он относится к типу Хордовые, подтипу Позвоночные, классу Млекопитающие, подклассу Плацентарные, отряду Приматы, семейству Гоминиды, роду Homo. Как считал Ч. Дарвин «...Животные — наши братья по боли, болезням, смерти, страданию и голоду, наши рабы в самой тяжелой работе, наши товарищи в наших удовольствиях — все они ведут, может быть, свое происхождение от одного общего с нами предка — нас всех можно было бы слить вместе».
Таблица 38
Сравнительные данные о структуре цитохрома С
у разных организмов
| Сравниваемые виды организмов
|
Количество аминокислотных различий
|
| Человек — обезьяна
|
1
|
| Человек — лошадь
|
12
|
| Человек — собака
|
10
|
| Лошадь — крупный рогатый скот
|
3
|
| Млекопитающие — куры
|
10-15
|
| Млекопитающие — тунец
|
17-21
|
Однако человек имеет существенные отличия от животных, на что также обращали внимание еще древние, например Анаксагор (500—428 гг. до н. э.) и Сократ (469—399 гг. до н. э.) считали, что специфическим признаком человека является наличие руки, которая выделила человека из всего мира. Называя человека «животным общественным», Аристотель ссылался на такие отличия, как двуногое хождение, больший по величине мозг, способность к речи и мышлению. Позднее К. Линней в качестве специфических отличий человека от обезьян называл речевую способность, а также способность накапливать и передавать в поколениях опыт, письменность, печать. По этой причине он и называл человека разумным. А. Н. Радищев обращал внимание на такие отличительные свойства человека, как способность к прямохождению, наличие рук, речи, разума.
Современные представления относительно отличий человека от животных основываются, прежде всего, на данных о различиях в развитии мозга и на способности человека к абстрактному мышлению. Средняя масса мозга человека составляет 1350-1500 г, тогда как гориллы и шимпанзе — всего лишь 460 г. Масса мозга человека составляет в средним около 1/40 общей массы тела, тогда как у обезьян — 1/60 — 1/200. Поверхность мозга человека составляет около 1200 см2
, шимпанзе — 400 см2
.
Среди других отличительных признаков человека следует назвать особенности челюстей, а также строение и расположение зубов, которые являются иными по сравнению с зубами животных. Важнейшим отличием является дифференцировка верхних и нижних конечностей, характерные изгибы позвоночника, широкий таз. Только человек способен к балансированию на двух конечностях (к двуногой локомоции). У человека довольно мощными являются кости, причем самой мощной является бедренная кость, выдерживающая нагрузку до 1650 кг. Исключительное развитие получила дифференциация кисти, обеспечивающая хватательные движения, значительные размеры приобрел первый палец. Из-за расположения глаз в передней части головы человек обладает бинокулярным зрением, которое позволяет ему различать (видеть) предметы в трех измерениях.
Однако важнейшей отличительной особенностью человека является то, что рассудочная деятельность, имеющаяся у многих видов животных, у человека достигает наивысшего развития, ибо он обладает сознанием, способностью к абстрактному мышлению, общению с помощью речи (2-й сигнальной системы) и абстрактных символов (письма), а также к передаче и восприятию информации.
Благодаря высокому уровню интеллектуального развития человек создал культуру, стал производить орудия труда для воздействия на природу, развил технологию, создал города, литературу, музыку и т. д. Являясь социальным существом, человек способен думать о прошлом, анализировать прошлое и планировать будущее. Этими свойствами животные не обладают.
Филогенез человека представляют, исходя из дарвиновского положения, в соответствии с которым предками человека были обезьяноподобные существа, а развитие человека и человекообразных обезьян представляло собой не последовательные степени, а параллельные ветви эволюции, характеризующиеся глубоким расхождением с эволюционной точкой зрения. Ч. Дарвин называл современных человекообразных обезьян двоюродными братьями и сестрами человека. Однако это сравнение не совсем верно, ибо в наше время гоминид и понгид считают дальними родственниками, у которых вымерли промежуточные ветви, т. е. человек произошел от каких-то животных, одна ветвь которых ведет к современным высшим обезьянам, вторая — к человеку.
Чтобы правильно понять самые ранние этапы эволюции человека необходимо начать с эволюции отряда приматов, в состав которого входят семейства понгид и гоминид. Находки палеонтологов позволяют проследить развитие животных, приведшее к появлению первых приматов, начиная с мезозойской эры, в частности, с триаса (230 млн лет назад), когда из тогдашних видов пресмыкающихся возникли первые млекопитающие, получившие название Protoinaectivara. Предполагают, что на основе именно этих примитивных млекопитающих в дальнейшем возникли более совершенные млекопитающие — насекомоядные, ставшие предками тупай —современных низших приматов (рис. 170), от которых через разные промежуточные формы одна из эволюционных ветвей повела к приматам, впервые появившимся в меле (рис. 171).
Отряд приматов классифицируют на подотряды полуобезьян (Prosimii) и обезъян (Antropoidea). Наиболее ранние приматы из подотряда Antropoidea были многочисленными в начале палеоцена (65 млн лет назад). В олигоцене (40 млн лет назад) приматы этого подотряда в результате эволюции дали начало 3 большим группам (надсемействам) живых существ, а именно:
 1. Надсемейство Geboidea. Представители этого надсемейства в ходе эволюции в свою очередь дали начало обезьянам Нового света (широконосым), которые эволюционировали в игрунков и капуцинов. Это надсемейство представляет собой приматов, не имеющих значения в эволюции человека. 1. Надсемейство Geboidea. Представители этого надсемейства в ходе эволюции в свою очередь дали начало обезьянам Нового света (широконосым), которые эволюционировали в игрунков и капуцинов. Это надсемейство представляет собой приматов, не имеющих значения в эволюции человека.
2. Надсемейство Cercopithecoidea. Представители этого надсемейства дали начало обезьянам Старого света (узко- носым), которые эволюционировали в макак и павианов. Это надсемейство тоже не имеет значения для эволюции человека.
3. Надсемейство Hominoidea (гоминоиды). Представители этого надсемейства появились в верхнем плиоцене и дали начало человекообразным обезьянам и человеку. Эволюция гоминоидов происходила независимо от других приматов. В пределах этого надсемейства выделяют семейства Pliopithecidae, представители которого полностью вымерли, Oreopithecidae, представители которого тоже полностью вымерли, Pongidae (человекообразные обезьяны), представители которого частично вымерли и Hominidae (люди), представители которого тоже частично вымерли. Понгид делят на подсемейства гиббонов (Hilolatenae) и настоящих человекообразных обезьян (Pongidae), к которым относятся орангутаны, гориллы и шимпанзе.
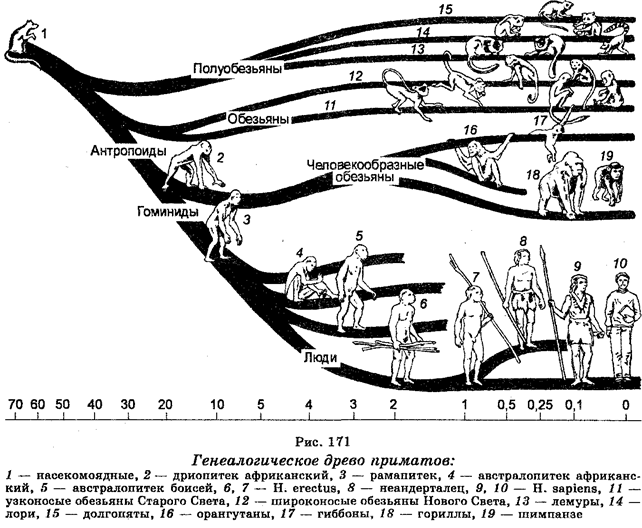
Начало эволюции семейства понгид (Pongidae) и гоминид (Hominidae) датируется временем, когда одна из ветвей эволюции повела к дриопитековым (древесным человекообразным обезьянам), сходным по костям с гориллами, шимпанзе и человеком. Наибольшая концентрация ископаемых останков дриопитеков обнаружена в Восточной Африке, однако их формы были найдены и в Европе и Азии, на территориях ФРГ, Франции, России, КНР, Турции. Возраст дриопитеков составляет около 25 млн. лет. Считают, что дри-опитековые представляют собой узел родословного древа приматов от которого берут начало 3 линии развития — к орангутанам, человекообразным обезьянам и человеку.
У истоков расхождения путей эволюции понгид и гоминид стоит один из дриопитеков, названный в честь индийского бога Рамы рамапитеком, челюсть которого была найдена Д. Льюисом в 1934 г. в Индии. В последующем ископаемые остатки рамапитека были найдены на территории Кении, Венгрии, Турции, КНР. Возраст рамапитека определен в 14 млн лет. Антропологи считают важным признаком эволюционное уменьшение размеров клыков гоминид. Сравнение размеров, формы и расположения зубов человека, человекообразных обезъян и рамапитека показывает, что человекообразные обезьяны имеют крупные клыки и так называемое семиальное пространство между резцами и клыками в зубной аркаде, тогда как у человека клыки по размерам меньше, чем у обезьян, семиальное пространство отсутствует, а зубная арка имеет специфическое закругление (рис. 172). Поскольку у рамапитека тоже нет этого пространства, его клыки имеют меньшие размеры, а зубная аркада закруглена, то часто считают, что он больше напоминает человека. Поскольку приматы используют их клыки для раскалывания орехов, разжевывания плодов или в качестве оружия, то пред полагают, что члены рода Ramapithecus уже не использовали клыки в качестве оружия. Вместо клыков для этих целей они использовали палки, камни. Однако рамапитек все же больше походил на обезьян, чем на человека. Его рост составлял около 1 м, обитал он на земле, питался зернами и корнями, мог частично пе полагают, что члены рода Ramapithecus уже не использовали клыки в качестве оружия. Вместо клыков для этих целей они использовали палки, камни. Однако рамапитек все же больше походил на обезьян, чем на человека. Его рост составлял около 1 м, обитал он на земле, питался зернами и корнями, мог частично пе редвигаться на задних конечностях, хотя большую часть времени он проводил на деревьях (как шимпанзе). Вопрос о сознании у рамапитеков спорен. Однако известны данные, в соответствии с которыми расхож дение эволюционных путей гоминид и понгид датируется временем между 6-7 млн лет назад. Поскольку у человека 23 пары хромосом, а у обезьян 24 пары, поскольку установлено, что гены недостающей обезьяньей пары локализованы на 2-1 паре хромосом человека, то предполагают, что расхождение эволюционных путей было детерминировано большой хромосомной транслокацией, следствием которой явилось включение обезьяньей хромосомы во вторую хромосомную пару человека. редвигаться на задних конечностях, хотя большую часть времени он проводил на деревьях (как шимпанзе). Вопрос о сознании у рамапитеков спорен. Однако известны данные, в соответствии с которыми расхож дение эволюционных путей гоминид и понгид датируется временем между 6-7 млн лет назад. Поскольку у человека 23 пары хромосом, а у обезьян 24 пары, поскольку установлено, что гены недостающей обезьяньей пары локализованы на 2-1 паре хромосом человека, то предполагают, что расхождение эволюционных путей было детерминировано большой хромосомной транслокацией, следствием которой явилось включение обезьяньей хромосомы во вторую хромосомную пару человека.
После рамапитека современная антропология не располагает какими-либо ископаемыми останками, в результате чего еще невозможно составить полный эволюционный ряд для человека, хотя на территории Эфиопии и обнаружен, вероятно, самый ранний гоминид A. ramidus, возраст которого составляет около 4,4 млн лет. Тем не менее условно филогенез человека можно представить в виде ряда последовательных, сменяющихся одна за другой стадий. В частности, различают несколько таких стадий, а именно:
1. Предшественник — предки рода Homo, представляющие собой ископаемых обезьянолюдей (австралопитеков).
2. Архантропы — древнейшие люди, представленные ископаемыми остатками.
3. Палеантропы — древние люди, непосредственные предки Н. sapiens. Представлены ископаемыми останками.
4. Неоантропы — ископаемые формы, связанные с культурами позднего палеолита (Н. sapiens fossilis) и ныне живущие формы (Н. sapiens ricens).
Предполагают, что эволюция предков человека была постепенной и с разной скоростью в разные периоды.
Стадия предшественников
. В 1924 г. профессор Иоганнесбург-ского университета Р. А. Дарт обнаружил в Южной Африке череп ископаемого гоминида, который по строению костей лицевого отдела и зубов принадлежал существу, занимавшему среднее положение между обезьяной и человеком. Р. Дарт назвал это существо австралопитеком или южной обезьяной (от лат. australis — южный, pithecus — обезьяна). Возраст южно-африканского австралопитека составляет около 3,0—2,3 млн лет. Спустя несколько лет Р. Дарт повторно нашел в Южной Африке ископаемые останки аналогичного существа, названного Australopithecua africanus. Изучение скелетных останков этого существа (рис. 173) показано, что его масса составляла 35—55 кг, зубы почти сходны с человеческими, а объем головного мозга — до 500 мл. Хотя объем головного мозга не достигает и половины объема мозга человека, он все же превышает объем мозга высших обезьян. Австралопитеки обладали способностью к прямохождению (рис. 174).
Ископаемые останки гоминид, которые характеризовались уменьшенными клыками, увеличенным мозгом и способностью к прямохождению, были обнаружены в 1978 г. в Восточной Африке (Танзания и Эфиопия). Их возраст составляет около 3,9-3,4 млн лет. Эти останки, названные останками Люси, были отнесены к виду A. afarensis.
 В 1994 г. в Эфиопии были обнаружены останки существа, названного A. ramidus. Возраст этих останков 4,4 млн лет, а его зубы более примитивны по сравнению с A. afarensis. Считают, что это существо обитало во время между расхождением эволюционных ветвей гоминид и пон-гид и появлением A. afarensis. В 1994 г. в Эфиопии были обнаружены останки существа, названного A. ramidus. Возраст этих останков 4,4 млн лет, а его зубы более примитивны по сравнению с A. afarensis. Считают, что это существо обитало во время между расхождением эволюционных ветвей гоминид и пон-гид и появлением A. afarensis.
В 1938 г. в Южной Африке были обнаружены ископаемые останки «сильного» (по нижней челюсти) австралопитека, которого называют сейчас A. robustus. Его рост составлял 150—155 см, масса — до 70 кг, а возраст — около 1,9-1,6 млн лет.
В 1959 г. в Олдовайском ущелье (Танзания) английский антрополог Л. Лики (1903—1972) нашел ископаемые останки существа с объемом мозга около 650 мл (как у австралопитеков), массой около 49 кг и галечные орудия, древность которых по данным калий-аргонового метода составляет 2 млн лет. Это существо вначале было названо Zinjanthropus boisei (от Зиндзе — арабское название В. Африки и греч. anthropus — человек), но позднее этот «гиперсильный» австралопитек был переименован в A. boisei.
На основании изучения тазовых и бедренных костей австралопитеков многие антропологи считают, что они были двуногими существами, способными стоять и передвигаться подобно человеку. Поскольку ископаемые останки австралопитеков находили в пещерах, где одновременно обнаруживали и зазубренные (подправленные) камни, то считают, что они жили в пещерах, которые использовали в качестве убежищ, и изготовляли простейшие орудия (рис. 175). Австралопитеки были охотниками на мелких животных, но использовали также в пищу зерна, плоды диких деревьев, корни. Вегетарианская пища, обилие растений и их семян, возможно, способствовали эволюции более мелких клыков австралопитеков.

 В 1960 г. в Африке Л. Лики нашел череп другого существа, уже более близкого к человеку, чем австралопитек и древность которого составляет 2 млн лет. На основе изучения черепа было предположено, что рост этого существа равен 120 см, объем головного мозга — около 650 мл. Первый палец его стопы уже не был отведен в сторону, что указывает на полное завершение прямохож-дения. Кости кисти этого существа сочетали как примитивные (обезьяньи), так и прогрессивные (человеческие) черты. Поскольку вместе с костными останками этого существа были найдены грубые галечные орудия и расколотые кости (для добывания мозга), то Л. Лики заключил, что это существо было и творцом этих орудий. Чтобы подчеркнуть способность этого существа изготовлять простейшие орудия, Л. Лики назвал его Н. habilis (человек умелый). В 1960 г. в Африке Л. Лики нашел череп другого существа, уже более близкого к человеку, чем австралопитек и древность которого составляет 2 млн лет. На основе изучения черепа было предположено, что рост этого существа равен 120 см, объем головного мозга — около 650 мл. Первый палец его стопы уже не был отведен в сторону, что указывает на полное завершение прямохож-дения. Кости кисти этого существа сочетали как примитивные (обезьяньи), так и прогрессивные (человеческие) черты. Поскольку вместе с костными останками этого существа были найдены грубые галечные орудия и расколотые кости (для добывания мозга), то Л. Лики заключил, что это существо было и творцом этих орудий. Чтобы подчеркнуть способность этого существа изготовлять простейшие орудия, Л. Лики назвал его Н. habilis (человек умелый).
В 60-70-е гг. к востоку от озера Рудольф (Кения) Л. Лики нашел остатки галечной культуры, возраст которых составляет 2,6 млн лет. Здесь же было обнаружено множество скелетных останков австралопитеков и более прогрессивных гоминид древностью тоже 2,6 млн. лет. В 1972 г. Л. Лики нашел в районе к востоку от озера Рудольф новый череп, изучение которого позволило предположить, что объем мозга этого гоминида составлял 800 мл, что на 150 мл крупнее мозга Н. habilis. Кроме того, череп этого гоминида был лишен надглазничного валика, характерного для всех ископаемых гоминид.
Принципиальный вопрос заключается в эволюционных отношениях между плио-плейстоценовыми гоминидами. В современной литературе опубликовано несколько филогении основных таксонов гоминид. Л. Лики считал, что Н. habilis является результатом эволюции A. africanus. В 1966 г. Л. Лики с сотрудниками опубликовали результаты изучения 22 особенностей черепа KNM 17 000 и низшей челюсти KNM-WT 16005, принадлежавших A. boisei и найденных в плиоценовых отложениях (возраст около 2,5 млн лет) к западу от озера Туркана в Кении, в сравнении с ископаемыми остатками A. afarensis, A. africanus, A. robustus и A. boisei. В соответствии с этими результатами предполагается, что A. africanus эволюционировал независимо от A. robustus и A. boisei, которые родственны между собой и что эволюционная линия, ведущая от австралопитека к роду Homo, начинается с A. africanus (рис. 176).
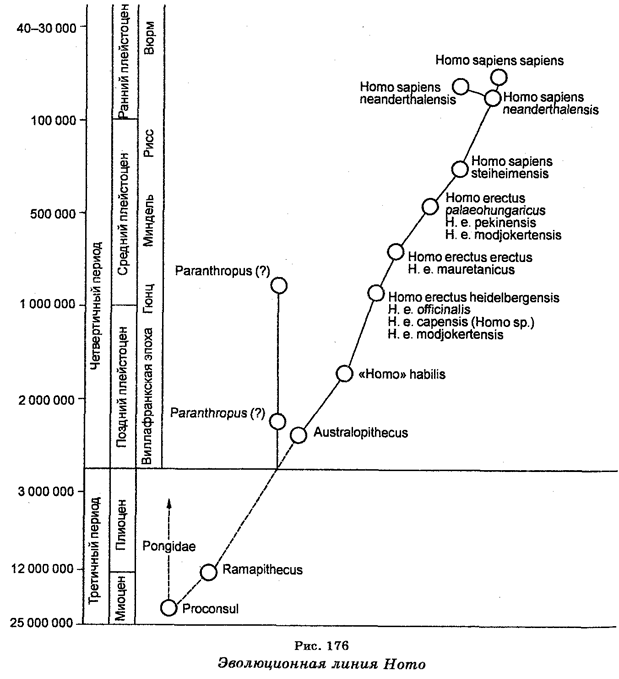
Итак, палеонтологические данные в совокупности свидетельствуют о том, что непосредственными животными предками человека служат австралопитеки — высокоразвитые двуногие обезьяноподобные существа. Их рост составлял 120-130 см, вес — около 50 кг, объем головного мозга — около 500 мл. Их коренные и передние зубы были человеческими по форме, а тазобедренные кости имели то же строение, что и аналогичные кости человека. Предполагают, что двуногость (прямохождение), являющаяся выдающимся эволюционным преобразованием, развивалась в результате естественного отбора в связи с жизнью австралопитеков на открытой равнине или по краям лесов на протяжении нескольких миллионов лет. Прямым следствием прямохождения явилось развитие хватательных движений и держание за счет использования рук. Между тем использование рук требовало рассудочной деятельности. Поэтому появление прямохождения и освобождение передних конечностей сопровождалось развитием головного мозга, увеличением его размеров. У австралопитеков начался также процесс потери волосяного покрова, в связи с чем они стали использовать пещеры для укрытия. Они, вероятно, начали использовать и огонь. Пища была частично вегетарианской, частично мясной. Таким образом, миновав 3 млн лет назад главный этап своей эволюции, заключавшийся в переходе от использования передних конечностей для передвижения по деревьям и земле к свободному передвижению в вертикальном положении при прямом торсе на сводчатой стопе, а также в использовании рук для изготовления простейших орудий, австралопитеки пересекли, по существу, границы звериной сущности. Можно сказать, что они уже не были обезьянами, но они еще не были и людьми.
Стадия архантропа (древнейшие люди).
Архантропы занимают среднее положение между австралопитеками и современным человеком. С зоологической точки зрения они входят в один комплекс или вид Н. erectus (человек выпрямленный).
Открытие архантропов произошло в 1891 гг., когда голландский врач Е. Дюбуа обнаружил на острове Ява в плейстоценовых отложениях на глубине 15м коренной зуб, бедренную кость и часть черепной коробки ископаемого существа, названного им Pithecanthropes erectus (от греч. pithecus — обезьяна, anthropos — человек), или яванским человеком. Для этого питекантропа были характерны мозг объемом 900 мл, что в два раза больше мозга современных человекообразных обезьян и больше мозга австралопитеков, вертикальное положение тела, почти человеческий скелет, зубы, сходные с человеческими, доминирование черепной коробки над лицевым отделом черепа, но толстые кости черепа. Рост питекантропа составлял, вероятно, 170 см, масса — около 70 кг, возраст — около 650 тыс лет. В послед.укмцем на Яве в более поздних слоях были обнаружены также грубые каменные орудия, что свидетельствует о способности яванского питекантропа к трудовой деятельности.
В 1927 г. вблизи Пекина в Китае канадским анатомом были найдены останки ископаемого существа, названного синантропом (пекинским человеком) и занимающего более высокое место по сравнению с яванским питекантропом. Объем мозга этого существа равен 1055 мл, рост — 150 см, возраст — 400 тыс лет. Для этого существа характерным были двуногое хождение, прямой торс. В пещере вместе с останками синантропа были найдены простейшие орудия из кварцита и известняка, а также кости. Эти находки свидетельствуют о том, что синантропы производили орудия труда, пользовались огнем. Предполагается, что они были каннибалами.
Костные останки ископаемых людей, сходных с питекантропами и синантропами, были обнаружены на территориях ФРГ (гей-дельбергский человек), Алжира (атлантроп), Венгрии, Чехии, Марокко и других стран. Абсолютный возраст этих людей составляет 400 тыс лет, хотя на Британских островах обитали люди, делавшие простые каменные орудия 500 тыс лет назад (средний плейстоцен). Древнейшие люди умели изготовлять орудия из камня и гальки. В 1985 г. Л. Лики и сотрудники нашли почти полный скелет 12-летнего Н. erectus к западу от озера Туркана в Кении. Возраст этого скелета составляет около 1,6 млн лет.
У всех Н. erectus над глазницами проходил сплошной, толстый (сильно выдающийся вперед) костный валик (надглазничный валик).

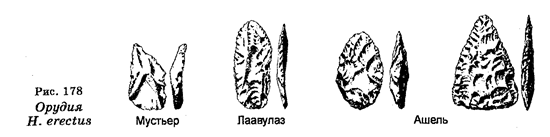
Древнейшие люди были более крупными, чем австралопитеки, могли ходить более долгое время прямо и без напряжения, полностью использовать руки (рис. 177). Их орудия были совершеннее орудий австралопитеков (рис. 178). Превосходя Н. habilis, они охотились на них и убивали их. Обитая в разных географических районах (рис. 179), древнейшие люди жили в пещерах, использовали огонь, но не могли его добывать самостоятельно, а лишь поддерживали. Судя по увеличенному размеру мозга, в котором все еще оставалась уплощенной теменная доля, узкими височные доли, они должны были обладать речью (рис. 180). Считают, что деятельность Н. erectus по добыванию пищи путем охоты имела коллективный характер. Архантропы уже планировали и координировали свои действия, в результате чего могли вести охоту на крупных животных, включая мамонтов. Уже на этом этапе возникло разделение труда между теми, кто охотился, и теми, кто оставался в лагере для наблюдения за детьми, стариками и выполнения мелких работ. Вероятно, на этом этапе существовали у людей и болезни, многие из которых перешли в наше время.
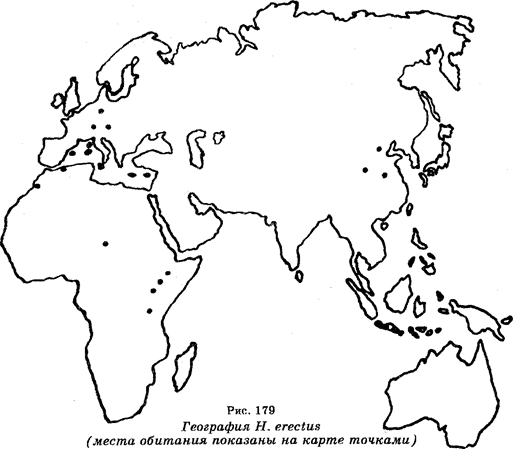
В частности, как свидетельствуют новейшие данные, у Н. erectus встречался сифилис.
Предполагают, что.представители Н. erectus берут начало от одной из ветвей австралопитеков, возможно Н. habilis, которые 2-1,5 млн лет назад начали расселяться по Африке, а затем проникли в Средиземноморье, Южную, Центральную и Юго-Восточную Азию. Имея долгую эволюционную историю, все представители Н. erectus тем не менее характеризуются недостаточным темпом прогрессивных морфологических изме-ненний. Их морфологические преобразования завершились появлением лишь форм с менее угловатым черепом и более тонкими костными структурами, но эти изменения не сопровождались резким совершенствованием методов изготовления каменных орудий. На этой стадии в эволюции человека в качестве фактора эволюции главенствовал естественный отбор. Предполагают также, что популяции Н. erectus были прямыми предками ранних представителей палеантропов, хотя точное время, способ трансформации в палеантропов и география процесса остаются дискуссионными.
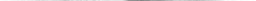 Стадия палеантропа (древние люди). В 1856 г. в долине Не-андерталь реки Дюссель в ФРГ во время добычи известняка были обнаружены останки скелета (череп и другие костные останки) древнего человека, которые были описаны К. Фульроттом, а в 1864 г. У. Кингом названы неандертальцем (Homo sapiens neanderthalensis). В последующем остатки неандертальцев были обнаружены в более чем 400 местах на европейском, африканском и азиатском континентах, что позволило воссоздать общую картину их строения и образа жизни (рис. 181). Стадия палеантропа (древние люди). В 1856 г. в долине Не-андерталь реки Дюссель в ФРГ во время добычи известняка были обнаружены останки скелета (череп и другие костные останки) древнего человека, которые были описаны К. Фульроттом, а в 1864 г. У. Кингом названы неандертальцем (Homo sapiens neanderthalensis). В последующем остатки неандертальцев были обнаружены в более чем 400 местах на европейском, африканском и азиатском континентах, что позволило воссоздать общую картину их строения и образа жизни (рис. 181).
Череп этих древних людей был значительно удлинен, с характерным низким сводом и сложными выступающими надбровными валиками, хорошо развитыми лобными пазухами. Объем мозга составляет 1200-1600 мл (как у современного человека), рост — около 160 см. Для неандертальцев характерны широкие зубы. Считают, что они уже обладали речью, но, вероятно, нечленораздельной. Новые данные свидетельствуют в пользу того, что неандертальцы были уже людьми, мало отличающимися от людей современного типа.
Многие антропологи предполагают, что неандертальцы произошли от каких-то архантропов около 300-200 тыс лет назад, причем их было две группы — ранние, или пренеандертальцы, и поздние (классические или западноевропейские). Ранние неандертальцы произошли от одной из популяций Н. erectus, жили в течение 300-250 тыс лет. Многие находки ископаемых останков в разных местах свидетельствуют о значительной изменчивости популяций ранних неандертальцев. Они являлись предками как поздних (классических) неандертальцев, так, возможно, и современных людей. Широкое распространение поздних неандертальцев на территориях, ныне занимаемых Францией, Бельгией, Испанией, Италией, приходится на период 200-35 тыс лет назад. У них были сильно развитое надбровье в виде соединения надбровных дуг у основания носа костным наростом, приплюснутый сверху затылок, резко увеличенная емкость черепной коробки. Средние объемы головного мозга составляли 1400—1450 мл, средний рост 155—165 см. Многочисленные находки скелетов неандертальцев свидетельствуют о том, что их средний возраст составлял 30 лет, хотя отдельные из них доживали до 50 лет. Нельзя не заметить, что такой средний возраст людей был и в период средневековья. Практически отсутствовал кариес зубов. Время жизни неандертальцев совпало с периодом очередного (последнего) наступления ледников. Они строили простые жилища, защищались от холода с помощью одежды из шкур зверей, изготовляли более совершенные кремниевые и костяные орудия, что позволяло им успешно вести борьбу с мамонтами (рис. 182). Неандертальцы хоронили умерших или погибших соплеменников.
Неандертальцы внезапно исчезли 40-30 тыс лет назад. Предполагают, что они были либо поглощены, либо частично истреблены людьми современного типа. Больше того, поскольку у них были недоразвитыми лобные доли головного мозга, которые обычно ответственны за поведение, то предполагают, что конфликты между неандертальцами способствовали распаду их групп (общин), вели к самоистреблению. На территории Израиля найдены ископаемые останки неандертальца, возраст которых составляет 50 тыс лет. Это означает, что люди современного типа и неандертальцы в этом регионе жили одновременно. Однако современные данные не подтверждают предположения о том, что неандертальцы были предками современного человека. Считают, что эволюционное расхождение между неандертальцами и людьми современного физического типа является очень большим. Больше того, известен взгляд о том, что Н. erectus, H. neandertaleusis и Н. sapiens являются разными видами рода Homo.
Стадия неоантропа.
Под этим названием понимают как ископаемые формы человека современного физического типа, так и ныне живущих людей. В 1868 г. в пещере Кро-Маньон (Франция) был найден скелет человека, объем черепной коробки которого составляет около 1800 см8
. Для этого человека характерно отсутствие надбровных валиков, т. к. это костное образование разделилось с помощью костного углубления на две дуги, идущие по верхнему краю глазниц. Оно превратилось в надбровные дуги, характерны также наличие подбородка, прямой лоб. Рост его составлял около 180 см. Этого человека назвали кроманьонцем. В дальнейшем сходные ископаемые формы были найдены также на территориях Китая, Палестины, Чехии, России и других местах Европы, Африки и Азии.
Средний возраст кроманьонцев составлял около 30 лет, хотя отдельные из них могли жить до 50 лет и более.
Палеонтологические находки указывают на то, что в период появления современных людей доживали свой век и последние неандертальцы. В частности, на территории Палестины найдены скелетные останки гибридов между неандертальцами и людьми современного типа.
В соответствии с новыми данными, представители Н. sapiens современного физического типа появились 100—200 тыс лет назад.
Кроманьонцы не отличаются от людей современного физического типа. Если бы кроманьонец, одетый в современную одежду, шел по улице, его трудно было бы сейчас выделить в толпе. В их пещерах были найдены стрелы, гарпуны, иглы, что указывает на усовершенствование методов изготовления орудий. Кроманьонцы, как и неандертальцы, были искусными охотниками. Подсчитано, что они во многом ответственны за истребление многих млекопитающих. В частности, они ответственны за истребление мегатериев, мамонтов, саблезубых кошек, бизонов. Больше того, примерно около трети родов млекопитающих было уничтожено еще 50 тыс лет назад. Предполагают, что резкое уменьшение диких животных способствовало переходу людей от охотничьего сообщества к сельскохозяйственному.
На стадии неоантропа на развитие человека стала оказывать все большее и большее влияние культура, социальные закономерности.
Вопрос о прародине человека, под которой понимают место, где протекал процесс самой ранней стадии антропогенеза, возник еще в прошлом веке. Палеонтологи, антропологи и археологи в качестве возможного центра зарождения человечества называли самые различные части земного шара (почти все континенты). Однако Ч. Дарвин в качестве прародины человека назвал Африку, отмечая, что именно в Африке обитают обезьяны, стоящие по морфологии и развитию наиболее близко к современному человеку. Действительно, наибольшая концентрация и разнообразие форм высокоорганизованных обезьян были достигнуты вначале в Африке и Южной Азии. На этих континентах были обнаружены и рамапитеки, имеющие прогрессивные признаки. Тем не менее вопрос этот обсуждают и в наше время. Одновременно обсуждают вопрос и о заселении людьми современного физического типа других континентов.
Многие антропологи считают, что судьба процесса гоминиди-зации оказалась разной на африканском и азиатском континентах вследствие, главным образом, климатических условий. В плиоцене (13 млн лет назад) в Южной Азии резко изменились климатические условия в неблагоприятном направлении, стали часто возникать засухи, происходило поднятие Гималаев, которое сопровождалось образованием пустынных пространств. Складывавшиеся там чрезвычайно неблагоприятные условия жизни привели к вымиранию или переселению гоминид в другие районы земного шара, что остановило процесс их эволюции на азиатском континенте. Напротив, в Африке сложились совершенно другие климатические условия. В частности, в Восточной Африке и особенно в ее приэкваториальной части сформировался очень благоприятный для жизни климат, сложился чрезвычайно богатый ландшафт, обеспечивающий существование гоминид. Поэтому именно в Восточной Африке продолжился процесс гоминидизации, в результате которого она оказалась прародиной человека. Можно сказать, что здесь развитие гоминид достигло такого уровня, когда они уже полностью отделились от животных, стали приобретать специфические человеческие черты. Таким образом, Восточная Африка оказалась прародиной человека.
Если в вопросе о прародине человека разногласий среди антропологов почти нет, то совершенно отсутствует согласие о времени движения первых гоминид из Африки на другие континенты, в частности в Азию. Тем не менее новейшие данные позволяют предполагать, что первые гоминиды пришли из Африки в Азию около 2 млн лет назад, но они были более примитивными по сравнению с Н. erectus. В Азии первые гоминиды эволюционировали в Н. erectus, представители которого вернулись затем в Африку и в Европу.
Считают далее, что первые люди современного физического типа мигрировали из Африки в другие районы, замещая там архаичные человеческие группы. Человек современного физического типа вступил из Африки в Ю.-В. Европу 37—44 тыс лет назад, откуда он перешел в 3. Европу около 34 тыс лет назад. Такой хронологической последовательности в миграции человека на заре его истории придерживаются очень многие антропологи. Однако на Британских островах недавно найдена бедренная кость человека, возраст которого составляет порядка 500 тыс лет, что заслуживает особого внимания и пока не имеет убедительных объяснений.
В последние годы для выяснения времени появления человека современного физического типа стали использовать молекулярно-генетические методы.
Как свидетельствуют результаты исследований мДНК индивидов, принадлежащих к разным европейским, американским, африканским и азиатским популяциям, трансформация архаичных форм в Н. sapiens произошла в Африке в период между 100-150 тыс лет назад. На основе этих данных сейчас считают, что современные люди являются потомками «митохондриальной Евы», жившей в Африке 140-280 тыс лет назад.
В пользу африканского происхождения человека современного физического типа свидетельствуют также результаты молукуляр-но-генетических исследований повторов ДНК длиной 2000—6000 азотистых оснований, которые локализованы в том генетическом районе хромосомы Y, который контролирует мужской пол. Эти результаты показали, что дивергенция хромосомы Y человека от Y-хромосомы его предка произошла в интервале 500—400 тыс лет назад. Предполагают, что «молекулярный» Адам, который первый стал обладателем Y-хромосомы человека, гулял по Земле в то же время, что и «митохондриальная» Ева. Данные об отдельных ДНК-последовательностях подтверждают мысль о том, что современная Y-хромосома стала распространяться в разные районы мира после того, как ее носители мигрировали из Африки.
Молекулярно-генетические исследования показали, что в мтДНК папуасов Новой Гвинеи имеется делеция, которая обнаруживается также в мтДНК людей многих африканских популяций, но отсутствует в мтДНК людей азиатских популяций. По этой причине есть предположения о том, что папуасы, возможно, пришли в Новую Гвинею из Африки через Индийский океан. В Северную Америку они пришли из Северного Китая через Берингию и Аляску. Что касается Южной Америки, то есть данные, что люди современного физического типа жили уже 32 тыс лет назад на той территории, которую в наше время называют Бразилией.
Однако существует альтернативный взгляд, который заключается в том, что представители Н. sapiens обитали одновременно около 1 млн лет назад как в Африке, так и в Азии, и что трансформация архантропов в современных людей шла во многих местах Старого Света. Между тем палеонтологические данные не подтверждают мысль о том, что азиатские формы Н. erectus являются предками Н. sapiens. Скорее можно предполагать, что популяции неафриканских форм Н. erectus замещались сапиентами, мигрировавшими из Африки. Люди современного физического типа появились в Восточной Азии 75-150 тыс лет назад. Молекулярно-генетические данные также не подтверждают эту гипотезу.
Осмысливая происхождение человека, Ч. Дарвин придавал большое значение таким факторам антропогенеза, как изменчивость телесных и психических свойств предков человека, использование ими тех или иных органов, естественный и половой отбор.
Естественный отбор действовал в популяциях как древнейших, так и древних людей. Однако он шел не только но физическим признакам. Он благоприятствовал таким чертам, как умение производить и использовать орудия, защита стариков, коллективные способы вести охоту и др. Можно сказать, что наряду с индивидуальным шел и групповой отбор.
Но действие естественного отбора полностью не прекратилось и в современную эпоху, о чем свидетельствуют данные, в соответствии с которыми в настоящее время в отдельных географических районах мира идет контрселекция индивидов на устойчивость к различным болезням. Например, среди жителей Африки и Средиземноморья распространена серповидноклеточная анемия, но гетерозиготы по этой анемии устойчивы к малярии. Точно так же, дефицит по глюкозе-6-фосфатдегидрогеназе также сопровождается повышением у таких индивидов резистентности к малярии.
В Центральной Азии очень низка встречаемость групп крови I (0), но очень высокая встречаемость группы III (В). В этом «повинны» былые эпидемии чумы, к которой обладатели группы крови I обычно характеризуются пониженной сопротивляемостью. Лица с группой крови II (А) очень редки в эндемичных районах по оспе, в Индии, Китае, Африке, поскольку они более восприимчивы к оспе.
Современные представления о факторах антропогенеза в общем сводятся к пониманию того, что человек является продуктом действия тех же факторов эволюции, которые создали живой мир, однако с учетом специфики человека должны были действовать также и специфические факторы антропогенеза.
Основы учения о специфических факторах антропогенеза были заложены Ф. Энгельсом еще в последней трети XIX в. Считая, что появление человека не является лишь ступенью в эволюции природы, ибо с человеком появляется новая сила в виде труда, который, воздействуя на природу, не только изменяет ее, но изменяет и человека. Ф. Энгельс назвал важнейшим специфическим фактором антропогенеза труд.
Можно сказать, что труд отделил человека от животных, а люди, благодаря совместной деятельности руки, органов речи и мозга, стали обладать способностью выполнять все усложняющиеся операции, ставить перед собой каждый раз все более высокие цели и достигать этих целей.
Однако признанию труда в качестве фактора антропогенеза мешает незнание причин, заставивших австралопитеков и более поздние формы прибегнуть к труду. В поисках объяснения этих причин некоторые исследователи предполагают, что происходившие в плиоцене бурные тектонические процессы, активный вулканизм, горообразование, землетрясения и другие факторы привели в Восточной Африке к обнажению урановых руд и повышению там уровня ионизирующей радиации. Являясь мутагенным фактором, ионизирующая радиация индуцировала у австралопитеков мутации, которые в совокупности сделали их настолько слабыми, что они оказались неспособными добывать пищу присущими им животными методами. Единственным в этих условиях путем выжить был переход части или отдельных австралопитеков от использования камней и палок к изготовлению более совершенных орудий нападения и защиты.
В последующем изготовление орудий обеспечило древнейшим людям выход на господствующее положение в природе. Поскольку для общения необходим был язык, то в головном мозге древнейших людей стали развиваться теменные и лобные доли (центры речи), стали совершенствоваться анализаторы.
Важным специфическим фактором антропогенеза явилась также мясная пища. Как предполагают, австралопитеки были хищными существами. Больше того, Н. habilis со своими орудиями, вероятно, оказался более опасным хищником и для самих австралопитеков. Возможно, что уже на этой стадии возник каннибализм.
Современные представления о роли этого фактора антропогенеза сводятся к тому, что употребление мяса обеспечивало предков современного человека незаменимыми аминокислотами.
Многие антропологи уделяют большое внимание такому возможному фактору антропогенеза, как альтруизм. Изготовление эффективных орудий древнейшими и древними людьми возможно было только на основе навыков, наследуемых от предыдущих поколений, что вело к повышению в первобытных сообществах роли и значения людей старшего поколения, хранящих опыт изготовления орудий, охоты и т. д. Чем лучше охранялись слабые, тем больше оставалось шансов на выживание. Видимо в этот период стала прекращаться внутривидовая борьба за существование, падать значение естественного отбора. Напротив, важным фактором антропогенеза становились возникавшие социальные закономерности.
Таким образом, создав социальные отношения в группах древнейших и древних людей, естественный отбор в этот период стал уступать свое место социальным факторам. В борьбе за существование стали побеждать не только те, которые были самыми сильными, но и те, которые охраняли слабых' (стариков, женщин и детей).
В результате этого в борьбе за существование преимуществ стали достигать те группы или племена, в которых больше заботились о поддержании стариков, ибо последние сохраняли накопленный опыт в изготовлении орудий, поисках мест охоты и съедобных растений, в ориентировке на местности и т. д.
Следовательно, если у ранних австралопитеков в основном действовал естественный отбор особей, лучше владеющих руками, а также отбор на повышение роли и эффективности орудийной деятельности, то в возникновении, скажем, Н. habilis большое значение имел альтруизм, ставший фактором эволюции и дававший многие преимущества в условиях коллективной жизни. Возникновение социальной среды стало необратимым. Популяции, лучше изготовлявшие и использовавшие орудия, вытесняли отстававшие в развитии популяции в районы, менее благоприятные для жизни, что вело там к их уничтожению.
Наконец, в антропогенезе действовали факторы, связанные с возникновением сельского хозяйства. Например, введение в питание молока вело к отбору индивидов, обладающих способностью утилизировать (расщеплять) лактозу. Известно, что в популяциях современных людей, не использующих молоко, концентрация генов, контролирующих расщепление лактозы, ничтожно мала.
Расы — это исторически сложившиеся группировки (группы популяций) людей разной численности, характеризующиеся сходством морфологических и физиологических свойств, а также общностью занимаемых ими территорий. Развиваясь под влиянием исторических факторов и принадлежа к одному виду (Н. sapiens), раса отличается от народа, или этноса, который, обладая определенной территорией расселения, может состоять из нескольких расовых комплексов и наоборот, к одной и той же расе может принадлежать ряд народов и носителей многих языков.
О существовании рас люди знали еще до нашей эры. Тогда же были предприняты и первые попытки объяснить их происхождение. Например, в мифах древних греков происхождение людей с черной кожей объяснялось неосторожностью Фаэтона (сына бога Гелиоса), который на солнечной колеснице так приблизился к Земле, что обжег стоявших на ней белых людей. Греческие философы в объяснениях причин возникновения рас большое значение придавали климату.
В соответствии с библейским описанием родоначальниками белой, желтой и черной рас были сыновья Ноя — Яфет, Сим и Хам (соответственно).
Стремления научно систематизировать представления о физических типах народов, населяющих земной шар, датируется XVII в., когда опираясь на различия людей в строении лица, цвета кожи, волос, глаз, а также особенностей языка и культурных традиций, французский врач Ф. Бернье впервые в 1684 г. классифицировал человечество на три расы — европеоидную, негроидную и монголоид-ную. Сходную классификацию, как известно, предлагал и К. Линней, который, признавая человечество в качестве единого вида, выделил дополнительно (четвертую) лапландскую расу (население северных районов Швеции и Финляндии).
В 1775 г. И. Ф. Блуменбах (1752-1840) разделял род человеческий на основе цвета кожи на 5 рас — кавказскую (белую), монгольскую (желтую), эфиопскую (черную), американскую (красную) и малайскую (коричневую). Почти через 200 лет У. Бойд (1953) на основе результатов изучения антигенов крови классифицировал человечество на пять рас, а именно:
1. Европейская группа, включающая лопарей, южных европейцев и северных африканцев.
2. Африканская группа.
8. Азиатская группа, включая жителей индийского субконтинента.
4. Американская группа, включая всех аборигенов.
5. Тихоокеанская группа (меланезийцы, полинезийцы, австралийцы).
На основе понимания расогенеза в качестве процесса, продолжающегося и в настоящее время, Т. Добжанский (1962) классифицировал человечество на 34 расы (рис. 183), а именно:
1. Северо-западная европейская — жители Скандинавии, северной части Германии, Северной Франции, Великобритании и Ирландии.
2. Северо-восточная европейская — жители Польши, европейской части бывшего СССР, большинство существующих популяций в Сибири.
3. Альпийская — жители территорий, простирающихся от Центральной Франции, южной части Германии, Швейцарии, Северной Италии до побережья Черного моря.
4. Средиземноморская — популяции по обе стороны Средиземноморья, от Танжера до Дарданелл, Аравии, Турции, Ирака.
5. Хинду — жители Индии, Пакистана.
6. Тюркская — жители Туркестана, Западного Китая.
7. Тибетская — жители Тибета.
8. Северо-китайская — жители Северного и Центрального Китая, Манчжурии.
9. Классическая монголоидная — жители Сибири, Монголии, Кореи, Японии.
10. Эскимосская — жители арктической Азии и Америки.
11. Юго-восточная азиатская — жители Южного Китая до Таиланда, Бирмы, Малайи и Индонезии.
12. Айну — популяция аборигенов в Северной Японии.
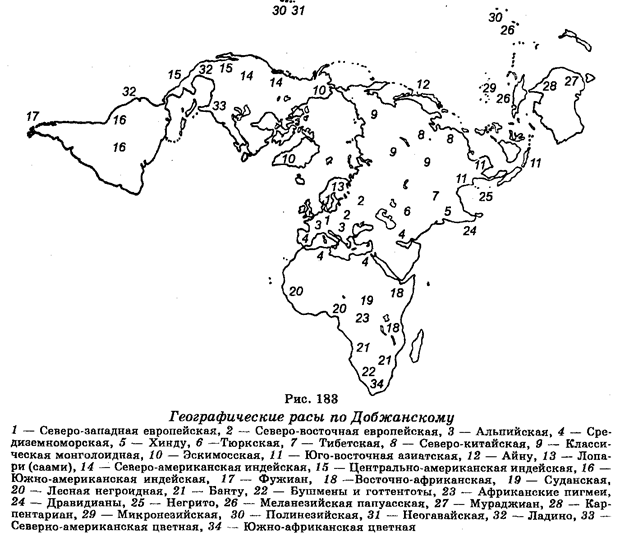
13. Лопари (саами) — аборигены арктической Скандинавии и Финляндии.
14. Северо-американская индейская — туземные популяции Канады и США.
15. Центрально-американская индейская — популяции, обитающие на территориях, простирающихся от юго-западных районов Северной Америки через Центральную Америку до Боливии.
16. Южно-американская индейская — популяции Перу, Боливии и Чили, занимающиеся главным образом сельским хозяйством.
17. Фужиан — аборигены юга Южной Америки, не занимающиеся сельским хозяйством.
18. Восточно-африканская — популяции Восточной Африки, Эфиопии, части Судана.
19. Суданская — популяции большей части Судана.
20. Лесная негроидная — популяции, обитающие в лесах Западной Африки и вдоль большей части р. Конго.
21. Банту — аборигены Южной Африки и части Восточной Африки.
22. Бушмены и готтентоты — аборигены, населяющие Южную Африку.
23. Африканские пигмеи — популяция малорослых людей, обитающих в лесах Экваториальной Африки.
24. Дравидианы — аборигены Южной Индии и Цейлона.
25. Негрито — популяция малорослых людей с вьющимися волосами на территории от Филиппин до Андаманса, Малайи и Новой Гвинеи.
26. Меланезийская папуасская — аборигены Новой Гвинеи до Фиджи.
 27. Мураджиан — популяция аборигенов Юго-Восточной Австралии. 27. Мураджиан — популяция аборигенов Юго-Восточной Австралии.
28. Карпентариан — популяция аборигенов Северной и Центральной Австралии.
29. Микронезийская — популяции островов западной части Тихого океана.
30. Полинезийская — популяции островов центральной и восточной части Тихого океана.
31. Неогавайская — популяция, недавно возникшая на Гавайских островах.
32. Ладино — популяция, недавно возникшая в Центральной и Южной Америке.
33. Северно-американская цветная — негритянская популяция в США.
34. Южно-африканская цветная — негритянская популяция Южной Африки.
Наряду с этими расами, получившими название локальных рас, иногда выделяют 9 географических рас, поскольку они приурочены к определенным географическим территориям (европейская, индийская, азиатская, индейская, африканская, австралийская, меланезий-ско-папуасская, микронезийская и полинезийская). Использование классификации рас по географическому признаку удобно, однако такая классификация не отражает сущности расовых группировок, сформировавшихся на тех или иных географических территориях.
В практическом плане очень популярна классификация рода человеческого на европеоидов, негроидов, монголоидов и австрало-идов(рис. 184).
Европеоиды светлокожи, для них характерны прямые или волнистые светло-русые или темно-русые мягкие или средней жесткости волосы, серые, серо-зеленые и каре-зеленые широко открытые глаза, умеренно развитый подбородок, широкий таз, узкий и сильно выступающий нос, нетолстые губы и довольно обильный волосяной покров тела и лица. Для женщин этой расы характерны полусферические груди и выпуклые ягодицы. Люди, принадлежащие к этой расе, занимают большую часть Европы, а также прилегающие к ней области.
Негроиды темнокожи, для них характерны курчавые или шерстистые темные волосы, толстые губы, очень широкий и плоский нос, очень крупные зубы, карие или черные глаза, длинная голова, редкая растительность на лице и теле, узкий таз, большие ступни. Для женщин характерны груди конической формы и маловыпуклые ягодицы. Люди, принадлежащие к этой расе, занимают практически весь экваториальный пояс от Африки до островов Тихого океана. К этой расе принадлежит население Африки, а также негрито (пигмеи), океанические негроиды (меланезийцы), южноафриканские бушмены и готтентоты.
Монголоиды смуглокожи, обладают желтой или желто-коричневой кожей. Для них характерны прямые жесткие иссиня-чер-ные волосы, плоское крупное скуластое лицо, узкие и слегка раскосые карие глаза со складкой верхнего века (третьим веком или эпикантусом) во внутреннем углу глаза, плоский и довольно широкий нос, редкая растительность на лице и теле. Люди, принадлежащие к этой расе, занимают Восточную Сибирь и Монголию, Дальний Восток, Центральную и Юго-Восточную Азию. Смешанную монголоидную расу представляют индонезийцы и американские индейцы.
Часто в виде отдельной расы выделяют австралоидов, которые почти так же темнокожи (их кожа имеет шоколадный цвет), как и негроиды, но для них характерны темные волнистые волосы, крупная голова и массивное лицо с очень широким и плоским носом, выступающим подбородком, значительный рост волос на лице и теле. Австралоиды являются аборигенами Австралии. Однако австралоидов часто считают негроидами. Иногда выделяют америка-ноидов, для которых характерны смуглая кожа, скуластое лицо, достаточно выступающий нос и эпикантус, иссиня-черные волосы. Однако часто американоидов относят к монголоидам.
Распределение групп крови и типов отпечатков пальцев в популяциях, принадлежащих к разным расам, показано в табл. 39 и табл. 40.
Правильное понимание процессов расогенеза зависит от подходов к определению сущности и классификации рас. Длительное время господствовал так называемый типологический подход, в соответствии с которым определение расы проводили на основе стереотипов, якобы отражающих все признаки расы. Поэтому, руководствуясь характеристиками отдельных индивидов, считали, что между расами существуют абсолютные различия. Между тем развитие популяционной генетики показало, что типологический подход к пониманию природы рас не имеет достаточной аргументации.
Таблица 39
Распределение групп крови в разных популяциях (в % )
| Популяция
|
А
|
АВ
|
В
|
0
|
| Европеоиды
|
5-40
|
1-37
|
4-18
|
45-75
|
| Негроиды
|
8-30
|
1-8
|
10-20
|
52-70
|
| Монголоиды
|
0-45
|
0-5
|
16-25
|
39-68
|
| Американские индейцы
|
0-20
|
около 0
|
0-4
|
68-100
|
Таблица 40
Распределение типов отпечатков пальцев в разных популяциях
(в % )
| Популяция
|
Типы отпечатков
|
| арковидные
|
петлевидные
|
закругленные
|
| Европеоиды
|
0-9
|
63-76
|
20-42
|
| Негроиды
|
3-12
|
53-73
|
20-40
|
| Бушмены
|
13-16
|
66-68
|
15-21
|
| Китайцы, японцы и родственные популяции
|
1-5
|
43-56
|
44-54
|
| Американские индейцы
|
2-8
|
46-61
|
35-57
|
| Австралийские аборигены
|
0-1
|
28-46
|
52-73
|
| Микронезиипы
|
2
|
49
|
49-60
|
Научно обоснованный подход к классификациям рас является популяционным подходом. Эффективность классификаций рас зависит от количества используемых признаков, ибо чем больше признаков используют для проведения различий между расами,- тем более эффективной оказывается классификация. Поскольку расы являются популяциями, то, описывая какую-либо расу, описывают фактически большую группу людей, но не отдельного индивида. Поскольку в пределах каждой расы имеются громадные вариации наследственных характеристик, то невозможно найти критерии, на основе которых можно было бы подтвердить или исключить принадлежность всех индивидуумов к разным расам. Точно так же, как невозможно найти индивидуумов со всеми признаками, присущими одной расе. Расы должны рассматриваться как группы, но не как индивиды. Например, сравнение черепных костей шведов и русских показало, что между этими национальными группами имеются существенные различия, но они являются средними различиями, а между индивидуумами в каждой группе имеется перекрываемость. Таким образом, определение расовых различий по популярным стереотипам (типологически) является неправильным, поскольку абсолютных различий между расовыми группами не существует.
Представления о происхождении рас и первичных очагов расо-образования отражены в нескольких гипотезах. В соответствии с гипотезой полицентризма или полифилии (Ф. Вайденрайх, 1947) существовало четыре очага расообразования — в Юго-Восточной Азии и на Больших Зондских островах, в Восточной Азии, в Африке южнее Сахары и в Европе и Передней Азии. В Европе и Передней Азии сложился очаг расообразования, где на основе европейских и переднеазиатских неандертальцев возникли европеоиды.
В Африке на основе африканских неандертальцев возникли негроиды, в Восточной Азии на основе синантропов возникли монго-лоиды, а в Юго-Восточной Азии и на Больших Зондских островах развитие на основе питекантропов и яванских неандертальцев привело к формированию австралоидов. Следовательно, европеоиды, негроиды, монголоиды и австралоиды имеют свои собственные очаги расообразования. Предполагается, что главным в расогенезе были мутации и естественный отбор. Однако эта гипотеза вызывает возражения, поскольку неизвестны случаи, когда бы идентичные эволюционные результаты воспроизводились несколько раз (эволюционные изменения всегда являются новыми); неизвестны научные данные, подтверждающие то, что каждая раса имеет свой собственный очаг расообразования.
В рамках гипотезы дицентризма Ф. Г. Дебец (1950), А. Тома (1960) предложили два объяснения происхождения рас. По одному объяснению очаг расообразования европеоидов и африканских негроидов существовал в Передней Азии, тогда как очаг расообразования монголоидов и австралоидов был приурочен к Восточной и ЮгО-Восточной Азии. Европеоиды передвигались в пределах европейского материка и прилегающих к нему районов Передней Азии. Напротив, негроиды расселялись в основном вдоль тропического пояса. Монголоиды вначале заселяли Юго-Восточную Азию, затем часть их перешла на американский континент. По другому объяснению европеоиды, африканские негроиды и австралоиды составляют один ствол расообразования, тогда как азиатские монголоиды и американоиды составляют другой ствол. В обоснование своей гипотезы сторонники дицентризма приводят данные о сходстве строения зубов монголоидов и австралоидов. Однако эта гипотеза имеет те же возражения, что и гипотеза полицентризма.
В соответствии с гипотезой моноцентризма или монофилии (Я. Я. Рогинский, 1949), которая основана на признании общности происхождения, социально-психического развития, а также одинакового уровня физического и умственного развития всех рас, расы возникли от одного предка, в одном месте, но протяженность этого места была очень большой, измеряемой многими тысячами километров. Предполагается, что формирование рас произошло на территориях Восточного Средиземноморья, Передней Азии, возможно, Южной Азии. По этой гипотезе одна ветвь архаичных палеантропов дала начало монголоидам, которые как раса сформировались в несколько этапов. Архаичные палеоантропы, придя в Восточную Азию, встретили там архантропов, которых истребили или поглотили, унаследовав от них совкообразные резцы. Затем в Азию пришли люди современного типа. В результате метисации сложились протомонголоиды, часть которых 25-30 тыс. лет назад двинулась из Восточной Азии через Берингию в Северную Америку, дав начало американским индейцам. Сохранив свой основной тип, они приобрели новые особенности. Из части прото-монголоидов, оставшихся на азиатском континенте, несколько позже сформировались настоящие монголоиды. Другая ветвь архаичных палеоантропов дала начало европеоидным и негроид-но-австралоидным группам. Предки европеоидов и негроидов занимали территории Восточного Средиземноморья и Южной Европы от Испании до г. Воронежа. Формирование европеоидов и негроидов произошло в палеолите. Позднее в результате изоляции, миграции и расселения возникли более мелкие расовые группировки.
Выделяют четыре этапа расообразования (В. П. Алексеев, 1985). На первом этапе расообразования происходило выделение первичных очагов расообразования (территорий, на которых происходит этот процесс) и формирование основных расовых стволов — западного (европеоиды, негроиды и австралоиды) и восточного (азиатские монголоиды и американоиды). Хронологически этот этап датируется временем около 200 тыс. лет назад, т. е. совпадает с этапом формирования человека современного типа. Следовательно, оформление основных расовых сочетаний в западных и восточных районах Старого Света происходило одновременно с формированием признаков, присущих современному человеку, а также с переселением части человечества в Новый Свет. На втором этапе происходило выделение вторичных очагов расообразования и формирование эволюционных ветвей в пределах основных расовых стволов. Хронологически этот этап приходится на время около 15—20 тыс. лет назад. На третьем этапе выделялись третичные очаги расообразования, в пределах которых формировались локальные расы. По времени этот этап имел место около 10-12 тыс. лет назад. На четвертом этапе возникли четвертичные очаги расообразования, в которых формировались популяции с углубленной расовой дифференциацией, сходной с современной. Этот этап осуществлялся в эпоху бронзы и раннего железа (IV-III тысячелетия до нашей эры).
Факторами расогенеза являются естественный отбор, дрейф генов, изоляция и смешение популяции, причем наибольшее значение имел естественный отбор, который выполнял формообразущую роль на ранних стадиях формирования рас.
Действие естественного отбора проявлялось в разных направлениях, а его эффективность зависела от селективного значения признака. Например за цвет кожи ответственны кожные клетки, содержащие меланин, синтез которого детерминируется генетически. Наличие пигмента определяется наличием гена, контролирующего тирозиназу, обеспечивающую конверсию тирозина в меланин. Однако только этого гена еще недостаточно для объяснения цвета кожи. На пигментацию кожи оказывает влияние еще один ген, который детерминирует другой фермент, содержащийся в клетках людей с белой кожей и влияющий на количество меланина. Когда этот фермент синтезируется, то меланин образуется в малых количествах, и кожа является белой. Напротив, когда он отсутствует (не синтезируется), то меланин образуется в больших количествах, и кожа становится черной. Имеются данные, позволяющие считать, что в определении цвета кожи принимает участие также и меланинстимулирующий гормон. Таким образом, в контроле цвета кожи принимает участие как минимум три гена.
Действие естественного отбора в случае такого расового признака, как цвет кожи, объясняется связью между солнечным освещением и продукцией противорахитного витамина D, который необходим для поддержания в организме баланса кальция. Избыток этого витамина сопровождается отложением кальция в костях и-ведет к их хрупкости, тогда как недостаток кальция ведет к рахиту. Между тем количество витамина D, синтезируемого нормально в организме человека, контролируется количеством солнечного облучения, которое проникает в клетки глубже слоя меланина. Чем больше меланина в коже, тем меньше проникает света в клетки глубже меланина. Поэтому люди зависимы в продукции из витамина в от солнечного света. Чтобы витамин в продуцировался в количествах, достаточных для поддержания нормального баланса кальция, люди со светлой кожей должны находиться подальше от экватора. Напротив, люди с черной кожей должны быть ближе к экватору. Как видно, территориальное распределение людей с разной пигментацией кожи является функцией географической широты. Посветление кожи у европеоидов способствует более глубокому прохождению солнечного света в ткани человека, что ускоряет синтез противорахитного витамина D, который обычно медленно синтезируется в условиях недостатка солнечной радиации. Миграция людей с интенсивно пигментированной кожей к отдаленным широтам, а людей с недостаточно пигментированной кожей к тропическим широтам могла вести к дефициту витамина в у первых и избытку у вторых с вытекающими отсюда последствиями. Следовательно, цвет кожи имел селективное значение, являясь фактором контрселекции. Естественный отбор шел на цвет кожи, который возникал в качестве приспособительного признака к условиям географической среды обитания.
Другим расовым признаком, детерминирующим действие отбора, является тип телосложения. Например, эскимосы более низкорослы и плотны, чем зулусы. У эскимосов площадь поверхности тела является меньшей по сравнению с ее массой. Напротив, у зулусов, для которых характерно тонкое тело с длинными руками, площадь поверхности тела превалирует над его массой. В результате этого зулусы обладают преимуществом в выделении избытка тепла из тела, что очень важно в условиях жаркого климата. Поскольку отношение длины конечностей к весу тела изменяется у индивидов в направлении к экватору, то считают, что длинные конечности имеют преимущество по сравнению с массой тела в условиях жаркого климата. Длинные конечности обеспечивали индивидов дополнительной площадью тела, способствующей более быстрой потере тепла. Следовательно, большее значение этого отношения имело преимущество в жарком климате. Напротив, меньшее значение этого отношения имело преимущество в более прохладном климате, поскольку при меньшей площади тела в нем лучше сохраняется тепло. Таким образом, подобно цвету кожи тип телосложения коррелирует с географической широтой, на которой расселена раса, и в прошлом имел селективное значение, т. е. отбор шел на телосложение, т. е. на отношение между длиной конечностей и массой тела (табл. 41).
Далее, выступающий узкий нос у европеоидов удлиняет носоглоточный путь, что способствует нагреванию холодного воздуха и защищает от переохлаждения гортань и легкие. Напротив, очень широкий и плоский нос (большее развитие слизистых) у негроидов способствует большей теплоотдаче. Их вытянутая высокая голова слабее нагревается, а курчавые волосы лучше предохраняют голову от перегрева, т. к. создают воздухоносную прослойку. Естественный отбор тоже прошел на эти признаки, ибо они также обеспечивали приспособление к условиям географической среды обитания.
Дрейф генов способен и зменять генетическую структуру популяций. Подсчитано, что в результате дрейфа генов облик популяций может измениться на протяжении 50 поколений, т. е. около 1250 лет. Поэтому предполагают, что дрейф генов в качестве фактора расогенеза действовал очень медленно.
Таблица 41
Изменчивость отношений между длиной конечностей и массой тела
в отдельных группах и популяциях
| Популяция
|
Количество исследованных индивидов
|
Среднее отношение длины конечностей / вес тела
|
Географическая широта (примерная)
|
| Парижские рабочие
|
73
|
4,88
|
50° сев. ш.
|
| Французские солдаты
|
60
|
4,91
|
50° сев.ш.
|
| Французские студенты
|
8
|
4,94
|
50° сев. ш.
|
| Финны
|
47
|
4,89
|
60° сев. ш.
|
| Британские солдаты
|
300
|
5,0
|
55° сев.ш.
|
| Украинцы
|
504
|
5,08
|
50° сев. ш.
|
| Сицилийцы
|
120
|
5,09
|
40° сев. ш.
|
| Арабы (йеменцы)
|
31
|
5,63
|
20° сев.ш.
|
| Нунгусы (Мозамбик)
|
119
|
5,66
|
15" ю. ш.
|
| Дароды (Сомали)
|
123
|
5,74
|
5' сев. ш.
|
| Гобавины (Сомали)
|
51
|
6,90
|
5° сев. ш.
|
| Антумбы (Мозамбик)
|
26
|
6,06
|
15° ю. ш.
|
| Кораны (Ю. Африка)
|
18
|
8,21
|
25° ю. ш.
|
| Индийцы (Мадрас)
|
35
|
6,41
|
10" ю. ш.
|
| Пигмеи Ака (Конго)
|
95
|
6,98
|
5° ю. ш.
|
| Пигмеи Васуа (Конго)
|
115
|
7,03
|
5° ю. ш.
|
Изоляция проявлялась в различных формах и объемах. В частности, географическая изоляция первобытных коллективов в эпоху палеолита сопровождалась дифференцировкой их генетического состава, прерыванием контактов с другими коллективами. Изоляция с помощью искусственных барьеров тоже оказывала влияние на генетическую дифференциацию популяций. Однако она влияла и на концентрацию культурных традиций. Например, изоляция в США индейцев от белых, а ирландцев и евреев от других национальных групп американцев вела и ведет к формированию обособленных расовых свойств. В популяциях индейцев племени хопи, проживающих на юго-западе США, всегда изолировали мужчин-альбиносов, оставляя их в жилищах на период полевых работ. Однако это сопровождалось большей свободой в половых связях с женщинами племени, поскольку мужчины-неальбиносы работали в поле, что приводило к повышению частоты рождения детей-альбиносов, к изменению генетической конституции племени. Следовательно, результатом изоляции была селекция носителей генов альбинизма.
Смешение популяций вело к смешению генотипов их членов, к формированию новых рас. Например, северо-американская цветная раса (негритянское население США) возникла в результате смешения лесной негроидной расы с расами банту, а также с северо-западной европейской, альпийской, средиземноморской и, возможно, с другими расами. Южноафриканская цветная раса возникла в результате смешения банту, бушменов и готтентотов. В настоящее время на Гавайских островах формируется новая расовая группа на основе метисации европеоидов, монголоидов и полинезийцев.
Будущее рас определяется рядом действующих в наше время факторов. Продолжается рост численности населения земного шара, увеличивается миграция, повышается частота межрасовых браков. Предполагают, что в результате подвижности популяций человека и межрасовых браков в будущем начнет формироваться единая раса человечества. В то же время предполагают и другое возможное последствие межрасовых браков, связанное с формированием новых популяций с собственными специфическими сочетаниями генов.
Обсуждая проблему расогенеза, следует остановиться на расизме. Как в прошлом, так и в настоящее время основу расизма составляют извращенные представления о природе человека в результате преувеличения роли биологических факторов в его индивидуальном и историческом развитии.
Расизм зародился еще в рабовладельческом строе, однако основные расистские теории, которые положили начало расизму в современном понимании, были сформулированы в прошлом веке. В этих теориях обосновывались либо преимущества одних рас над другими в пределах европейских рас, либо превосходство белых над черными, либо пропагандировался сионизм. Без каких-либо научных обоснований расисты считают, что расы являются самостоятельными видами, происходящими от разных обезьяноподобных предков, либо признают общность происхождения рас, но утверждают, что расы различаются по скорости и уровню развития. Расисты делят расы на «низшие» и «высшие», полагая, что высокие цивилизации якобы созданы «высшими» расами.
В XIX в. в Европе возник культ арийской расы, который берет начало от санскритской легенды о светлых покорителях темнокожих индусов и персов. В 1854 г. в качестве «высшей» расы французский социолог Ж. Гобино назвал «германскую» расу, выступив одновременно против браков представителей «высших» и «низших» рас.
В последующем расизм оказался связанным с евгеникой и социал-дарвинизмом. Некоторые положения евгеники оказались привлекательными для расистов в качестве орудия улучшения расовых «достоинств». Что касается социал-дарвинизма, то он оказался выгодным для расистов по той причине, что открывал возможность рассматривать общественные явления на биологической основе. В начале нашего века для обоснования расового превосходства стали использовать данные генетики. Расисты стали утверждать о превосходстве в США белых над неграми и о биологической неполноценности негров, о вредности межрасовых браков.
В 20-е гг. XX в. в расизме появилось стремление найти автономные генетические факторы, детерминирующие признаки «высшей» расы, а также связать расизм с необходимостью геноцида. Это стремление привело несколько позднее к объединению расизма с фашизмом. В фашистской Германии «расоведение» было возведено в ранг государственной политики с геноцидом на оккупированных территориях.
Современный расизм — это смесь старых расистских и евгенических идей, а также их «доказательств» с современными био-логизаторскими концепциями человека. Характерная особенность современного расизма заключается в том, что из расистского словаря совершенно исчезли старые доводы в пользу большей чувствительности «низших» рас к болезням. Напротив, появились доводы в пользу превосходящих умственных способностей «высших» рас.
В научном плане расизм не выдерживает критики. Используемое для обоснования существования «высших» рас понятие о «чистых» расах, представители которых якобы гомозиготны по всем локусам (имеют одинаковые аллели на каждом генетическом локу-се), — это расистское понятие. Генетической идентичности нет не только в случае рас, но даже в отдельных семьях. Полная гомози-готность существует только в случае однояйцевых близнецов. Нет также никаких оснований считать, что чистые расы существовали в прошлом, ибо в пользу этого допущения нет ни одного доказательства. Напротив, вся эволюционная история человечества — это непрерывное смешение рас. Таким образом, утверждение о том, что раса является хорошо дифференцированной генетической группой, неверно.
Расисты утверждают, что «чистые» расы (которых фактически нет) обладают лишь «хорошими» генами, в том числе «хорошо» детерминирующими психические свойства. Между тем наука не располагает данными о связи между признаками, на основе которых идентифицируют расу, и психическими свойствами индивидуумов. Ни одна из рас не обладает монопольно «хорошими» генами и не является полностью свободной от «плохих» генов. Гены, которые не желательны в одних географических и культурных условиях, могут быть желательными в других условиях. Наконец, многие из так называемых «превосходящих» и «отсталых» характеристик, иногда связанных с определенными расовыми группами, могут быть в основном или полностью результатом воздействия среды. Никто и никогда не показывал существования особых генов, контролирующих психическую деятельность людей, принадлежащих к той или иной расе. В формировании интеллекта, видимо, участвуют многие факторы. Каждый индивидуум должен оцениваться на основе его собственных характеристик, а не характеристик расы, к которой он принадлежит. Изучая местное население Новой Гвинеи, Микронезии, Меланезии, Филиппин и других районов земного шара, русский ученый Н. Н. Миклухо-Маклай (1846-1888) еще в конце XIX в. доказал, что аборигены разной расовой принадлежности ничем не отличаются по умственным способностям от европеоидов. Поэтому утверждения о различиях между расами по умственным способностям основаны на извращениях понятий равенства и неравенства, идентичности и разнообразия, недооценке роли среды. Равенство и неравенство — это понятия социальные и этические, тогда как идентичность и разнообразие — это понятия, применимые к характеристикам генотипов людей. Любой индивидуум после рождения оказывается в определенной среде. Поскольку гены появляются в одной среде и не проявляются в другой, то с учетом нормы реакции это означает, что не все гены, контролирующие физические свойства человека, могут проявляться в условиях той или иной среды, и что сходные гены проявляются в разных сре-довых условиях. Но в отличие от среды животных среда человека является средой социальной. Благодаря сознанию он действует в сфере общественной формы движения материи, подчиняет себе среду обитания и, в конечном итоге создает надбиологическую сферу. Гены интеллектуальности, если они есть, не обладают особенностями, позволяющими им определить верхние или нижние границы в развитии духовного облика индивидуума, группы индивидуумов, расы. Разные особенности в проявлении духовного потенциала расы определяется не генетическими недостатками или дефицитами, а генетическим разнообразием, которое складывается на основе генетических механизмов и проявляется в зависимости от разнообразия социальной среды.
Любой индивид независимо от расовой принадлежности развивается по генетической программе в результате взаимодействия с социальной средой. Следовательно, в онтогенезе людей индивид любой расы есть продукт собственной наследственности и социальной среды, тогда как в филогенезе человек является продуктом общественного развития, а расы являются результатом действия генетических механизмов лишь на основе полиморфизма человека (цвет кожи, группа крови, строение наружного уха, форма носа, структура волос и т. д.). Следовательно, расовые различия касаются чисто физических признаков, не имеющих отношения к чисто человеческим признакам, т. е. расы не имеют различий по чисто человеческим признакам. Для каждой расы характерно генетическое разнообразие, которое создает потенциал расы. Между расами есть различия лишь в общественно-экономическом развитии, однако эти различия не основываются на биологических закономерностях. Они являются результатом исторического развития, что было тоже показано Н. Н. Миклухо-Маклаем на основе изучения аборигенов Новой Гвинеи, Микронезии, Меланезии и других районов земного шара.
Морфологические и физиологические особенности человека, т. е. его конституция и функциональная активность давно представляют всеобщий интерес, который восходит к далекому прошлому. Еще великий врач древности Гиппократ различал сильную, плотную, влажную и жировую конституцию человека, причем считал, что люди разных конституциальных типов склонны и к разным болезням.
Позднее Клавдий Гален (130—200) выделял четыре конституциальных типа людей, связывая каждый тип с определенным характером движения «соков» в организме. В частности, он различал такие «соки» как sangua — (кровь), phlegma (холодная слизь), chole (желчь), melan chole (черная желчь). Преобладание в организме того или иного «сока» определяло тип темперамента людей. Поэтому по типу темперамента Гален различал сангвиников (веселых, подвижных, постоянно стремящихся к переменам), флегматиков (медлительных, сдержанных, сохраняющих спокойствие в любой ситуации, но инертных), холериков (напористых, энергичных, но гневных и пристрастных) и меланхоликов (раздражительных, нерешительных, но способных к большой системности в работе). Такой классификации конституциональных типов придерживался и И. П. Павлов.
В начале XX в. французский врач Сиго на основе учета характера питания, дыхания, движения и нервных реакций предложил выделять четыре конституциональных типа — церебральный, ди-гестивный, мышечный и респираторный.
К церебральному типу он относил людей с относительно крупной головой и удлиненной грудной клеткой. К дигестивному типу были отнесены люди с большим животом, короткой и широкой грудной клеткой, развитыми челюстями. К мышечному типу были отнесены люди с развитой мускулатурой, широкими плечами, длинными конечностями. Наконец, респираторный тип по этой классификации был представлен людьми с развитой грудной клеткой и носовой полостью, с длинной грудной клеткой и длинными конечностями.
В 20—30 гг. нашего века немецкий врач Кречмер определил три конституциональных типа — астенический, пикнический и атлетический. Астенический тип — люди с утолщенной грудной клеткой, узкими плечами, отсутствием жировых отложений, резкими переменами настроения. Пикнический тип — люди с большими размерами головы, груди и живота, плотной фигурой, склонностью к ожирению, к плавной смене настроения. Атлетический тип представлен людьми, для которых характерны массивный грудной отдел скелета, широкие плечи, хорошо развитые мышцы, медленная (постепенная) смена настроения.
Выделение конституциональных типов людей не потеряло своего значения и в наше время. В зависимости от целей типизации используют каждую из названных выше классификаций. С генетической точки зрения можно сказать, что конституциональный тип того или иного индивида есть категория фенотипическая, являющаяся результатом взаимодействия генотипа и среды.
Однако человечество чрезвычайно расселено по земному шару, занимая районы, различающиеся по климату, ландшафту, геохимическим и другим особенностям. Между тем действие климатических особенностей, а также влияние гравитации, электромагнитного поля, радиации, патогенных организмов и других факторов сопровождается географической изменчивостью морфологических и физиологических свойств людей. Приуроченность этих свойств к определенным территориям свидетельствует о географической (экологической) изменчивости современного человека. В контексте этой изменчивости различают арктические, высокогорные и тропические группы людей, а также группы людей, проживающих в условиях умеренного климата.
Население арктических групп (эскимосы, чукчи и др.) представлено людьми в основном мускульного типа с повышенной массой тела и цилиндрической грудной клеткой. Все члены этих групп характеризуются также повышенным уровнем основного обмена, поглощения кислорода, энергетических процессов. У индивидов большинства арктических групп отмечается высокое содержание холестерина в крови. Однако жители континентальных районов Сибири, по сравнению с аборигенами Арктики, чаще принадлежат к астеническому и пикническому типам телосложения. Они характеризуются относительной коротконогостью и длиннорукостью, более плоской грудной клеткой, увеличением жирового компонента тела. Для них характерна более высокая, по сравнению с жителями умеренных районов, теплопродукция, но одинаковый уровень холестерина в крови.
Люди высокогорных групп (горцы Кавказа, Памира и Тяныпаня, коренные жители Эфиопии и Индии, индейцы Перу и др.) характеризуются увеличенной емкостью грудной клетки и увеличением костно-мускульной массы тела. Для них характерен повышенный уровень эритроцитов (гемоглобина) и иммуноглобулинов, но пониженный уровень холестерина.
Люди тропических групп (коренные жители Африки, Австралии, Океании, Индии и Америки) характеризуются удлиненной формой тела, недостаточно развитой мускулатурой, повышенны!,? количеством потовых желез (на 1 см2
тела), усиленной теплоотдачей и пониженным уровнем энергетических процессов. Кроме того, для них характерны повышенный уровень иммуноглобулинов и пониженный уровень холестерина в крови. У коренных жителе!-! тропических широт найден белок трансферин, регулирующий температурный режим тела. Для коренного населения пустынь характерны высокорослый тип тела, более низкое кровяное давление. повышенное содержание эритроцитов в крови.
Население зон умеренного климата по морфологическим и. функциональным свойствам занимает среднее положение между жителями арктических и тропических групп. Жители умеренных зон. подвержены влиянию химических свойств почвы, воды и высоты над уровнем моря. Например, минерализация их скелета зависит от содержания макро- и микроэлементов в почве и воде.
На основе зональной зависимости морфофункциональной изменчивости разных популяций человека предполагают существование адаптивных типов, которые независимы ни от расовой, ни от этнической принадлежности и определяются нормой реакции, обеспечивающей равновесие популяций со средой. Адаптация человека к среде связана с изменением его морфологических и физиологических свойств. Поэтому одинаковые черты приспособленности к условиям тропических зон характерны как для коренных жителей Африки (негроидов), так и для европеоидов Индии, австралийцев. Единые черты приспособленности характерны также для жителей Крайнего Севера (ненцы, чукчи, эскимосы, саами).
Адаптивноеть человека имеет исторический характер. Предполагают, что у австралопитеков приспособительные реакции заключались в адаптации их к климату тропической зоны, а у архантро-пов эти реакции развились в направлении формирования приспособленности к влажному тропическому и высокогорному климатам. Расселение палеантропов в Европе сопровождалось формированием адаптивного типа умеренного пояса (эпоха среднего палеолита). Арктические адаптивные типы возникли, вероятно; в эпоху верхнего палеолита.
Считают, что физический тип человека не изменился за послед ние 35--40 тыс лет. Почти не изменился и интеллект человеке. Однако экологические факторы сейчас воздействуют на человека больше, чем даже в прошлом веке. Поэтому современной тенденцией физического облика человека сейчас стали акселерация и се-кулярный тренд.
Акселерация (от лат. acceleratio) — это ускорение роста людей и проявления их физиологических функций. Термин предложен в 1935 г. немецким врачом Е. Кохом. Примеры акселерации многочисленны.
Так, в начале века длина тела у мужчин достигала своих обычных размеров к 25-26 годам, в настоящее время — к 18-19 годам. Начало менструального цикла за последние годы сократилось с 14,5 лет до 12,5 лет. По обобщенным данным в развитых странах вес при рождении увеличился на 100—300 граммов. Половое созревание подростков происходит на 2 года раньше.
Объяснения причин акселерации весьма противоречивы. Одни специалисты считают, что основу акселерации составляют улучшение условий жизни и повышение уровня медицинского обслуживания населения. Другие же считают, что акселерации способствует появление новых сочетаний генов. Ни одно, ни другое из этих объяснений не является убедительным. Природа акселерации остается невыясненной, но ясно, что акселерация имеет и негативные особенности. Например, среди современного населения повысилась частота близорукости, кариеса, различных неврозов и т. д.
Секулярный тренд (от лат. secular trend — вековая тенденция) — это увеличение длины тела, репродуктивного периода, продолжительности жизни и других важнейших свойств человека в определенные (длительные) интервалы времени. Например, в нашей стране было отмечено увеличение длины тела на 3,5 см у всех родившихся в 1920-1935 гг. по сравнению с прошлым столетием. Как и природа акселерации, природа секулярного тренда также не имеет удовлетворительного объяснения.
В археологии четвертичный период подразделяют по-другому в сравнении со шкалой геологического времени. Археологи в пределах четвертичного периода вместо эпох выделяют века, в частности древнекаменный (палеолит), среднекаменный (мезолит) и новокаменный (неолит), а также еще более поздние другие века, приуроченные к современному человеку и его культуре. Между геологическим и археологическим временем имеется некоторое совпадение. В целом палеолит соответствует плейстоцену, а мезолит и неолит — периоду после плейстоцена.
Данные археологии свидетельствуют о том, что появление предков современного человека сопровождалось формированием начал культуры. Биологическая эволюция человека заняла около 14 млн лет, тогда как начало его культурного развития датируется временем порядка 2—3 млн лет от наших дней, т. е. четвертичным периодом. Тем не менее биологическое развитие и культурное развитие человека взаимосвязаны. Биологическое развитие оказывало влияние на культурное развитие и наоборот. Культурное развитие влияло на физическое и психическое развитие человека и было очень большим с самых первых шагов культуры, но на последних этапах эволюции человека оно приобрело решающее значение.
Начало культурного развития человека связано с его орудийной деятельностью. Рассматривая первые результаты орудийной деятельности человека (каменные и костяные орудия), нельзя не заметить, что в разных географических районах она характеризовалась разным уровнем обработки. Это объясняют обычно географическим местом обитания популяций, влиянием среды их обитания и степенью изолированности. Умение в изготовлении орудий и качество изготавливаемых орудий послужили основанием для квалификации их в качестве культур.
Характерная особенность этих культур заключается в их сменяемости. Но нельзя не отметить, что сменяемость отдельных из них задерживалась на очень долгое время по ходу истории человечества. Например, уровень культур некоторых аборигенов на африканском и австралийском континентах остается и сейчас почти на уровне каменного века.
Считают, что самые первые простые орудия из камня и кости были изготовлены Н. habilis, которому были известны орудия как из каменных отщепов (рубил), так и из крупных галек (олдован-ская культура). Для Н. erectus культура отщепов и культура ручных рубил также были хорошо известны в раннем палеолите, когда шло непрерывное совершенствование орудий, возникали стоянки.
В нижнем палеолите каменные орудия (ручные рубила) характеризовались лучшей обработкой, о чем свидетельствуют находки в пещере Азых (Азербайджан). Этот тип культуры известен под названием ашельской и шельской, которые значительно выше ол-дованской культуры.
Развитие культур нарастало в среднем палеолите. Тогда неандертальцами была создана так называемая мустьерская культура (на территории современной Франции), которая характеризовалась более высоким уровнем изготовления каменных орудий. Одновременно появляются орудия и из кости (остроконечники, скребки и др.). Мустьерская культура длилась примерно 100 тыс лет.
Особенно бурное развитие культуры приходится на поздний палеолит (35-40 тыс лет назад). Увеличение числа стоянок и жителей в них вело к совершенствованию методов охоты, к дифференциации каменного инвентаря. Появляются инструменты для изготовления орудий.
В конце последнего ледникового периода неандертальцами были разработаны довольно совершенные методы выделки кожи, появились начала искусства. В верхнем палеолите искусство поднимается на новый уровень, о чем свидетельствуют образцы пещерной живописи (изображения животных и различных сцен на стенах пещер), найденные в Африке и Европе.
К тому времени относятся первые погребения и начало формирования ритуальных церемоний.
Для древнейших и древних людей характерен каннибализм и ритуальный характер каннибализма. Считают, что неандертальцы имели представления о посмертной жизни. Главным оружием человека в мезолите была рогатина (пика) из тисса с острым каменным наконечником. Достигли значительного совершенства способы охоты, рыболовства. Неандертальцы и ранние кроманьонцы уничтожали мамонтов, бизонов, мегатериев. Появляются лук и стрелы. Стали строиться жилища.
В неолитическом веке человек стал уходить от стадного образа жизни. Этим временем датируется изобретение керамики, переход к оседлому образу жизни, образование дифференцированных общественных ячеек. От собирания и охоты он стал переходить к сельскому хозяйству (выращивание растений и разведение животных). На европейском континенте начала сельского хозяйства, сопровождаемые доместикацией растений и животных, а также неолитической технологией, восходят к 9-6 тысячелетиям до нашей эры.
Каждая популяция людей вносила свой вклад в создание культур на ранних этапах цивилизации. Последующие поколения людей создавали более совершенные цивилизации. Благодаря распределению работы между разными социальными группами, созданию новых инструментов и технологий, доместикации животных и растений, а также развитию экономических, социальных и политических институтов люди современного физического типа создали чрезвычайно высокую цивилизацию, передаваемую от одних поколений к другим посредством языка (речи), письма, печати, а в новое время посредством телефона, радио, телевидения и компьютерной техники. Каждая социальная группа стала действовать как часть экологического сообщества людей. В результате объединения и взаимодействия люди стали производить, хранить и транспортировать пищу в больших количествах, строить железные и шоссейные дороги, производить самолеты и космическую технику, запускать искусственные спутники Земли, обеспечивать общественный порядок, санитарное благополучие и т. д. Адаптация, одежда и жилища позволили развиваться жизни человека в самых различных климатических условиях.
В заключение важно еще раз подчеркнуть, что биологическое и культурное развитие человека шло синхронно. Тем не менее в развитии региональных сообществ имеются различия. Одни сообщества людей ушли далеко вперед, другие отстали в своем развитии. Эти различия еще не получили достаточного объяснения, хотя в числе причин, обеспечивавших отсталость отдельных народов, часто называют сверхиспользование природных ресурсов или исходный недостаток природных ресурсов, тяжелые климатические условия, войны, а также недостаточность регулирующих механизмов развития. Возможны и другие причины, но их обсуждение выходит за рамки данного учебника.
1. Когда стали интересоваться происхождением человека? Почему проблемы антропогенеза и расогенеза привлекают такое большое внимание сейчас?
2. В чем заключается концепция животного происхождения человека? Имеет ли она доказательства? Известны ли другие (альтернативные) концепции?
3. Назовите свойства, по которым человека отличают от животных. Могут ли современные человекообразные обезьяны эволюционировать в сторону человека?
4. Каково значение ископаемых останков в изучении антропогенеза? Какова роль других биологических наук в изучении антропогенеза?
б. Назовите основные этапы антропогенеза. Можно ли построить эволюционные ряды в применении к человеку?
6. Какое значение в эволюции человека имело появление прямохож-дения и развитие головного мозга?
7. В чем заключаются особенности Н. habilis, каков его возраст и можно ли считать его эволюционной линией на пути к Н. sapiens?
8. Перечислите факторы антропогенеза, обратив внимание на специфические.
9. Когда и как естественный отбор действовал в эволюции человека? Действует ли сейчас естественный отбор в популяциях человека?
10. Какова роль труда в антропогенезе?
11. Как вы понимаете биологическую и социальную сущность человека?
12. В чем заключается ошибочность типологического подхода к определению рас? Приведите научное определение расы. Какова роль по-пуляционного подхода к научному определению расы?
13. Есть ли разница между терминами «вид» и «раса»? Если имеется, то покажите ее на двух-трех примерах.
14. Приведите существующие классификации рас. Назовите основные расы рода человеческого. Что вкладывают в понятия «старые» и «новые» расы? С какой скоростью происходит расогенез?
16. Перечислите факторы расогенеза. В чем заключается различие в действии естественного отбора при видообразовании и расогенезе?
16. Существуют ли «чистые» расы? В чем заключается научная несостоятельность расизма?
17. Что понимают под конституциональными типами людей? Какие вы знаете классификации конституциональных типов и насколько они совершенны?
18. Что вы понимаете под географической изменчивостью человека? Есть ли разница между географической и экологической изменчивостью человека?
19. Какие вы знаете группы людей, возникшие в результате экологической изменчивости?
20. Дайте определение понятиям «акселерация» и «секулярный тренд» и приведите соответствующие примеры.
21. Не угаснет ли в будущем вид Н. sapiens подобно тому, как в ходе эволюции угасли многие виды других живых существ?
Алексеев В. П. Человек (эволюция и таксономия). М.: Наука. 1985. 285 стр.
Дубинин Н. П. Что такое человек. М.: Мысль. 1983. 319 стр.
Елинек Я. Большой иллюстрированный атлас первобытного человека. Прага: Артия. 1985. 569 стр.
Рогинский Я. Я. Проблемы антропогенеза. М.: Высшая школа. 1977. 261 стр.
Чебоксаров Н. Н., Чебоксарова М.А. Народы, расы, культуры. М.: Наука. 1985.
Gibson К. R., Ingold T. (Ed.) Tools, Language and Cognition in Human Evolution. Cambridge University Press. 1995. 482 p.
Marks J. Human Biodiversity: Genes, Races and History. Aldine de Qruyter. 1995. 321 p.
Способность живых существ к жизни в различных условиях является результатом эволюции не только их самих, но и их систем органов. Ниже мы рассмотрим это заключение на примерах эволюции систем органов животных, начиная с простейших и заканчивая млекопитающими.
Внешние покровы животных (кожа или интегумент) выполняют защитную функцию, предохраняют тело от излишней потери воды и помогают регулировать температуру тела. Кроме того, у ряда организмов они являются дополнительным органом, принимающим участие в обмене веществ (дыхании и выделении).
У беспозвоночных покровы тела не достигают значительного развития, будучи представленными эктодермой и ее производными. Эволюция покровов у них шла в направлении развития мерцательного эпителия в плоский эпителий, что отмечается у турбелля-рий и других плоских червей, а также у круглых и кольчатых червей. Мерцательный эпителий служит примитивным органом движения, тогда как плоские этой способностью не обладают.
У членистоногих поверхностный слой эпителия превращается в хитинизированную кутикулу, которая развивается в панцирь у ракообразных. У моллюсков поверхностный слой эпителия развивается в раковину, содержащую известь. У животных отдельных групп (кольчатые черви, членистоногие и моллюски) в эпидермисе локализованы одно- или многоклеточные железы (слюнные, паутинные, ядовитые и другие).
У хордовых эволюция кожных покровов шла в направлении замены однослойного эпителия многослойным, в частности, в формировании двух слоев и в превалирующем развитии собственно кожи (кориума).
Кожа хордовых состоит из поверхностного слоя эктодермаль-ного происхождения, или эпидермиса, иногда называемого кутикулой, и нижнего слоя мезодермального происхождения — собственно кожи (кориума), или дермы. У туникат (асцидий, пиросом и других морских животных) наружний покров представляет собой студенистую или хрящевую оболочку, являющуюся продуктом эк-тодермального эпителия и пронизанную мезодермальными клетками и сосудами. Покровы ланцетников имеют двуслойное строение. Первый слой представлен цилиндрическим однослойным эпителием эктодермального происхождения, второй — студенистой соединительной тканью, выполняющей роль кориума.
Покровы у позвоночных достигли большого развития и представляют собой кожу, которая построена из двух различных слоев. Один из них является поверхностным и состоит из многослойного эпителиального эпидермиса эктодермального происхождения, второй — нижним, собственно кожей, которая прилегает к подкожной ткани. Последняя, в свою очередь, прилегает к мышцам и костям.
Поскольку у земноводных кожа используется в качестве дополнительного органа дыхания, но при этом необходима водная пленка, то это привело к развитию в коже этих животных многослойных желез, продуцирующих слизь и увлажняющих ее. Кожа земноводных содержит также ядовитые железы, выполняющие защитную функцию.
У млекопитающих развились производные кожи (поверхностный каратин, волосы, копыта и рога), которые выполняют важные функции в жизни организмов. Большое значение в терморегуляции имеют кожные потовые железы.
Скелет выполняет опорную и защитную функции. У беспозвоночных скелетом являются ригидные структуры, имеющие самый разнообразный характер и подразделяемые на эндос-келет и экзоскелет (рис. 185).
Эндоскелет построен из органических фибрилл или кристаллов неорганических соединений. У простейших эндоскелетной структурой является аксостиль ряда жгутиковых, который имеет фиб-риллярную структуру, у радиолярий и фероминифер эндоскелет представлен кристаллами неорганического происхождения, у губок — спикулами (иглами, состоящими из отложений карбоната  кальция или силикона вокруг клеточных фибрилл), у морских звезд — различными хрящеподобными тканями. Некоторые членистоногие имеют хорошо развитый эндоскелет, к которому прикрепляются мышцы. Известковые пластинки и иглы иг-локожих имеют мезодермаль-ное происхождение. кальция или силикона вокруг клеточных фибрилл), у морских звезд — различными хрящеподобными тканями. Некоторые членистоногие имеют хорошо развитый эндоскелет, к которому прикрепляются мышцы. Известковые пластинки и иглы иг-локожих имеют мезодермаль-ное происхождение.
Экзоскелет имеется у многих беспозвоночных организмов и характеризуется чрезвычайным разнообразием, начиная от пелликулы (утолщенной клеточной стенки) и заканчивая такими образованиями, как крылья бабочек. Импрегнация эк-зоскелета неорганическими солями повышает его ригидность. Гибкость экзоскелета, которая необходима для его мобильности, достигается у многих нематод вследствие гибкости кутикулы, а у коловраток и особенно у членистоногих благодаря наличию гибких районов между ригидными участками кутикулы. У членистоногих кутикула является результатом секреции клеточного эпидермиса и состоит из тонкой наружной эпикутикулы и толстой эндокутикулы, в состав которой входят хитин и белок.
У низших хордовых развит осевой скелет, которым является хорда. У бесчерепных эндоскелетом является хорда, а экзоскеле-том — стержневидные образования, состоящие из плотного студенистого вещества (непарные плавники и опора жаберного аппарата). У кругл оротых хорда также сохраняется на протяжении всего онтогенеза. Однако у них под хордой уже появляются парные закладки позвонков в виде хрящевидных образований (верхние дуги). У беспозвоночных кальциевый скелет представлен кальцитом.
У позвоночных эндоскелет достигает высокого развития, являясь хрящевым и (или) костным скелетом. У этих организмов различают осевой скелет, скелет головы (черепная коробка и висцеральный скелет) и скелет конечностей. Экзоскелет представлен волосами, перьями, чешуей, рогами, отростками рогов, копытами, когтями, косточками в коже.
У примитивных рыб наряду с верхними дугами появляются закладки низших дуг. У высших рыб из тканей, окружающих хорду, и оснований верхних и низших дуг, развиваются позвонки. Позвоночный столб состоит из туловищного и хвостового отделов. Благодаря сращению концов верхних дуг образуется канал для спинного мозга. К низшим дугам прикрепляются ребра.
У земноводных процесс замены хорды позвонками углубляется еще в большей степени и происходит в ранние периоды развития. Развиваются четыре отдела позвоночника — шейный, грудной, крестцовый и хвостовой. Шейный отдел очень короток (один позвонок), грудной отдел насчитывает пять позвонков, к которым прикрепляются свободно заканчивающиеся ребра (не доходящие до грудины), крестцовый отдел также состоит из одного позвонка, к которому прикрепляются кости таза. Хвост хвостатых амфибий состоит из нескольких позвонков, бесхвостых — из одного позвонка, называемого уростилем.
У амниот (рептилий, птиц и млекопитающих) различают осевой скелет, скелет головы (череп) и скелет конечностей. Осевой скелет состоит из шейного, грудного, поясничного, крестцового и хвостового отделов.
Количество шейных позвонков варьирует от 1 (у современных земноводных) до 76 (у ископаемых форм — морских плезиозавров). У мниот и лабиринтодонтов первые два шейных позвонка специализированны, обеспечивая поддержание и движение головы. Количество шейных позвонков зависит от длины шеи. У некоторых рептилий их количество достигает 8, у птиц — до 25. Современные млекопитающие имеют 7 шейных позвонков.
Количество грудных позвонков различно у разных групп позвоночных. У черепах 10 грудных позвонков слиты со связками и костями и формируют панцирь, у пресмыкающихся к грудным позвонкам прикрепляются ребра, что формирует грудную клетку. Птицы имеют по 4—6 грудных позвонков. У млекопитающих чаще развивается 10-20 грудных позвонков. У человека грудную клетку формируют также ребра (12 пар) и грудина.
Поясничный отдел развит по-разному. У рептилий поясничный отдел выражен плохо. У птиц поясничные позвонки обычно слиты с грудными, крестцовыми и урокрестцовыми, формируя сложный крестец. У млекопитающих количество поясничных позвонков составляет от 3 до 24 в зависимости от вида.
Количество хвостовых позвонков зависит от длины хвоста. У лягушек, например, хвостовые позвонки сливаются в одиночную кость (уростиль), у птиц имеется 4-6 хвостовых позвонков, которые сливаются также в уростиль. Некоторые ископаемые птицы (археоптерикс) имели длинный хвост, состоящий из 20 позвонков. У млекопитающих количество позвонков в хвосте зависит от вида. Прямохождение человека вело к искривлению позвоночника, т. е. это оказалось «платой» за Прямохождение (рис. 186).
Скелет головы состоит из черепа и висцерального (лицевого) скелета.
Черепная коробка различается не только по форме, но и по количеству входящих в нее костей, которые очень колеблются  в зависимости от принадлежности животных к тому или иному классу. У костных рыб, например, количество костей черепа превышает 100, в то время как у земноводных оно равно 15-19, у пресмыкающихся — от 40 до 70, у птиц — 20, у примитивных млекопитающих (Didelphis) — 48, у шимпанзе — 22, у человека — 27 (рис. 187). Снижение количества костей черепа в эволюции от рыб до млекопитающих является результатом потери индивидуальных костей или их слияния с другими костями. в зависимости от принадлежности животных к тому или иному классу. У костных рыб, например, количество костей черепа превышает 100, в то время как у земноводных оно равно 15-19, у пресмыкающихся — от 40 до 70, у птиц — 20, у примитивных млекопитающих (Didelphis) — 48, у шимпанзе — 22, у человека — 27 (рис. 187). Снижение количества костей черепа в эволюции от рыб до млекопитающих является результатом потери индивидуальных костей или их слияния с другими костями.
Эволюция висцерального скелета шла в направлении его редукции. Он очень уменьшен у четвероногих позвоночных, дышащих легкими.
Первоначальная функция висцеральных дуг заключалась в поддержке жабер. Это подтверждается и наличием жаберных щелей у низших рыб (кроссоптериг, осетровых и др.). У рептилий и птиц первая жаберная дуга развилась в скелет гортани. Жаберная крышка, представлявшая собой костный клапан, покрывающий жабры, и большинство жабер были утеряны наземными позвоночными. Многие кости мозгового черепа, которые разделены у низших четвероногих, слились у млекопитающих. Млекопитающие отличаются от других животных наличием трех небольших костей в полости среднего уха. Одна из этих костей — стремечко — гомологична второй висцеральной дуге рыб, другая — наковальня — гомологична кости нижней челюсти других позвоночных, а третья — молоточек — гомологична кости верхней челюсти других позвоночных.
Скелет конечностей имеет особенности, причем ключевым является вопрос о переходе плавников в конечности. Конечности классифицируют на непарные и парные. Примером непарных конечностей является спинной и хвостовой плавники у рыб, парных — свободные конечности, роль которых выполняют плавники у рыб и пятипалые конечности у наземных животных. Парные плавники кистеперых рыб содержат гомологи плечевой, локтевой и лучевой костей пятипалой конечности наземных позвоночных, что свидетельствует о происхождении пятипалой конечности от плавника кистеперых рыб. Свободные конечности четвероногих обычно состоят из трех сегментов, которые организованы по-разному у разных животных. Главные вариации отмечаются в строении костей плеча и бедра. В частности, плечевая и бедренная кости укорочены у дельфинов и некоторых ископаемых форм (ихтиозавров).
 Существенные колебания отмечаются в количестве костей запястья и плюсны. Некоторые из примитивных ископаемых земноводных имели от 20 до 30 костей в этих частях скелета, но современные земноводные имеют лишь от 3 до 9 в запястье и от 3 до 12 в плюсне. Число костей в каждом пальце также колеблется у разных позвоночных. Фаланговая формула (количество костей от первого пальца до последнего) у современных амфибий, у которых имеется обычно четыре пальца, составляет 2—2-3—2 (саламандры), 2-2—3-3 (лягушки). Ихтиозавры имели от трех до восьми пальцев. Крылья современных птиц имеют по три пальца (пониженное число фаланг). Существенные колебания отмечаются в количестве костей запястья и плюсны. Некоторые из примитивных ископаемых земноводных имели от 20 до 30 костей в этих частях скелета, но современные земноводные имеют лишь от 3 до 9 в запястье и от 3 до 12 в плюсне. Число костей в каждом пальце также колеблется у разных позвоночных. Фаланговая формула (количество костей от первого пальца до последнего) у современных амфибий, у которых имеется обычно четыре пальца, составляет 2—2-3—2 (саламандры), 2-2—3-3 (лягушки). Ихтиозавры имели от трех до восьми пальцев. Крылья современных птиц имеют по три пальца (пониженное число фаланг).
Общее количество костей у человека составляет 206. В ходе эволюции сформировался механизм остеофикации скелета эмбрионов, в котором существенное значение имеют кальций, фосфор, витамины Д и ряд ферментов.
У позвоночных кальциевый скелет представлен не кальцитом, а гидроксиапатитом.
В ходе эволюции существенное развитие приобрела и скелетная мускулатура. Выработался также механизм использования химической энергии пищи для сокращения мышц (20-35% от общего количества энергии) и для использования остальной энергии в качестве тепла, обеспечивающего тепловой режим организма.
У простейших органами пищеварения являются пищеварительные вакуоли. Для этих животных характерно внутриклеточное пищеварение, которое является самым древним.
У кишечнополостных и червей из эмбрионального бластопора развивается ротовое отверстие. У кишечнополостных рот открывается в гастральную полость, которая является полостью первичной кишки и заканчивается слепо (анального отверстия нет). В этой полости наряду с внутриклеточным пищеварением появляется внеклеточное. На более высоких уровнях эволюции внутриклеточное пищеварение теряется полностью, уступая место внеклеточному (полостному) как более прогрессивному способу. Однако внутриклеточное пищеварение все же сохраняется у организмов на всех ступенях развития, но лишь в виде фагоцитоза, выполняющего защитную роль.
У ресничных и плоских червей строение пищеварительной системы еще сохраняет сходство с пищеварительной системой кишеч-нополостных. Анального отверстия нет, поэтому выброс пищеварительных остатков происходит через рот. Но у круглых червей пищеварительная система уже несколько усложняется, поскольку пищеварительная трубка заметно подразделяется на передний, средний и задний отделы, из которых передний и задний отделы имеют эктодермальное происхождение, средний — энтодермальное. Впервые развивается анальное отверстие за счет впячивания эктодермы в каудальную часть тела и соединения ее с полостью первичной кишки. У кольчатых червей пищеварительный тракт уже представлен глоткой, пищеводом, зобом, желудком, средней и задней кишками.
Дальнейшее усложнение пищеварительного тракта продолжается у членистоногих и моллюсков, у которых развиваются ротовые придатки вокруг рта, которые служат для измельчения пищи, в кишечнике развиваются отростки, что увеличивает его поверхность. Развиваются пищеварительные железы, которые являются аналогом печени позвоночных.
У хордовых в ходе эволюции продолжается дифференциация кишечной трубки, причем более глубокая, в результате чего происходит увеличение кишечной поверхности для всасывания продуктов пищеварения. Возникают железы, участвующие в пищеварении. Пищеварительная система всех хордовых в основном имеет энтодермальное происхождение (кроме ротового отдела и заднего кишечника, имеющих эктодермальное происхождение). У позвоночных кишечная трубка дифференцируется на ротовую полость, глотку, пищевод, желудок, тонкий и толстый кишечники, которые, однако, не у всех организмов (классов) дифференцированы полностью. Начиная с круглоротых, у позвоночных развивается печень, а начиная с рыб — поджелудочная железа.
У рыб развиваются костные пластинки и зубы. У земноводных впервые появляются слюнные железы, заканчивается обособление желудка, тонкой и толстой кишок, клоаки, развиваются однорядные зубы. Очень интенсивно развивается язык. У рептилий намечается некоторая дифференцировка зубов, часть слюнных желез преобразуется в ядовитые, углубляется обособление желудка, в кишечнике развивается зачаток слепой кишки. У птиц способность к полету сопровождается изменениями в пищеварительной системе. Вместо челюстей и зубов у них появляется клюв, а в пищеводе образуется растяжимое расширение, называемое  зобом. Желудок разделен на железистый и мускульный отделы, в которых происходит хжлическая и механическая переработка пищи (соответственно). Увеличивается в длину толстый отдел кишечника, но редуцируется язык. зобом. Желудок разделен на железистый и мускульный отделы, в которых происходит хжлическая и механическая переработка пищи (соответственно). Увеличивается в длину толстый отдел кишечника, но редуцируется язык.
У млекопитающих развитие пищеварительной системы достигает верхнего предела. Имея значительную длину, она характеризуется большей расчлененностью. Дифференцируются зубы, развивается пищевод. Желудок, состоит из нескольких слоев и содержит много пищеварительных желез. У некоторых травоядных (жвачных копытных) желудок является многокамерным. Кишечник дифференцирован на тонкий, толстый и прямой отделы. Значительной дифференцировки достигают печень, поджелудочная железа, их протоки впадают в передний отдел тонкого кишечника. Происходит удлинение толстого кишечника, появляются слепая кишка, и аппендикс. У многих млекопитающих получила значительное развитие слепая кишка, достигая '/g длины всего кишечника. Толстая кишка, заканчивается анусом, отделенным от мочеполового отверстия промежностью. У яйцекладущих толстая кишка заканчивается клоакой.
У человека в состав пищеварительной системы входит рот, глотка, пищевод, желудок, кишечник, слюнные железы, печень, желчный пузырь и панкреатическая железа (рис. 188). Все отделы же-лудочно-кишечного тракта построены из четырех слоев.
Человек является всеядным существом, поскольку его пищеварительная система обеспечивает метаболизирование как растительной, так и животной пищи (табл. 42).
Главными углеводами, содержащимися в растительной пище, являются крахмал и целлюлоза, а в жквотной пище —гликоген.
Таблица 42
Функции отделов пищеварительной системы человека
| Отделы пищеварительной системы
|
Ротовая полость
|
Пищевод
|
Желудок
|
Тонкий кишечник
|
Толстый кишечник
|
| Основные функции
|
Поступление пищи, частичное переваривание крахмала
|
Переход пищи в
желудок
|
Переваривание белков
|
Переваривание и всасывание
|
Реадсорб-пия солей и воды, формирование каловых масс
|
| Ферменты
|
Амилаза
|
Пепсин
|
Трипсин, липаза
|
| Источники
|
Слюнные
железы
|
Желудочные железы
|
Панкреас, тонкий кишечник
|
| Образуемые
|
Сложные сахара
|
Крупномолекулярные фрагменты белков, некоторые аминокислоты
|
Жирные кислоты, глице-рол, простые сахара, аминокислоты, фрагменты белков
|
Переваривание углеводов начинается в полости рта, где слюнная амилаза, (птиалин) действует на крахмал и гликоген, превращая их в смесь Д-глюкозы, мальтозы и олигосахаридов. Переваривание углеводов завершается в тонком кишечнике, где переваренный под действием птиалина крахмал подвергается дальнейшему воздействию панкреатической амилазы, конвертируется в мальтозу. В конечном итоге вся мальтоза под действием маль-тазы конвертируется в глюкозу.
Все реакции происходят с участием воды (гидролиз) и ферментов:
Рот, желудок:
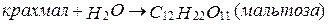
Тонкий кишечник:
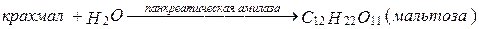

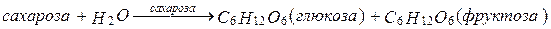
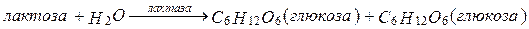
Непереваренная целлюлоза способствует нормальной перистальтике кишечника у млекопитающих.
Переваривание жиров начинается в тонком отделе кишечника, где панкреатическая липаза с помощью желчных кислот и белка колипазы конвертирует жиры в глицерол и жирные кислоты. Одна молекула жира дает одну молекулу глицерола и три молекулы жирной кислоты:

Перевариванию липидов в тонком отделе кишечника способствуют соли желчных кислот, которые являются производными холевой кислоты. Эти соли поступают из печени в желчь, которая затем изливается в переднюю часть тонкого кишечника.
Белки начинают перевариваться (гидролизуются) в желудке под воздействием пепсина и соляной кислоты, выделение которой клетками желудка происходит под действием гормона — гастри-на. Образованные продукты подвергаются повторному гидролизу трипсином и хемотрипсином, предшественники которых вырабатываются в клетках поджелудочной железы, до полипептидов, после чего последние гидролизуются пептидазами до аминокислот. Однако в желуд очно-кишечном тракте человека не все белки перевариваются полностью. Например, такой белок, как кератин, переваривается лишь частично. Многие белки злаковых также перевариваются не полностью, поскольку они покрыты непереваривающейся целлюлозой.
В ходе эволюции у организмов развилась высокоорганизованная система органов дыхания.
У многих беспозвоночных животных, например, у кишечно-полостных, плоских и круглых червей еще нет специальных органов дыхания. У них газообмен со средой осуществляется всей поверхностью тела (рис. 189). Впервые органы дыхания встречаются у морских кольчатых червей и водных членистоногих в виде пористых жабер, располагающихся по обеим сторонам тела и обильно снабжаемыми кровью. У моллюсков в мантийной полости развиваются пластинчатые жабры. Однако уже у наземных членистоногих в углублениях тела имеются трахеи или листовидные легкие.
У водных хордовых дыхательная система связана с кишечником, причем наиболее просто она организована у ланцетника, у которого стенка глотки (передний отдел кишечника) пронизана жаберными щелями, пронизывающими весь передний отдел кишечника и открывающимися в околожаберную полость. У наземных хордовых жаберные щели развиваются лишь в эмбриональный период, после чего исчезают. Дыхание у них осуществляется легкими, которые развиваются из выпячиваний кишечной стенки.
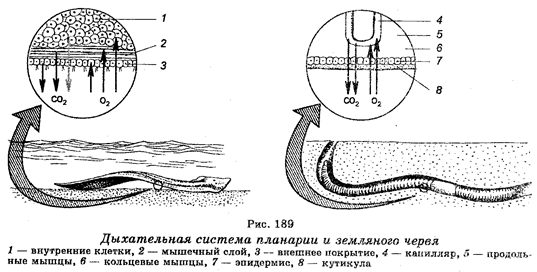
Жаберный аппарат у хордовых эволюционировал в направлении образования жаберных лепестков. В частности, у рыб развилось 4-7 жаберных мешков, являющихся щелями между жаберными дужками и содержащими большое количество лепестков, которые пронизаны капиллярами (рис. 190). У рыб в дыхании участвует также воздушный пузырь.
Легкие эволюционировали в направлении увеличения дыхательной поверхности, что привело к образованию бронхов и бронхиол. Впервые легкие появляются у земноводных, представляя собой полые мешки. Однако у них в дыхании еще участвует кожа. У рептилий происходит дальнейшая дифференцировка органов дыхания, В частности, строение легких усложняется, в результате чего в них образуются ячеистые структуры (перекладины). Появляются бронхи. У птиц легкие представляют собой губчатые образования, развиваются разветвления бронхов.
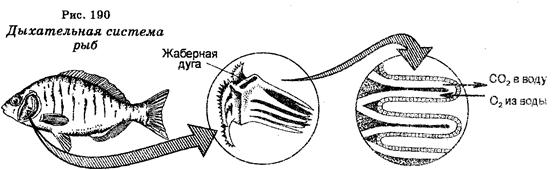
У млекопитающих развитие дыхательной системы достигает вершины (рис. 191). Наряду с дальнейшим совершенствованием легких чрезвычайному развитию подвергаются воздухоносные пути.
 Развиваются бронхи второго, третьего и четвертого порядков, а также бронхиолы и альвеолы. Развивается диафрагма, которая отделяет грудную полость от брюшной. Наличие высокоспециализированных органов дыхания обеспечивает очень эффективный газовый обмен в легких (внешнее дыхание) и в тканях (внутреннее дыхание). Появляются гортанные хрящи. Развиваются бронхи второго, третьего и четвертого порядков, а также бронхиолы и альвеолы. Развивается диафрагма, которая отделяет грудную полость от брюшной. Наличие высокоспециализированных органов дыхания обеспечивает очень эффективный газовый обмен в легких (внешнее дыхание) и в тканях (внутреннее дыхание). Появляются гортанные хрящи.
Эволюция дыхательной системы зависима от колебаний в содержании атмосферного кислорода и углекислоты, поскольку жабры не чувствительны к изменению газового состава воздуха, а кожное дыхание не компенсирует этого недостатка жабер. Начиная с позднего палеозоя, содержание атмосферного кислорода колебалось в разные времена. Хотя фактические величины этих колебаний не определены, тем не менее, предполагают, что максимум содержания кислорода в атмосфере достигал 35%, а затем падал до 15% (при 21% в настоящее время).
Повышенное содержание кислорода и одновременное уменьшение углекислоты повышало эффективность легочного дыхания, помогая позвоночным в завоевании суши. Более того, есть основания к допущению влияния кислородного ритма даже на эволюцию основных групп животных.
У многих беспозвоночных животных сосудистой системы как таковой нет. В частности, у губок, кишечнополостных и плоских червей транспорт питательных веществ и кислорода в разные части тела осуществляется путем диффузии их тканевых жидкостей. Однако у организмов других групп появляются сосуды, обеспечивающие циркуляцию тканевой жидкости в определенных направлениях. В начале развиваются примитивные сосуды, но в результате развития в сосудах мышечной ткани они в дальнейшем оказываются способными к сокращению. Параллельно развивается в кровь жидкость, заполняющая сосуды.
Кровеносная система впервые встречается (развивается) у кольчатых червей, у которых она является замкнутой, представляя собой систему, состоящую из верхнего спинного и нижнего брюшного сосудов, объединенных кольцевыми сосудами, опоясывающими кишечник. Кроме того, от спинного и брюшного сосудов отходят более мелкие сосуды к стенкам тела. К головному отделу кровь идет по спинной стороне, а к заднему — по брюшной.
У членистоногих, кровеносная система не замкнута, т. к. сосуды открываются в полость тела. Спинной сосуд разделяется перегородками (клапанами) на отдельные камеры — сердца, сокращения которых заставляют проходить кровь в артерии, а из последних — в пространства между органами. Из этих пространств кровь затем поступает в околосердечную полость.
У моллюсков кровеносная система тоже не замкнута и представлена сердцем, состоящим из нескольких предсердий и желудочка, а также артериальными и венозными сосудами. Вены впадают в предсердие, тогда как артерии отходят от желудочка.
Кровь у беспозвоночных несет кислород к тканям, удаляет двуокись углерода и транспортирует питательные вещества, гормоны, а также шлаковые продукты азотистого обмена. Кровь беспозвоночных по сравнению с кровью позвоночных содержит меньшее количество белков и форменных элементов. Единственным форменным элементом крови большинства беспозвоночных являются лейкоциты. Поэтому очень простой состав крови у беспозвоночных компенсируется разными способами транспорта кислорода, при которых используются гемоглобин, гемоцианин или другие дыхательные пигменты.
Предполагают, что развитие гемоглобина беспозвоночных шло независимо от развития гемоглобина позвоночных, поскольку этот пигмент у них обладает несколько отличной функцией. В частности, у беспозвоночных он выполняет запасательную функцию, обеспечивая защиту против недостатка кислорода в неблагоприятной среде.
По мере усложнения организации живых существ отмечается повышение количества кислорода в крови, т. е. повышается кислородная способность дыхательных пигментов (табл. 43).
Считают, что гемоглобин имеет древний возраст. Поскольку некоторые бактерии синтезируют гемопротеин, содержащий участки с аминокислотным составом, близким к последовательностям глобинов высших эукариотов, то предполагают, что гемоглобин впервые возник еще у прокариот. Однако не исключено, что у эукариот и прокариот имела место конвергентная эволюция. Гемоглобин встречается у плоских, круглых и кольчатых червей, членистоногих, моллюсков, иглокожих, рыб, земноводных, рептилий, птиц, млекопитающих. Молекулярная масса гемоглобина беспозвоночных колеблется в пределах 17 000-2 750 000. У многих членистоногих и моллюсков встречается гемопианин, представляющий собой медьсодержащий белок молекулярной массой 400 000—5 000 000. Дыхательный пигмент обеспечивает у беспозвоночных буферную способность крови. Кроме того он ответственен и за осмотический баланс кровяной жидкости. Большинство насекомых не содержит совсем дыхательного пигмента, имея в крови растворенный кислород.
Таблица 43
Кислородная способность различных дыхательных пигментов
| Животные
|
Пигмент
|
Цвет пигмента
|
Метал-
лоэле-мент
|
Локализация пигмента
|
Количество кислорода (в мл на 100 мл крови)
|
| Млекопитающие
|
Гемоглобин
|
Красный
|
Железо
|
Эритроциты
|
25
|
| Птицы
|
18,5
|
| Рептилии
|
9
|
| Земноводные
|
12
|
| Рыбы
|
9
|
| Аннелиды
|
Плазма
|
6,5
|
| Моллюски
|
1,5
|
| Моллюски
|
Гемоцианин
|
Голубой
|
Медь
|
Плазма
|
2,8
|
| Аннелиды
|
Хлоро-круорин
|
Зеленый
|
Железо
|
Плазма
|
9
|
| Аннелиды
|
Гемоэрит-рин
|
Красный
|
Железо
|
Эритроциты
|
2
|
У полухордовых кровеносные сосуды еще не выстланы эндотелием. Сердце урохордовых представляет собой мускульное выпячивание около желудка. У хордовых кровеносная система характеризуется дальнейшим совершенствованием. У них развиваются сердце, сосуды, кровь.
У ланцетника кровеносная система является замкнутой, но сердца еще нет; его функцию выполняет передний отдел крупного сосуда в виде брюшной аорты. Впервые сердце появляется у водных позвоночных. В частности, у рыб сердце двухкамерное с предсердием и желудочком.
Круг кровообращения один, в котором артериальная и венозная кровь не смешиваются. Из сердца венозная кровь идет к жабрам, где, окисляясь, она становится артериальной, после чего расходится по артериям ко всем частям тела. К сердцу кровь вновь доставляется венами. У пресмыкающихся желудочек разделен неполной перегородкой на правую (венозную) и левую (артериальную) половины. У крокодилов желудочек разделен полностью. Однако
у пресмыкающихся разделения артериального и венозного тока крови еще не происходит.
 У наземных позвоночных развивается трех-, а затем четырехкамерное сердце, что явилось результатом очень крупного ароморфоза. У птиц и млекопитающих сердце построено из мышц и разделено на четыре камеры в виде двух предсердий и двух желудочков (рис. 192). Имеются два круга кровообращения, один из которых большой, второй малый. Благодаря этим кругам артериальная и венозная кровь не смешиваются. Артериальная кровь выходит из сердца, венозная вступает в сердце. У наземных позвоночных развивается трех-, а затем четырехкамерное сердце, что явилось результатом очень крупного ароморфоза. У птиц и млекопитающих сердце построено из мышц и разделено на четыре камеры в виде двух предсердий и двух желудочков (рис. 192). Имеются два круга кровообращения, один из которых большой, второй малый. Благодаря этим кругам артериальная и венозная кровь не смешиваются. Артериальная кровь выходит из сердца, венозная вступает в сердце.
Сердце позвоночных относят к миогенному типу, поскольку его сокращения начинаются внутри за счет сократительных элементов, называемых миофибриллами. У рыб, земноводных и пресмыкающихся сокращения сердца начинаются в тонкой мышечной стенке венозного синуса и продолжаются посредством внутренней проводящей системы через предсердие, потом к желудочкам. У птиц и млекопитающих венозного синуса нет.
У позвоночных усложняются строение и функции крови. Кровь состоит из форменных элементов и плазмы, увеличивается их масса. Например, в крови млекопитающих клеточная масса крови составляет 45% , плазменная — 55% . Гемоглобин содержится в эритроцитах, которые у всех позвоночных, кроме млекопитающих, содержат ядра. У земноводных клетки крови очень крупные по сравнению с пресмыкающимися, птицами и млекопитающими. В 1 мл крови человека содержится 5 млн эритроцитов и 8—10 тыс лейкоцитов. Группы крови современных людей также являются результатом их эволюции (см. гл. XVI). В плазме крови содержатся ферменты, гормоны и другие соединения, необходимые для метаболизма клеток.
В ходе эволюции у высших животных взаимосвязи обмена веществ разных органов, обеспечиваемые кровью, получили дальнейшее усложнение. Одна из важнейших функций крови заключается в транспорте кислорода от легких к тканям и в транспорте СОд, являющимся конечным продуктом дыхания, от тканей к легким, причем эта функция крови развивалась в направлении способности транспортировать большие объемы кислорода. Основная часть кислорода, потребляемого организмом человека, транспортируется гемоглобином эритроцитов, масса которого составляет примерно треть массы эритроцитов. Как неоднократно отмечалось ранее, гемоглобин представляет собой очень сложный белок, состоящий из двух а-полипептидных цепей и двух b-полипептид-ных цепей, каждый из которых присоединяет по одной гемовой группе. Его происхождение является очень древним (см. § 49).
Эритроциты участвуют также в транспорте СО2
из тканей в легкие, откуда он выводится в процессе выдыхания. Эволюционным приобретением млекопитающих, в том числе человека, является то, что их гемоглобин приспособлен как для транспорта кислорода, так и для транспорта СОд, причем оба эти транспорта взаимоусиливаемы.
Кровь ответственна и за перенос питательных веществ из тонкого кишечника в печень и другие органы, а также за выведение шлаков из тканей в почки. Эти функции тоже развивались в направлении увеличения количественной способности в связи с увеличивающейся интенсификацией метаболизма по мере усложнения организации живых существ.
Диффузия кислорода, двуокиси углерода, питательных веществ и метаболитов происходит лишь в капиллярах.
Параллельно с развитием кровеносной системы шло развитие лимфатической системы, которая впервые появилась у хордовых. Эта система в ходе эволюции обособилась из венозной системы.
Лимфатическая система состоит из лимфатических сосудов, лимфатических узлов и лимфы.
Лимфатические сосуды построены из эластичной и мышечной тканей и выстланы внутри эндотелием. Для них характерен такой же внутренний объем, как и у кровеносных капилляров. Лимфатические сосуды очень разветвлены. Они в виде очень тонких капилляров начинаются с межклеточных пространств, формируя затем более крупные сосуды. С помощью лимфатических сосудов лимфа проходит от тканей в венозное русло. Все лимфатические сосуды сливаются в один общий сборный сосуд (млечный путь). Одиночные лимфатические узлы обнаруживаются в толще слизистой оболочки и подслизистой основы кишечника у рыб, земноводных, пресмыкающихся, птиц и млекопитающих, но групповые лимфатические узлы развиваются лишь у млекопитающих.
У позвоночных по мере усложнения их организации наступает приближение лимфатических капилляров к клеткам и лимфоид-ным узелкам. Например, расстояние между лимфатическими капиллярами, кишечными эпителиоцитами и лимфоидными узелками у рыб составляет 150-180 мкм, но затем прогрессивно уменьшается у земноводных, пресмыкающихся и птиц, составляя у млекопитающих всего лишь 50-30 мкм.
У простейших выделение продуктов обмена осуществляется через всю поверхность тела путем диффузии. Этот же механизм выделения действует также у губок и иглокожих, однако у некоторых из них освобождение от продуктов обмена осуществляется с помощью фагоцитоза.
У плоских червей, ротифер и некоторых немертин развивается примитивная выделительная система в виде протонефридий, которые представляют собой слепые канальцы, разветвленные по телу и снабженные на конце пламенной клеткой, проникающей в канадец (рис. 193). Продукты выделения поступают в цитоплазму пламенных клеток, а затем в канальцы, объединяющиеся и открывающиеся наружу в виде пор. Через эти поры продукты выделения и выходят в окружающую среду.
 У круглых червей выделительная система в виде протонефридий представлена внутрицитоплазматическими каналами, проникающими в полость тела. У круглых червей выделительная система в виде протонефридий представлена внутрицитоплазматическими каналами, проникающими в полость тела.
У кольчатых червей, у которых появляется вторичная полость (целом), развивается выделительная система в виде метанефридий, представляющих собой извитые канальцы, располагающиеся по два в каждом сегменте тела (рис. 194). Эти канальцы открываются во вторичную полость тела одним концом (нефро-стомом), снабженным мерцательными ресничками, а наружу (на боковой стороне гельминта) — другим (нефри-диопорой).
У кольчатых червей метанефридий очень многочисленны. У насекомых и других членистоногих выделительная система представлена слепыми каналь-цами (мальпигиевыми сосудами), включенными в целом и открывающимися в анальное отверстие. Эти канальцы выстланы полусферическим эпителием.
У моллюсков также имеются метанефридий, но они представлены всего лишь одной парой, каждая из которых открывается нефростомом в околосердечную полость, а другим концом в бронхиальную полость. Та ким образом, выделительную функцию осуществляет структура, формируемая канальцами каждого метанефридия. ким образом, выделительную функцию осуществляет структура, формируемая канальцами каждого метанефридия.
У хордовых выделительная система характеризуется дальнейшим усложнением и разнообразием, определяемым переходом от нефридий у низших хордовых к почкам трех «поколений» у позвоночных. У бесчерепных, в частности у ланцетника, выделительная система состоит из располагающихся в районе жаберной щели сегментированных канальцев, в которые жидкость поступает из це-лома через нефростомы и выводится в околожаберную полость через не-фридиоспоры. У ланцетника количество таких нефридий достигает до 100 пар.
У позвоночных в период эмбриогенеза из мезодермы развивается три «поколения» почек — предпочка, или головная почка (пронефрос) у круглоротых (миксин), первичная, или туловищная почка (мезонефрос) у круглоротых (миног), рыб и земноводных и вторичная, или тазовая почка (метанефрос) у пресмыкающихся, птиц и млекопитающих.
Усложнение выделительной системы отмечается в первую очередь в строении почечных канальцев. В предпочке почечные канальцы еще похожи на метанефридий, т. е. они являются канальцами, открывающимися мерцательным нефростомом в целом, а количество канальцев составляет 4-10. У миксин пара предпочечных протоков тянется от шейной части до клоаки.
Дальнейшее усложнение связано с развитием большей эффективности фильтрующей системы внутри канальцев. Для первичной почки характерно увеличение извилистости капилляр внутри канальцев, которых может быть около 100, концентрация их в клубочки и локализация в особых камерах (капсулах), образуемых стенкой канальцев, что приводит к образованию так называемых мальпигиевых телец. Первичные почки являются овальными телами.
Вторичная почка представляет собой уже дольчатый орган, канальцы которого (их количество доходит до 1 млн) открываются в мочеточник, переходящий в мочевой пузырь (у птиц последнего нет). У земноводных, пресмыкающихся и птиц протоки 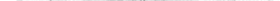 выделительной системы открываются непосредственно в клоаку, тогда как у млекопитающих мочевой пузырь переходит в мочеиспускательный канал, открывающийся наружу. выделительной системы открываются непосредственно в клоаку, тогда как у млекопитающих мочевой пузырь переходит в мочеиспускательный канал, открывающийся наружу.
Почки разных поколений развиваются из мезодермы. Пронефрос закладывается из парных зачатков. Разрастаясь, они сливаются в так называемый предпочечный проток. Закладка мезонефроса происходит сзади пронефроса. В процессе развития проток мезонефроса расщепляется на так называемые вольфов и мюллеров каналы.
Метанефрос развивается позади мезонефроса. Канальцы этой почки открываются в мочеточник, который отделяется от вольфова канала, развивающегося в семяпровод у самцов, тогда как мюллеров канал сохраняется, выполняя роль яйцевода у самок.
У взрослых форм круглоротых, рыб и земноводных функционируют первичные почки, но у миксин еще сохраняются некоторые особенности структуры предпочки, у личиночных форм рыб и земноводных предпочки сохраняются полностью. У взрослых форм пресмыкающихся функционируют только вторичные почки. Они представляют собой парные сплошные или дольчатые тела, располагающиеся под позвоночником.
У млекопитающих выделительная система помимо почек (рис. 195) представлена мочеточниками, мочевым пузырем и мочеиспускательным каналом. Резко возрастает «производительность» выделительной системы. Почки здорового человека образуют ежедневно около 2 л мочи. Половина твердых веществ суточного количества мочи составляет мочевина (около 30 г), которая является главным шлаковым продуктом белкового метаболизма.
У простейших нервной системы в той форме, как ее обычно понимают, не существует, однако в их цитоплазме имеется аппарат возбудимости и движения. В частности, у парамеций центральная нейромоторная масса цитоплазмы связывается фибриллами с основанием каждой реснички, благодаря чему образуется примитивный координирующий центр и проводящая система.
Начало нервной системы простейшего типа отмечается у ки-шечнополостных (рис. 196). Причем первым этапом в ее возникновении является дифференциация эпителиальных клеток в нейро-сенсорные элементы. Поверхность этих клеток выполняет роль рецептора, в то время как от их основания разветвляются нервные нити в сторону подлежащих мышечных клеток. Истинные нервные клетки, обладающие функцией проводимости, наблюдаются впервые также у кишечнополостных, у которых впервые появляются нейроны, секретирующие катехоламины.
Дальнейшее развитие нервной системы оказалось связанным с переходом организмов от радиальной симметрии к билатеральной и заключалось в концентрации нервных клеток в разных частях тела.
 У плоских червей концентрация нервных клеток завершилась формированием парных головных нервных узлов с отходящими от них нервными волокнами к органам чувств и парных нервных стволов, проходящих по телу от головной части к каудальной. У круглых червей уже встречается окологлоточное нервное кольцо, образуемое за счет слияния головных нервных узлов, а у кольчатых червей даже развивается нервная цепочка за счет образования парных нервных узлов в сегментах тела. У плоских червей концентрация нервных клеток завершилась формированием парных головных нервных узлов с отходящими от них нервными волокнами к органам чувств и парных нервных стволов, проходящих по телу от головной части к каудальной. У круглых червей уже встречается окологлоточное нервное кольцо, образуемое за счет слияния головных нервных узлов, а у кольчатых червей даже развивается нервная цепочка за счет образования парных нервных узлов в сегментах тела.
У членистоногих отмечается дальнейшая концентрация нервных клеток, в результате чего обособляются нервные центры, развиваются органы чувств.
Эволюция центральной нервной системы у беспозвоночных и позвоночных проходила, главным образом, в направлении топографической и ультраструктурной перестроек базисных нервных и нервно-эндокринных структур. Она генерирована из одного источника — нейрогенного эпителия, который формирует вначале нервную пластинку, а затем и нервную трубку.
У низших хордовых центральная нервная система представлена трубкой, в головной части которой развивается расширение, представляющее собой зачатки мозга, а полость в расширении (невроцель) — желудочек. Периферическая нервная система представлена отходящими нервами. Нейросекреторные клетки осуществляют сенсорную, проводящую и секреторную функции.
По мере усложнения организмов нервная система эволюционировала в направлении цефализации и повышения ассоциации нейронов (рис. 197). У позвоночных нервная система представлена головным и спинным мозгом и периферическими нервами. Головной и спинной мозг составляют центральную нервную систему. Нервы составляют периферическую нервную систему (соматическую и автономную).
У позвоночных центральная нервная система имеет существенные вариации в зависимости от филогенетического положения организма.
Головной мозг состоит из пяти отделов — переднего, промежуточного, среднего, мозжечка и продолговатого, которые у животных, принадлежащих к разным классам, характеризуются различной степенью развития. Для позвоночных характерна повышающаяся дифференциация серого и белого вещества. Основная функция спинного мозга заключается в интеграции различных сенсомоторных механизмов.
Наименее развит головной мозг у круглоротых, у которых все отделы этого мозга располагаются один за другим.
У рыб центральная нервная система имеет вид трубки, расширяясь в головной части. Дифференциация головного мозга более выражена. В частности, увеличен передний мозг, хорошо выражены зрительные доли среднего мозга, развит мозжечок. Имеется 10 пар черепномозговых нервов.
 У земноводных в связи с выходом их на сушу нервная система характеризуется более сложным строением по сравнению с рыбами, в частности, большим развитием и полным разделением мозга на полушария. Передний мозг (полушария) содержит серое вещество. Дно желудочка, бока и крыша также содержат серое вещество, что свидетельствует о появлении у земноводных первичного мозгового свода (архепаллиума). У земноводных в связи с выходом их на сушу нервная система характеризуется более сложным строением по сравнению с рыбами, в частности, большим развитием и полным разделением мозга на полушария. Передний мозг (полушария) содержит серое вещество. Дно желудочка, бока и крыша также содержат серое вещество, что свидетельствует о появлении у земноводных первичного мозгового свода (архепаллиума).
Из головного мозга как и у рыб выходят 10 пар черепно-мозговых нервов. Симпатическая нервная система хоро-
шо развита. Более совершенное зрение. Наряду с внутренним ухом, развитым у рыб, у них появляется среднее ухо. Большего развития достигает орган обоняния.
У рептилий особенностью нервной системы является прогрессивное развитие всех отделов головного мозга, характерное для наземных животных. В частности, значительно увеличены полушария мозга и мозговой свод. Увеличивается архепаллиум, дифференцируются закладки вторичного мозгового свода (неопаллиума). На поверхности полушарий впервые появляется кора, увеличивается мозжечок, становясь более выпуклым, продолговатый мозг формирует изгиб. Из головного мозга  выходят 12 пар черепно-мозговых нервов. Усложняются нейронные сети. Еще в большей мере развиваются органы чувств. выходят 12 пар черепно-мозговых нервов. Усложняются нейронные сети. Еще в большей мере развиваются органы чувств.
У птиц прогрессивное развитие головного мозга заключалось в увеличении полушарий и зрительной доли, дальнейшем развитии мозжечка.
У млекопитающих увеличение полушарий головного мозга сопровождалось развитием коры, образованием в ней извилин и борозд, завершением развития вторичного мозгового свода (неопаллиума), прогрессивным развитием мозжечка (рис. 198). Основной структурой и функциональной единицей нервной системы является нейрон, развитие которого у человека достигает наивысшего уровня. Происходит дифференциация нейронов на сенсорные, моторные и соединительные. Нейроны синтезируют большое количество нейропептидов, ответственных за коммуникацию клеток и другие важные функции.
Высокого уровня достигает развитие сенсорной системы, в которой наиболее сложными являются органы зрения и слуха. В ходе эволюции зрение впервые появляется у членистоногих. У них оно представлено парой сложных фасеточных глаз, разделенных на омматидии, каждая из которых может различать лишь часть объекта. Насекомые обладают цветовым и объемным зрением. Дальнейшее совершенствование органа зрения характерно для рыб и земноводных.
У рептилий уже отмечается возможность изменения кривизны хрусталика, что ведет к улучшению зрения. В ресничном теле уже развита поперечно-полосатая мускулатура. У земноводных в среднем ухе содержится слуховая косточка, а у рептилий уже увеличилась улитка. Органы зрения и слуха достигают совершенства у млекопитающих, особенно у человека.
Предполагают, что высокая скорость эволюции млекопитающих по сравнению с другими организмами объясняется наличием у них мозга и крупных полушарий. Особенно быстро шло развитие головного мозга у гоминид в плейстоцене, в течение которого объем мозга возрос у них в три раза. Это является примером наиболее быстрых макроэволюционных явлений. Появление нервных клеток означало качественно новый этап эволюции, позволивший высшим животным и человеку регулировать условия среды обитания, а с этим и выживание.
Эндокринной системы в том виде, как ее понимают в применении к высшим организмам, у беспозвоночных нет. Однако у членистоногих в особых железах синтезируются гормоны, которые влияют на линьку (у ракообразных), на созревание яиц и протекание диапаузы, линьку и развитие взрослых форм (у насекомых) и другие свойства.
Таблица 44
Эндокриннная система человека
| Железы
|
Гормоны
|
Гормональные эффекты
|
| Гипофиз
|
Соматотропин, адренокорти-костероидный гормон
|
Рост
|
| Гонадотропный гормон
|
Половые железы
|
| Тиротропный гормон
|
Щитовидная железа
|
| Вазопрессин
|
Почки
|
| Щитовидная железа
|
Тироксин
|
Метаболизм
|
| Паращитовидная железа
|
Паращитовидный гормон
|
Кальций крови
|
| Надпочечники
|
Разные гормоны (более 30)
|
Метаболизм
|
| Островки Лалгерганса
|
Инсулин
|
Метаболизм углеводов
|
| Яичники
|
Эстрогены, прогестероны
|
Гениталии и половые
признаки
|
| Тестисы
|
Тестостерон
|
 Относительно совершенная система появляется у позвоночных. Уже у земноводных имеется гипофиз, надпочечники, щитовидная, поджелудочная и половые железы. Относительно совершенная система появляется у позвоночных. Уже у земноводных имеется гипофиз, надпочечники, щитовидная, поджелудочная и половые железы.
Чрезвычайного развития эндокринная система достигает у млекопитающих, у которых она представлена совокупностью желез внутренней секреции, выделяющих в кровь гормоны — химические регуляторы (табл. 44, рис. 201).
Гипофиз вырабатывает гормоны, которые не только влияют на свойства организма, но и на синтез гормонов Другими железами внутренней секреции. В частности, гипофизарный гормон со-матотропин повышает частоту роста клеток, стимулирует рост и развитие тканей. Вазонрессин оказывает влияние на реабсорбцию воды почечными канальцами. В то же время гипофиз стимулирует секрецию тироксина, который повышает частоту метаболизма в клетках и тканях, а гипофизарный гонадотропин влияет на синтез половых гормонов.
Надпочечники секретируют ряд гормонов, среди которых наиболее важными являются кортизол, альдостерон и эпинефрин (адреналин). Кортизол регулирует метаболизм белков, жиров и углеводов, влияет на беременность, альдостерон — метаболизм натрия и калия, а эпинефрин стимулирует распад гликогена печени и повышает кровяное давление. Гормоны, получившие название эстро-генов, синтезируются в яичниках и тестисах из холестерола в ответ на сигналы, поступающие из мозга и других органов. Попадая в кровь, они разносятся затем к молочным железам и репродуктивным органам. Поскольку эстрогены растворимы в липидах, обычно содержащихся в мембранах, то они легко преодолевают мембранные барьеры и входят в ядро, где связываются с присутствующим в ядерном содержимом белком-рецептором. В дальнейшем комплекс эстроген + белковый рецептор связывается с регуляторными районами ряда генов и изменяют их экспрессию путем активирова-ния или ингибирования действия генов. Комплекс эстроген + рецептор регулирует также экспрессию гена, кодирующего синтез белка-рецептора для прогестерона.
Эндокринная система у высших организмов ответственна за гуморальную регуляцию роста и развития. Все гормоны представляют собой химические соединения, обладающие функциями сигналов, связывающих внутренние коммуникационные системы организма, чем помогают клеткам «чувствовать» и реагировать на изменяющиеся физиологические условия. Различают пептидные гормоны (все гормоны гипоталамуса, гипофиза, инсулин и глюка-гон), стероидные (гормоны коры надпочечников, андрогены и эстрогены) и гормоны типа аминов (эпинефрин и тиреоидные гормоны).
В ходе эволюции у высших организмов сформировалась сложная система регуляторных взаимосвязей между железами, клетки которых продуцируют гормоны, и клетками (органами)-мише-нями. Главным центром развитой эндокринной системы является гипоталамус. В ответ на сигналы, которые получает гипоталамус из центральной нервной системы, его клетки секретируют ряд гипоталамических регуляторных гормонов, каждый из которых в свою очередь регулирует секрецию одного из гипофизарных гормонов. Последние, оказавшись в крови, подходят к надпочечникам, щитовидной железе и стимулируют выработку разных гормонов, которые, попав в кровь, транспортируются к рецепторам гормонов, расположенных на поверхности клеток или тканей-мишеней этих гормонов. В клетках существуют внутриклеточные сигнальные посредники (передатчики), которые передают затем сигнал с клеточного (тканевого) рецептора гормона на какую-либо из специфических внутриклеточных структур или какой-либо фермент.
У простейших специальной репродуктивной системы нет. У губок и кишечнополостных такой системы тоже нет, их половые клетки выходят наружу через стенки тела. Репродуктивная система в виде половых желез и протоков для выведения половых клеток впервые развивается у плоских червей. Вопреки гермафродитизму, у плоских червей формируются семяпроводы и семяизвергатель-ные каналы для мужских половых клеток, а также яйцеводы, матка и влагалище для женских половых клеток. Половые клетки развиваются в семенниках и яичниках.
У круглых червей уже возникает раздельнополость. Отмечается развитие копулятивного аппарата, необходимого для внутреннего оплодотворения.
У хордовых прогрессивное усложнение репродуктивной системы шло в направлении специализации женских и мужских половых желез и формирований приспособлений для внутреннего осеменения. Для ланцетника характерно наличие большого количества половых желез (26 пар), располагающихся в стенках околожаберной щели. Эти железы еще не имеют протоков для выведения половых клеток. Поэтому последние благодаря разрывам стенки желез вначале попадают в околожаберную щель, а затем наружу через атрио-пор. У круглоротых уже имеется непарная половая железа, но протоков еще нет. После созревания половые клетки оказываются в полости тела, откуда через половые отверстия выводятся наружу.
У рыб характерно увеличение половых желез. Яичник обычно непарный, а семенники часто парные. Выводные протоки половых желез у рыб уже связаны с выделительными канальцами почек. Сложность оплодотворения яйцеклеток в водной среде компенсируется выработкой огромного количества половых клеток (икры и молоки).
У земноводных половые железы являются парными как у самцов (семенники), так и у самок (яичники). У части амфибий (бесхвостых) еще сохраняется наружное оплодотворение, тогда как у хвостатых оплодотворение является внутренним. Их развитие происходит с метаморфозами.
У пресмыкающихся женские и мужские половые железы сходны по строению с такими железами земноводных, но у них сильнее развит правый яичник. Крокодилы продуцируют большое количество яиц, но мелких.
У птиц репродуктивная система тоже асимметрична, но больше развит левый яичник. Яйцевод является очень большим, причем делится на передний и задний отделы. Яичник очень увеличен в период кладки яиц.
После завершения откладывания яиц он уменьшается. Яйца относительно крупные.
Наибольшего усложнения репродуктивная система достигает у млекопитающих, для которых характерно только внутреннее оплодотворение. Происходит развитие совокупительных органов, приспособлений для развития плода в организме матери. Развиваются придаточные железы. Характерным является также сложное половое поведение организмов этого класса
Исключительное совершенствование отмечается в организации половых клеток, особенно мужских. У большинства беспозвоночных половые клетки обладают структурными компонентами, характерными для одноклеточных простейших и соматических клеток многоклеточных. В частности, акросома является первичной лизосомой. Имеются центриоли, митохондрии и жгутик с аксоне-мой. У организмов, приспособившихся к жизни на суше, после развития внутреннего оплодотворения сперматозоиды развились в вытянутые структуры, а в их хвосте появились дополнительные цитоскелетные элементы. У высших наземных организмов развиваются разные формы подвижности сперматозоидов и разные типы акросом, что зависит от строения яйцевых оболочек.
1. Какое место занимает эволюция систем органов в эволюции организмов и как важно знать об этом?
2. Назовите основные черты эволюции покровов тела.
3. Как эволюционировал осевой скелет, в чем заключались эволюционные изменения этого скелета?
4. В каком направлении эволюционировала пищеварительная система?
5. Как связаны между собой пищеварительная и дыхательная системы эволюционно?
6. Назовите особенности, характеризующие развитие дыхательной системы позвоночных животных, принадлежащих к разным классам.
7. В каком направлении шла эволюция выделительной системы?
8. С чем было связано усложнение кровеносной системы в ходе эволюции у наземных животных?
9. В каком направлении шла эволюция нервной системы?
10. Что вы знаете об эволюции продуктивной системы? Вовлекались ли в эволюцию половые клетки?
11. Можно ли считать, что эволюция систем органов человека продолжается и сейчас? Если да, то какие системы органов продолжают эволюционировать?
Яблоков А. В., Юсуфов А. Г. Эволюционное учение. М.: Высшая школа. 1989. 335 стр.
Long J. The Rise of Fishes: 500 Million Years of Evolution. The Johns Hopkins Uni versity Press. 1995. 233 p.


«Государству
не может быть безразлично,
как ведется хозяйство
на его земле»
Е. Н. ТРУБЕЦКОЙ,
1906 г.
Живые организмы связаны между собой не только происхождением, но и различными отношениями в процессе их жизни. Кроме того, они связаны и с окружающей их средой (неживой природой). Изучение закономерностей взаимоотношений организмов между собой и с окружающей их средой является задачей науки, впервые названной в 1866 г. Э. Гек-келем в его основном труде «Всеобщая морфология» экологией (от греч. oikos — жилище, logos — наука).
Однако в качестве самостоятельной науки экология стала лишь в XX в. В наше время экология стала чрезвычайно дифференцированной наукой. Ее классифицируют в основном на общую экологию, физиологическую экологию, экологию поведения, экологию популяций и сообществ, экологию экологических систем, а также экологию ландшафтов и статистическую экологию. Задачами общей экологии являются изучение закономерностей существования живых существ во времени и пространстве, численности организмов, круговорота веществ и энергии с участием живых организмов, а также изучение форм взаимодействия организмов между собой и с факторами среды. Задачей экологии поведения является изучение поведенческих реакций организмов (выбор брачных партнеров, других отношений к организмам своего вида, выбор пищи и т. д.) в разных экологических условиях. Задачей экологии популяций является изучение частоты рождаемости и смертности в популяциях, динамики и особенностей регуляции их численности, а также процессов конкуренции, хищничества, мутуализма и других форм взаимоотношений между разными организмами.
Задачей экологии сообществ является изучение групп организмов разных видов, обитающих на определенной территории, а также факторов, определяющих видовое разнообразие и взаимодействие между видами в сообществах. Задачей экологии экосистем является изучение организмов в экологических системах с акцентом на абиотические факторы, действующие в этих системах. Кроме того, эта экология изучает закономерности круговорота веществ и энергии в экологических системах. Задачей экологии ландшафтов является изучение экологических закономерностей на различных ланд-шафных территориях (садах, парках). Задачей статистической экологии является изучение закономерностей распределения организмов. Нельзя не отметить, что границы между этими разделами экологии довольно размыты.
Экологию классифицируют также в зависимости от изучаемых объектов. Различают экологию растений, животных, микроорганизмов и человека. Экология является биологической наукой, однако следует обратить внимание на то, что в наше время в связи с научно-техническим прогрессом она стала междисциплинарной наукой. Экология тесно связана с генетикой, поскольку данные генетики позволяют осуществлять генетический мониторинг в популяциях организмов, включая человека. Экология имеет традиционные связи с биохимией. В частности, биохимия помогает выяснению путей метаболизирования растениями различных токсинов. Экология связана также с физиологией, поскольку она учитывает последствия регуляции жизненных функций. Экология также связана с химией, математикой, географией и другими естественными науками. Больше того, экологию можно считать и единой биосоциальной или социально-биологической наукой, ибо в ее предмет изучения вошли взаимодействие и взаимосвязи надорганизменных систем различного иерархического уровня друг с другом и с окружающей средой.
В совокупности интерес всех экологических наук составляет ряд принципиальных вопросов. В частности, ими являются вопросы о причинах распространения видов, о причинах и механизмах варьирования среды во времени и пространстве, о механизмах влияния организмов на среду и об «ответах» организмов на изменения среды. Далее интересом экологии является познание механизмов изменения сообществ организмов во времени и пространстве и связей между разными экологическими системами. Наконец, задачей экологии является изучение круговорота веществ и энергии в разных экологических системах и нахождение путей допустимой интенсификации этих круговоротов.
Экологи и другие специалисты, интересующиеся проблемами экологии, по-разному подходят к изучению экологических проблем. Одни экологи изучают экологию диких зверей или рыб, экологию природных заповедников, парков и садов, решая при этом как общие, так и частные вопросы. Другие опираются на эволюционный подход, связывая экологические процессы с эволюционными и выясняя, каким образом естественный отбор отражается на состоянии экологических систем и каким образом экологические факторы оказывают влияние на эволюционный процесс. Медиков интересуют экологические проблемы, связанные с распространением возбудителей инфекционных и паразитарных болезней, а также проблемы, связанные с молекулярными экологическими факторами (плазмиды). Наконец, в изучении экологических проблем широко используют математическое и компьютерное моделирование, создавая модели, которые очень полезны для решения глобальных проблем, например, причин и последствий потепления.
Научно-технический прогресс выдвинул на первое место экологию человека и по-новому определил ее содержание. Экологию человека сейчас следует рассматривать в качестве науки о реакциях человека на факторы среды обитания, с одной стороны, и о путях и результатах воздействий самого человека на среду обитания, с другой. Далее, в содержание современной экологии входят также вопросы, касающиеся индивидуумов как самостоятельных экологических систем с их микробными и другими биоценозами, внутрипопуля-ционных общественных отношений в связи с окружающей средой и во взаимодействии с окружающей средой. Продолжая эту мысль, можно сказать далее, что экология человека становится, по существу, социальной экологией, предметом изучения которой служат производственные отношения и их взаимодействие с абиотическими и биотическими факторами среды.
Особой задачей всех разделов экологии в связи с научно-техническим прогрессом является необходимость предсказаний последствий деятельности человека в биосфере.
Несмотря на то, что экология стала междисциплинарной наукой, ее важнейшей задачей остается выяснение общих закономерностей, присущих организму и среде, в которой он обитает.
Пространство, в котором живут организмы, и все то, что их окружает в этом пространстве, есть их окружающая среда. В этой среде присутствуют факторы, которые определяют ее содержание, делают ее специфичной для организмов каждого вида, т. е. каждый вид имеет свою среду. Для человека пространство, в котором он живет, является средой обитания. Территория, на которой располагается город, деревня или другой какой-либо населенный пункт, в котором живет человек, есть его среда обитания.
Различают абиотические, биотические и антропогенные (в составе биотических) факторы среды. Под абиотическими (греч. а — не, bios — жизнь) факторами понимают факторы неживой природы. Характер этих факторов определяется их физической и химической природой. Биотическими (греч. bios — жизнь) факторами являются живые организмы растительной и животной природы, обитающие в среде. Эти организмы составляют совокупность биотических факторов. Для абиотических и биотических факторов (растения, животные) характерны многообразие и безграничные связи, а также их взаимодействие в процессе жизни. Антропогенные (греч. anthropos — человек, genos — рождение) факторы — это факторы, возникающие в результате деятельности человека в среде.
Абиотическое содержание среды определяется климатическими, почвенными и водными условиями. Поэтому в соответствии с одной из популярных классификаций абиотические факторы среды классифицируют на физические (температура, свет, влажность,  барометрическое давление), химические (состав атмосферы, органические и минеральные вещества почвы, рН почвы и др.) и механические факторы (рельеф местности, движения почвы и воды, ветер, оползни и др.). Значение этих факторов состоит в том, что они существеннейшим образом определяют распространение видов, т. е. они определяют ареал видов, под которым понимают географическую зону, являющуюся местом обитания (распространения) организмов того или иного вида. барометрическое давление), химические (состав атмосферы, органические и минеральные вещества почвы, рН почвы и др.) и механические факторы (рельеф местности, движения почвы и воды, ветер, оползни и др.). Значение этих факторов состоит в том, что они существеннейшим образом определяют распространение видов, т. е. они определяют ареал видов, под которым понимают географическую зону, являющуюся местом обитания (распространения) организмов того или иного вида.
Для живых организмов характерен диапазон переносимости действия абиотических факторов, причем это определяется их нормой реакции. Одни организмы способны переносить колебания факторов среды в очень широких пределах. Они получили название эв-рибиотных организмов (от греч. eurys — широкий). Другие выдерживают влияние абиотических факторов в очень узких пределах. Их называют стенобионтными организмами (от греч. stenos _ узкий). Эврибионтные и стенобионтные организмы встречаются как среди растений, так, и среди животных.
Физические факторы
составляют значительную часть абиотических факторов. Особое значение принадлежит температуре, поскольку она является важнейшим фактором, ограничивающим жизнь. Различают термические пояса — тропический, субтропический, умеренный и холодный, к которым приурочена жизнь организмов в тех или иных температурных условиях. Верхний и нижний уровни температурного диапазона легальны для организмов. Температуру, которая благоприятна для жизни организмов, называют оптимальной. Большинство организмов способно к жизни в диапазоне от 0° до 50°С.
На основе способности организмов существовать в условиях разных. температур их классифицируют на эвритермные организмы, которые способны существовать в условиях значительных колебаний температур, и сте-нотермные организмы, которые могут существовать лишь в узком диапазоне температур (рис. 202). Эвритермны-ми являются организмы, обитающие в основном в условиях континентального климата. Примером их являются животные многих видов, обитающие в пресных водоемах и способные выдерживать как промерзание воды, так и ее нагревание до 40-45 С. Эвритермные организмы выдерживают самые жесткие температурные условия. Например, личинки многих двукрылых могут жить в воде при температуре 50°С. В горячих источниках (гейзерах) при 85 °С и более обитают многие виды бактерий, водорослей, гельминтов. С другой стороны, арктические виды бактерий и водорослей обитают в очень холодной морской воде. Для многих эвритермных организмов характерна способность впадать в состояние оцепенения, если действие температурного фактора «ужесточается». В этом состоянии резко снижается уровень обмена веществ. Примерами оцепенения являются оцепенение насекомых или рыб при значительном падении температуры. У млекопитающих (медведи, барсуки и др.) оцепенение проявляется в виде зимней спячки, когда резко снижается обмен веществ, но температура тела при этом падает незначительно.
От оцепенения следует отличать анабиоз (от греч. ana — вновь, bios — жизнь, anabiosis — оживление), который представляет собой явление, заключающееся в том, что у организмов под влиянием разных причин может резко снижаться уровень обмена веществ вплоть до отсутствия видимых признаков жизни. Например, у растений высушенные семена сохраняют всхожесть в течение многих лет. Инцистирование инфузорий позволяет сохраняться им живыми до 6 лет, а яйца Diaptomus sanguires сохраняются свыше 300 лет.
Стенотермные организмы встречаются как среди животных, так и растений. Например, многие морские животные способны выдерживать повышение температуры лишь до 30°С. Некоторые кораллы выживают при температуре воды не более 21°С.
Многие виды животных способны или неспособны к собственной терморегуляции, т. е. поддерживать постоянную температуру. По этому признаку их делят на пойкилотермных (от греч. poikiloi —-различный, переменный и therme — жар) и гомойотермных (от греч. homoios — равный и therme — жар). Первым присуща непостоянная температура, тогда как вторым — постоянная. Гомойотермны-ми являются млекопитающие и некоторые виды птиц. Они способны к терморегуляции, которая обеспечивается физическими и химическими путями. Физическая терморегуляция осуществляется за счет накапливания подкожного жирового слоя, ведущего к сохранению тепла, или за счет учащенного дыхания. Химический путь терморегуляции заключается в потоотделении. Пойкилотермными являются все организмы, кроме млекопитающих и нескольких видов птиц. Температура их тела приближается к температуре среды. Лишь некоторые виды этих животных способны к изменению температуры своего тела, притом в определенных условиях. Например, этой способностью обладают тунцы. Важным для пойкилотермных организмов является то, что повышение температуры их тела происходит, когда увеличивается их активность, их обмен веществ.
В ходе эволюция гомойотермные животные развили способность защищаться от холода (миграции, спячка, мех и т. д.).
Свет является важнейшим абиотическим фактором, особенно для фотосинтез ирующих растений (фототрофов). Уровень фотосинтеза зависит от интенсивности солнечной радиации, качественного состава света, распределения света во времени. Однако для других организмов его значение по сравнению с температурой является меньшим, поскольку известны многие виды бактерий и грибов, которые могут длительно размножаться в условиях полной темноты. Различают светолюбивые, теплолюбивые и тепловыносливые растения. Для многих животных зоопланктона свет является сигналом к вертикальной миграции, в результате чего днем они остаются на глубинах, тогда как ночью поднимаются в теплые, богатые кормом верхние слои воды. Для животных, обладающих зрением, наиболее успешно добывание пищи в светлое время.
У животных многих видов длительность светового дня оказывает влияние на их половую функцию, стимулируя ее в период увеличения светового дня (фотопериода) и угнетая ее при уменьшении светового дня (осенью или зимой). У птиц фотопериод влияет на яйцеклетки (рис. 203). Укорочение светового дня служит сигналом к миграции.
Результатом изменения светового режима (длительности светового дня) является фотопериодизм (от греч. photos — свет, periodos — круговращение), под которым понимают годовые циклы развития у многих видов растений и животных. Например пшеница, овес, ячмень и другие культуры зацветают при длинном световом дне на севере, тогда как кукуруза, хлопчатник — при коротком световом дне на юге.
Влажность — это комплексный фактор и представлен количеством водяных паров в атмосфере и воды в почве. Влажность измеряют путем определения относительной влажности воздуха в виде процентного отношения давления водяного пара к давлению насыщенного пара при одинаковой температуре. Важность влажности для жизни организмов определяется тем, что потеря ее клетками ведет к их гибели
 . .
Обычно растения поглощают воду из почвы. Что касается животных, то потребность в воде они реализуют путем ее питья, либо всасыванием через покровы тела, либо с пищей, либо путем окисления жиров.
В зависимости от влажности происходит распределение видов. Например, земноводные, дождевые черви и некоторые моллюски способны жить только в очень влажных местах. Напротив, многие животные предпочитают сухость.
Влажность почвы зависит от количества осадков, глубины залегания почвенных вод и других условий. Она важна для растворения в воде минеральных веществ.
Большое значение в качестве абиотического фактора имеет комбинированное воздействие на организмы температуры и влажности (рис. 204).
Химические факторы
, по своему значению не уступают физическим факторам. Например, большую роль играет газовый состав атмосферы и водной среды. Большинство организмов нуждается в кислороде, а некоторые организмы — в азоте, метане или сероводороде.
Газовый состав чрезвычайно важен для организмов, обитающих в водной среде. Например, в воде Черного моря очень много сероводорода, что делает этот бассейн не очень благоприятным для жизни в нем многих организмов. Что касается наземных организмов, то они малочувствительны к газовому составу атмосферы, поскольку он постоянен.
Соленость очень важна также в водной среде. Например, из числа водных животных наибольшее число видов обитает в соленых водах (морских и океанических), меньшее — в пресной воде и еще меньшее — в солоноватой воде. Способность поддерживать солевой состав внутренней среды влияет на распространение водных животных.
Существенную роль для жизни организмов, особенно растений, играет значение рН. Одни растения способны жить в кислой среде, другие — в щелочной, причем изменения в концентрации водородных ионов очень губительны для них. В среде, рН которой составляет 0, жизни почти нет. При таком рН растут лишь отдельные виды микроскопических грибов и водорослей.
Механические факторы характеризуются тем, что их действие сопровождается образованием свободных от жизни участков, которые затем заселяются, но содержание новых «поселенцев» будет отличаться от исходного вплоть до формирования новых сообществ живых существ.
Образование свободных от жизни участков происходит в результате стихийных бедствий (пожаров, наводнений и др.), различных геологических процессов, действий человека в природе и т. д. Примером механических факторов является обмеление Аральского моря. Вслед за этим на освобожденных от воды территориях появились новые виды животных и растений.
Характерная особенность видов в контексте их отношений к абиотическим факторам заключается в том, что каждый вид обладает определенным диапазоном толерантности (устойчивости) к тому или иному фактору, причем толерантность определяется нормой реакции, т. е. детерминируется генетически. В том случае, если действие абиотического фактора происходит за пределами диапазона толерантности, организм погибает. Оптимальными условиями для жизни вида является средняя часть диапазона его толерантности к тому или иному фактору. В этой части диапазона происходит также размножение организмов вида. Крайние границы диапазона толерантности неблагоприятны для жизни вида.
С другой стороны, характерной особенностью любого абиотического фактора является то, что он может ограничивать не только жизнь, но в той или иной степени и численность вида, действуя при этом в качестве регулирующего фактора. Регулирующее влияние абиотических факторов особенно возрастает на фоне взаимодействия организмов между собой (губительное воздействие на популяции анабиоза, хищничества, паразитизма), которое само по себе является регулирующим.
Следующей характерной особенностью абиотических факторов является их ограничивающая способность, которая заключается в том, что при оптимальном действии всех возможных абиотических факторов на организмы недостаток одного из факторов все же окажет ограничительное воздействие на популяцию. Например, внесение удобрений в почву при отсутствии какого-либо микроэлемента не приводит к ожидаемому улучшению (значительному повышению урожая культуры). Следовательно, данный микроэлемент является ограничивающим (лимитирующим) фактором. Понимание природы ограничивающих факторов имеет важное практическое значение для сельского хозяйства.
Абиотические факторы находятся в постоянном взаимодействии между собой, причем чувствительность организмов к зависимости одного фактора от другого также определяется нормой реакции, т. е. контролируется генетически. Например, губительный результат повышения температуры в наибольшей мере проявляется в случае повышения влажности среды. Другими словами, при «ужесточении» действия на организмы одного абиогенного фактора сужается диапазон устойчивости их к другому абиотическому фактору.
В ходе исторического развития организмы в ответ на смену дня и ночи, на смену времен года, т. е. в ответ на основные ритмы Земли, обусловленные ее вращением вокруг Солнца, выработали в процессе адаптогенеза способность к ритмической жизнедеятельности, что получило название биоритмоа. Характерная особенность биоритмов заключается в том, что они осуществляются синхронно с процессами, протекающими в среде периодически. Различают суточные ритмы (24-часовые) и околосуточные, которые протекают во время от 20 до 28 часов и которые называют циркадными (от лат. cirka — вокруг, около, dies — день).
В случае человека суточные и циркадные ритмы затрагивают многие свойства и физиологические процессы (температура тела, артериальное давление, митотическая активность клеток, биоэлектрическая активность мозга, количество тромбоцитов в периферической крови и т. д.). Предполагают, что эти ритмы генетически контролируются, т. к. у нейроспоры и дрозофилы установлены так называемые «часовые» гены, которые детерминируют циркадные ритмы этих организмов. Установлено также, что циркадный ритм присущ синтезу мРНК на «часовых» генах и что существуют «часовые» белки, способные ингибировать экспрессию «часовых» генов.
Наука, изучающая биоритмы, носит название хронобиология (от греч. chronos — время, logos — наука). Знание закономерностей биоритмов имеет большое значение в сельском хозяйстве и в профилактической медицине.
Все организмы, независимо от их видовой принадлежности, не только испытывают влияние среды, но и сами прямым образом или косвенно влияют на нее. Поэтому среди биотических факторов различают фитогенные, зоогенные и антропогенные факторы.
Фитогенное воздействие на среду чрезвычайно многообразно. Назовем лишь некоторые примеры и, прежде всего, фотосинтез, который определяет газовый состав среды. В результате фотосинтеза зеленые растения выделяют в атмосферу кислород. Они же и поглощают углекислый газ. Наземные растения оказывают влияние на среду также путем изменения структуры и состава почвы. Далее, азотфиксирующие бактерии, извлекая азот атмосферы, также влияют на ее газовый состав.
Зоогенные воздействия на среду также многообразны. Например, потребляя для дыхания кислород, животные также изменяют газовый состав атмосферы. Животные, обитающие в почве, оказывают значительное влияние на ее механический состав.
Антропогенные воздействия мы рассмотрим в гл. XVIII.
Все организмы находятся между собой в самых различных отношениях, которые, в частности, проявляются в форме симбиоза (мутуализма, комменсализма, хищничества, паразитизма), конкуренции и антагонизма.
Симбиоз (от греч. symbiosis — сожительство) — это очень распространенная форма во взаимоотношениях организмов, принадлежащих к разным видам. В рамках симбиоза различают мутуализм, комменсализм, хищничество и паразитизм.
Мутуализм — это взаимодействие между двумя организмами разных видов, которое выгодно для каждого из них. Например, азотфиксирующие клубеньковые бактерии обитают на корнях бобовых растений, конвертируя атмосферный азот в форму, доступную для усвоения этими растениями. Следовательно, бактерии обеспечивают растения азотом. В свою очередь растения обеспечивают клубеньковые бактерии всеми необходимыми питательными веществами. Мутуализмом можно считать также взаимодействие между микроорганизмами, обитающими в толстом отделе кишечника человека, и самим человеком. Для микроорганизмов выгода определяется тем, что они обеспечивают свои питательные потребности за счет содержимого кишечника, а для человека выгода состоит в том, что микроорганизмы осуществляют дополнительное переваривание пищи и еще синтезируют крайне необходимый для него витамин К. В мире цветковых растений мутуализмом является опыление насекомыми растений и питание насекомых нектаром растений. Мутуализм значим и в «переработке» органических веществ. Например, переваривание целлюлозы в желудке (рубце) крупного рогатого скота обеспечивается содержащимися в нем бактериями.
Комменсализм — это межвидовое взаимодействие между организмами, при котором один организм получает выгоду за счет другого, не повреждая его, тогда как другой организм от этого взаимодействия не имеет ни выгоды, ни ущерба. Например, некоторые виды морских полипов поселяются на поверхности тела крупных рыб, питаясь их выделениями, но для рыб данное сожительство является индифферентным, т. е. не имеет никакого значения.
Хищничество — это особый способ жизни, при котором один организм (хищник) живет за счет другого (жертвы), убивая его. Хищничество наблюдается уже у простейших. Например, известна инфузория дидиний (Didinium nasutum), которая является хищником для других простейших, в частности, парамеций. Плавая в воде, дидиний парализует парамецию, прикрепляется к ней, а затем заглатывает. Процесс переваривания одиночной парамеции составляет 2 часа (рис. 205).
Примеры хищничества высших животных в природе многочисленны. Убивая и поедая жертвы, хищники приносят вред популяциям организмов-жерт.в. Но хищничество иногда оказывается и полезным. Например, волки, поедая ослабленных (больных) лосей, способствуют оздоровлению лосиных популяций.
Уменьшение численности хищников может привести к резкому возрастанию численности жертв. Иногда имеет место синхронный рост численности и хищников и жертв. Например, в северных широтах эта синхронность отмечается каждые несколько лет между численностями леммингов, мышей, с одной стороны, и питающихся ими песцов, лис и полярных сов, с другой.
Паразитизм — это форма взаимоотношений организмов, при • которой один (паразит) живет за счет другого (хозяина). Как правило, хозяину присущи большие размеры тела, чем паразиту. В отличие от хищников паразиты не должны убивать хозяев, иначе тем самым они убьют и себя. Поэтому паразиты вызывают болезнь хозяина, но смерть хозяина от этой болезни бывает лишь в очень редких случаях.
Паразитизм очень широко распространен в природе, но его следует отличать от сапробиоза, при котором организм питается мертвым материалом другого организма, хотя некоторые сапро-бионты иногда могут стать паразитами. С эволюционной точки зрения наиболее результативными являются те паразиты, которые вызывают незначительные повреждения хозяина
В процессе эволюции паразитизма паразиты приобрели исключительную специализацию. Они потеряли, прежде всего, те органы, которые в условиях паразитизма перестали быть ценными для их существования. Напротив, у них развились новые органы, которые обеспечивают их способность существовать за счет хозяев, прикрепляясь к его клеткам, обитая в его полостях и жидкостях.
Среди морфологических и биологически специализированных модификаций паразитов наибольшую важность представляет способность к физиологической адаптации в организме хозяина и наличие сложных жизненных циклов, которые обеспечивают распространение их среди новых хозяев. Физиологическая адаптация к хозяину характеризуется хозяйской специфичностью, т. е. паразит может существовать лишь в хозяевах, принадлежащих к определенным видам. Иногда диапазон хозяйской специфичности является очень широким, в результате чего паразит может паразитировать на организмах многих видов, причем родственные виды хозяев иногда имеют и родственных паразитов. Однако часто хозяйская специфичность настолько узка, что паразит может использовать в качестве хозяина лишь организмы одного единственного вида.
Предполагают, что эволюция видов паразитов и их хозяев шла синхронно (правило Фаренгольца). Хозяева оказывают влияние на эволюции паразитов, но и паразиты влияют на эволюцию хозяев. Например, резистентность к малярии, вызываемой в Западной Африке Р. falciparum, связана с тем, что этот паразит является причиной появления новых аллелей в комплексе гистонесовместимос-ти аборигенов.
Хозяев паразитов классифицируют на дефинитивных и промежуточных.
Организм-хозяин, в котором паразит приобретает половую зрелость, является дефинитивным (постоянным) хозяином, а организм-хозяин, в котором паразит проходит лишь некоторые стадии своего развития, служит промежуточным хозяином. Чтобы облегчить собственный перенос от одного хозяина к другому, паразиты часто используют промежуточных хозяев. Иногда завершение жизненного цикла паразита связано с последовательной сменой нескольких промежуточных хозяев.
Распространение паразитов определяется, прежде всего, географическим распространением их хозяев и переносчиков, а также климатическими условиями, в которых обитают их хозяева. Поскольку многие паразиты могут переходить от хозяина к хозяину, то онтогенез паразитов связан с модификацией их репродуктивной системы в направлении повышения ее мощности. В результате этого у паразитов развилась способность продуцировать потомство в огромных количествах. Повышение количества потомства часто связано с бесполым размножением, гермафродитизмом.
Паразитов классифицируют на временных (комары, мухи-жигалки, блохи и др.) и постоянных (простейшие, гельминты и др.), на эндопаразитов (простейшие, гельминты) и эктопаразитов (вши, блохи и др.). Паразитизм имеет важное значение в сельском хозяйстве и медицине, так как многие паразиты являются возбудителями болезней животных и человека.
Конкуренция — это взаимоотношение между организмами при использовании одинаковых жизненных ресурсов (пищи, света, территории и т. д.), которых, однако, недостаточно в определенных средах при значительном росте популяций. Поэтому она ведет к снижению плодовитости, повышению смертности и к другим неблагоприятным последствиям. Конкуренция бывает как внутривидовой, так и межвидовой и сопровождается она борьбой за существование. Внутривидовая конкуренция является более ожесточенной, поскольку организмы нуждаются в одних и тех же ресурсах. Голод иногда у отдельных видов ведет к каннибализму. Конкуренция и борьба за существование играют большую роль в ограничении численности популяции. Их следствием является естественный отбор (см. гл. XIV).
Антагонизм — это форма взаимоотношений, когда один вид в процессе жизни угнетает другой. Например, известны виды растений, которые выделяют биологически активные вещества (фитон-циды), угнетающие развитие растений других видов. Формой антагонизма является антибиоз, который имеет место в случае бактерий и микроскопических грибов, когда организмы одного вида подавляют рост организмов другого вида или сразу нескольких видов. Эта подавляющая способность зависит от продукции организмами антибиотических видов веществ, получивших название антибиотиков. Наиболее известными антибиотиками являются ампициллин, стрептомицин, тетрациклин, хлорамфеникол и другие.
В ходе эволюции в процессе взаимодействия между собой животные организмы выработали многие факторы собственной защиты от проникновения в них вирусов, бактерий, простейших и других патогенных факторов, которые действуют либо механически, либо вырабатывают токсины, оказывающие неблагоприятное влияние на клетки и ткани хозяина.
Факторами естественной защиты человека являются кожа, поверхностные структуры (слизистые оболочки) дыхательного тракта, соляная кислота в желудке, лизопим, интерферон, белые форменные элементы крови и антитела. Защитная функция кожи, слизистых оболочек дыхательного тракта, соляной кислоты, лизоцима, пропердина и интерферона заключается в том, что они препятствуют проникновению патогенов в организм, действуя бактерицидно. Однако если патогены все же проникают в организм, тогда вступает в действие и иммунная система посредством механизмов иммунитета в виде фагоцитарной и гуморальной защиты.
Иммунитет (от лат. immunitas — освобождение от чего-либо) — это невосприимчивость организма к болезнетворным агентам, продуктам их жизнедеятельности и к генетически чужеродным веществам, обладающим антигенными свойствами. Можно сказать, что иммунитет представляет собой способность организма отличать чужеродный материал от своего, например, чужеродный белок от своего и нейтрализовать этот материал. В отличие от наследственности, которая охраняет организмы от резких изменений на протяжении поколений, иммунитет осуществляет охрану на протяжении индивидуальной жизни организма (онтогенеза).
Охранительная роль иммунитета распространяется не только на вирусы, простейшие, грибы, гельминты, но и на чужеродные трансплантанты тканей и органов. Она распространяется также на аутоиммунные процессы, возникающие в организме. Например, в механизме возникновения сахарного диабета у человека имеют значение аутоиммунные процессы против белков, содержащихся в клетках островков Лангерганса панкреатической железы.
Различают иммунитет инфекционный и неинфекционный.
Инфекционный иммунитет классифицируют на противовирусный, антимикробный (антибактериальный) и антитоксический. В случае противовирусного иммунитета (при гриппе, полиомиелите человека и других вирусных инфекциях) происходит разрушение вирусных частиц, при антимикробном иммунитете (при дизентерии человека, бруцеллезе человека и животных) происходит обезвреживание бактериального возбудителя, тогда как при антитоксическом (при столбняке, ботулизме человека и животных и др.) имеет место разрушение токсина, продуцируемого микробами в организме. В рамках инфекционного иммунитета различают также врожденный и приобретенный иммунитет. Врожденный иммунитет — это естественный иммунитет, возникший в ходе исторического развития организмов и передающийся по наследству. Например, куры не восприимчивы к возбудителю сибирской язвы, причем эта невосприимчивость передается по наследству от одного поколения к другому. Приобретенный иммунитет приобретается организмом в ходе его жизни. Такой иммунитет классифицируют на активный и пассивный иммунитет. Активный иммунитет возникает после перенесения организмами болезни или после их вакцинации, тогда как пассивный — после введения в организм готовых антител (сыворотки, содержащей антитела).
Неинфекционный иммунитет является результатом исторически сложившейся генетической несовместимости. Например, неинфекционным иммунитетом является несовместимость донора и реципиента по группам крови, проявляющаяся в виде тяжелых осложнений при переливании несовместимой крови. Неинфекционным иммунитетом является также трансплантационный иммунитет, развивающийся при пересадке сердца и других органов у человека. Трансплантационный иммунитет возникает, когда ткани донора и реципиента неидентичны. Этот иммунитет проявляется в виде разрушения (отторжения) пересаженной ткани или органа уже через 8-23 дня после пересадки.
Различают клеточный, гуморальный и тканевой иммунитет.
Клеточный иммунитет заключается в фагоцитозе, т. е. в захва-тывании и переваривании фагоцитами (специализированными клетками) бактерий, проникших в организм (рис. 206). Фагоцитами являются клетки белой крови, называемые микрофагами (эозинофилы, нейтрофилы и базофилы) и макрофаги (подвижные клетки крови — моноциты, клетки лимфатических узлов и селезенки, эндотелий кровеносных сосудов).
 Гуморальный иммунитет — это иммунитет, связанный с выработкой антител и обусловленный взаимодействием антигенов и антител. Антигены — это чужеродные для организма вещества. Ими являются белки, липопротеи-ды, белки в соединении с полисаха-ридами, нуклеиновые кислоты. Антигенами являются также ферменты, токсины, яды змей и пауков, вирусы, бактерии, простейшие. Характерными свойствами антигенов являются их чужеродность и специфичность (видовая, групповая, органная и тканевая), а также то, что они вызывают иммунный ответ в виде образования антител. Можно сказать, что антигены выполняют роль генераторов антител. Известны также аутоантигены, образующиеся в самом организме в результате повреждения тканей. Например, аутоантигены, связанные с множественным склерозом человека, присутствуют в миелиновой оболочке, окружающей нервные волокна центральной нервной системы, представляя собой белок р-кристаллин. Гуморальный иммунитет — это иммунитет, связанный с выработкой антител и обусловленный взаимодействием антигенов и антител. Антигены — это чужеродные для организма вещества. Ими являются белки, липопротеи-ды, белки в соединении с полисаха-ридами, нуклеиновые кислоты. Антигенами являются также ферменты, токсины, яды змей и пауков, вирусы, бактерии, простейшие. Характерными свойствами антигенов являются их чужеродность и специфичность (видовая, групповая, органная и тканевая), а также то, что они вызывают иммунный ответ в виде образования антител. Можно сказать, что антигены выполняют роль генераторов антител. Известны также аутоантигены, образующиеся в самом организме в результате повреждения тканей. Например, аутоантигены, связанные с множественным склерозом человека, присутствуют в миелиновой оболочке, окружающей нервные волокна центральной нервной системы, представляя собой белок р-кристаллин.
Антитела — это белки, в частности, иммуноглобулин, содержащийся в сыворотке крови. Известно около десяти групп разных антител, среди которых у человека наиболее часто встречаемыми являются антитела групп IgG, IgM, I^D, IgE и IgA. Иммуноглобулины (антитела) каждой из этих групп состоят из полипептидных цепей разной молекулярной массы. Например, антитела IgG состоят из четырех полипептидных цепей, две из которых имеют молекулярную массу 22 500, а две другие — 50 000. В соответствии со способом действия антител на антигены различают нейтрализующие, лизирующие, коагулирующие и способствующие фагоциту антитела. Помимо антител, циркулирующих в плазме крови, существуют клеточные антитела, связанные с поверхностью клетки.
В ходе эволюции выработался комплекс между способностью животных отвечать немедленно на какой-либо один из миллионов антигенов и «мощностью» иммунитета. Молодые млекопитающие животные способны распознавать и отвечать на внедрение в организм многих антигенов, но эта их способность является небольшой. Способность животных отвечать на антиген быстрее и мощнее возникает лишь со временем. Однако иммунная система может быть мобилизована до того, как чужеродный фактор попал в организм. Эту особенность используют в иммунизации человека и животных вакцинами, содержащими антигены возбудителей тех или иных инфекций с целью профилактики против этих инфекций. Иногда для увеличения продукции специфических антител используют двухтрехкратную вакцинацию в интервалы через несколько недель.
Появление антител отражает одну сторону иммунного ответа, тогда как другая сторона связана с включением в «работу» лимфоцитов, поскольку морфологическим субстратом антител являются лимфоидные клетки лимфоузлов, селезенки, костного мозга, миндалин, тимуса, а за развитие иммунного процесса и за его специфичность ответственны лимфоциты, плазматические клетки и моноциты (макрофаги) и поскольку центральная роль в иммунном ответе принадлежит лимфоцитам. Они развиваются из так называемых плюропотентных стволовых клеток, которые являются родоначальниками всех клеток крови (эритроцитов, лейкоцитов и тромбоцитов).
Различают долгоживущие тимусзависимые Т-лимфоциты и ко-роткоживущие В-лимфоциты (рис. 207). Т-лимфоциты развиваются в тимусе, тогда как В-лимфоциты — в костном мозге взрослых или в печени плодов. В тимусе Т-клетки развиваются из клеток-предшественников, поступающих в тимус из кроветворных органов. В-клетки у млекопитающих возникают непосредственно из стволовых клеток в кроветворных органах, а у птиц — в фабрициевой бурсе (сумке) из клеток-предшественников, поступающих в бурсу с кровью из кроветворных тканей. Кроветворные ткани, тимус и фабрициеву бурсу называют первичными лимфоидными органами.
После дифференцировки в первичном лимфоидном органе часть лимфоцитов с током крови переносится во вторичные лимфоидные органы (лимфатические узлы, селезенка, аппендикс, миндалины, аденоиды и пейеровы бляшки тонкого кишечника). Именно здесь Т-клетки и В-клетки реагируют с антигенами. Т-лимфоциты первоначально распознают чужеродный антиген, а затем становятся хранителями иммунологической памяти и переносчиками этой информации антителообразующими клетками. В-лимфоциты образуются в огромном количестве (ежедневно по нескольку миллионов). Они активируются Т-клетками и дифференцируются или трансформируются в плазматические клетки, непосредственно образующие антитела (растворимые Иммуноглобулины) против распознанных антигенов.
Лимфоциты — это иммунокомпетентные клетки. Они способны переносить в другой организм свойства иммунного организма — противотканевый иммунитет, иммунологическую память, аллергическую активность. Лимфоциты Т и В осуществляют иммунологи-ческий надзор и память в отношении чужеродного антигена. Реализация гуморального иммунного ответа связана с В-лимфоцитами, которые становятся продуцентами антител, а клеточного — с Т-лим-фоцитами, причем в обоих случаях с участием макрофагов.
Почему аутоантитела не распознают собственные макромолекулы? Считают, что распознание аутоантителами собственных макромолекул не происходит по причине гибели лимфох^итов, выполняющих эту функцию. Несомненно, что данное явление имеет место после развития иммунной системы организма, но до встречи организма с чужеродными макромолекулами.
Иммунный ответ организма координирует белки — цитокины, которые являются своеобразными иммунологическими гормонами.
Антитела, возникшие в результате иммунизации, называют иммунными, тогда как в сыворотках крови нормальных людей и животных могут находиться их естественные нормальные антитела. Схема развития лимфоцитов является относительно простой. Предполагают, что в ходе дробления оплодотворенной яйцеклетки возникает одна клетка, которая является предшественником клеток Т и В. Последующие деления этой клетки продуцирует только клетки Т и В, потомство каждой из которых развивает способность распознавать только один антиген (одни макромолекулы). Однако некоторые из этих антигенов являются собственными макромолекулами организмов. Лимфоциты, распознающие собственные антигены, погибают или делаются «нераспознающими», причем время, когда это происходит, зависит от вида организмов. Например, у мышей это происходит в период рождения, а у человека — еще в период эмбрионального развития.
Известно также, что при размножении в клетках нервной системы вирусов полиомиелита, клещевого энцефалита и др. образуются антигены, вызывающие синтез аутоантител. Эти реакции аутоантителообразования ведут к аутоиммунным болезням (гломерулонефриту, некоторым гемолитическим анемиям, множественному склерозу человека).
Известно, что организм животных обладает способностью синтезировать гигантское разнообразие антител. Например, мыши могут синтезировать антитела около 2´106
специфических типов. Исходя из того, что каждый антиген кодируется только одним геном, а размер одного гена составляет примерно 1000 нуклеотидов, часть мышиного генома, ответственного за синтез антигенов, равна 2´109
нуклеотидным парам.
Для объяснения механизмов образования антител предложено несколько теорий, из которых наиболее популярной является «кло-нально-селекционная» теория (Ф. Вернет, 1961). В соответствии с этой теорией в организме имеется исключительно многочисленная популяция лимфоидных клеток, порядка 1 х 10". Эта популяция очень гетерогенна генетически, в результате чего каждая группа лимфоцитов обладает разным родством к антигену. Вследствие контакта с антигеном лимфоциты, обладающие на своей поверхности белками-рецепторами антигена, подвергаются пролиферации и трансформации в плазматические клетки, продуцирующие антитела. Таким образом, антиген избирательно стимулирует те клетки, у которых к нему есть рецепторы. Давая затем потомство, они образуют множество клонов, состоящих из Т-клеток или В-клеток и обладающих одинаковой антигенной специфичностью.
Клетки определенных клонов лимфоцитов могут сохранять память об антигенном стимуле в течение нескольких делений. Это носит название иммунологической памяти., и на этом явлении основано длительное поддержание иммунитета.
Поскольку апоптоз клеток сопровождается экскрецией в межклеточные пространства деградированной ДЦК итистонов, то считают, что последние являются аутоантигенами, определяя аутоим-мунные реакции.
Иногда иммунная система не способна реагировать на собственные антигены. Это называют естественной иммунологической толерантностью. Если же толерантность к собственным антигенам нарушится, то результатом будет развитие аутоиммунологических болезней. Избыточная антигенная стимуляция приводит к открытому в 1953 г. австралийским ученым П. Медаваром и чешским ученым М. Гашеком явлению приобретенной иммунологической толерантности (от лат. tolerantia — терпение).
Как уже отмечено, антитела представляют собой белки-имму-ноглобулины. Количество их в плазме крови составляет примерно 20% всех плазменных белков. Для них характерно наличие анти-генсвязывающих участков, причем каждый клон В-клеток продуцирует антитела со специфическими антигенсвязывающими участками. Эти участки принимают участие во взаимодействии антигена с антителом (рис. 208).
Основными реакциями иммунитета являются нейтрализация токсинов (антитоксинами), преципитация бактерий (преципитинами), агглютинация бактерий (агглютининами), лизис бактерий (лизина-ми), связывание комплемента, опсонизация (опсонинами). Эти естественные реакции иммунитета широко используют в лабораториях для диагностики многих болезней человека и домашних животных.
Иммунный ответ клеточного типа детерминируется реакциями Т-клеток, которые у позвоночных чрезвычайно важны в защите от вирусов и грибов. Различают цитотоксические Т-клетки, убивающие соматические клетки, инфицированные вирусом, Т-хелперы, помогающие В-лимфоцитам в образовании антител, и Т-супрессоры, подавляющие иммунные реакции. Считают, что Т-хелперы и Т-супрессоры являются главными регуляторами иммунных ответов организмов.
Тканевой иммунитет у животных и человека обеспечивается кожей, слизистыми оболочками, лимфатическими узлами, тканями мышц, мускулатуры кишечника и матки, сывороткой крови и другими жидкостями. Этот иммунитет является неспецифическим.
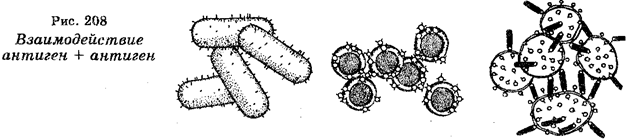
Часто иммунологическая реактивность организмов может изменяться. У человека и животных различают явление, называемое аллергией. Аллергия (от греч. allos — другой, ergon — действие) — это измененная реактивность организма, наступающая под влиянием микробов, токсинов, лечебных препаратов и других веществ, называемых аллергенами. Аллергенами могут служить пыль подушек, перхоть собак, кошек, лошадей, пыль шерсти, хлопка, пыльца растений, яйца, кофе, цитрусовые и др. Как правило, она возникает в результате повторного введения или попадания в организм того или иного аллергена.
Аллергия проявляется либо очень быстро в форме анафилаксии (судороги, выделение мочи и кала, повышение температуры), сывороточной болезни и острого ревматизма либо в замедленной форме (лекарственная болезнь, вызываемая антибиотиками и другими препаратами).
Аллергические, инфекционные и неинфекционные болезни с выраженным проявлением воспалительных реакций, а также иммуно-дефицитные состояния и аутоиммунные заболевания человека в настоящее время довольно широко распространены. Считают, что в основе иммунологических нарушений лежит ряд причин, а именно:
1. Снижение эффективности естественного отбора на устойчивость к инфекциям, т. е. снижение роли отбора в совершенствовании иммунной системы организмов, включая человека.
2. Снижение стимуляции лимфоцитов Т и В, равно как и клеток иммунной системы, антигенами бактерий, обитающих в кишечнике и других полостях тела животных и человека, из-за широкого использования антибиотиков, которые убивают бактерий.
3. Интенсификация воздействия на животных и человека экологических факторов, обладающих иммунологическим эффектом. Иммунитет является недостаточным либо совершенно отсутствует у животных и человека против гельминтов.
Жизнь сосредоточена в океанах (морях) и на материках. Однако пространство океанов и материков измеряется очень большими расстояниями, на которых могут существовать совершенно различные живые формы. Поэтому в целях упорядочивания пространство океанов и материков районируют, выделяя фаунистические и флористические области.
В океане различают 10 фаунистических (зоогеографических) областей, в частности. Арктическую, Атлантическую, Тихоокеаническую, Западноатлантическую, Восточноатлантическую, Магелланову и другие. Что касается материков, то на них различают таких 6-9 областей. По одной из классификаций выделяют Палеаркти-ческую (Европа, Средняя Азия, Северная Азия и Северная Африка), Неоарктическую (Северная Америка), Кругоантарктическую (Антарктика), Неотропическую (южная Америка), Эфиопскую (Африка), Восточную (Юго-Восточная Азия) и Австралийскую области. Для каждой из этих областей характерна. своя фауна. Существуют и другие классификации материковых фаунистических областей, но они не имеют особых отличий от приведенной классификации.
Выделение флористических областей на пространствах морей и материков имеет некоторое отличие, т. к. ботаники предпочитают применять термин «флора» (на практике) лишь к сосудистым растениям. В случае низших растений чаще используют такие словосочетания, как биофлора, альгофлора и т. д. Поэтому флористическое районирование распространяется практически лишь на материки, на территориях которых выделяет ряд областей (фитохорионов) с присущими им определенным видовым составом растений. В выделении областей учитывают эндемизм растений, т. е. приуроченность видов, родов и других систематических единиц растений к определенным территориям, границы которых иногда бывают несколько размытыми. Растения видов, родов, семейств и других систематических единиц, приуроченные в своем распространении к определенным территориям, являются эндемиками.
В районировании различают прежде всего флористические царства, которые подразделяют на флористические области, флористические области — на флористические провинции, а последние — на флористические округа.
В соответствии с наиболее распространенной флористической классификацией различают голарктическое, палеотропическое, неотропическое, капское, австралийское и голантарктическое царства (рис. 209). Наша страна укладывается в границы Голарктического царства, которое охватывает Европу, Северную Африку, Северную Америку и ту часть Азии, которая не является тропической. Голарктическая флора представлена такими семействами растений, как лютиковые, гвоздичные, барбарисовые, березовые, ивовые, магнолиевые, розоцветные, крестоцветные и др.
Видовой состав разных флор очень различен. Наиболее богатыми по видовому составу являются флоры тропических широт. Если в Южной Америке насчитывают около 40 тыс видов растений, то в Гренландии их лишь около 400, а на Шпицбергене около 130. В тропических широтах преобладают орхидные (в основном во влажных тропических лесах), молочайные, мареновые, злаки, бобовые, осоковые. В зонах умеренного климата преобладают сложноцветные, бобовые, розоцветные, злаки, осоковые и крестоцветные. Для засушливых районов характерны обычно мареновые.
Местообитание — это, по существу, место, где живет тот или иной организм. Им может быть участок поверхности земли, водоема или воздушного пространства. Размеры местообитания чрезвычайно варьируют, что зависит от вида организма. Например, ме-стообитанием термитов может служить нижняя поверхность гниющего бревна, тогда как местообитанием лососевых является почти весь Тихий океан.
Различают наземные, воздушные и водные местообитания. Наземные местообитания представляют собой поверхность и поверхностные слои Земли. Наиболее богатой является наземная флора тропических широт. Воздушные местообитания являются временными, т. к. все летающие животные являются все же наземными организмами- Наиболее объемными и населенными являются водные местообитания, которые делят на морские и пресноводные.
Морские местообитания занимают около 78% поверхности земного шара и представляют собой слои воды глубиной до 4000 м, в которых еще активна жизнь. Моря и океаны неоднородны по физическим свойствам, и это ведет к тому, что в разных районах Мирового океана существуют и разные температуры. Температуpa морей зависит от места моря, глубины, течения и времени года. Если в полярных и тропических морях она более или менее постоянна, то в морях континентальной зоны она непостоянна, особенно в разные времена года. На глубинах более 2000 м она всегда и везде составляет около 2°С. Между тем, температура имеет ведущее значение в определении зональной распространенности организмов.
Соленость морской воды также ограничивает распространение живых форм. Например, в Мертвом море из-за высокой концентрации солей вообще нет живых организмов.
Свет чрезвычайно важен для организмов, но на глубинах 2000 м и более устанавливается сплошная темнота. Первичные производители органического вещества в море сосредоточены в прибрежных областях и представлены фитопланктоном, который состоит в основном из диатомовых водорослей и динофлагеллят. Но в открытых тропических морях планктона очень мало. Зоопланктон встречается во всех областях моря и представлен мелкими членистоногими, а также яйцами и личинками разных животных.
Установлено, что основные морские сообщества организмов формируются в зависимости от температуры, глубины и солености того или иного моря. Их классифицируют на тропические и полярные морские сообщества, хотя границы между этими сообществами чрезвычайно размыты.
Пресноводные местообитания составляют 2% от всей площади суши. Они представлены реками, озерами, прудами, водохранилищами. Местообитания организмов здесь ограничиваются температурой, мутностью воды, течениями, кислородом и углекислым ва-зом, солями и осмотическим давлением. Наиболее богатыми по видовому содержанию животных и растений являются прибрежные зоны.
Фитопланктон пресных водоемов представлен зелеными и диа-томовыми водорослями, а также цианобактериями.
Растения, животные, физический субстрат и климат находятся в постоянном взаимодействии между собой. В результате этого в местах обитания формируются крупные комплексы (совокупности) организмов, получившие название биомов.
Различают наземные, морские и пресноводные биомы. Каждый из наземных биомов характеризуется определенным набором растительности (деревья, кустарники, травы и т. д.), с которой связаны живущие в этих местообитаниях животные. Биомами являются тундра, хвойные леса, лиственные леса, влажный тропический лес, степь, чаппараль, пустыня.
В одном биоме может обитать одно или несколько сообществ животных и растений, причем видовое разнообразие сообществ зависит от его возраста, климата, продуктивности и др.
Для биомов, населяющих местообитания, характерна определенная климаксная растительность и определенные виды животных. Между обоими компонентами (группировками) биома существует тесная связь.
Эти группировки растений и животных получили название сообществ. Часто сообщества называют по большинству встречающихся в нем организмов.
Размеры сообществ очень колеблются, тем не менее их структура характеризуется упорядоченностью, которая выражается в определенных отношениях между видами, одни из которых встречаются чаще, другие реже. С другой стороны, для сообществ присуща пространственная структура (размещение в пространстве) и видовая. На состав сообществ влияют пищевые связи. Например, использование степи в качестве пастбища сопровождается ростом той же растительности. Но прекращение выпасов заканчивается ростом только ковыля.
Сообщества состоят из популяций организмов разной видовой принадлежности. Тропические сообщества характеризуются большим количеством видов, но численность видов является не очень большой.
Напротив, сообщества полярных морей характеризуются меньшим количеством видов, но численность их является большей.
Например, в море Лаптевых встречается 400 видов животных, в Карском — 1200 видов, в Баренцевом — 2500 видов, в Северном — 3000-4000 видов, в Средиземном — 6000-7000 видов, в Черном — 1200 видов, в Белом — 1000 видов.
Видовой состав сообществ может быть разным, но некоторые виды в них имеют сходные фенотипы и функционально одинаковы. Эти виды называют экологическими двойниками (экологическим эквивалентами).
Облик современных сообществ является результатом эволюции. В свою очередь сообщества влияют на эволюцию видов.
Сообщества организмов состоят из видов, а виды — из популяций (см. гл. XIV). Вид может состоять из одной или более популяций, причем разной численности. Между популяциями практически не бывает резких разграничении, хотя степень их изоляции зависит от географических условий, от местообитания, от численности соседних популяций. В результате интенсивного размножения границы между популяциями вообще могут размываться. В случае многих млекопитающих популяции имеют стадный характер.
Что же касается структуры самих популяций, то она может быть пространственной, возрастной, половой и генетической.
Пространственная структура определяется размещением особей в пространстве и зависит от биологических свойств вида, от характера местообитания, от времени года. Например, организмы многих видов предпочитают держаться стаями (птицы) либо стадами (млекопитающие).
Возрастная структура популяции определяется количеством в популяции организмов того или иного возраста, а половая — количеством самцов и самок.
Генетическая структура является отражением количественных отношений между разными генотипами в популяции.
Популяции в сообществах взаимодействуют между собой. Одни из них создают среду для других организмов. Например, деревья служат местом для гнездовий птиц или, затеняя почву, мешают развитию травянистой растительности. Это так называемые типические связи. Но часто организмы связаны между собой пищевыми связями, когда один организм служит пищей для других. Например, лисы используют в пищу мышей-полевок, что и определяет их довольно тесную связь.
Для популяций характерна такая величина, как численность или поголовье. Эта величина не является постоянной, ибо зависит от очень многих факторов (темпов размножения, гибели особей в результате старости, болезней, уничтожения хищниками, миграции).
Если по каким-то причинам невозможно определить численность популяции, тогда определяют ее плотность, выражаемую как количество особей на единицу площади (в наземных местообитани-ях) или объема (в водных местообитаниях).
Ареал и численность особей в популяции определяется географическими, физическими и экологическими условиями их обитания. Как и в случае видов, распространение популяций является неравномерным. Поэтому различают «сгущенную» и «островную» формы распределения популяций. В случае «сгущенной» формы распространения популяций их ареал характеризуется наличием особенно плотно заселенных зон. Например, «сгущенное» распределение характерно для популяций берез на русской равнине, где они формируют так называемые высокоплотные на единицу площади чистые березняки наряду с уменьшенным количеством растений в других местах ареала. Напротив, «островное» распределение заключается в очаговом распределении берез в Западной Сибири.
Для популяций характерны различия по возрасту. Например, древесные растения в лесах характеризуются разным возрастом, вследствие чего в перекрестном опылении участвует пыльца разновозрастных растений. Напротив, популяции отдельных видов всегда являются молодыми. Например, популяции дальневосточных лососей в море всегда представлены организмами одного возраста.
Половой состав популяций (соотношение полов) организмов (животных и двудомных растений) разных видов также характеризуется различиями (рис. 210), что отражается на интенсивности их размножения. У тех же организмов, у которых существует партеногенез (ракообразные, насекомые), интенсивность размножения является очень высокой.
Численность популяции представляет собой величину, часто меняющуюся в зависимости от условий обитания, частот рождения, гибели и притока организмов из одной популяции в другую. Иногда разные популяции одного вида объединяются или разделяются на более мелкие.
 И животные и растения способны очень быстро увеличить свою численность в течение определенного промежутка времени. Эту способность, т. е. их плодовитость, называют биотическим потенциалом вида, который особенно велик, например, у бактерий. Несколько меньшим, но все же очень большим он является у насекомых и некоторых ракообразных, численность которых за год может возрасти примерно в 1030
раз. Что касается млекопитающих, то их биотический потенциал является небольшим. Например, одна пара овец за год в среднем может дать лишь одного ягненка. И животные и растения способны очень быстро увеличить свою численность в течение определенного промежутка времени. Эту способность, т. е. их плодовитость, называют биотическим потенциалом вида, который особенно велик, например, у бактерий. Несколько меньшим, но все же очень большим он является у насекомых и некоторых ракообразных, численность которых за год может возрасти примерно в 1030
раз. Что касается млекопитающих, то их биотический потенциал является небольшим. Например, одна пара овец за год в среднем может дать лишь одного ягненка.
Если рост популяции происходит в условиях избытка пищи, достаточного места и других благоприятных факторов, то рост численности происходит в геометрической прогрессии или экспоненциально. Классическим примером экспоненциального роста численности являются вспышки численности саранчи (волн жизни). Однако во времени
экспоненциальный рост численности обычно является коротким, после чего он значительно замедляется.
Установлено, что повышение плотности популяции сопровождается уменьшением условий для ее роста и размножения, в результате чего рост численности замедляется (рис. 211). Очень хорошо это положение иллюстрируют также кривые размножения культур бактерий (рис. 212).
 В мире микроорганизмов прирост численности, т. е. плотности культуры зависит от скорости деления клеток. Что же касается многоклеточных организмов, то рост численности зависит от рождаемости и от смертности. По существу, коэффициент рождаемости отражает степень плодовитости. В мире микроорганизмов прирост численности, т. е. плотности культуры зависит от скорости деления клеток. Что же касается многоклеточных организмов, то рост численности зависит от рождаемости и от смертности. По существу, коэффициент рождаемости отражает степень плодовитости.
Различают абсолютную и удельную рождаемость, причем под первой понимают количество особей, рождающихся в популяции в единицу времени, тогда как под второй понимают количество родившихся особей на определенное число организмов. В случае человека удельную рождаемость выражают коэффициентом рождаемости. Например, если рождается 5,6 детей на 100 жителей, коэффициент рождаемости составит 5,6%.
В противоположность рождаемости смертность определяют как скорость уменьшения численности популяции вследствие гибели отдельных организмов в результате старости, болезней, хищников и т. д.
Изменение численности организмов в замкнутых популяциях зависит от соотношения смертности и рождаемости. При смертности, большей рождаемости, рост численности становится отрицательным. Напротив, при рождаемости, превышающей смертность, рост численности становится положительным, т.е. численность популяции увеличивается.
Численность популяций всегда подвержена колебаниям, частота которых наиболее высокая у насекомых. Например, колебания численности саранчи в годы ее интенсивного размножения (волн жизни) составляют сотни тысяч раз. У млекопитающих колебания численности являются небольшими.
Эффективность действия одних факторов на численность зависит от плотности, действие же других с плотностью не связано. Например, обеспеченность пищей или распространение инфекций зависит от плотности организмов, тогда как губительный характер снежных зим для птиц не зависит от их плотности.
Жизнь организмов зависит от совокупности факторов, действующих в их среде обитания. Однако в ходе исторического развития организмы развили множество приспособительных механизмов, которые придали им чрезвычайную способность к приспособлению в окружающей среде. Именно это обеспечивает способность вида к расширению своего ареала, под которым понимают область земного шара, где встречается данный вид. Ареал может составлять всего лишь несколько квадратных километров либо целый материк и даже больше. Кроме того наличие приспособительных механизмов обеспечивает уход вида от неблагоприятных воздействий среды, чем изменяется его ареал.
Одно из приспособлений, которое позволило животным завоевать сушу, Мировой океан и атмосферу, заключается в том, что им присуща чрезвычайная способность к перемещению, к отысканию областей, в которых имеется больше возможностей для лучшего питания и размножения. Например, многие киты в осенний период года передвигаются в северные широты, где достаточно корма, а с приближением зимы вновь уходят в экваториальные широты. Тунцы и лососевые преодолевают огромные расстояния, чтобы найти места для размножения и откорма. Личинки речных угрей мигрируют из Сар-гассова моря в реки Евразии и Северной Америки, а половозрелые особи (самки) возвращаются на нерест снова в Саргассово море.
Известны также случаи успешного преднамеренного «переселения» животных. Например, из Азовского моря в Каспийское по экономическим причинам были перемещены полихеты и двустворчатые моллюски, а из дальневосточных морей в моря, омывающие Южную Америку и Новую Зеландию, — дальневосточные лососи. Успешным оказалось также «переселение» колорадского жука из Америки в Европу, который стал вредителем.
Растения не обладают самостоятельной способностью к передвижению, хотя ветром их семена могут переноситься на огромные расстояния. Тем не менее для распространения растений характер на зональность, выражающаяся в смене одних сообществ растений на другие по мере продвижения их от экваториальных широт к полюсам. Различают ряд зон, а именно тундры, хвойные леса, лет-незеленые широколиственные леса, степи, пустыни, саванны, вечнозеленые тропические леса и др.
Одно из важнейших приспособлений растений к температуре заключается в форме роста. Например, в холодных районах (Арктика, высокогорье) встречается много стелющихся форм. В то же время южные растения, будучи перенесенными на север, обычно произрастают на прогреваемых склонах. Многочисленные примеры из мира животных и растений свидетельствуют о том, что организмы могут существовать не только в границах природных ареалов, но и далеко за их пределами, давая многочисленное потомство.
В результате приспособления организмов к абиотическим и биотическим факторам создаются условия для существования видов и их связей с другими видами, причем сообщества разных районов населяют эквивалентные формы организмов. Например, кактусы, которые растут в штате Аризона (США), эквивалентны молочаям Европы, внешне не очень сходным и филогенетически очень далеким от них. Но и кактусы и молочаи в своих сообществах выполняют сходную роль, т. е. занимают сходные ниши. Можно сказать, что местообитание организма есть его «адрес», тогда как экологическая ниша, образно говоря, представляет собой «профессию» организма. Организмы двух видов не могут обладать одной и той же нишей (принцип конкурентного взаимоисключения).
Различают фундаментальные и реализованные экологические ниши. Фундаментальная экологическая ниша представляет собой комплекс условий, в которых есть принципиальная возможность для существования того или иного вида. Напротив, реализованная экологическая ниша представляет собой комплекс условий, в которых реально вид обитает. Например, фундаментальной нишей аскариды человеческой является кишечник млекопитающих, тогда как реальной нишей этого организма является кишечник человека.
Систематически подвергаясь воздействиям среды, организмы, в свою очередь, влияют на среду. Примеры этого влияния очень многочисленны. Например, растения изменяют газовый состав атмосферы, т. к. в нее поступает кислород в результате осуществляемого растениями фотосинтеза. Одновременно растения извлекают из атмосферы углекислый газ, а азотфиксирующие бактерии — азот (см. § 95).
Далее, организмы изменяют физические и химические свойства почвы. Например, микроорганизмы участвуют в образовании почвы, тогда как черви и роющие животные изменяют структуру почвы, делают ее более рыхлой. Деятельность растений и животных определяет содержание органических и минеральных веществ в воде (см. § 95).
1. Дайте определение экологии и назовите причины, определившие междисциплинарный характер этой науки.
2. Что вы понимаете под средой организмов? Есть ли разница в понятиях «окружающая среда» и «среда обитания»?
3. Назовите абиотические факторы среды и каково их значение в применении к человеку?
4. Что понимают под циркадными ритмами и какова их роль в жизни растений и животных?
5. Что собой представляет симбиоз, каковы его формы и как часто он распространен в природе?
6. Дайте определение иммунитета. В чем заключается биологическая сущность иммунитета?
7. Перечислите формы иммунитета и охарактеризуйте их.
8. Определите содержание понятий «антиген» и «антитело».
9. Каковы принципы выделения фаунистических и флористических областей?
10. Что представляют собой биомы? Как они формируются?
11. Можно ли изменять ареалы животных и растений искусственно?
Агаджанян Н. А., Торшин В. И. Экология человека. М.: Изд. фирма «Крук». 1994. 256 стр.
Дре Ф. Экология. М.: Атомиздат. 1976. 164 стр.
Пехов А. П. Биология и общая генетика. М.: РУДН. 1993. 439 стр.
Реймерс Н. Ф. Экология. М.: Россия молодая. 1994. 363 стр.
Хаитов Р. М., Пинегин Б. В., Истамов X. И. Экологическая иммунология М.: ВНИРО. 1995. 218 стр.
Brown J. Н. Macroecology. University of Chicago Press. 1995. 265 pp. Elgert K. Immunology. Willey. 1996. 448 pp.
Биосфера (от греч. bios — жизнь, sphaira — пленка) — живая оболочка Земли.
Термин «биосфера» впервые был использован известным австрийским ученым Э. Зюссом (1831-1914) в его книге «Лик Земли» (1875).
Позднее термин «биосфера» использовали и другие ученые, но учение о биосфере в современном понимании было сформулировано В. И. Вернадским (1863-1945) в его знаменитой книге «Биосфера» (1926). По В. И. Вернадскому «Биосфера представляет собой определенную геологическую оболочку, резко отличную от всех других оболочек нашей планеты...» И далее: «Живое вещество проникает всю биосферу и ее в значительной степени создает».
Как отмечал В. И. Вернадский, геохимические процессы на Земле и формирование лика Земли связаны с живыми существами, поэтому биосфера включает в себя собственно живую оболочку Земли (живой материал в виде живых организмов, населяющих Землю в каждый момент) и былые живые оболочки (былой живой материал), границы которых определяются распределением биогенных осадочных пород.
По В. И. Вернадскому «Живое вещество биосферы, единственной области планеты, закономерно связанной с космическим пространством, есть совокупность ее живых организмов, ее живого вещества как планетного явления».
Отмечая, что «Века и тысячелетия прошли, пока человеческая мысль могла отметить черты единого связного механизма в кажущейся хаотической картине природы», В. И. Вернадский сумел подняться на вершины научного творчества. Развитые им положения стали одним из крупнейших естественно-научных обобщений XX в.
В. И. Вернадский подразделил биосферу на тропосферу, литосферу и гидросферу.
Тропосфера — нижняя часть атмосферы, высота которой доходит до 20 км, где жизни уже нет, но происходят миграция и обмен биогенных газов. Объем атмосферы, в которой обнаруживается жизнь, составляет 7 444 600 км8
.
Литосфера — это твердая поверхность Земли, представленная ее верхними водонепроницаемыми слоями глубиной до 2-5 км, ниже которых уже лежат осадочные породы, а еще ниже — переплавленные породы гранитной оболочки. Объем почвенного слоя, в котором обнаруживают жизнь, составляет около 100 000 км3
. Почва заселена такими организмами как бактерии, корненожки, инфузории, черви (нематоды), олигохеты, насекомые, клещи, мелкие млекопитающие, растения.
Гидросфера — это водная часть биосферы, представленная реками, озерами,морями и океанами. Объем морей и океанов составляет около 1 млрд 370 млн км3
, тогда как объем озер, рек, водохранилищ и учтенных подземных вод составляет около 8 млн км3
. Моря и океаны являются одним из основных биотопов, хотя около 90% их объема представлены глубинами, для которых характерен полный мрак. С другой стороны, для глубин более 4000 м характерно также очень высокое давление, составляющее около 400 атмосфер. Можно сказать, что часть Мирового океана является естественной барокамерой, заполненной живыми организмами.
Население гидросферы представлено планктоном, бентосом и нектоном. Планктон представляет собой совокупность мелких организмов животной и растительной природы, которые либо не способны к самостоятельному движению, передвигаясь вместе с водой, либо способны, двигаясь в воде самостоятельно. Различают фитопланктон, который в морях представлен одноклеточными водорослями (диатомовыми), цианобактериями и другими организмами, и зоопланктон, представленный одноклеточными форами-ниферами, радиоляриями и многоклеточными кишечнополостными, а также червями, ракообразными, личинками беспозвоночных животных и т. д. В планктоне пресных вод встречаются в основном низшие ракообразные и коловратки.
Бентос представляет собой совокупность животных (зообентос) и растений (фитобентос), ведущих придонный образ жизни (губки, кишечнополостные, черви, моллюски, ракообразные, иглокожие, асцидии, водоросли и др.). В пресноводном бентосе содержатся личинки некоторых насекомых, брюхоногие моллюски, пиявки, губки (бадяги) и др. Нектон представлен крупными плавающими организмами (морские млекопитающие, рыбы, кальмары и др.). Нектон пресных вод представлен карповыми рыбами.
Зона морских приливов и отливов (осушная зона), которая может составлять всего лишь несколько метров, носит название лито-рали. Она заселена ракообразными, червями, моллюсками.
Глубины водной части биосферы зависят от водоема. В океане они доходят до 10 км и более, причем жизнь встречается на самых разных глубинах.
Считают, что со времени появления жизни на Земле живые существа непрерывно перерабатывали вещество литосферы, тропосферы и гидросферы. Поэтому мощность биосферы определяется биомассой живущих одновременно на Земле организмов. Подсчитано, что биомасса живых существ составляет 2,423´1012
тонн, из которых на долю сухопутных организмов приходится 2,42´1012
тонн, водных — 0,003´1012
. Подсчитано также, что одну треть биомассы Земли составляют одноклеточные организмы, бактерии и простейшие. Кислород в живом веществе составляет 65-70%, водород — 10%, остальные более 60 элементов — 20-25%.
Жизнь и деятельность человека связана с нижними слоями тропосферы (несколько метров), верхним слоем литосферы (биогеоце-нотический покров с почвой и подпочвой, где сосредоточены корневые системы растений) и гидросферой. Теперь мы знаем, что жизнь человека длительное время возможна и в космосе.
Впервые представления об экологических системах сформулированы в 30-е гг. А. Тенсли (1935). В нашей стране близкое понятие о биогеоценозах сформулировал в 1944 г. В. Н. Сукачев (1880—1967). В наше время под экологическими системами понимают совокупность живых и неживых элементов на определенной территории. Экологические системы состоят из живых организмов (биоценозов) и среды обитания — косной (атмосфера) и бикосной (почва, водоем и т. д.). Они иногда отделены одна от другой, но часто между ними имеются переходы. Примерами экологических систем являются озеро, лесной массив и т. д. От экосистем следует отличать биомы, под которыми, как показано выше, понимают значительные сообщества организмов, приуроченные к определенным географическим районам с их климатическими и почвенными зонами.
Экологические системы являются элементарными единицами биосферы. В то же время они являются элементарными единицами биогеохимической активности, протекающей в биосфере. Любая экологическая система имеет энергетический ввод, через который в нее
 поступает энергия солнечного света. Световая энергия, поступающая в экосистему через ввод, поддерживает порядок в этой системе, предупреждая повышение энтропии (рис. 213). поступает энергия солнечного света. Световая энергия, поступающая в экосистему через ввод, поддерживает порядок в этой системе, предупреждая повышение энтропии (рис. 213).
Рассмотрим экологическую систему на примере озера (рис. 214). Как и любая экологическая система, озеро состоит из абиотической и биотической частей.
Абиотическая (неживая) часть экосистемы представлена воздухом, почвой, водой, растворенными в воде кислородом, двуокисью углерода, неорганическими солями (фосфаты и хлориды натрия, калия и кальция) и органическими соединениями. Абиотической частью является также температура, свет, ветер и гравитация, которые оказывают влияние на живую часть.
Биотическая (живая) часть озера представлена организмами-производителями (продуцентами), организмами-потребителями (коксумен-тами) и организмами-разрушителями (редуцентами). Организмами-производителями являются автотрофы — прибрежная растительность, водные многоклеточные и одноклеточные плавучие растения (фитопланктон), живущие до глубин, куда еще проникает свет. За счет энергии, поступающей через ввод, организмы-производители в процессе фотосинтеза синтезируют органическое вещество из воды и углекислого газа. Основным показателем мощности экосистемы является ее продуктивность, под которой понимают массу органического вещества в телах организмов-продуцентов. Продуктивность экосистемы зависит от количества света, воды, богатства почвы или воды органическими и минеральными соединениями.
Организмами-потребителями (коксументами) органического вещества служат гетеротрофы, среди которых различают потребителей первого и второго порядка. Первичными потребителями служат травоядные животные, вторичными — плотоядные, которые питаются первичными потребителями. Организмы-разрушители — это бактерии и грибы, которые разлагают «мертвую» протоплазму (органические соединения) клеток погибших организмов-производителей и организмов-потребителей вплоть до низкомолекулярных органических и неорганических соединений. Органические соединения затем используются самими организмами-разрушителями, тогда как неорганические — зелеными растениями. Итак, в экологической системе в процессе ее функционирования происходит круговорот веществ и энергии.
Мы рассмотрели в качестве экологической системы природную систему (озеро), не уделяя внимания участия в ней человека. Однако большинство экологических систем функционирует с участием человека. В связи с этим различают экологию отдельных индивидуумов и сообществ людей.
Экология отдельных индивидуумов заключается в том, что каждый индивидуум должен «подогнать» свою внутреннюю физиологию к меняющимся условиям среды обитания. Индивидуум получает энергию с пищей и расходует ее для обеспечения своих физических и интеллектуальных усилий, метаболических процессов, протекающих в организме, роста и т. д. Благодаря нейрогуморальной регуляции в организме индивидуума поддерживается постоянная температура тела, оптимальные концентрации воды, кислорода, двуокиси углерода, NaCI, углеводов, белков и других важных соединений. Проникновению в  организм индивидуума патогенных факторов препятствует кожа, антитела, фагоциты и другие факторы защиты. Органы чувств, нервная система и локомоторные органы позволяют индивидууму обезопасить пищу, найти друзей, избегать врагов, создавать ситуацию, наиболее благоприятную для выживания. Каждый индивидуум способен адаптироваться к измененным климатическим условиям. Все это организм индивидуума патогенных факторов препятствует кожа, антитела, фагоциты и другие факторы защиты. Органы чувств, нервная система и локомоторные органы позволяют индивидууму обезопасить пищу, найти друзей, избегать врагов, создавать ситуацию, наиболее благоприятную для выживания. Каждый индивидуум способен адаптироваться к измененным климатическим условиям. Все это  приводит к тому, что между внутренней физиологией индивидуума и условиями окружающей среды устанавливается динамический эквилибриум. приводит к тому, что между внутренней физиологией индивидуума и условиями окружающей среды устанавливается динамический эквилибриум.
Однако люди объединены в сообщества. В состав этих сообществ входят также окружающие их растения и животные, которые являются источником пищи и других необходимых материалов для людей. Следовательно, с учетом абиотических факторов экологическую систему, в которой функционирует ^человек, составляют сообщества людей и среда их обитания. Экологические системы, в которых человек занимает важное место, чрезвычайно разнообразны по размерам, содержанию и организации, что чрезвычайно затрудняет классификацию этих систем. Тем не менее, они являются экологическими системами, в которых центрами являются деревни, города и другие населенные пункты.
Все элементы экологических систем составляют едисоставляют единую совокупность, и это оп ределяется тем, что они объединены между собой так называемыми цепями питания, под которыми понимают передачу от организмовпотребителей заключенной в пище энергии первоначального источника (Солнца) через организмы-потребители (в ряде цепей питания конечным звеном является человек) к организмам-разрушителям.
Важнейшей особенностью цепей питания является то, что их количество в каждой экосистеме ограничено, поскольку в каждом звене каждой цепи питания происходит потеря энергии при ее передаче. В результате этого продукция вещества понижается на каждом звене цепи. Например, 10 000 кг водорослей достаточно для накопления вещества в количестве 1000 кг водных членистоногих, а 10 кг рыбы — для накопления 1 кг вещества человека. Таким образом, пищевая цепь представляется в виде пирамиды, состоящей из нескольких трофических уровней (рис. 215). У основания расположены фотосинтезирующие бактерии, которые являются пищей для следующего уровня, а эти организмы являются пищей для последующего уровня и т. д.
Химические механизмы, лежащие в основе пищевых цепей, представляют собой круговороты (циклы) веществ. С другой стороны, круговороты веществ обеспечивают самоподдержание популяций.
Круговороты веществ не являются замкнутыми. Часть органических и неорганических веществ вытесняется за пределы сообществ, но при этом происходит их пополнение за счет внешних источников (осадки, фиксация азота атмосферы и т. д.). Рассмотрим в качестве примеров круговороты углерода, кислорода и азота.
Круговорот углерода
, входящего в состав всех органических соединений, начинается с конверсии двуокиси углерода (формы углерода в атмосфере) и воды в органическое вещество (пищу). Часть этого вещества используется живыми организмами при дыхании, в результате чего часть двуокиси углерода снова возвращается в атмосферу.
Подсчитано, что весь углерод атмосферы проходит через живые организмы за время, равное 7-8 годам. Другая часть двуокиси углерода запасается в протоплазме клеток. После смерти организмов протоплазма их клеток разлагается, в результате чего двуокись углерода также освобождается и уходит в атмосферу. Основная часть углерода содержится в форме COg в морях и океанах. Его круговорот является таким же, как и в случае атмосферного углерода.
В экологических системах, где принимает участие человек, двуокись углерода поступает в атмосферу также и в результате сжигания растений в качестве топлива.
Круговорот кислорода
заключается в том, что атмосферный кислород используется растениями и животными при дыхании (сжигании пищи), в результате которого освобождается энергия, вода и двуокись углерода в фотосинтезе, при котором освобождается кислород, после чего цикл начинается снова.
Более сложным является круговорот азота, самым большим резервуаром которого служит атмосфера (около 80%). Поскольку большинство растений и животных не может использовать атмосферный азот (N2
), то он конвертируется почвенными азотфиксирущими бактериями, корневой системой бобовых растений и цианобактериями в нитриты (NO-
2
), а затем в нитраты (NО-
3
). Этот процесс получил название нитрификации. Растения восстанавливают нитраты, т. е. уеваивают азот и синтезируют белки. Круговорот азота далее заключается в том, что почвенные микроорганизмы разрушают животные отходы и остатки мертвых организмов, в результате чего освобождается аммоний, который конвертируется нитрифицирующими бактериями в растворимые соли нитратов, используемые в производстве белков в растениях. В результате поедания растений травоядными животными растительные белки в их организме превращаются в животные.
В процессе гниения трупов растений и животных денитрифи-цирующие бактерии превращают нитраты в свободный азот (NO2
® NО-
2
® N2
O ® N2
), который уходит в атмосферу, но азотфиксирую-щие бактерии снова конвертируют атмосферный азот в органические соединения, доступные для усвоения растениями.
Свободный азот конвертируется в нитраты также электрическими зарядами (молнией). Искусственное добавление азотных соединений в почву связано с использованием химических удобрений.
Большое значение в природе имеет круговорот воды. Он осуществляется за счет солнечной энергии, но регулируется со стороны организмов. Под влиянием Солнца моря и океаны подвергаются испарению. Эта вода в форме «испарений» (пара) поступает в атмосферу, а затем выпадает в виде дождей на сушу, после чего через реки и грунтовые воды снова попадает в моря и океаны. Значительная часть воды, имеющейся на суше, поглощается и испаряется растениями. Но некоторая часть воды в клетках растений подвергается фотолизу, в результате чего она разлагается на кислород и водород. Кислород уходит в атмосферу, а водород включается в состав органических соединений клеток.
Важнейшим свойством экологических систем является их устойчивость, т. е. постоянство, которое поддерживается цепями питания. Именно благодаря постоянству цепей питания в природе поддерживается экологический гомеостаз. При этом важно заметить, что устойчивость экологических систем имеет исторический характер, а регуляция тех немногих экологических систем, в которых человек не имеет значения, обеспечивается такими факторами, как конкуренция, миграция, хищничество, недостаток корма или питательных веществ в почве, болезни, температура и другими естественными факторами.
Напротив, в экологических системах, где сообщества людей имеют важное значение, в качестве регулирующих механизмов действуют не только механизмы, названные выше, но и механизмы, действующие непосредственно в человеческом обществе. Ими являются законы, общественное мнение, поощрения и наказания, снабжение и потребление, конкуренция, образование, воспитание навыков экологической культуры, здравоохранение и т. д. Благодаря этим и другим экономическим, социальным и политическим механизмам каждая социальная группа сохраняет свой рабочий баланс в изменяющихся условиях окружающей среды, что оказывает существеннейшее влияние на устойчивость экологических систем.
Следовательно, природные и общественные механизмы способны поддерживать экологическое равновесие и, следовательно, поддерживать динамический баланс между видами в экологических системах и между разными экологическими системами, в разных географических зонах.
Наряду с устойчивостью, экологическим системам присуща так называемая экологическая сукцессия, заключающаяся в смене сообществ организмов в ходе исторического развития природы, т. е. в замене одних сообществ растений и животных (их видов) в экосистеме растениями и животными других сообществ (видов).
Для экологической сукцессии, как исторического процесса, характерен ряд закономерностей. Прежде всего, процесс экологической сукцессии весьма длителен во времени, поскольку он заключается не в быстрой и внезапной замене одних сообществ другими, а в медленной и нерегулярной замене одних видов животных или растений в сообществах другими видами, но после того, как начался процесс изменения абиотических факторов, т. е. изменяются условия абиотической среды.
Условия, создаваемые организмами видов, начинающих сук-цессий первыми, благоприятствуют затем для внедрения в экологическую систему организмов других видов, которые, как правило, оказываются лучше адаптированными к изменившимся условиям. Благодаря этому они быстрее заменяют организмы, ставшие менее приспособленными к новым условиям существования. Таким образом, развитие новых экологических систем (в результате сукцессии) начинается с замены первичного сообщества растений и животных более совершенными сообществами растений и животных. В конечном итоге в экологической системе устанавливается постоянное сообщество, которое разрушается лишь при воздействии на экосистему каких-либо сильнодействующих факторов.
Сообщества организмов, которые в течение длительного периода существования экологических систем не сменяются на другие сообщества, называют климаксными. Те же сообщества, которые появляются в результате сукцессии, получили название сериальных.
Обычно сукцессии совершаются в течение длительных промежутков времени, составляющих сотни или тысячи лет. Однако если же экологические системы подвергаются внешним воздействиям, то сукцессии происходят быстрее. Можно сказать, что в результате внешних воздействий на экологическую систему нарушается экологическое равновесие, т. е. происходят изменения (нарушения) в природе.
Таким образом, нарушения в природе являются результатом нарушения экологического равновесия, разрушения исторически сложившихся экологических систем.
Изменения сообществ в результате сукцессии заключаются в изменении видового состава растений и животных. Видовое разнообразие в новых сообществах зависит от многих факторов, начиная от устойчивости отдельных видов К сукцессиям и вхождения их в новые сообщества и заканчивая растениями и животными, легко заселяющими территории, подвергавшиеся хозяйственному воздействию со стороны человека.
Вернемся к озеру, избранному в качестве примера экологической системы. Типичным примером сукцессии является подсыхание и загрязнение озера травянистой растительностью, превращение его в болото, а затем в неудобье, загрязненное кустарником.
Несомненно, что наряду со сменой растительности здесь происходит и смена животного мира. Очень иллюстративным, возможно, глобальным примером сукцессии является подсыхание Аральского моря, которое сопровождается появлением на месте бывшего морского дна новых сообществ растений и животных, включая мелких млекопитающих.
На примитивных стадиях своего развития человек был одним из равных видов (среди растений и животных) в экологических системах. По этой причине регулирующие механизмы в тогдашних экологических системах действовали так, как будто бы в них нет человека. Однако с тех пор, как человек стал важным, а затем и доминирующим видом в экологических системах, регулирующие механизмы стали ослабевать или совсем разрушаться. Причины заключаются в воздействии человека на биосферу, начала которых восходит к неолиту.
Однако на ранних этапах истории человека эти воздействия были незначительными. В последующем же они стали нарастать. Обратив на это внимание, В. И. Вернадский назвал ту часть биосферы, на которой особенно сильно сказывается деятельность человека, ноосферой.
Особенно прогрессирующий характер воздействий на биосферу отмечается в новейшее время, когда деятельность человека в биосфере по многим направлениям необратимо стала глобальной, когда его жизнь стала определяться потреблением и выбросами в гигантских размерах. Ниже показано, какими будут потребление и выбросы в ближайшее десятилетия на одного человека, начиная с 1972 г.
Предполагается Предполагается
потребить: выбросить
:
Пища — 50 т. Бутылки — 27 000
Вода — 98 280 000 л Бутылочные пробки —27 000
Железо и сталь — 52 т Мусор — 126 т
Бумага — 650 кг Использованные
Удобрения — 5000 кг автомобили — 2
Оценка непрерывно повышающихся все время темпов потребления и выбросов приводит к заключению, что сохранение имеющейся тенденции может даже увеличить показатели, приведенные выше.
Направлений воздействия человека на биосферу (антропоген-ных факторов) очень много. Воздействия человека на биосферу настолько значительны, что они, как считал В. И. Вернадский, создают новую оболочку Земли — ноосферу. Однако здесь мы рассмотрим лишь некоторые из этих направлений, обратив внимание на их «результативность» и на то, что человек еще не сумел изобрести механизмы, которые бы поддерживали стабильность в модифицированных им экологических системах, членом которых он является сам и которые обеспечивают его пищей и материалами, позволяющими дальнейшее развитие цивилизации.
Среди важнейших направлений в деятельности человека в биосфере следует назвать в первую очередь производство пищи, производство энергии, производство промышленных материалов и химический синтез, транспорт и хозяйственную деятельность. Особое значение имеет военная деятельность в виде войн и различных вооруженных конфликтов. Заслуживают особого внимания вопросы, касающиеся возможного использования ядерного оружия.
Производство пищи. В ходе своей истории человечество всегда сталкивалось с необходимостью обеспечения себя пищей, причем эта проблема решалась разными способами. В течение первых тысячелетий своей истории наши предки были хищниками и травоядными, а Земля в начальный период земледелия могла прокормить лишь 10 млн человек (рис. 219). Недостаток продовольствия сопровождается голодом.
Впервые голод возник еще в древнем Риме примерно 3,5 тыс. лет до н. э., а его вспышки сопровождают всю историю человечества, включая и наше время. В соответствии с существующими расчетами для обеспечения нормальной жизнедеятельности одного человека на протяжении 70 лет его жизни необходимо 50 т продовольствия со значительной долей белкового содержания. Сейчас население мира составляет свыше 6 млрд человек, но количества производимого белка достаточно для удовлетворения потребностей лишь половины мирового населения. Между тем по данным ООН численность населения Земли к 2020 г. превысит 7 млрд, а к 2025 г. — 8,46 млрд.
В частности, к 2025 г. население Китая составит 1,492 млрд человек, Индии — 1,445 млрд, Нигерии — 301 млн, США — 300 млн. По этой причине необходимо будет иметь продовольствия, как минимум, в два раза больше, чем сейчас.
Одно из традиционных направлений в производстве пищи заключается в рубке лесов и распахивании новых земель. Уже сейчас пахотные земли занимают 1,3 млрд гектаров (10% поверхности Земли). Однако неправильное распахивание земель ведет к эрозии почвы, к зарастанию ее сорняками. Больше того, введение в севооборот новых культур изменило содержание агроценозов, в результате чего в плодах некоторых культур стали обнаруживать ядовитые вещества. Например, в арахисе, зараженном Aspergillus flavus, обнаружен афлатоксин.
Для достижения высоких урожаев прибегают к обильному орошению посевов, но это ведет к засолению почв. Кроме того, прибегают к химическим удобрениям, вносимых в почву в больших количествах, но не использованные растениями удобрения с дождевыми или вешними водами выносятся в водоемы и «удобряют» их, вследствие чего в них усиленно размножается планктон, что изменяет водные экосистемы, ведет к экологической сукцессии с ее неблагоприятными последствиями.
Для защиты растений в сельском хозяйстве широко используют различные химические соединения в качестве пестицидов, гербицидов и дефолиантов, которых в мире сейчас производится более 2 млн тонн ежегодно.
Однако эти химические вещества, среди которых многие являются мутагенами, загрязняют среду (почву и воду), проникают в клетки растений и животных, а затем с пищей растительного и животного происхождения попадают в организм людей, вызывая нарушения в здоровье.
Химические вещества, загрязняющие почву, воду, атмосферу называют загрязнителями среды обитания.
Производство энергии. Чтобы обеспечить себя энергией (рис. 220), человек прибегает к добыче и сжиганию энергоносителей. В частности, с середины XIX в. началось бурное потребление каменного угля, а позднее и нефти. Однако при сжигании энергоносителей образуется множество веществ-загрязнителей, которые широко распределяются в биосфере, пересекая границы стран и континентов. Например, по некоторым подсчетам ежегодно в мире в результате сжигания жидкого и твердого топлива лишь на электростанциях, ТЭЦ и в домовых котельных в атмосферу выбрасывается около 200 млн тонн окислов азота.

В то же время в атмосфере уменьшается количество кислорода, но увеличивается доля углекислого газа.
Подсчитано, что за последние 100 лет концентрация углекислоты в атмосфере увеличилась на 12%, создавая так называемый парниковый эффект.
Если в дождевые воды попадают двуокись серы и окислы азота, образующиеся в результате сжигания угля и нефти, то они превращаются практически в разбавленные растворы серной и азотной кислот, которые с осадками в виде кислотных дождей или снега выпадают на Землю, где они становятся ядовитыми для животных, растений и человека.
Подсчитано, что за последние 100 лет количество кислотных осадков увеличилось более чем в 30 раз.
В результате производства энергии в атмосферу попадает также огромное количество частиц золы, которые содержат мутагенные и одновременно канцерогенные вещества (пирен, перилен и др.).
Предполагается, что к 2000 г. около 50% или более энергии будет производиться АЭС, а это позволяет предполагать, что биосфера и дальше будет загрязняться радиоактивными отходами. Особенно опасны аварии на АЭС. Итак, производство энергии неизбежно ведет к загрязнению биосферы.
Производство промышленных материалов и химический синтез. Это направление в деятельности человека сопровождается не только использованием невосполнимых запасов минеральных веществ и воды, в том числе питьевой, но и образованием при производстве материалов (стали, чугуна, цемента, тканей и т. д.) в огромных количествах различных химических соединений. В почву и воду попадают соли тяжелых металлов, а через пищу — и в организм человека.
Производство различных металлов, как и энергии, сопровождается также запылением атмосферы, особенно минеральной пылью и сульфатными аэрозолями. Например, цементная пыль содержит окислы кальция и магния.
Большое распространение получило производство бытовых аэрозолей, чистящих и моющих средств, а также химических соединений, придающих материалам водонепроницаемость и другие свойства. Их использование также сопровождается загрязнением среды обитания. Минеральные пыли способствуют повышению температуры в верхних слоях атмосферы, а также вызывают другие неблагоприятные последствия в тропосфере.
Опасность представляют хлорфторуглеводороды в связи с их использованием в холодильной промышленности. Они разрушают озоновый слой, защищающий живой мир от коротковолнового ультрафиолетового излучения. Особую опасность для атмосферы представляет насыщение ее углекислым газом, который создает так называемый «парниковый эффект».
Ежегодно в странах, имеющих химическую лабораторную базу и химическую промышленность, синтезируется несколько десятков тысяч новых химических соединений, из которых около 500 идет на рынок для использования в промышленности, сельском хозяйстве и медицине. Однако использование продуктов химического синтеза не всегда является благоприятным, что подтверждается использованием в практике ДДТ и ДЭС. ДДТ был синтезирован впервые еще в 1874 г., но начиная с 1930 г. его стали использовать в качестве пестицида и препарата против малярийных комаров. Однако, уже в 60-е годы стали замечать, что использование сопровождалось уменьшением численности птиц в Европе, а в 70-е годы было установлено, что он обладает «феминизирующим» воздействием на животных.
В частности, было установлено, что под воздействием этого соединения у аллигаторов уменьшается в размерах совокупительный орган, а воздействие метаболитов этого соединения на мужчин сопровождается снижением концентрации сперматозоидов в их эйякулятах, а также развитием крипторхизма. Детальное изучение механизмов действия ДДТ и других сходных химических соединений, загрязняющих окружающую среду, показало, что в организме эти соединения и продукты их распада, будучи по химической структуре непохожими на естественные эстрогены, все же действуют в качестве эстрогенов или блокаторов андрогенов. Кроме того, не вызывая структурных изменений в генах, они вызывают изменения в экспрессии генов.
ДЭС (диэтилстилбестрол) был синтезирован в 1938 г. и длительное время использовался в животноводстве для стимуляции роста крупного рогатого скота, а в медицинской практике — для предупреждения выкидышей.
Однако в 70-е гг. стали отмечать, что у девочек, родившихся от матерей, принимавших этот препарат, развивается вагинальный рак.
Было установлено также, что ДЭС тоже обладает эстрогенной активностью, сопровождающейся неблагоприятными последствиями для людей.
Помимо названных химических соединений известны и другие синтетические соединения, которые опасны не только тем, что они загрязняют окружающую среду, но и своим механизмом действия на человека и животных. Будучи непохожими по химической структуре на гормоны, они, тем не менее, мимикрируют сигнализирующие действия естественных гормонов. В результате этого такие химические соединения-загрязнители получили название средовых гормонов. Образно говоря, токсичность некоторых химических загрязнителей окружающей среды является результатом «естественного» сигнала, посланного «неестественной» молекулой.
Особого внимания заслуживает рассмотрение вопроса о химических соединениях, используемых в качестве пищевых добавок, поскольку в последние годы у многих сложилось представление о том, что применение пищевых добавок является одним из условий массового производства продуктов питания.
Пищевые добавки классифицируют на несколько групп, а именно:
а) Красители. Эти химические соединения используют для улучшения товарного вида мяса, овощей и фруктов.
б) Консерванты, антиокислители, стабилизаторы и эмульгаторы. Эти химические соединения используют для обеспечения сохранности продуктов питания в течение длительного времени при разных условиях хранения. Ими обрабатывают продукты как животного, так и растительного происхождения.
в) Усилители вкуса и аромата. Эти химические соединения широко используют в производстве продуктов как животного, так и растительного происхождения.
г) Актифламинги. Эти добавки представляют собой химические соединения, препятствующие образованию пены при разливе соков, а также слеживанию сахара, соли, муки и других сыпучих продуктов питания. Хотя биологическая эффективность многих добавок неизвестна, тем не менее, продукты питания с пищевыми добавками нельзя считать экологически чистыми. Наконец, химический синтез сопровождается бесконтрольным выбросом в среду побочных продуктов химии в огромных количествах, часть которых обладает мутагенными (канцерогенными) свойствами. Некоторые химические соединения в обычных условиях кажутся безвредными. Однако попав в организм, гидролируются там и превращаются в мутагены.
Транспорт. Одним из важнейших направлений в деятельности человека является развитие к использование транспорта с целью перевозки людей, промышленной и сельскохозяйственной продукции, сырья и т. д. Однако транспорт, особенно автомобильный, является загрязнителем биосферы. Воздушный транспорт также не безразличен дли биосферы. Например, авиалайнер Москва-Ныо-Йорк расходует за один рейс свыше 50 тонн кислорода. Но вред также заключается и в том, что транспортировка различных сырьевых материалов часто сопровождается их потерями, загрязняющими литосферу и гидросферу. Например, в последние годы сброшено в моря и океаны около 0,1% нефти, перевезенной от мест добычи к нефтеперегонным заводам. Загрязнение суши нефтью ведет к полному прекращению роста растений на ее поверхности.
Перечисленные здесь направления в деятельности человека уже привели к резким изменениям в биосфере, к нарушениям равновесия во многих экологических системах, что создало для людей новое физическое, химическое и биологическое окружение.
Диалектика преобразующей деятельности человека в собственных интересах заключается в том, что возникли новые противоречия между биологическими особенностями человека и созданными им в результате преобразующей деятельности факторами среды, многие из которых опасны для его здоровья, являясь мутагенами и канцерогенами, а также другими патогенетическими факторами.
Хозяйственная деятельность. С давних времен хозяйственная деятельность человека в природе всегда сопровождалась разрушением местообитаний и изменением численности видов животных и растений. Например, в период с 1600 г. по 1947 г. с лица Земли .исчезло 63 вида и 55 подвидов млекопитающих. Среди них следует назвать таких животных как морские коровы, которые обитали у Командорских островов, квагги (зебра), обитавшие в Южной Африке, странствующие голуби — эндемики Северной Африки.
Продолжающееся разрушение мест обитания животных создало угрозу еще для около 450 видов позвоночных, а продолжающаяся чрезмерная добыча и отлов угрожают очень многим видам млекопитающих, птиц и рыб.
Считают, что в результате хозяйственной деятельности в настоящее время главными угрожающими факторами для существования позвоночных являются разрушение их местообитания, ухудшение или ликвидация кормовой базы, уничтожение этих животных с целью защиты посевов, промысел.
Следует отметить также роль интродукции (внедрения) новых видов, которая не всегда благоприятна для исторически сложившихся биоценозов.
Например, вселение во внутренние водоемы нашей страны некоторых видов рыб, питающихся планктоном, сопровождалось резким изменением структуры пресноводных сообществ. В 1960 г- на Тайвань был переселен из Аргентины один из видов улиток как потенциальный источник белка. Улитки этого вида распространились и в другие районы Ю. Азии, что сопровождалось снижением урожая риса.
Во всех промышленно развитых странах ежедневно в пересчете на одного человека выбрасывается около 3 кг домашнего, строительного, уличного и другого мусора. В мире ежегодно один человек выбрасывает в среднем несколько десятков консервных банок, несколько десятков стеклянных бутылок, несколько килограммов бумаги (упаковочной, оберточной, газетной и др.). Все это чрезвычайно загрязняет биосферу.
Особое значение приобретают биологические загрязнители, попадающие в среду в результате недостаточной санитарной культуры людей в процессе их хозяйственной деятельности, с одной стороны, а также в результате экологической сукцессии, с другой. Примерами первого типа загрязнений является загрязнение среды гельминтами, патогенными микроорганизмами, плазмидами, а второго типа — появление на территориях, освобожденных в результате высыхания водоёмов, животных, являющихся резервуарами природно-очаговых болезней.
Военная деятельность. Самая большая угроза для биосферы и человечества связана с перспективой ядерной войны. Как отмечают в одном из своих докладов эксперты Всемирной федерации научных работников, если произойдут взрывы двух ядерных бомб мощностью около 5-10 тыс мт (по одной на территории каждого из возможных противников), то в результате только действия ударной волны сразу погибнет 750 млн. человек, а в результате совместного действия ударной волны, светового излучения и проникающей радиации будет уничтожено около 1,1 млрд человек и еще около 1,1 млрд человек получат ранения и будут нуждаться в медицинской помощи. Таким образом, 30-50% мирового населения станут непосредственными жертвами войны только лишь в случае взрыва двух ядерных бомб.
Однако в случае ядерного конфликта неизбежны и долговременные биологические последствия. Пыль и сажа, образующиеся в результате взрыва, поглотят и рассеят солнечный свет, понизят температуру воздуха и почвы. Над северным полушарием интенсивность света может упасть до 1% нормы, а температура может понизиться до -40°С. Доза радиации на площади, составляющей 30% суши, повысится до 500 рад. В последующие несколько недель после взрыва более чем на половине территории средних широт северного полушария радиоактивные осадки создадут внешнюю дозу облучения, превышающую 100 рад. Радиоактивные вещества будут отлагаться в щитовидных железах, костях, желудочно-кишечном тракте людей, в молоке матерей. После оседания пыли частично разрушиться слой озона окислом азота, образующимся при ядерном взрыве. В южном полушарии минимальный уровень освещенности составит 10% нормы, температура поверхности Земли достигнет —18°С, а ультрафиолетовое излучение будет выше нормы на десятки процентов в течение нескольких лет. Затемнение приведет к прекращению фотосинтеза, многие растения погибнут из-за недостатка света, что вызовет нарушения в цепях питания в экологических системах. Снижение температуры вызовет гибель зерновых культур, ибо, как известно, летом при наличии заморозков посевы пшеницы гибнут уже при 5°С, а рис и сорго не образуют семян при 15°С. Кукуруза очень чувствительна к температуре ниже 10°С.
Поскольку 30% площади суши на средних широтах получит дозу проникающей радиации не менее 50 рад, то при губительной дозе для человека в 300—500 рад за 48 часов это приведет к гибели еще около 1 млрд людей. Кроме того, сажа и пыль будут поглощать УФ-излучение, что приведет к повреждению иммунной системы, роговицы глаз и к катаракте у людей, остающихся в живых.
Воздействие низких температур, пожаров, радиации, сильных ветров будет сопровождаться распадом экологических систем, размножением вредителей лесов, полей, садов и огородов. Животные погибнут от голода, морозов и отсутствия воды. В результате миграции животных начнется распространение болезней человека и животных. В конечном итоге действие радиоактивных веществ приведет к гибели животного мира. Особенно чувствительными окажутся тропические леса, ибо у растений тропиков и субтропиков нет периода покоя, позволяющего им выдерживать температуры даже выше нуля.
Четкие представления о последствиях ядерной войны являются мощным фактором дальнейшей активизации антивоенных движений, борьбы за выживание человечества в ядерный век.
Современные представления об охране природы и среды обитания человека основаны на идеях В. И. Вернадского об охране биосферы. В современной трактовке речь идет, прежде всего, о предупреждении изменений в количествах лучистой энергии, достигающей Земли, о поддержании достаточной устойчивости химических циклов, протекающих в биосфере.
Охрана природы и среды обитания человека в наше время приобрела общественный интерес. Можно сказать, что взаимоотношение общества с окружающей средой — это одна из наиболее глобальных проблем человечества.
Понятия «охрана природы» и «охрана среды обитания человека» сложны и обширны. Охрана природы — это комплекс государственных, общественных и научных мероприятий, направленных на рациональное природопользование, восстановление и умножение естественных ресурсов Земли. Охрана среды обитания человека — это охрана всего того, что непосредственно окружает человека, что составляет экологические системы, членом которых он является, а также недопущение в среде обитания факторов, губительно действующих на его здоровье. Эти понятия во многом сходны между собой, ибо их стратегический смысл состоит в нахождении путей регулирования взаимоотношений человеческого общества и природы (живой и неживой). Тем не менее эти понятия имеют и существенные различия.
Охранять природу — это не означает сохранять ее в нетронутом виде, ибо человек и дальше будет эксплуатировать природные ресурсы, причем по мере роста народонаселения еще в большей мере.
Речь идет об охране, которая должна обеспечить установление равновесия между использованием и восстановлением, а также непрерывное поддержание мощности биосферы. Поэтому главные задачи всех природ оохранительных мероприятий заключаются в том, чтобы не нарушать количественные и качественные характеристики круговорота веществ и трансформации энергии, т. е. не изменять исторически сложившуюся биопродуктивность биосферы.
Напротив, должны осуществляться систематические разработки мероприятий, направленных на интенсификацию биологических круговоротов в естественных и искусственных экосистемах, т. е. на резкое повышение производительности Земли. В частности, необходимо создание подлинно научных основ увеличения плотности зеленого покрова Земли с большой долей видов, для которых характерен высокий коэффициент полезного действия фотосинтеза. С другой стороны, важно сохранять редкие и исчезающие виды животных.
Наконец, нельзя наполнять среду радиационными и химическими загрязнителями, вредными для животных и растений. Итак, генеральная линия в охране природы есть охрана и воспроизведение живого мира.
Говоря об охране среды обитания человека, важно помнить, что будучи составным компонентом биосферы, человек в ходе исторического развития адаптировался к своему окружению, но не биологически, а социально с помощью технических и культурных средств. Поэтому, как живое существо, человек открыт для действия на него загрязнителей среды обитания. Поддерживать гигиену среды обитания — это значит поддерживать экологическое равновесие между человеком и его окружением в целях обеспечения благополучия человека, его здоровья. Поэтому в наше время возникли вопросы не только определения ущерба, уже причиненного генофонду человека, но и определения путей защиты наследственного материала человека от факторов, порождаемых его деятельностью в биосфере.
Решение названных вопросов в разных странах идет по нескольким направлениям, главные из которых заключаются в создании чувствительных тест-систем для оценки мутагенной активности загрязнителей окружающей среды и в поисках подходов в эффективному слежению за генетическими процессами, протекающими в популяциях человека (разработка основ генетического мониторинга популяций). Смысл и необходимость этих работ заключается в интегральном анализе динамики генетического груза, т. е. в изучении и оценке частоты мутаций генов и хромосом, индуцированных загрязнителями, по отношению к мутациям, исторически накопленным в процессе эволюции, эволюционно сложившимся системам сбалансированного генетического полиморфизма.
В настоящее время для регистрации изменений в генетической структуре популяций человека используют несколько подходов.
Один из этих подходов связан с учетом популяционных характеристик. В качестве показателя оценки генетического груза используют медико-статистические показатели (частота спонтанных абортов, частота мертворождений, вес детей при рождении, вероятность выживания, соотношение полов, частота заболеваний врожденных и приобретенных, показатели роста и развития детей).
Другой подход связан с учетом «сторожевых» фенотипов, т. е. с определением фенотипов, возникающих благодаря определенным мутациям, унаследованным доминантно. Примером такого фенотипа является вывих тазобедренного сустава. В отобранной популяции ведется слежение за динамикой частоты интересующих фенотипов среди новорожденных, например, за динамикой частоты вывиха тазобедренного сустава.
Еще один подход связан с использованием электрофореза белков сыворотки крови и эритроцитов для выявления мутантных белков на основе их подвижности в электрическом поле, поскольку причиной изменения заряда белковой молекулы может быть замена или вставка одного или нескольких азотистых оснований в гене. Наконец, используют подход, связанный с цитогенетическим исследованием спонтанно абортированных эмбрионов, мертворожденных, живорожденных и детей с врожденными пороками.
Несомненно, что часть ущерба, уже нанесенного биосфере, невозможно восстановить. Поэтому перед человечеством стоит задача создать условия сбалансированного развития. Важнейшая задача заключается в создании таких технологий, которые бы исключали полностью или ограничивали выброс в окружающую среду загрязнителей.
Речь идет о таких технологиях как в промышленности, так и сельском хозяйстве.
Во многих странах имеются национальные программы охраны природы и окружающей среды. Эти программы основаны на учете специфики местных условий. Однако какие бы меры не принимались в отдельных странах, они не могут обеспечить решения всего комплекса вопросов, связанных с загрязнением атмосферы, открытых морей, Мирового океана. Поскольку биосфера неделима политически, а загрязнение среды обитания человека влечет за собой глобальные последствия, огромное значение имеет международное сотрудничество в области охраны природы и среды обитания человека.
Помимо решений вопросов на правительственных уровнях, большое значение имеет деятельность Международного союза охраны природы, Всемирного фонда охраны природы, а также специализированных учреждений ООН.
5 июня — Всемирный день охраны окружающей среды. В 1986 г. ВОЗ приняла Глобальную стратегию сохранения здоровья для всех к 2000 г. В соответствии с этой стратегией непременным условием для выполнения поставленных задач является сохранение и упрочнение мира на Земле. В наше время речь идет о сохранении жизни на Земле.
1. Сформулируйте определение биосферы и назовите ее подразделения.
2. Что является элементарной единицей биосферы?
3. Что собой представляет экологическая система?
4. Объясните взаимоотношения между организмами-производителями, организмами-потребителями и организмами-разрушителями в экосистемах.
5. Чем обеспечивается постоянство в поддержании экосистем?
6. Что такое пищевая цепь? Как много этих цепей в экосистемах?
7. Объясните связь между энергией, порядком и энтропией.
8. Расскажите о потоке энергии через пищевую цепь.
9. Что такое экологическая пирамида?
10. Объясните значение первого и второго законов термодинамики для мира живых существ.
11. Назовите основные формы внутривидовых отношений организмов.
12. Перечислите основные формы межвидовых отношений организмов и покажите место и роль человека в этих отношениях.
13. Что такое паразитизм и в чем заключается его отличие от хищничества? Каково его значение в природе?
14. Перечислите основные направления деятельности человека в природе. Назовите благоприятные и неблагоприятные последствия этой деятельности.
15. Есть ли разница между понятиями «окружающая среда» и «среда обитания»?
16. Каковы основные биологические последствия ядерной войны?
17. Почему важны международные усилия в охране окружающей среды?
Агаджанян Н. А., Торшин В. И. Экология человека. М.: Изд. фирма «Круг». 256 стр.
Красная книга СССР. М.: Лесная промышленность. 1978. 459 стр.
Реймерс Н. Ф. Охрана природы и окружающей среды: словарь и справочник. М.: Просвещение. 1992.
Реймерс Н. Ф. Экология. М.: Россия молодая. 1994. 363 стр. Brown J. Н. Macroecology. University of Chicago Press. 1995. 265 pp.

«Познание определяется тем,
что утверждается нами
как Истина»
П. А. ФЛОРЕНСКИЙ.
1923
Современная биология коренным образом отличается от традиционной биологии не только большей глубиной разработки познавательных идей, но и более тесной связью с жизнью общества, с практикой. Можно сказать, что в наше время биология стала средством преобразования живого мира с целью удовлетворения материальных потребностей общества. Это заключение иллюстрируется прежде всего тесной связью биологии с биотехнологией, которая стала важнейшей областью материального производства, равноправным партнером механической и химической технологий, созданных человеком ранее. Чем же объясняется взлет биотехнологии?
С момента своего возникновения биология и биотехнология всегда развивались совместно, причем с самого начала биология была научной основой биотехнологии. Однако длительное время недостаток собственных данных не позволял биологии оказывать очень большое влияние на биотехнологию. Положение резко изменилось с созданием во второй половине XX в. методологии генетической инженерии, под которой понимают генетическое манипулирование с целью "конструкции новых и реконструкции существующих генотипов. Являясь по своей природе методическим достижением, генетическая инженерия не привела к ломке сложившихся представлений о биологических явлениях, не затронула основных положений биологии подобно тому, как радиоастрономия не поколебала основных положений астрофизики, установление «механического эквивалента тепла» не привело к изменению законов теплопроводности, а доказательство атомистической теории вещества не изменило соотношений термодинамики, гидродинамики и теории упругости.
Генетическая инженерия открыла новую эру в биологии по той причине, что появились новые возможности для проникновения в глубь биологических явлений с целью дальнейшей характеристики форм существования живой материи, с целью более эффективного изучения структуры и функции генов на молекулярном уровне, понимания тонких механизмов работы генетического аппарата. Успехи генетической инженерии означают переворот в современном естествознании. Они определяют критерии ценности современных представлений о структурно-функциональных особенностях молекулярного и клеточного уровней живой материи. Современные данные о живом имеют гигантское познавательное значение, ибо обеспечивают понимание одной из важнейших сторон органического мира и тем самым вносят неоценимый вклад в создание научной картины мира. Таким образом, резко расширив свою познавательную базу, биология через генетическую инженерию оказала также ведущее влияние на подъем биотехнологии.
Генетическая инженерия создает заделы на пути познания способов и путей «конструирования» новых организмов или улучшения существующих организмов, придавая им большую хозяйственную ценность, большую способность резкого увеличения продуктивности биотехнологических процессов.
В рамках генетической инженерии различают генную инженерию и клеточную инженерию. Под генной инженерией понимают манипуляции с целью создания рекомбинантных молекул ДНК. Часто эту методологию называют молекулярным клонированием, клонированием генов, технологией рекомбинантных ДНК или просто генетическими манипуляциями. Важно подчеркнуть, что объектом генной инженерии являются молекулы ДНК, отдельные гены. Напротив, под клеточной инженерией понимают генетические манипуляции с изолированными отдельными клетками или группами клеток растений и животных.
Генную инженерию составляет совокупность различных экспериментальных приемов (методик), обеспечивающих конструкцию (реконструкцию) и клонирование молекул ДНК (генов) с заданными целями.
Методы генной инженерии используют в определенной последовательности (рис. 221), причем различают несколько стадий в выполнении типичного генно-инженерного эксперимента, направленного на клонирование какого-либо гена, а именно:
1. Выделение ДНК из клеток интересующего организма (исходного) и выделение ДНК-вектора.
2. Разрезание (рестрикция) ДНК исходного организма на фрагменты, содержащие интересующие гены, с помощью одного из ферментов-рестриктаз и выделение этих генов из образованной рестрикционной смеси. Одновременно разрезают (рестрикциируют) векторную ДНК, превращая ее из кольцевой структуры в линейную.
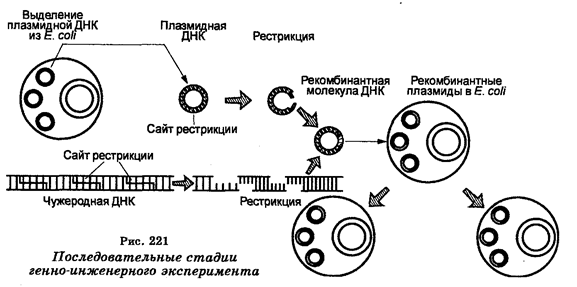
3. Смыкание интересующего сегмента ДНК (гена) с ДНК вектора с целью получения гибридных молекул ДНК.
4. Введение гибридных молекул ДНК путем трансформации в какой-либо другой организм, например, в Е. coli или в соматические клетки.
5. Высев бактерий, в которые вводили гибридные молекулы ДНК, на питательные среды, позволяющие рост только клеток, содержащих гибридные молекулы ДНК.
6. Идентификация колоний, состоящих из бактерий, содержащих гибридные молекулы ДНК.
7. Выделение клонированной ДНК (клонированных генов) и ее характеристика, включая секвенирование азотистых оснований в клонированном фрагменте ДНК.
ДНК (исходная и векторная), ферменты, клетки, в которых клонируют ДНК — все это называют «инструментами» генной инженерии.
Рассмотрим методику выделения ДНК на примере ДНК плаз-мид. ДНК из плазмидосодержащих бактериальных клеток выделяют с помощью традиционной техники, заключающейся в получении клеточных экстрактов в присутствии детергентов и последующем удалении из экстрактов белков фенольной экстракцией (рис. 222). Полная очистка плазмидной ДНК от белков, РНК и других соединений проводится в несколько стадий. После того как клетки разрушены, например, с помощью лизоцима (растворены их стенки), к экстракту добавляют детергент, чтобы растворить мембраны и инактивировать некоторые белки. Большинство хромосомной ДНК удаляют из получаемых препаратов обычным центрифугированием.
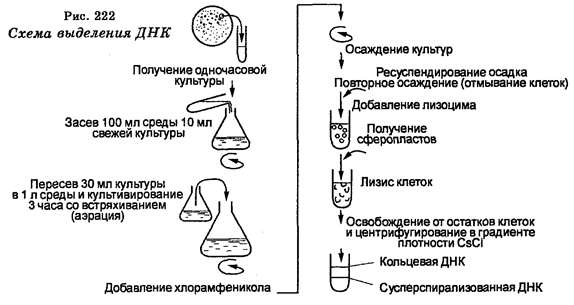
 Часто для полной очистки используют хроматографию. Если требуется очень тщательная очистка, используют высокоскоростное центрифугирование в градиенте плотности CsCI с использованием этидия бромида. Оставшаяся хромосомная ДНК будет фрагментирована в линейную, тогда как плазмидная ДНК останется ковалентно закрытой. Поскольку этидий бромид менее плотен, чем ДНК, то при ультрацентрифугировании в центрифужной пробирке будет «выкручиваться» два кольца — плазмидная ДНК и хромосомная ДНК (рис. 223). Плазмидную ДНК отбирают для дальнейшей работы, хромосомную ДНК выбрасывают. Часто для полной очистки используют хроматографию. Если требуется очень тщательная очистка, используют высокоскоростное центрифугирование в градиенте плотности CsCI с использованием этидия бромида. Оставшаяся хромосомная ДНК будет фрагментирована в линейную, тогда как плазмидная ДНК останется ковалентно закрытой. Поскольку этидий бромид менее плотен, чем ДНК, то при ультрацентрифугировании в центрифужной пробирке будет «выкручиваться» два кольца — плазмидная ДНК и хромосомная ДНК (рис. 223). Плазмидную ДНК отбирают для дальнейшей работы, хромосомную ДНК выбрасывают.
В ходе эволюции бактерии развили способность синтезировать так называемые рестрицирующие ферменты (эндонуклеазы), которые стали частью клеточной (бактериальной) системы рестрикции-модификации. У бактерий системы рестрикции-модификации являются внутриклеточной иммунной системой защиты от чужеродной ДНК. В отличие от высших организмов, у которых распознание и разрушение вирусов, бактерий и других патогенов происходит внеклеточно, у бактерий защита от чужеродной ДНК (ДНК растений и животных, в организме которых они обитают) происходит внутриклеточно, т. е. тогда, когда чужеродная ДНК проникает в цитоплазму бактерий. С целью защиты бактерии в ходе эволюции развили также способность «метить» собственную ДНК метилирующими основаниями на определенных последовательностях. По этой причине чужеродная ДНК из-за отсутствия в ней метальных групп на тех же последовательностях плавится (разрезается) на фрагменты разными бактериальными рестриктазами, а затем деградируется бактериальными экзонуклеазами до нуклео-тидов. Можно сказать, что таким образом бактерии защищают себя от ДНК растений и животных, в организме которых они обитают временно (как патогены) или постоянно (как сапрофиты).
Рестриктазы впервые были выделены из Е. coli в 1968 г- Оказалось, что они способны разрезать (плавить) молекулы ДНК на разных сайтах (местах) рестрикции. Эти ферменты получили название эндонуклеаз класса I. Затем у бактерий были обнаружены эндонуклеазы класса II, которые распознают в чужеродной ДНК сайты рестрикции специфически и на этих сайтах тоже осуществляют рестрикцию. Именно ферменты этого класса стали использовать в генной инженерии. Тогда же были открыты ферменты класса III, которые плавят ДНК рядом с сайтами распознания, но эти ферменты не имеют значения в генной инженерии.
Действие системы рестрикции-модификации «рационализуется» так называемыми палиндромными (распознающими последовательностями) азотистых оснований, которые являются сайтами рестрикции ДНК. Палиндромные последовательности — это последовательности оснований, которые одинаково читаются вперед и назад, как, например, последовательность букв радар. Поскольку цепи ДНК обладают антипараллельным направлением, то считают, что последовательность является палиндромной, если она идентична, когда читается в направлении от 5'- к 3'-концу на верхней и 3'- к о'-концу на нижней цепи, а именно:
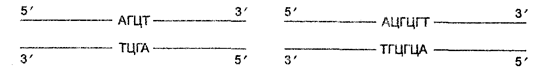
Палиндромы могут быть любых размеров, но большинство тех палиндромов, которые используют в качестве сайтов узнавания рестриктазами, состоят из 4, 5, 6 и реже 8 оснований.
Рестриктазы — это абсолютно необходимый инструмент в генной инженерии для вырезания интересующих фрагментов (генов) из больших молекул ДНК. Поскольку известно более 100 ферментов рестрикции, то это позволяет выбор рестриктаз и селективное вырезание фрагментов из исходной ДНК.
Замечательной особенностью рестриктаа является то, что они продуцируют разрезы молекул на несколько фрагментов (рестрик-тов) ДНК уступами, в результате чего в образующихся концах одна цепь длиннее другой, образуя своеобразный хвост. Такие концы (хвосты) получили название «липких» концов (рис. 224), т. к. они способны к самокомплементарности.
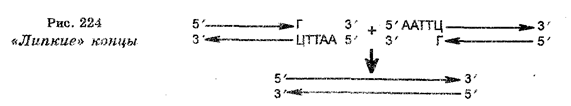
Рассмотрим результаты рестрикции на примере одной из наиболее известных рестриктаз Eco RI из системы рестрикция—модификация Е. coli. Вместо того, чтобы плавить ДНК в центре па-линдромной последовательности узнавания, этот фермент плавит ДНК за пределами центра и продуцирует 4 самокомплементарных («липких») конца, состоящих из разного количества нуклео-тидов, а именно:
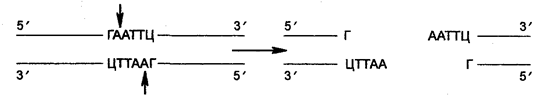
Эти «липкие» концы в генно-инженерных опытах полезны по той причине, что они могут быть воссоединены комплементарно при низких температурах, что позволяет эффективное смыкание ДНК-фрагментов.
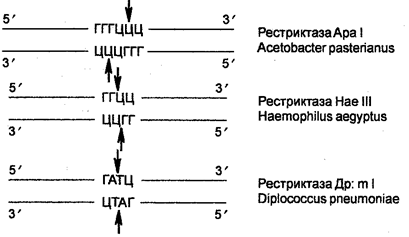
Сайты распознания и сайты плавления в случае других рестриктаз имеют другое содержание, а именно:
Вслед за рестрикцией ДНК из рестрикционной смеси выделяют рестрикционные ДНК-фрагменты (ДНК-рестрикты), которые необходимы затем для объединения с вектором. Для выделения ДНК-рестриктов прибегают к электрофорезу, поскольку с помощью этого метода рестрикцированную ДНК очень легко фракционировать благодаря размерам фрагментов-рестриктов и благодаря константным отношениям электрический заряд-масса. Фрагменты в электрическом поле мигрируют в ходе электрофореза при частоте, зависимой от их размеров (массы). Чем больше (длиннее) фрагмент, тем медленнее он мигрирует в электрическом поле. Материалом, в котором проводят электрофорез, являются незаряжающиеся агаро-за и полиакриламид. Для опознания фрагментов используют эти-дий бромид, который красит фрагменты, что ведет к их более легкому обнаружению (рис. 225).
 Результативность электрофореза очень высока, поскольку с его помощью могут быть разделены фрагменты, размеры которых составляют от 2 до 50 000 оснований. Результативность электрофореза очень высока, поскольку с его помощью могут быть разделены фрагменты, размеры которых составляют от 2 до 50 000 оснований.
После электрофореза фрагменты из агарозы выделяют с помощью разных методов. На основании результатов сравнения размеров рестриктов одной и той же ДНК, полученных с помощью разных рестриктаз, строят рестрикционные карты, на которых показывают сайты рестрикции каждой из использованных рестриктаз (см. § 107). В практическом плане рестрикционные карты позволяют определять не только размеры рестриктов, но и выяснять расположение в молекулах ДНК локусов тех или иных генов.
Поскольку у высших организмов в ходе транскрипции синтезируется гетерогенная ДНК, корректируемая процессингом, то в генной инженерии обычно используют комплементарную ДНК (кДНК), которую получают при использовании в качестве матрицы мРНК, на которой обратная транскриптаза синтезирует одноцепочечную ДНК (кДНК), являющуюся копией мРНК. В последующем эти одноцепочечные ДНК превращают в двухцепочечные ДНК. Считают, что кДНК содержит непрерывные нуклеотидные последовательности (транскрибируемые и транслируемые). Именно кДНК используют для рестрикции.
Выделенные после электрофореза из агарозных гелей фрагменты ДНК (рестрикты) можно предварительно подвергнуть секвени-рованию, т. е. определить в них нуклеотидную последовательность. Для этого используют химический и ферментативный методы сек-венирования.
Химический метод основан на получении меченных радиоактивным фосфором (32р) фрагментов и удалении из этих фрагментов одного из оснований с последующим учетом результатов радиоавтографии гелей, содержащих эти фрагменты. Ферментативный метод основан на том, что в конец анализируемого фрагмента вводят нуклеотид, используемый затем в синтезе разных фрагментов in vitro, анализируемых на нуклеотидную последовательность электрофоретически. Для изучения специфических последовательностей нуклеотидов в молекуле ДНК используют также гибридизацию ДНК-ДНК, РНК-РНК, ДНК-РНК, Нозерн- и Саузерн-блоттинги.
Сегмент ДНК (ген), который предназначен для молекулярного клонирования, должен обладать способностью к репликации при переносе его в бактериальную клетку, т. е. быть репликоном. Однако он такой способностью не обладает. Поэтому, чтобы обеспечить перенос и обнаружение клонируемых генов в клетках, их объединяют с так называемыми генетическими векторами. Последние должны обладать, как минимум, двумя свойствами. Во-первых, векторы должны быть способны к репликации в клетках, причем в нескольких копиях. Во-вторых, они должны обеспечивать возможность селекции клеток, содержащих вектор, т. е. обладать маркером, на который можно вести контрселекцию клеток, содержащих вектор вместе с клонируемым геном (рекомбинантные молекулы ДНК). Таким требованиям отвечают плазмиды и фаги. Плазмиды являются хорошими векторами по той причине, что они являются репликонами и могут содержать гены резистентности к какому-либо антибиотику, что позволяет вести селекцию бактерий на устойчивость к этому антибиотику и, следовательно, легкое обнаружение рекомбинантных молекул ДНК.
Поскольку природные плазмидные векторы неизвестны, то все известные к настоящему времени плазмидные векторы были сконструированы искусственно. Исходным материалом для создания ряда генетических векторов послужили R-плазмиды, в которых с помощью рестриктаз удаляли излишние последовательности ДНК, в том числе те, на которых располагались множественные сайты рестрикции. Это удаление определялось тем, что плазмидный вектор должен обладать только одним сайтом узнавания для одной рестриктазы, причем этот сайт должен лежать в функционально несущественном районе плазмидного генома. Например, плазмидный вектор pBR 322, который содержит гены резистентности к ампициллину и тетрациклину, что делает его очень удобным для селекции бактерий, содержащих клонируемый сегмент ДНК, обладает одиночными сайтами рестрикции для более 20 ферментов-рестриктаз, включая такие известные рестриктазы, как Eco R I, Hind III, Pst I, Pva II и Sal I (рис. 226).
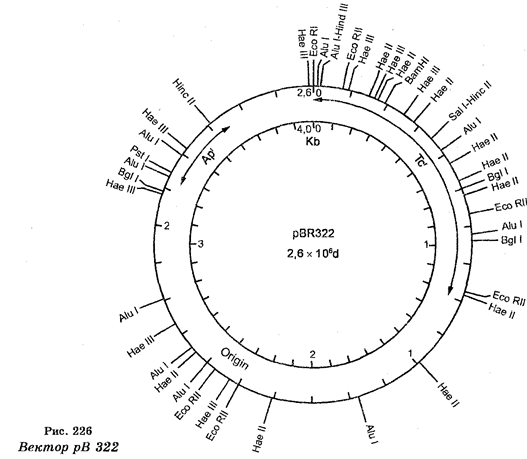
Фаговые векторы тоже обладают рядом преимуществ. Они могут включать в себя более крупные (более длинные) клонируемые фрагменты ДНК по сравнению с плазмидными векторами. Далее, перенос фагами клонируемого фрагмента в клетки в результате ин-фицирования ими последних является более эффективным, чем трансформация ДНК. Наконец, фаговые векторы позволяют более эффективный скрининг (распознание) на поверхности агара колоний, содержащих клетки, несущие клонируемый ген. Многие фаговые векторы сконструированы на базе фага лямбда.
Кроме фаговых используют и другие вирусные векторы, сконструированные на базе вируса герпеса, а также векторы, сконструированные на базе дрожжевой ДНК.
Если клонирование генов проводят, используя клетки млекопитающих или растений, то требования к векторам те же, что и в случае клонирования в бактериальных клетках.
Непосредственное конструирование рекомбинантных молекул ДНК следует после того, как получены рестрикты исследуемой ДНК и векторной ДНК. Оно заключается в смыкании сегментов-рест-риктов исследуемой ДНК с рестриктом векторной ДНК, которая в результате рестрикции превращается из кольцевой в линейную ДНК.
Чтобы сомкнуть фрагменты исследуемой ДНК с ДНК-вектора, используют ДНК-лигазу (рис. 227). Лигирование будет успешным, если смыкаемые структуры обладают 3'-гидроксил и 5'-фосфатной группами и если эти группы расположены соответствующим образом  одна относительно другой. Фрагменты объединяются через их «липкие» концы в результате са-мокомплементарности. При высоких концентрациях фрагментов последние время от времени становятся в правильное положение (напротив друг друга). Многие ре-стриктазы, такие как Eco R I, продуцируют «липкие» концы, состоящие из 4-х оснований. Процесс лигирования «липких» концов, состоящих из четырех оснований, происходит при пониженной температуре (до 12°С). одна относительно другой. Фрагменты объединяются через их «липкие» концы в результате са-мокомплементарности. При высоких концентрациях фрагментов последние время от времени становятся в правильное положение (напротив друг друга). Многие ре-стриктазы, такие как Eco R I, продуцируют «липкие» концы, состоящие из 4-х оснований. Процесс лигирования «липких» концов, состоящих из четырех оснований, происходит при пониженной температуре (до 12°С).
Если при рестрикции образуются фрагменты без «липких» концов, то их «насильственно» конвертируют в молекулы с
«липкими» концами, используя фермент трансферазу. Этот фермент добавляет нуклеотиды к 3'-концу ДНК. На одном фрагменте может быть добавлен поли-А-хвост, на другом — поли-Т-хвост. Для генерации любых желаемых концов ДНК используют также так называемую полимеразную цепную реакцию (ПНР). Принцип ПЦР основан на денатурации выделенной из клеток ДНК и «отжиге» ее с добавлением к ренатурирующимся цепям ДНК-олигонуклеотидов, состоящих из 15—20 нуклеотидов каждый. Эти олигонуклеотиды должны быть комплементарны последовательностям в целях, разделенных расстояниями в 50-2000 нуклеотидов. Будучи «затравкой» для синтеза ДНК in vitro, они позволяют ДНК-полимеразе копировать те участки, которые находятся между «затравками». Это копирование дает большое количество копий изучаемого фрагмента ДНК.
После смыкания интересующего фрагмента ДНК (гена) с генетическим вектором с помощью ДНК-лигазы образованные реком-бинантные молекулы вводят в клетки с целью добиться их репликации (за счет генетического вектора) и увеличения количества копий. Наиболее популярным способом введения в клетки рекомбинантных молекул ДНК, в которых вектором служит плазмида, является трансформация Е. coli. С этой целью бактериальные клетки предварительно обрабатывают кальцием или рубидием (ионами) для того, чтобы они стали «компетентными» в восприятии рекомби-нантной ДНК.
Чтобы повысить частоту проникновения ДНК в клетки, используют метод электропорации, заключающийся в кратком экспонировании клеток в интенсивном электрическом поле. Эта обработка создает полости в мембранах клеток, что способствует лучшему восприятию клетками ДНК- После введения рекомбинантных молекул ДНК в бактерии последние высевают на МПА, обогащенный антибиотиками для селекции желаемых клеток, т. е. клеток, содержащих рекомбинантные молекулы ДНК. Частота трансформации является невысокой. Обычно один трансформант возникает на 106
высеянных клеток. Если же вектор является фаговым, то прибегают к трансфекции клеток (бактерий или дрожжей) фагом. Что касается соматических клеток животных, то их трансфекцию осуществляют ДНК в присутствии химических веществ, облегчающих прохождение ДНК через плазматические мембраны. Возможны также прямые микроинъекции ДНК в овоциты лягушек, в культивируемые соматические клетки и в эмбрионы млекопитающих.
 Важнейшим моментом, связанным с молекулярным клониро-ванием, является поиск способа, позволяющего установить, действительно ли клонируемый фрагмент включился в вектор и вместе с вектором, образовав рекомбинантную молекулу ДНК, вошел в клетки. Если речь идет о бактериальных клетках, то один из способов основан на учете инсерционной инактивации плазмидного (векторного) гена резистентности. Например, в плазмидном векторе pBR322, детерминирующем резистентность к ампициллину и тетрациклину, единственный сайт для рестриктазы Pst I находится в локусе, занимаемом геном резистентности к ампициллину. Pst I —плавление на этом сайте генерирует липкие концы, позволяющие лигирование клонируемого фрагмента с векторной ДНК. Однако при этом плазмидный (векторный) ген ампициллинрезис-тентности инактивируется, тогда как ген тетрациклинрезистентнос-ти на векторе остается интактным. Именно, ген тетрациклинрезис-тентности и используется для селекции клеток, трансформируемых рекомбинантными молекулами ДНК. Чтобы убедиться, что клетки выросших колоний на среде с тетрациклином действительно содержат рекомбинантные молекулы ДНК, их проверяют с помощью так называемого «спот-теста» на паре чашек с плотной средой, одна их которых содержит ам-пициллин, тогда как другая лишена этого антибиотика. Клонируемые ДНК содержатся лишь в трансформантах, резистентных к тетрациклину (рис. 228). Что касается трансформантов, резистентных одновременно к ампициллину и тетрациклину (АрТс), то они содержат плазмидные (векторные) молекулы, которые спонтанно приобрели кольцевую форму без включения в них чужеродной (клонируемой) ДНК. Другой способ обнаружения инсерции чужеродных (клонируемых) фрагментов в плазмидный вектор основан на использовании вектора, содержащего ген (3-га-лактозидазы. Инсерция чужеродной ДНК в этот ген неизбежно инак-тивирует синтез р-галактозидаза, что может быть обнаружено посевом трансформированных клеток на среду, которая содержит субстраты р-галактозидазы. Эта среда позволяет селекцию окрашенных колоний клеток. Важнейшим моментом, связанным с молекулярным клониро-ванием, является поиск способа, позволяющего установить, действительно ли клонируемый фрагмент включился в вектор и вместе с вектором, образовав рекомбинантную молекулу ДНК, вошел в клетки. Если речь идет о бактериальных клетках, то один из способов основан на учете инсерционной инактивации плазмидного (векторного) гена резистентности. Например, в плазмидном векторе pBR322, детерминирующем резистентность к ампициллину и тетрациклину, единственный сайт для рестриктазы Pst I находится в локусе, занимаемом геном резистентности к ампициллину. Pst I —плавление на этом сайте генерирует липкие концы, позволяющие лигирование клонируемого фрагмента с векторной ДНК. Однако при этом плазмидный (векторный) ген ампициллинрезис-тентности инактивируется, тогда как ген тетрациклинрезистентнос-ти на векторе остается интактным. Именно, ген тетрациклинрезис-тентности и используется для селекции клеток, трансформируемых рекомбинантными молекулами ДНК. Чтобы убедиться, что клетки выросших колоний на среде с тетрациклином действительно содержат рекомбинантные молекулы ДНК, их проверяют с помощью так называемого «спот-теста» на паре чашек с плотной средой, одна их которых содержит ам-пициллин, тогда как другая лишена этого антибиотика. Клонируемые ДНК содержатся лишь в трансформантах, резистентных к тетрациклину (рис. 228). Что касается трансформантов, резистентных одновременно к ампициллину и тетрациклину (АрТс), то они содержат плазмидные (векторные) молекулы, которые спонтанно приобрели кольцевую форму без включения в них чужеродной (клонируемой) ДНК. Другой способ обнаружения инсерции чужеродных (клонируемых) фрагментов в плазмидный вектор основан на использовании вектора, содержащего ген (3-га-лактозидазы. Инсерция чужеродной ДНК в этот ген неизбежно инак-тивирует синтез р-галактозидаза, что может быть обнаружено посевом трансформированных клеток на среду, которая содержит субстраты р-галактозидазы. Эта среда позволяет селекцию окрашенных колоний клеток.
Как уже отмечено, рестрикционные линейные фрагменты векторной ДНК способны к восстановлению кольцевой структуры без включения в них клонируемых сегментов. Чтобы уменьшить частоту спонтанного образования таких кольцевых молекул векторной ДНК, рестрикты векторной ДНК обрабатывают фосфатазой. В результате этого образование кольцевых молекул ДНК становится невозможным, поскольку будут отсутствовать концы 5'-РО^, необходимые для действия лигазы.
Совокупность колоций-трансформантов, выросших на селективной среде, представляет собой совокупность клеток, содержащих клоны разных фрагментов (генов) клонируемой геномной или кДНК. Коллекции этих клонов формируют так называемые библиотеки ДНК, широко используемые в генно-инженерных работах.
Заключительной стадией клонирования генов является выделение и исследование клонированной ДНК, включая секвениро-вание.
Перспективные штаммы бактерий или соматических клеток, содержащих рекомбинантные молекулы ДНК, которые контролируют синтез интересующих белков, имеющих коммерческую ценность, передают в промышленность.
1. Что понимают под генной, клеточной и генетической инженерией? Есть ли разница между этими понятиями и понятием о молекулярном клонировании?
2. В чем заключается прогрессивность генетической инженерии по сравнению с другими методами, используемыми в биологии?
3. Перечислите основные «инструменты» генной инженерии.
4. Что представляют собой ферменты-рестриктазы, каковы их свойства и какова их роль в генной инженерии?
5. Все ли рестриктазы образуют «липкие» концы исследуемой ДНК и зависит ли структура «липких» концов от вида рестриктазы?
6. Дайте определение генетическим векторам. Существуют ли природные векторы?
7. Как получают генетические векторы в лабораторных условиях? Какие биологические объекты являются исходным материалом для получения векторов?
8. Какова предельная длина последовательностей азотистых оснований ДНК, которые еще могут включиться в генетический вектор? Различаются ли векторы по «мощности»?
9. Охарактеризуйте свойства ДНК-лигазы и определите ее роль в генной инженерии.
10. Как смыкают клонируемый сегмент ДНК (ген) с генетическим вектором?
11. Какова частота введения рекомбинантных молекул ДНК в бактериальные клетки?
12. На каком принципе основана селекция бактериальных клеток, содерасащих рекомбинантные молекулы ДНК? Приведите один из примеров такой селекции.
13. Можно ли считать штамм Е. coli, содержащий ген инсулина человека, новым видом бактерий?
14. Многие штаммы бактерий обладают одинаковыми ферментами, практически одинаково обеспечивающими их метаболизм. Между тем нуклеотидная специфичность систем рестрикции-модификации бактерий различна. Можете ли вы объяснить это явление?
15. Почему последовательности ДНК, представляющие собой сайты распознания рестриктазами, не могут содержать более восьми пар оснований?
16. Сколько раз последовательность ГГЦЦ, распознаваемая рестриктазой Нае III, будет встречаться в сегменте ДНК длиной в 50 000 пар оснований с 30%-, 50%- и 70%-процентным содержанием ГЦ?
17. Рестриктазы Ват Н I и Bgl I плавят последовательности Г-ГАТЦЦ и Т-ГАТЦА, соответственно. Можно ли включить в сайты Barn H I фрагменты ДНК, продуцированные Bgl 1-рестрикцией? Если да, то почему? Если используемая плазмида (вектор) содержит один сайт для рестрикции Barn H I, то на какой питательной среде можно осуществить селекцию бактерий, содержащих эту плазмиду?
18. Вычислите частоту трансформации бактерий на одну молекулу ДНК, если на 6000 плаамидных пар оснований образуется б х 106
трансформантов?
19. Можно ли клонировать 0-пункт репликации ДНК Е. coli и если да, то каким образом?
20. Можно ли определить, сколько необходимо молекул ДНК для трансформации одной клетки Е. coli?
21. Можно ли с помощью полимеразной цепной реакции определить сайт сплайсинга на мРНК?
22. Каким образом можно использовать полимеразную цепную реакцию для того, чтобы ввести желаемый сайт рестрикции в интересующее место на фрагменте ДНК, предназначенном для клонирования?
Альберте В., Брей Д., Льюис Дж.. Рэфф М., Роберте К., Уотсон Дж. Молекулярная биология клетки. М.: Мир. 3.984. Т. 1. 615 стр.
Маниатж Т., Фрич Э„ Сзмбрук Дж, Молекулярное клонирование. М.: Мир. 1984. 479 стр.
Пехов А. П. Основы плазмидологии. М.: РУДН. 1996. 229 стр. Hunter Ceuera J„ Belt A. Maintaining Cultures for Biotechnology and Industry. Academic Press. 1995. 242 pp.
Kaplitt M; Loewy A. Viral Vectors. Academic Press. 1995. 486 pp. Russo E., Cove D. Genetic Engineering. WH Preeman. 1995. 255 pp.
Как отмечено в начале раздела, клеточной инженерией называют генетические манипуляции с изолированными клетками животных и растений. Эти манипуляции часто осуществляют in vitro, а главной целью они имеют получение генотипов этих организмов с заданными свойствами, в первую очередь, хозяйственно полезными. Что касается человека, то клеточная инженерия оказалась применимой к его половым клеткам.
Предпосылкой к развитию клеточной инженерии у человека и животных явилась разработка методов культивирования их соматических клеток на искусственных питательных средах, а также получение гибридов соматических клеток, включая межвидовые гибриды. В свою очередь, успехи в культивировании соматических клеток оказали влияние на изучение половых клеток и оплодотворение у человека и животных. Начиная с 60-х гг., в нескольких лабораториях мира были выполнены многочисленные эксперименты по пересадке ядер соматических клеток в яйцеклетки, искусственно лишенные ядер. Результаты этих экспериментов часто были противоречивы, но в целом они привели к открытию способности клеточных ядер обеспечивать нормальное развитие яйцеклеток (см. гл. VIII).
На основе результатов изучения развития оплодотворенных яйцеклеток в 60-е гг. были начаты также исследования по выяснению возможности оплодотворения яйцеклеток вне организма матери. Очень быстро эти исследования привели к открытию возможности оплодотворения яйцеклеток сперматозоидами в пробирке и дальнейшего развития образованных таким путем зародышей при имплантации их в матку женщины. Дальнейшее совершенствование разработанных в этой области методов привело к тому, что рождение «пробирочных» детей стало реальностью. Уже к 1981 г. в мире было рождено 12 детей, жизнь которым была дана в лаборатории, в пробирке. В настоящее время этот раздел клеточной инженерии получил большое распространение, а количество «пробирочных» детей составляет уже десятки тысяч. В нашей стране работы по получению «пробирочных» детей были начаты в 1986 г. В 1993 году была разработана методика получения монозиготных близнецов человека in vitro, путем разделения эмбрионов на блас-томеры и доращивания последних до 32 клеток, после чего они могли быть имплантированы в матку женщины.
Под влиянием результатов, связанных с получением «пробирочных» детей, у животных тоже была разработана технология, получившая название трансплантации эмбрионов. Она связана с разработкой способа индукции полиовуляции, способов искусственного оплодотворения яйцеклеток и имплантации зародышей в организм животных — приемных матерей. Суть этой технологии сводится к следующему. Высокопродуктивной корове вводят гормоны, в результате чего наступает полиовуляция, заключающаяся в созревании сразу 10—20 клеток. Затем яйцеклетки искусственно оплодотворяются мужскими половыми клетками в яйцеводе. На 7-8-й день зародышей вымывают из матки и трансплантируют в матки другим коровам (приемным матерям), которые затем дают жизнь телятам-близнецам. Телята наследуют генетический статус своих подлинных родителей.
Другой областью клеточной инженерии у животных является получение трансгенных животных. Наиболее простой способ получения таких животных заключается во введении в яйцеклетки исходных животных линейных молекул ДНК. Животные, развившиеся из оплодотворенных таким образом яйцеклеток, будут содержать в одной из своих хромосом копию введенного гена. Больше того, они и будут передавать этот ген по наследству. Более сложный способ получения трансгенных животных разработан на мышах, различающихся по окраске шерстного покрова и сводится к следующему. Вначале из организма беременной серой мыши извлекают четырехдневных зародышей и измельчают их на отдельные клетки. Затем из эмбриональных клеток извлекают ядра, переносят их в яйцеклетки черных мышей, предварительно лишенные ядер. Яйцеклетки черных мышей, содержащие чужие ядра, помещают в пробирки с питательным раствором для дальнейшего развития. Развившиеся из яйцеклетки черных мышей зародыши имплантируют в матки белых мышей. В выполненных по этой методике экспериментах от пяти белых мышей («приемных матерей») было получено 36 мышей, среди которых трое были серыми. Таким образом, в этих экспериментах удалось получить клон мышей с серой окраской шерстного покрова, т. е. клонировать эмбриональные клетки с заданными свойствами. В § 35 мы рассмотрели результаты оплодотворения искусственно лишенных ядер яйцеклеток овец ядерным материалом соматических клеток животных этого же вида. В частности, из яйцеклеток овец удаляли ядра, а затем в такие яйцеклетки вводили ядра соматических клеток (эмбриональных, плодовых или клеток взрослых животных), после чего оплодотворенные таким образом яйцеклетки вводят в матки взрослых овец. Рождающиеся ягнята оказались идентичными овце-донору. Как было отмечено в § 35, такое получение трансгенных животных представляет собой прямой путь клонирования животных с хозяйственно-полезными признаками, включая особей определенного пола.
Трансгенные животные получены также при использовании исходного материала, принадлежащего разным видам, в частности, известен способ передачи гена, контролирующего гормон роста, от крыс в яйцеклетки мышей, а также способ комбинирования блас-томеров овцы с бластомерами козы, что привело к получению гибридных животных (ковец). Эти эксперименты указывают на возможность преодоления видовой несовместимости на самых ранних этапах развития. Особенно заманчивые перспективы открываются (если видовая несовместимость будет преодолена полностью) на пути оплодотворения яйцеклеток одного вида ядрами соматических клеток другого вида. Речь идет о реальной перспективе получения хозяйственно-ценных гибридов животных, которых невозможно получить путем скрещиваний.
Следует отметить, что ядерно-трансплантационные работы еще не очень эффективны. Эксперименты, выполненные на земноводных и млекопитающих, в целом показали, что их результативность является небольшой, причем она зависит от несовместимости между донорскими ядрами и реципиентными овоцитами. Кроме того, препятствием на пути к успехам являются также образующиеся хромосомные аберрации в трансплантированных ядрах в ходе дальнейшего развития, которые сопровождаются гибелью трансгенных животных.
На стыке работ по изучению гибридизации клеток и иммуноло-гических исследований возникла проблематика, связанная с получением и изучением так называемых моноклональных антител. Как отмечено выше (см. § 96), антитела, продуцируемые организмом в ответ на введение антигена (бактерии, вирусы, эритроциты и т. д.), представляют собой белки, называемые иммуноглобулина-ми и составляющие фундаментальную часть защитной системы организма против возбудителей болезней. Но любое чужеродное тело, вводимое в организм, представляет собой смесь разных антигенов, которые будут возбуждать продукцию разных антител. Например, эритроциты человека обладают антигенами не только для групп крови А (II) и В (III), но и многими другими антигенами, включая резус-фактор. Далее, белки клеточной стенки бактерий или капси-да вирусов также могут действовать в качестве разных антигенов, вызывающих образование разных антител. В то же время лимфо-идные клетки иммунной системы организма обычно представлены клонами. Значит, даже только по этой причине в сыворотке крови иммунизированных животных антитела всегда представляют собой смесь, состоящую из антител, продуцируемых клетками разных клонов. Между тем для практических потребностей необходимы антитела только одного типа, т.е. необходимы так называемые моноспецифические сыворотки, содержащие антитела только одного типа, или, как их называют, моноклональные антитела.
В поисках методов получения моноклональных антител швейцарскими исследователями в 1975 г. был открыт способ получения гибридов между лимфоцитами мышей, иммунизированных тем или иным антигеном, и культивируемыми опухолевыми клетками костного мозга. Такие гибриды получили название «гибридомы». От «лим-фоцитарной» части, представленной лимфоцитом одного клона, одиночная гибридома наследует способность вызывать образование необходимых антител, причем одного типа, а благодаря «опухолевой (миэломной)» части она становится способной, как и все опухолевые клетки, бесконечно долго размножаться на искусственных питательных средах, давая многочисленную популяцию гибридом. На рис. 229 показана схема выделения клеточных линий, синтезирующих моноклональные антитела. Линии мышиных клеток, синтезирующих моноклональные антитела, выделяют путем слияния миеломных клеток с лимфоцитами из селезенки мыши, иммунизированной за пять дней до этого желаемым антигеном. Слияние клеток достигают смешиванием их в присутствии полиэтиленгликоля, который индуцирует слияние клеточных мембран, а затем в высеве их на питательную среду, позволяющую рост и размножение только гибридных клеток (гибридом). Размножение гибридомы разводят в жидкой среде, где они растут далее и секретируют антитела в культуральную жидкость, причем только одного типа, к тому же в неограниченных количествах. Эти антитела получили название моноклональных.
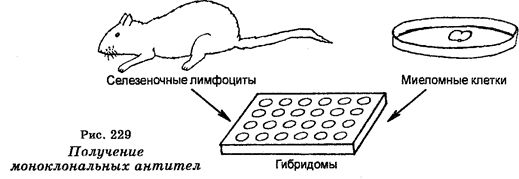
Чтобы повысить частоту образования антител, прибегают к клонированию гибридом, т. е. к селекции отдельных колоний гибридом, способных вызывать образование наибольшего количества антител желаемого типа. Моноклональные антитела нашли широкое применение в медицине для диагностики и лечения ряда болезней (см. § 115). В то же время важнейшее преимущество моноклональной технологии заключается в том, что с ее помощью могут быть получены антитела против материалов, которые невозможно очистить. Напротив, можно получить моноклональные антитела против клеточных (плазматических) мембран нейронов животных. Для этого мышей иммунизируют выделенными мембранами нейронов, после чего их селезеночные лимфоциты объединяют с миеломными клетками, а дальше поступают, как описано выше.
Клеточная инженерия у растений заключается в получении растений из одной клетки, а также в генетических манипуляциях с изолированными клетками, направленными на преобразование их генотипов.
Метод получения растений из одной клетки основан на способности тканей растений ряда видов к неорганическому росту на специальных искусственных средах, содержащих питательные вещества и регуляторы роста. При культивировании тканей растений на таких средах многие клетки оказываются способными к неограниченному размножению, образуя слои (массу) недифференцированных клеток, получивших название каллуса. Если затем каллус разделить на отдельные клетки и продолжить культивирование изолированных клеток на питательных средах, то из отдельных (одиночных) клеток могут развиться настоящие растения. Способность одиночных соматических клеток растений развиваться в настоящее (целое) растение, называют тотипотентностыо
. Возможно, тотипотентность присуща клеткам всех листостебельных растений. Но пока она обнаружена у растений ограниченного круга. В частности, эта способность обнаружена у клеток картофеля, моркови, табака и ряда других видов сельскохозяйственных культур. Этот метод клеточной инженерии растений уже вошел в широкую практику. Однако растения, развившиеся из одной клетки, характеризуются генетической нестабильностью, что связано с мутациями их хромосом. Поскольку генетическая нестабильность дает разнообразные формы растений, они очень полезны в качестве исходного материала для селекции.
Однако растения можно получить и из так называемых протопластов растительных клеток, под которыми понимают клетки, у которых искусственно с помощью гидролитических ферментов (пек-тиназы и целлюлазы) удалена клеточная стенка. Обычно протопласты получают из клеток листьев, корней, лепестков, прорастающей пыльцы, плодов и других структур растений. Способность протопластов давать начало растениям выявлена у очень большого количества видов.
Получение растений из одной клетки или протопласта часто называют клональным микроразмножением. Главнейшее преимущество этого метода заключается в том, что он позволяет резко сократить сроки размножения многих видов растений, а также очень быстро воспроизвести одно и то же растение в сотнях тысяч экземпляров, что имеет исключительно важное значение в селекционной работе и в получении посадочного материала, незараженного возбудителями болезней (рис. 230).
Генетические манипуляции, связанные с растительными клетками, направлены на преобразование генотипов клеток растений, что достигают либо путем соматической гибридизации (получения гибридных клеток) либо путем переноса в клетки генетического материала, происходящего от других организмов. Во всех случаях исходным материалом являются протопласты клеток.
Соматическую гибридизацию осуществляют в несколько этапов, а именно:
1. Получение и слияние протопластов, происходящих от клеток растений разных видов.
2. Культивирование гибридных протопластов, используя селективные питательные среды.
3. Регенерация растений из соматических гибридов (гибридов протопластов) через образование последними каллуса.
Как видно, соматическая гибридизация осуществляется по схеме растения — протопласты — каллус — растения. Ее практическая ценность заключается в том, что соматические гибриды используют в селекционной работе.
Перенос генетического материала от одних клеток к другим осуществляют путем трансформации протопластов чужеродной ДНК либо введением в протопласты чужеродной ДНК с помощью плаз-мид. Из образующегося затем каллуса выращивают растения, содержащие интересующий ген. Растения, полученные таким путем, называют трансгенными растениями (рис. 230).
1. Что понимают под клеточной инженерией и каково ее отличие от генной инженерии?
2. Каковы биологические основы выращивания растений из одной клетки?
3. Назовите методы клеточной инженерии в применении к растениям. Какова хозяйственная ценность растений, получаемых этими методами?
4. Назовите методы клеточной инженерии в применении к животным. Какова хозяйственная ценность животных, получаемых этими методами?
5. Дайте определение понятиям «трансгенные растения» и «трансгенные животные». Сохраняют ли трансгенные организмы свою видовую принадлежность или их можно считать организмами новых видов?
6. Что такое гибридомы и моноклональные антитела? Как их получают?
7. Применима ли клеточная инженерия к человеку?
8. Допустим, что инъекция чужеродной ДНК в яйцеклетку мыши и имплантация оплодотворенной таким путем яйцеклетки в организм мыши закончились ее беременностью и рождением мышат, содержащих в гено-ме копии инъецированной ДНК. Однако мышата оказались мозаиками, т. е. одни их клетки содержат копии инъецированной ДНК, другие лишены этой ДНК. Можете ли вы объяснить природу этого явления?
Альберте В., Брей Д., Льюис Дж„ Рэфф М., Роберте К., Уотсон Дж. Молекулярная биология клетки. М.: Мир. 1994. Т. 1. 615 стр.
Пехов А. П., Щипков В. П., Кривская К. С. Новейшие биотехнологические процессы и система современного высшего образования. Вестник РУДН, серия «Фундаментальное естественнонаучное образование», 1999, стр. 120-128.
Генетическая инженерия не только повысила эффективность старых отраслей биотехнологии, но и открыла новые направления в различных областях этой практической деятельности человека. Рассмотрим некоторые из этих направлений, начав с такого важнейшего направления для жизни людей, как производство пищи, тем более что именно в связи с острой необходимостью решения продовольственной проблемы в мире идет бурное развитие биологической технологии.
Рост народонаселения мира обостряет проблему обеспечения людей пищей. Как показывают расчеты, чтобы обеспечить хотя бы минимальные пищевые потребности населения мира, в ближайшие 20—25 лет необходимо удвоить количество продовольствия, резко увеличить производство пищевого белка, доведя его количество хотя бы до 40-50 млн тонн в год к началу XXI в. Например, по данным Всемирного продовольственного форума ООН, состоявшегося в ноябре 1996 г., производство только риса необходимо увеличить к 2030 г. на 70%.
Из биологических наук прямое отношение к производству пищи имели и имеют микробиология и биохимия, которые непосредственно вовлечены в разработку биотехнологических процессов, основанных на использовании биосинтезирующей способности микроорганизмов.
Уже давно в промышленных условиях осуществляется микробиологический синтез лимонной, щавелевой, итаконовой, глюко-новой и других органических кислот, которые используются в пищевой промышленности.
Как известно, белки растительного происхождения содержат отдельные аминокислоты в очень малых количествах. Это создает значительный дефицит качества кормов, потребляемых сельскохозяйственными животными и птицами. Поэтому еще в начале 60-х гг. в ряде стран был введен промышленный микробиологический синтез аминокислот, включая незаменимые. Сейчас микробиологическим путем получают лизин, аланин, аспараги-новую и глутаминовую кислоты, метионин, триптофан, лейцин и другие аминокислоты, которые широко используют для обогащения кормов.
Значительный удельный вес в микробиологической промышленности занимает производство ферментов. Промышленным путем производят амилолитические, протеолитические, пектиназы и другие ферменты, которые широко используют в пищевой промышленности с целью улучшения вкуса и аромата пищевых продуктов (изготовление сыров, различных кисломолочных продуктов), а также для ферментации соевых бобов и другого сырья.
Определенное значение имеет дешевое производство богатого незаменимыми аминокислотами кормового «одноклеточного» белка (кормовых дрожжей). Подсчитано, что тонны дрожжей, добавленной в корм кур, достаточно для получения дополнительно почти 35 тыс. яиц и 1,5 т куриного мяса. В больших количествах получают также биомассу хлореллы, пасты из которой идут в корм животных, птицы, шелковичного червя. Хлорелла используется также в качестве удобрения.
Микробиологическая промышленность производит в больших количествах различные витамины, которые добавляют в пищевые продукты, а в сочетании с белками добавляют и в корма (белково-витаминный комплекс) для повышения продуктивности животных. Наконец, отметим микробиологическое производство кормовых антибиотиков, используемых для добавки в корм скоту, а также гиб-берелинов и энтомопатогенных препаратов, применяемых в растениеводстве для регуляции роста растений и для защиты их от вредителей.
Сейчас все большее значение приобретают поиски культур микроорганизмов, которые позволили бы придать определенный (желаемый) вкус пищевым продуктам. По грубым подсчетам в мире имеется крупных домашних животных столько же, сколько и людей, т. е. на одного человека приходится одно крупное домашнее животное, а также одна домашняя птица (куры, гуси и Др.), что снижает белковые ресурсы биосферы. Поэтому возникла идея получения пищи из микроорганизмов, минуя животноводство и растениеводство. Специалисты считают вполне возможным получение различных продуктов из микроорганизмов, из которых при сочетании с традиционными видами пищи можно приготовить блюда, сбалансированные по количеству и аминокислотному составу белка и по другим соединениям.
Однако традиционных мер недостаточно в повышении количества и качества пищи. Именно по этой причине производство пищи стало важнейшим направлением генетической инженерии. Задачей этого направления является повышение на принципиально новой основе урожайности сельскохозяйственных растений и, в первую очередь, злаковых культур, как источника хлеба, а также повышение продуктивности сельскохозяйственных животных как источника мяса и мясопродуктов.
В течение длительного периода, начиная с доисторических времен, селекционеры отбирали и размножали растения со свойствами, имеющими агрономический интерес. Этот путь известен в качестве классической селекции. Однако и древние и современные селекционеры были всегда ограничены в своих успехах, ибо это ограничение определялось ограничениями используемых методов отбора.
Генетическая инженерия распространила свои претензии на создание условий для управления такими процессами, как фиксация азота атмосферы, фотосинтез, цветение растений, водный режим, минеральное питание, транспорт веществ и др., которые, как предполагают, имеют важное значение в формировании агрономически ценных признаков растений.
Говоря о фиксации азота, отметим, что ежегодно во всех странах для повышения урожайности полей в почву вносят около 6 млрд тонн удобрений. Высокоурожайные сорта пшеницы, риса и других культур, внедрение которых в практику в ряде стран вызвало в свое время так называемую «зеленую революцию», нуждаются в огромных количествах азотных удобрений. Однако их производство очень обременено для экономики. Недостатком их является и то, что они усваиваются растениями лишь на 40-50%. Значительная часть их смывается в водоемы дождями и вешними водами, что загрязняет питьевую воду и создает условия для развития в водоемах водорослей, поглощающих кислород. Между тем, атмосфера состоит на 70% из азота. Но, как известно, главный источник зерна, злаковые, не способны усваивать азот атмосферы. Поэтому возникла идея переноса генов фиксации азота от клубеньковых бактерий, являющихся симбионтами бобовых и способных фиксировать азот атмосферы, к почвенным микроорганизмам, способным обитать в ризосфере злаковых. Результаты уже выполненных генно-инженерных опытов показывают перспективность этой задачи и реальность ее решения в производственном масштабе.
Увеличение продуктивности растений посредством интенсификации фотосинтеза — это одна из старых идей сельскохозяйственной биологии. Однако на пути ее реализации даже генно-инженерным путем имеется много трудностей. Одна из них заключается в том, что мы ничего не знаем о механизмах, которые ограничивают фотосинтез. Именно это пока препятствует использованию генетических манипуляций применительно к фотосинтезу.
Важной задачей генной инженерии являются поиски путей обеспечения растений генами, контролирующими сроки цветения. Такие гены должны контролировать цветение растений лишь в определенное время года, что позволит уменьшить зависимость земледелия от климатических и метеорологических факторов. Этот вопрос решают введением генов от быстрозацветающих растений в клетки растений, зацветающих в более поздние сроки. Например, установлено, что введение генов, контролирующих цветение растений Arabidopsis, в клетки осины сопровождалось развитием трансгенных осин, зацветающих в 7-месячном возрасте. Осины обычно зацветают лишь в 8-летнем возрасте. Известно, что некоторые растения содержат гены резистентности (R-гены), которые обеспечивают их устойчивость к вирусам, бактериям или грибам, являющимся возбудителями болезней растений. Эти гены кодируют в растениях рецепторные белки. Связываясь с продуктами генов, обеспечивающими патогенность микроор1"анизмов, R-контролиру-емые рецепторные белки включают в действие факторы защиты растений. Однако естественной защиты недостаточно во всех случаях для предупреждения болезней культивируемых растений. Поэтому не меньший интерес представляют также генно-инженер-ные разработки с целью создания растений с повышенной устойчивостью к заболеваниям. Как известно, ресурсы пахотных земель в мире давно исчерпаны, а сейчас даже уменьшаются в связи с деятельностью человека. Между тем одну треть земной поверхности составляют пустыни, полупустыни и сухие саванны. Оскудение ресурсов и рост народонаселения заставляет искать новые сорта и виды сельскохозяйственных растений, которые можно было бы культивировать в пустыне, орошаемой морской водой (неопресненной). Поэтому уже давно ведутся работы, направленные на создание растений — галофитов, способных использовать морскую воду, или растений с меньшей потребностью в воде, когда их культивирование не связано с орошением.
На генетическую инженерию возлагают надежды в плане разработки способов обеспечения растений генами, которые контролируют их устойчивость к заморозкам, засухе и другим неблагоприятным факторам в сельском хозяйстве разных географических зон.
Одно из направлений генетической инженерии в области производства пищи связано с разработкой методов введения генов от животных в геном растений. Как предполагают, это позволит добиться качественного изменения хлеба, сходство его с продуктами животного происхождения. То, что уже удалось ввести гены ин-терферона в табачные и другие растения, делают это направление перспективным.
Как отмечено выше, генетические манипуляции с изолированными клетками растений (клональное микроразмножение растений) позволяют резко сократить сроки размножения многих видов растений, а также вопроизвести одно и то же растение в сотнях тысяч экземпляров, причем незараженных возбудителями болезней, что исключительно важно для сельскохозяйственной практики. Получение сельскохозяйственных растений из одной клетки и дальнейшее клонирование их обещает выведение сортов растений, устойчивых к вирусам, бактериям и грибам, способным вызывать болезни.
Известно, что в сельскохозяйственной практике давно и с успехом используют для повышения урожайности гибридизацию растений и, в частности, отдаленную гибридизацию, под которой понимают скрещивание растений, принадлежащих к разным видам и родам. В разработку теории отдаленной гибридизации и в получение практических результатов в этой области существенный вклад внесли русские ученые Н. И. Вавилов, Г. Д. Карпе-ченко, А. И. Державин, Н. В. Цицин и другие. Однако отдаленная гибридизация растений связана с затратой многих лет работы, ценность генетической инженерии здесь заключается в том, что она открыла перспективы для значительного сокращения сроков получения отдаленных гибридов, а также почти неограниченного использования видового И родового разнообразия растений в качестве исходных форм. Получение отдаленных гибридов стало возможным на основе слияния протопластов отдельных растительных клеток либо на основе трансформации клеток с помощью чужеродной ДНК или с помощью плазмид.
По существу, речь идет о чрезвычайно быстром преодолении генетической несовместимости растений, т. е. о перспективах гибридизации растений отдельных видов или родов с помощью современных методов. Реализация этих перспектив уже привела к некоторым результатам, в частности, к получению новых форм в виде так называемых соккура (соя + кукуруза), сотаба (соя + табак) и табапета (табак + петуния). Осуществлены также эксперименты по получению картомидора (картофель + помидор). Растения, развившиеся из отдельных гибридных клеток, еще не имеют той хозяйственной ценности, которой от них ожидают. Поэтому предстоит еще многое сделать для доработки как самих методов получения отдаленных гибридов, так и сохранения хозяйственно-полезных признаков этих гибридов. Но то, что в ряде стран уже культивируют на значительных площадях генно-инженерные сорта кукурузы, сои и томатов, порождает надежду, что в ближайшие годы в сельскохозяйственное производство войдут принципиально новые методы получения растений с заданными свойствами, включая такие растения, которые не существовали в природе раньше.
Поиски новых способов повышения продуктивности животноводства на основе методов генетической инженерии проходят в трех направлениях, а именно: «конструирование» животных с заданными свойствами путем пересадки генов, клонирование животных путем клонирования клеток после разделения эмбрионов и трансплантаций эмбрионов.
Возможность «конструирования» животных с заданными свойствами путем пересадки генов показана в экспериментах, в которых удалось пересадить крысиный ген гормона роста в оплодотворенные яйцеклетки мышей, которые затем были имплантированы в матки мышей — приемных матерей. Некоторые из появившихся на свет мышат продуцировали чужой гормон роста в очень больших количествах и по этой причине значительно переросли своих собратьев, став мышами-гигантами. Исследования в этом направлении углубляются с каждым годом, принося новые результаты. С помощью пересадки генов предполагают вывести коров, вес которых будет в несколько раз большим по сравнению с весом коров существующих пород, а удои будут достигать до 20 000 кг молока в год.
Большие возможности таит в себе метод клонирования животных, который уже применяют в случае крупного и мелкого рогатого скота. Обычно измельчают эмбрионы, состоящие из 60—80 клеток, которые имплантируют в организм приемных матерей. Следовательно, в принципе можно получать из одного эмбриона несколько десятков животных.
Трансплантации эмбрионов — это, вероятно, наиболее развитое направление в рассматриваемой области, успехи которого связаны, как уже отмечено выше, с разработкой способов индукции полиовуляции, искусственного оплодотворения клеток и имплантации эмбрионов в организм животных. С помощью этого метода уже произведены сотни тысяч телят.
Новые методы обладают чрезвычайно важными преимуществами, обеспечивающими несомненный прогресс в этой области. Они позволяют прежде всего ускорить разведение животных с высокими хозяйственными качествами. Кроме того, они позволят сохранить ценный генофонд, т. к. «вымытые» эмбрионы можно консервировать замораживанием и хранить неопределенно долгий срок. Этот способ удешевляет транспортировку животных, ибо рентабельнее транспортировать замороженные эмбрионы в пробирке, нежели самих животных.
Это направление генетической инженерии связано с решением другой глобальной проблемы, которая касается производства энергии и материалов. В связи с неизбежным истощением мировых запасов природных энергоносителей исключительное значение приобрело создание индустрии, связанной с использованием растительного сырья для получения моторного топлива. В обозримом будущем, вероятно, будут найдены заменители нефтепродуктов. Предполагается, что бензин будет полностью заменен этиловым спиртом. Поэтому идут поиски микроорганизмов, пригодных для создания более эффективной технологии получения дешевого этилового спирта из растительного сырья на основе ферментации. Заметим, что дешевый этиловый спирт необходим также для производства растворителей, красок, смазочных и клеящих материалов, детергентов, смол для синтетических волокон, лекарственных веществ и т. д.
Известны попытки создать технологию получения этилового спирта из бумажной макулатуры с помощью генно-инженерных штаммов микроскопических грибов.
Генетическая инженерия вовлечена в разработку способов получения искусственной сырой нефти, а также способов получения «биогаза» из сырья различного происхождения.
С генетической инженерией связывают прогресс в области создания технологии новых материалов. Уже найдены микроорганизмы, которые размножаются в аэрируемой среде, представляющей собой смесь метилового спирта, аммония и других неорганических соединений.
Некоторые из утилизирующих метиловый спирт микроорганизмов способны синтезировать полимер, напоминающий синтетический полиэфир, но отличающийся от него хрупкостью. Сейчас проводятся исследования с целью «уподобить» структуру этого полимера структуре полиэфира и использовать затем этот полимер для изготовления из него тканей с особыми свойствами. Речь идет о реальной перспективе разработки научных и технологических основ текстильной промышленности нового типа, в которой производителями тканей станут микроорганизмы, сконструированные генно-инженерными методами.
В настоящее время ищут методы ферментативного синтеза окиси пропилена, необходимого в производстве пластических масс. Это направление имеет ряд преимуществ по сравнению с химическим синтезом. Уже найдены микроорганизмы, синтезирующие ферменты для этих целей.
Генная инженерия оказалась очень перспективной для медицины, прежде всего, в создании новых технологий получения физиологически активных белков, используемых в качестве лекарств (инсулин, соматостатин, интерфероны, соматотропин и другие). Инсулин используют для лечения больных диабетом, который стоит на третьем месте (после болезней сердца и рака) по частоте вызываемых смертельных случаев. Мировая потребность инсулина составляет несколько десятков килограммов. Традиционно его получают из панкреатических желез свиней и коров, но гормоны этих животных слегка отличаются от инсулина человека. Инсулин свиней различается по одной аминокислоте, а коровий — по трем. Считают, что инсулин животных часто вызывает побочные эффекты. Хотя химический синтез инсулина осуществлен давно, но до сих пор промышленное производство гормона оставалось очень дорогим. Сейчас получают дешевый инсулин с помощью генно-инже-нерного метода путем химико-ферментативного синтеза гена инсулина с последующим введением этого гена в кишечную палочку, которая затем синтезирует гормон. Такой инсулин более «биологичен», т. к. химически идентичен инсулину, вырабатываемому клетками поджелудочной железы человека.
Интерфероны — белки, синтезируемые клетками главным образом в ответ на заражение организма вирусами. Интерфероны характеризуются видовой специфичностью. Например, у человека установлены три группы интерферонов, продуцируемых различными клетками под контролем соответствующих генов. Интерес к интерферонам определяется тем, что их широко используют в клинической практике для лечения многих болезней человека, особенно вирусных.
Имея крупные размеры, молекулы интерферона малодоступны для синтеза. Поэтому большинство интерферонов сейчас получают из крови человека, но выход их при таком способе получения небольшой. Между тем потребности в интерфероне исключительно велики. Это поставило задачу изыскать эффективный метод производства интерферона в промышленных количествах. Генетическая инженерия лежит в основе современного производства «бактериального» интерферона.
Усилилось влияние генетической инженерии на технологию тех лекарственных веществ, которые уже давно создаются по биологической технологии. Еще в 40-50-е годы была создана биологическая промышленность для производства антибиотиков, которые составляют наиболее эффективную часть лекарственного арсенала современной медицины. Однако в последние годы отмечается значительный рост лекарственной устойчивости бактерий, особенно к антибиотикам. Причина заключается в широком распространении в микробном мире плазмид, детерминирующих лекарственную устойчивость бактерий, именно поэтому многие знаменитые ранее антибиотики утратили свою былую эффективность Единственный пока путь преодоления резистентности бактерий к антибиотикам — это поиски новых антибиотиков. По оценкам специалистов в мире ежегодно создают около 300 новых антибиотиков. Однако большинство из них либо неэффективно, либо токсично. В практику же каждый год вводится лишь несколько антибиотиков, что заставляет не только сохранять, но и увеличивать мощности антибиотической промышленности на основе генно-инженерных разработок.
Для лечения паразитарных и многих неинфекционных болезней в течение длительного времени используют лекарственные вещества, полученные в процессе химического синтеза, однако некоторые из них вредны для организма. Поэтому наметилась тенденция расширить производство микробных метаболитов (других, кроме антибиотиков), перспективных в качестве лекарственных веществ. Еще в начале XX в. В. Л. Омелянский разработал теоретические основы учения о трансформирующей активности микроорганизмов. Вызывая окисление, восстановление, гидратацию, дегидратацию, полимеризацию и другие реакции, микроорганизмы многих видов способны изменять субстрат, на который они воздействуют, трансформировать химические соединения. В настоящее время на трансформирующей способности микроорганизмов основан ряд производств, выпускающих из стероидного сырья растительного происхождения вещества с фармакологическими свойствами. Например, с помощью микроскопических грибов трансформируют стероидные соединения в кортизон и гидрокортизон. Актиномицеты и бактерии используются для производства преднизолона, диакабола и т. д., имеющих большое значение в медицине.
Основные задачи генной инженерии в тех технологиях лекарственных веществ, в которых продуцентами лекарств являются микроорганизмы, определяются необходимостью генно-инженерной реконструкции последних с целью повышения их активности. В то же время началась реализация идеи создания лекарств в виде малых молекул, что способствует их большей эффективности.
Иммунная биотехнология связана с производством, прежде всего вакцин нового поколения для профилактики инфекционных болезней человека и животных.
Первыми коммерческими продуктами, созданными с помо1цью генетической инженерии, стали вакцины против гепатита людей', ящура животных и некоторые другие. Исключительно важное направление в этой области связано с производством моноклональных антител, реагентов, необходимых для диагностики возбудителей болезни, а также для очистки гормонов, витаминов, белков различной природы (ферментов, токсинов и других).
Значительный практический интерес представляет метод получения искусственного гемоглобина путем введения гемоглобино-вых генов в растения табака, где под контролем этих генов продуцируются а- и р-цепи глобина, которые объединяются в гемоглобин. Синтезируемый в клетках табачных растений гемоглобин полностью функционален (связывает кислород). Клеточная инженерия в применении к человеку связана не только с решением фундаментальных проблем биологии человека, но и с преодолением прежде всего женского бесплодия. Поскольку частота положительных случаев имплантации в матку женщин эмбрионов, полученных in vitro, является небольшой, то получение монозиготных близнецов-эмбрионов in vitro также имеет значение, поскольку увеличиваются возможности повторных имплантаций за счет «запасных» эмбрионов.
В использовании генетической инженерии применительно к проблемам медицины особое значение приобрела задача разработки генно-инженерных методов радикального лечения наследственных болезней, которые, к сожалению, еще не поддаются лечению существующими методами. Содержание этой задачи заключается в разработке способов исправления (нормализации) мутаций, результатом которых являются наследственные болезни, и в обеспечении передачи «исправлений» по наследству. Считают, что успешной разработке генно-инженерных методов лечения наследственных болезней будут способствовать данные о геноме человека, получаемые в результате выполнения международной научной программы «Геном человека».
Подняв на новый уровень биотехнологию, генетическая инженерия нашла также применение в разработке способов определения и устранения загрязнений окружающей среды. В частности, сконструированы штаммы бактерий, которые являются своеобразными индикаторами мутагенной активности химических загрязнителей. С другой стороны, генно-инженерным способом сконструированы штаммы бактерий, которые содержат плазмиды, под контролем которых происходит синтез ферментов, способных разрушать многие химические соединения-загрязнители среды обитания. В частности, некоторые плазмидосодержащие бактерии способны разлагать до безвредных соединений нефть и нефтепродукты, оказавшиеся в среде в результате различных аварий или других неблагоприятных причин.
Однако генетическая инженерия — это превращение генетического материала, которое в природе отсутствует. Следовательно, продукты генной инженерии — это абсолютно новые продукты, не существующие в природе. Поэтому она сама по себе из-за неизвестности ее продуктов таит опасность как для природы и среды обитания, так и для персонала, работающего в лабораториях, где используют методы генетической инженерии или работают со структурами, созданными в ходе генно-инженерных работ.
Поскольку возможности клонирования генов безграничны, то еще в самом начале этих исследований среди ученых возникли вопросы о природе создаваемых организмов. Одновременно были высказаны предположения о ряде нежелательных последствий этой методологии, причем эти предположения нашли поддержку и среди широкой общественности. В частности, появились вопросы о свойствах бактерий, получивших в генно-инженерных экспериментах гены животных. Например, сохраняют ли бактерии Е. coli свою видовую принадлежность из-за содержания введенных в них генов животного происхождения (например, гена инсулина) или их следует считать новым видом? Далее, насколько долговечны такие бактерии, в каких экологических нишах они могут существовать? Но самое главное стало заключаться в появлении опасений, что в ходе производства и манипуляций с рекомбинантными молекулами ДНК могут быть созданы генетические структуры со свойствами, непредвиденными и опасными для здоровья человека, для исторически сложившегося экологического равновесия. Тогда же начались и призывы к мораторию на генетическую инженерию. Эти призывы вызвали международный резонанс и повели к международной конференции, которая состоялась в 1975 г. в США и на которой широко обсуждались возможные последствия исследований в этой области. Затем в странах, в которых стала развиваться генетическая инженерия, были выработаны правила работы с рекомбинантными молекулами ДНК. Эти правила направлены на исключение попадания в среду обитания продуктов деятельности генно-инженерных лабораторий.
Другой аспект нежелательных последствий генно-инженерных работ связан с опасностью для здоровья персонала, работающего в лабораториях, где используют методы генетической инженерии, поскольку в таких лабораториях используют фенол, этидий бромид, УФ-излучение, которые являются вредными для здоровья факторами. Кроме того, в этих лабораториях существует возможность заражения бактериями, содержащими рекомбинантные молекулы ДНК, контролирующие нежелательные свойства, например, лекарственную резистентность бактерий. Эти и другие моменты определяют необходимость повышения уровня техники безопасности в генно-инженерных работах.
1. Каковы научные и социальные предпосылки формирования разных направлений в использовании генетической инженерии?
2. Что вы можете сказать о результативности генетической инженерии в растениеводстве?
3. Можно ли с помощью методов генетической инженерии повысить продуктивность животноводства? Если да, то каким образом?
4. Действительно ли реальны надежды на генную инженерию в создании новых источников энергии и новых материалов?
б. Что Вы знаете о роли генетической инженерии в создании лекарств и иммунопрепаратов? Приведите примеры успехов в этой области.
6. Каковы возможности генетической инженерии в охране среды обитания?
7. Существуют ли опасности в работе персонала генно-инженерных лабораторий? Если да, то назовите их.
Бирс Р.. Басит Э. (ред). Рекомбинантные молекулы: значение для науки и практики (пер. с англ.). М.: Мир. 1960. 624 стр.
Holland H.,Petersen U. Living Dangerously. Prinston University Press. 1995. 210 pp.
Hunter-Cevera J.. Belt A. Maintaining Cultures for Biotechnology and Industry. Academic Press. 1995. 242 pp.
Russo E., Cove D. Genetic Engineering. WH Freeman. 1995. 255 pp.
На протяжении многих веков у всех, кто интересовался проблемами жизни, всегда были различия во взглядах на живой мир, в оценках свойств, характерных для живых существ, в обсуждении бытия человека, причем характер этих взглядов зависел от исторической эпохи, в которой они формировались.
Древнегреческие философы, будучи стихийными материалистами, рассматривали мир в качестве единого материального целого, которое находится в вечном возникновении и исчезновении, в непрерывном движении и в изменениях. Эти взгляды в истории философии получили название эллинского мировоззрения.
Особенно большое внимание в эллинском мировоззрении уделялось человеческой природе. Оценивая очень высоко человека вообще, древнегреческая философия придавала большое значение его телу и образу жизни. Греки отстаивали гармоничное развитие всех природных способностей, заложенных в человеке с целью формирования его нравственного поведения. Однако с упадком античной цивилизации оптимистический взгляд на природу человека уступил место новому взгляду, в соответствии с которым тело человека есть оболочка его души, причем тело ничтожно, но душа чиста.
Пренебрежение к телу и возвеличивание души стало одним из важнейших моментов христианства. В IV—V вв. н. э. высшим идеалом стремлений человека в христианстве стало подавление чувственной стороны природы человека. С этого времени латинская церковь ввела даже безбрачие, которое достигло высшего уровня к концу Х — началу XI в. В более поздние времена положение вновь изменилось. Эпоха реформации способствовала возрождению эллинских взглядов на природу и бытие человека. Стала провозглашаться необходимость развития всех естественных свойств человека. Природа человека для французских материалистов оказалась основой их учения о нравственности, причем одинаковой независимо от типа религиозных убеждений людей.
В XIX в. роль философии стала еще большей. Ч. Дарвин анало-гизировал нравственность с «общим благом», под которым он понимал развитие возможно большего числа здоровых и сильных особей, со всеми способностями, в наиболее совершенной степени развития. Однако из-за нерешенности проблема природы человека перешла и в наше время, превратившись в проблему взаимоотношений социальных и биологических факторов в развитии человека (см. ниже).
В древнейшие времена зародилось также учение, названное витализмом. Виталисты считали, что в основе жизни лежит особое сверхъестественное непознаваемое начало («жизненная сила», «органическая сила», «архей» и Др.), которое возникает раньше организма, но направляет его. Мысль о создании живого творцом перешла в средневековье (V-XV вв.). В то же время на основе идеализма возникает креационизм (от лат. creator — творец) — учение о создании живого путем акта творения и о неизменяемости всего живого. Исходя из того, что изначальность всего живого создана творцом, креационисты объясняли многообразие живых существ в соответствии с Библией и положениями теологии (от греч. teos — бог, logos — наука), тогда как органическую целесообразность они объясняли теологически (от греч. teleos — стремящийся к цели) — «мудрой предусмотрительностью», «высшей мудростью» творца. Теологические и телеологические взгляды в средневековье совпали с виталистическими представлениями о природе. Эти представления укрепились в период средневековья по той причине, что в то время сложность организации живых существ казалась не поддающейся объяснению. Однако, несмотря на господство креационизма, в средние века все же шла жестокая борьба между рациональными представлениями и теолого-мисти-ческими взглядами на природу, между проявлениями интересов к природе как реально существующей и символико-мистическим восприятием мира.
В XVII-XVIII вв. формируются материалистические взгляды на живую природу среди французских философов. Доказывая материальность души, зависимость психики от тела, т. е. отстаивая первичность материи и вторичность сознания, французские философы защищали материализм. В частности, Р. Декарт (1569-1650) ввел понятие о «животном-машине», считая, что животные являются механизмами, возникающими по законам механики, а Ж. Ламет-ри (1709-1751) ввел и обосновал понятие о «человеке-машине». Однако, понимая организмы в качестве суммы частей, действующих в соответствии с законами физики и химии, французские материалисты все же не замечали специфики живого. Несомненно, что их взгляды означали существенное продвижение вперед, но их материализм все же был механистическим.
Чрезвычайно быстрое накопление научных данных о живых организмах в XVII-XVIII вв. вело к дифференциации наук, которая была очень полезной в изучении природы вообще и сопровождалась выяснением закономерностей, которые были очень полезны для развития производительных сил. Но дифференциация наук имела тогда и негативные последствия, ибо вела к безвозвратному расчленению природы в сознании естествоиспытателей и философов. В рамках представлений, обусловленных дифференциацией наук, им казалось, что природа состоит из отдельных предметов и процессов, не связанных между собой, что между природными явлениями нет ни единства, ни взаимосвязи. В результате таких оценок возникали мысли о неизменяемости природы, об отсутствии в ней развития и, самое главное, о том, что природа всегда была такой, какой она есть сейчас. Так формировались метафизические представления. Сейчас можно отметить, что, возможно, это был самый трудный этап в развитии знаний о живой природе. Положение осложнилось еще и тем, что Ф. Бэкон (1561-1629) и Д. Локк (1632-1704) перенесли метафизический способ познания природы из естествознания в философию, а это означало укрепление метафизического способа мышления, который, как отмечал еще И. И. Мечников, ничего не имеет общего с наукой.
На основе креационизма в XVII-XVIII вв. возник и значительно распространился преформизм, сторонники которого полагали, что в яйцеклетке (или сперматозоиде) уже содержится преформи-рованный (полностью сформированный) организм и что в дальнейшем все сводится лишь к его росту, увеличению размеров. Будучи метафизическим от начала до конца, преформизм сразу же сомкнулся с распространенными тогда общими метафизическими представлениями о постоянстве (неизменяемости) видов, о неизменяемости органического мира. В практическом плане все это значительно тормозило развитие биологии.
В XVII-XVIII вв. широко распространилось учение и об эпигенезе, сторонники которого полагали, что организм возникает из гомогенной бесструктурной массы. Для своего времени учение об эпигенезе было прогрессивным, ибо близко подходило к материалистическому пониманию развития, что в значительной мере способствовало возникновению эволюционизма в биологии. Однако оно было односторонним, т. к. в нем не принималось во внимание взаимодействие внутреннего и внешнего в развитии зародышей (организмов).
В противоположность креационизму в XVII в. возник трансформизм (от лат. transformatio — развертываю), в соответствии с которым допускалась изменяемость живого, но вне связи с поступательным характером развития (от низших организмов к высшим). В XVIII в. и в первой половине XIX в. между креационизмом и трансформизмом проходила очень острая борьба, переросшая в борьбу креационизма с эволюционизмом. Поэтому во второй половине XIX в. борьба идей в биологии пошла в основном вокруг идеи развития вообще и дарвиновской теории эволюции, в частности. Благодаря Ч. Дарвину во второй половине XIX в. из биологии был изгнан креационизм или, по крайней мере, ему был нанесен сокрушительный удар, в результате которого была сокрушена господствовавшая веками метафизическая доктрина неизменяемости видов. Это сопровождалось тем, что идея развития проникла во все биологические науки, вызвав новые подходы к пониманию научной картины мира. Дарвинизм оказался мощнейшим естественнонаучным обоснованием диалектического материализма, который стал философским фундаментом биологии.
Однако в XIX в. значительное распространение получили и антидарвиновские концепции эволюции. С дарвинизмом стали конкурировать концепции неоламаркизма (механоламаркизм, орто-ламаркизм, психоламаркизм). Благодаря Э. Геккелю возник социал-дарвинизм, сторонники которого признавали роль естественного отбора в развитии человеческого общества и возникновении классов. Появилась также теологическая концепция эволюции, по которой органический мир является результатом развития, стремящегося к высшей цели и руководимого разумом (духовным началом).
В XX в. на смену витализма пришел неовитализм, основатель которого немецкий биолог Г. Дриш (1876-1941) считал, что строение и жизнедеятельность организма предопределяется и управляется особой жизненной силой, идеей, существующей вне пространства, но действующей в пространстве и названной заимствованным у Аристотеля термином «энтелехия». Г. Дриш утверждал также, что «энтелехия» является ведущей, если не единственной причиной эволюции. Новая вспышка неовитализма произошла в наше время в результате абсолютизации наиболее эффективных методов изучения молекулярного уровня организации живого, недооценки сущности этого уровня и специфики биологических макромолекул. Неовитализм стал проявляться в стремлении рассматривать макромолекулы нуклеиновых кислот и белков в качестве «живых» молекул. Переоценкой элементарных молекулярных структур современные неовиталисты стараются опровергнуть материалистическую сущность мира, т. е. извратить научную картину мира. На виталистической основе возникли представления о «жизненном плане», носителем которого является «номофор», представляющий собой нематериальный фактор организма. На виталистической основе в последние годы вновь стал проявляться и механицизм, заключающийся в сведении живой материи к простым элементам и в признании существования якобы единой унифицированной структуры материи, гомогенной по своим свойствам и не претерпевающей развития. Многообразие и единство мира механицистами превращается в единообразие, а качественные различия заменяются представлениями о простых количественных различиях в сочетаниях одинаковых молекулярных структур. Как видно, речь идет об отрицании идеи развития.
Виталистические и механистические толкования молекулярного уровня организации живой материи ведут к идеализму. Между тем современные естественно-научные данные подтверждают положение о материальном единстве мира. Структура, рост и размножение клеток происходит по тем же законам, которые определяют поведение различных молекул, находящихся в клетке. Но именно рост и размножение являются важнейшими свойствами живого.
Креационизм, как и витализм, тоже оказался очень живучим. В наше время он вновь возродился, причем даже возникли стремления придать креационизму статус официальной науки, сделать его «научным» и опять противопоставить теории эволюции. Современные креационисты полагают, что Земля и Вселенная (вещество, энергия и жизнь) возникли внезапно и относительно недавно (10 тыс. лет назад), причем «из ничего». Они придают большое значение в истории Земли катастрофам, включая всемирный потоп. Признавая недостаточность мутаций и естественного отбора в эволюции, считают, что она компенсируется действием сверхъестественных сил. «Научные» креационисты развивают положение о раздельном происхождении человека и обезьяны, чем пытаются опровергнуть положение о естественном происхождении человека. Недостатки биологического образования населения позволили в наше время довольно широкое распространение креационизма. Например, в США по опросам 1991 г. оказалось, что в креационизм верят около 47% опрошенных. Больше того, бывший президент США Р. Рейган в одной из своих речей высказал сомнения относительно справедливости даже теории эволюции. Между тем новейшие достижения биологических наук дали новые подтверждения справедливости теории эволюции, новые свидетельства в пользу естественного происхождения человека. ' Достижения биологических наук обострили ряд старых проблем и выдвинули новые проблемы социального значения. Если в конце XIX — начале XX в. в числе обсуждавшихся проблем социального звучания в нравственном аспекте называли целесообразность продления жизни человека, возможность или невозможность : экспериментов на животных и человеке, а также необходимость а лечения венерических болезней, то в наше время обострился ин-| терес к проблеме, связанной с ростом народонаселения и вытекающими из этого последствиями. На протяжении последнего тысячелетия и особенно в период новейшей истории население мира неуклонно росло. Например, если в 1961 г. оно составило более 3 млрд людей, то в 1996 г. оно уже достигло почти 6 млрд. Удвоение населения мира предполагается и в ближайшие 35 лет. Каковы последствия этого явления?
Еще до нашей эры выдающийся представитель греческой цивилизации Платон (около 428-348 гг. до н. э.) в своем сочинении «Законы» предлагал регулировать численность популяций (населения), устанавливая его количество соответственно размерам занимаемой территории и наличию достаточного количества жилищ. Платон рассматривал популяции людей в качестве структурных единиц, а граждан государства — в качестве структурных единиц популяции, причем предлагал регулировать численность популяции путем воздействия факторов, повышающих или понижающих размеры популяции. Повышение частоты рождаемости Платон считал «премией», а понижение рождаемости должно было быть результатом устанавливаемого общественным мнением «позора». С точки зрения современной науки рассуждения Платона представляют собой чистейшую спекуляцию, ибо они не могут быть подтверждены научным подходом в виде наблюдения или эксперимента. Тем не менее эта спекуляция оказалась полезной в позднейшие времена для формирования взглядов на структуру и свойства популяций людей.
 Аристотель считал полезным принятие закона, ограничивающего рождаемость путем абортов. Следует вспомнить, что аборты не запрещались ни в Древней Греции, ни в Древнем Риме. Аристотель считал полезным принятие закона, ограничивающего рождаемость путем абортов. Следует вспомнить, что аборты не запрещались ни в Древней Греции, ни в Древнем Риме.
В христианском мире к регулированию народонаселения обращались неоднократно. Особенно интерес к регулированию народонаселения обострился, когда в 1798 г. англичанин Томас Мальтус (1766-1834) представил новый подход к пониманию природы популяций людей. Задавшись целью выяснить, всегда ли подвержены люди бедности, голоду, болезням и войнам и может ли человек улучшить свою жизнь, Т. Мальтус сформулировал представления о факторах (силах), которые регулируют размеры популяции людей, причем его представления основывались на признаниях двух важнейших допущений, а именно: человеку для существования необходима пища, с одной стороны, человек будет репродуцировать себя непрерывно, с другой. Руководствуясь этими допущениями, Т. Мальтус предположил, что репродукция популяции человека идет в геометрической прогрессии, тогда как количество пищи производится популяцией в арифметической прогрессии (рис. 231). В результате этого размеры популяции неизбежно оказываются ограниченными количеством пищи, т. е. из-за недостатка пищи размеры популяции не могут увеличиваться далее геометрически. В доказательство своих заключений Т. Мальтус приводил данные о росте населения в английской колонии (США) в Америке и в самой Великобритании. В английской колонии (США) население в период с 1643 г. по 1760 г. выросло с 21 200 человек до 500 тыс, причем размеры популяции удваивались каждые 25 лет, т. е. росли в геометрической прогрессии. Т. Мальтус подчеркивал, что это была популяция, имевшая достаточное жизненное пространство и достаточное количество пищи. За это же время популяция людей (население) на Британских островах увеличилась лишь незначительно, что определилось недостатком пищи и эмиграцией. Кроме того, проследив судьбу свыше 250 известнейших английских семей на протяжении длительного времени и увидев снижение их численности, Т. Мальтус заключил, что размеры популяции зависят от количества пищи, а рост популяции контролируется голодом и болезнями, т. е. идет борьба за существование.
Теория Т. Мальтуса была использована Дарвином для формирования представлений о роли борьбы за существование в естественном отборе. Тем не менее она все время вызывала возражения, т. к. не могла решить вопрос о последствиях роста населения.
В начале нашего века И. И. Мечников также пришел к заключению, что превышение потребностей организмов над средствами к их удовлетворению составляет основной разлад в. природе, который все же может гармонизироваться либо приспособлением организмов к новым условиям, либо уменьшением их плодовитости. Однако в случае человека эта гармонизация может достигаться сознательно либо путем увеличения средств к существованию, либо ограничением рождаемости. Как писал И. И. Мечников в 1907 г. «...по законам природы человек в состоянии очень сильно размножаться, однако идеал его благоденствия требует ограничения плодовитости».
В настоящее время достигнута ясность, что рост мирового народонаселения в современных масштабах, действительно, вызовет трудности в обеспечении людей продовольствием и доброкачественной водой. Важнейшая причина здесь заключается в том, что жизненное пространство (размеры Земли) никогда не увеличится, а рост производительности сельского хозяйства неизбежно будет остановлен на определенном уровне, поскольку конечная урожайность сортов растений и продуктивность пород животных, которая может быть достигнута в обозримом будущем, неизбежно будет ограничена их нормой реакции. Следовательно, если предположить даже максимально возможный уровень производства продовольствия, то этому уровню могут соответствовать лишь определенные размеры популяций людей. Следовательно, возникнет необходимость регулирования численности населения путем ограничения рождаемости. Между тем решение проблемы регулирования роста народонаселения мира путем ограничения рождаемости встречает серьезные затруднения. Одни считают регулирование нравственным, другие — безнравственным решением. В одних странах ограничение рождаемости невозможно из-за существующих стандартов жизни и образования, тогда как в других, напротив, по религиозным причинам рождаемость стимулируется.
Другой социальной проблемой глобального значения является проблема сохранения экологического равновесия на Земле (сохранения окружающей среды). Сейчас совершенно ясно, что индустриально развитые страны, используя достижения научно-технической революции и безудержно эксплуатируя природные ресурсы, смогли обеспечить высокий уровень потребления. Но при этом нарушение ими экологического равновесия поставило мир перед катастрофой. Стремясь решить свои проблемы, менее развитые страны также увеличивают активность в биосфере, что еще в большей мере повышает экологическую угрозу. Стало ясно, что все действия в мире должны осуществляться в свете понимания этой угрозы и разработки мер по ее недопущению.
В связи с непреодолимо углубляющейся деятельностью человека в биосфере возникла проблема, которая может быть определена как проблема «человек, природа и общество». Речь идет о том, как человек и общество в целом должны строить свои отношения с природой, чтобы не оказаться в какой-то момент лицом к лицу с безнадежно разрушенной природой. В разное время вопросы взаимоотношений человека (общества) с природой решались по-разному, но часто не в пользу природы.
Современные данные биологии вообще и экологии в частности помогают формированию мысли о том, что в наше время основной парадигмой мирового развития должно стать обеспечение экологической безопасности. Однако это может быть обеспечено только в результате тесного международного сотрудничества. Между тем разные страны подходят к этой проблеме по-разному. Поэтому их разрозненность сопровождается малой эффективностью, что не позволяет решать проблему на достаточном уровне. Необходимы новые международные отношения, ибо сохранение окружающей среды — это самая приоритетная задача человечества. Заглядывая в будущее человечества, можно сказать, что оно будет связано с качеством жизни, со здоровьем человека, с подлинным осуществлением мер по сокращению среды обитания.
Возвращаясь к природе человека, которая в наше время обсуждается на уровне определения роли биологических и социальных факторов в развитии человека, нельзя не отметить, что здесь по-прежнему существуют разные подходы к решению проблемы. Отдельные философские концепции человека (природы человека) основаны на преувеличении биологических факторов. В рамках этих концепций человек рассматривается часто в качестве иррационального животного, а социальные условия в качестве фона, способствующего развитию человека. Как и в старом социал-дарвинизме признается, что различия между классовыми группами людей также определяются биологическими факторами, в частности, естественным отбором. Сторонники превалирующего значения биологических факторов в развитии человека считают, что генетическая информация современных людей якобы недостаточна для программирования поколений, способных к полному овладению результатами, порожденными научно-технической революцией сейчас и ожидаемыми в будущем. Больше того, утверждается, что человек как биологический вид, подобно многим видам, вымершим в ходе эволюции, тоже угаснет. Однако чтобы это не произошло, человека необходимо «переделать» на биологической основе, используя достижения генетической инженерии. При этом речь идет не об ортобиозе по И. И. Мечникову, а о глубокой реконструкции генетического аппарата человека, последствия которой кажутся непредсказуемыми. Преувеличивая биологические особенности человека, сторонники биологизаторской идеи объясняют многие общественные пороки, включая преступность, биологическими недостатками людей, «звериными» генами, якобы унаследованными ими от своих далеких предков и от предыдущих поколений.
Напротив, в соответствии с другими взглядами, человек есть часть живого мира, есть существо, действующее в сфере общественной формы движения материи. «Живая природа» и социальная сущность человека взаимно связаны нерасторжимыми узами. Специфические человеческие действия и социальное положение человека не предопределяются только биологическими основами. Жизнь людей определяется не естественным отбором, а условиями жизни. Генетическая программа человека обладает не только устойчивостью, но и громадным потенциалом, обеспечивающим программирование поколений, способных к овладению результатами научно-технической революции, к социальному совершенствованию в интересах большинства. Следовательно, нет необходимости в глубокой генетической реконструкции современного человека. Напротив, важнейшим условием всестороннего развития и совершенствования человека является социальная основа и образ жизни, которые определяются общественным бытием и общественным сознанием.
Критически оценивая обе концепции и признавая выдающуюся роль социальных условий в развитии и поведении людей, в формировании личности, нельзя забывать при этом основополагающую роль биологических факторов, в частности, здоровья людей, их природных способностей. Объективное, адекватное современному уровню знаний заключение состоит в том, что природа человека является биосоциальной. Другими словами, человек есть биосоциальное существо.
В связи о появлением генетической инженерии, а также в связи с успехами молекулярно-генетического изучения генома человека возникли новые проблемы не только социального, но и этического звучания. Попытки разрешения этих проблем означают начало формирования нравственности в ядерный век, т. е. формирования совокупности принципов и норм поведения людей в условиях научных достижений, непосредственно затрагивающих их бытие. Одна из таких социально-этических проблем касается определения генетического статуса людей. Речь идет о введении в практику генетического обследования людей, создания их нуклеотидных карт и о стремлениях использовать молекулярно-генети-ческие карты для определения профессиональной ориентации и занятости людей. Поскольку создание нуклеотидных карт может выявить индивидов с неблагоприятными генотипами, в обществе возникли вопросы о степени гласности результатов генетического обследования и о защите лиц с неблагоприятными генотипами в плане выбора ими профессии или найма на работу. Лица, содержащие в генотипе мутации, не могут нести ответственность за эти мутации и, следовательно, не должны иметь ограничений в выборе профессии.
Следующая этическая проблема, заслуживающая обсуждения, касается этики экспериментов, связанных с оплодотворением в пробирке (in vitro) яйцеклеток и созданием «пробирочных» детей. Обсуждение этой проблемы идет по нескольким другим направлениям.
Во-первых, опираясь на принятый в 1947 г. Нюрнбергский кодекс медицинской этики, в соответствии с которым экспериментам на человеке должны предшествовать адекватные эксперименты на животных, а риск, связанный с экспериментами на человеке, должен быть полностью оправдан, многие считают, что преодоление бесплодия путем оплодотворения яйцеклеток в пробирке есть пример экспериментов на людях, которые осуждаются в течение многих веков. Следовательно, такие эксперименты не должны проводиться.
Во-вторых, по мнению многих специалистов оплодотворение яйцеклеток в пробирке может сопровождаться нарушениями количества или структуры половых хромосом, что будет сопровождаться некоторым повышением генетического груза человечества за счет вхождения в популяции человека «пробирочных» людей. Поскольку в пользу этого предположения уже есть некоторые доказательства, то оно также представляет собой возражение против этих экспериментов.
Далее, в соответствии с доктриной римско-католической церкви душа вселяется в эмбрион во время беременности в определенное, хотя и точно не установленное время.
Следовательно, оплодотворенная яйцеклетка уже представляет собой человеческую жизнь и даже личность. Между тем не все оплодотворенные яйцеклетки сохраняются при имплантации их в организм матерей. По ряду данных оплодотворенные яйцеклетки лишь в 40—50% случаев приживаются в матке матерей. Поэтому возник дополнительный довод против этих экспериментов, по которому неудачные результаты, связанные с прекращением жизни оплодотворенной яйцеклетки, есть грубейшее нарушение прав человека. Наконец, критики метода «пробирочных» детей считают, что этот метод таит в себе угрозу использования его для решения евгенических программ, которые давно не вызывают согласия в обществе. Рассмотрение всей проблематики, связанной с оплодотворением in vitro, показывает, что разработка способов оплодотворения яйцеклеток в пробирке и способов, обеспечивающих рождение «пробирочных» детей, является выдающимся достижением биологии XX в. Можно сказать, что биологи стали подлинными архитекторами жизни.
С другой стороны, решение такой фундаментальной задачи имеет практический результат, связанный с преодолением женского бесплодия. Поэтому сторонники технологии «пробирочных» детей выдвигают весьма важный довод в ее пользу, заключающийся в том, что таким путем решается большая общечеловеческая задача — любовь к ближнему. Что же касается аргументации нарушения прав человека в этой области, то приводимые контраргументы сводятся к тому, что у эмбрионов, образованных оплодотворением в пробирке, еще нет головного мозга и, следовательно, они не могут считаться человеческой жизнью. Следовательно, препятствий для экспериментирования на эмбрионах по этой причине не должно быть.
Итак, оценивая успехи в области оплодотворения яйцеклеток человека in vitro в качестве выдающегося научного достижения XX в., нельзя не отметить, что в этой области возникли серьезные социальные и этические проблемы, но они пока не поддаются решению.
В заключение важно обсудить проблему этики возможных экспериментов, направленных на создание с помощью генетической инженерии новых видов бактериологического оружия. Как известно, идеи использования массовых заболеваний для решения конфликтов уходят в глубокую древность. Например, еще в Ветхом завете сообщается о том, что Иегова наказал египетского фараона тем, что наслал на него мух, вшей и блох. Как повествуется в Ветхом завете, Иегова заслал к ассирийцам и ангела, который распространил чуму, что привело к гибели большого количества людей.
Известно также, что древние римляне забрасывали колодцы враждебной стороны трупами животных, павших от повальных болезней, а воины Александра Македонского перебрасывали трупы павших от повальных болезней животных и людей через стены осажденных ими крепостей.
В более поздние времена также отмечались случаи применения подобного оружия. Например, осаждая в XIV в. крепость Кафу, татары катапультировали через крепостные стены в крепость трупы своих воинов, погибших от бубонной чумы. В XVIII в. англичане в Индии для подавления сопротивления местного населения передавали им одеяла, зараженные возбудителем оспы, что вызвало значительную эпидемию.
С тех пор как было установлено, что инфекционные болезни человека и животных вызываются микроорганизмами, появилось стремление использовать в качестве оружия штаммы вирусов и бактерий, вызывающих особо опасные болезни человека и животных. Поэтому, исходя из чрезвычайной опасности этого оружия, еще в 1925 г. в Женеве был подписан Протокол о запрещении при^ менения на войне удушливых, ядовитых или других подобных газов и бактериологических средств. Однако это международное соглашение не создало запретов бактериологического оружия. Документы свидетельствуют, что во Второй мировой войне в фашистской Германии проводились работы по подготовке к бактериологической войне на Восточном фронте. Однако военные успехи нашей страны не позволили фашистам развязать бактериологическую войну. Но во Второй мировой войне бактериологическое оружие было использовано в широких пределах японскими милитаристами в Китае.
В послевоенный период вопросы запрета биологического оружия, которое стали квалифицировать в качестве оружия массового поражения, неоднократно рассматривались на Генеральной Ассамблее ООН (резолюции 2162 (XXI), 2454А (XXII) от 1966 и 1968 гг. соответственно). Важным явилось подписание в 1972 г. международного соглашения о запрещении разработки, производства и накопления запасов бактериологического и токсинного оружия. Эти документы оказались серьезным сдерживающим фактором. Однако угроза создания и использования биологического оружия не прошла.
Бактериологическим оружием могут быть культуры возбудителей особо опасных болезней (чумы, холеры, тулеремии, бруцеллеза, сапа). В качестве такого оружия могут быть использованы токсины разных бактерий, например, ботулизма и столбняка, а также вирусы ряда болезней (энцефалитов, желтой лихорадки, геморрагической лихорадки). Сюда же относят и гормоны. Однако это бактериологическое оружие относится к оружию «устаревших» видов, хотя его поражающая способность является очень большой (табл. 45).
Значительную опасность в рассматриваемой области таит в себе генная инженерия, т. к. с ее помощью не исключена реальная возможность создания бактериального оружия нового поколения. Методология генной инженерии позволяет создание штаммов бактерий и вирусов, которые уклоняются от диагностики, резистент-ны ко всем современным лекарственным веществам, обладают повышенной вирулентностью и способностью долго находиться в окружающей среде, а также способностью легко приживаться в организме человека и животных и вызывать болезни с неизвестной клинической картиной. Что касается токсинов, то в качестве токсинного оружия могут быть созданы супертоксины. Таким образом, бактериологическое оружие, которое может, быть создано генно-инженерным путем, может иметь принципиально новые признаки, повышающие его способность к массовому поражению.
Таблица 46
Сравнительные характеристики поражающего действия бактериологического, химического и ядерного оружия, которое может быть взято на борт одним стратегическим бомбардировщиком
(по О. В. Варояну, 1971)
| Критерий оценки
|
Вид оружия
|
| ядерное
|
химическое
|
биологическое, бактериологическое
|
| Зона поражающего действия
|
200-300 KM2
|
до 250 км2
|
до 100 000 км2
|
| Время «проявления» поражающего действия
|
секунды
|
от 7 сек до 30 мин
|
От нескольких дней до 2 недель
|
| Поражающее действие
|
Летальность (90%)
|
Поражаемость до 30%
|
Заболеваемость до 25-75%
|
| Возможность выживания
|
Нет
|
Любая
|
Любая
|
| Вред сооружениям
|
Уничтожение на площади до 100 км2
|
Нет
|
Нет
|
| Дополнительные последствия
|
Радиоактивные осадки в зоне 2000 км2
в течение 6 месяцев
|
Заражение местности на срез до нескольких недель
|
Распространение эпидемии
|
| Возможность использования зоны применения
|
Через 3—6 месяцев
|
Ограниченная
|
После окончания инкубации
|
| Скрытность
|
Отсутствует
|
Достижимая
|
Большая
|
| Сравнительная стоимость
|
Очень большая
|
Достаточно велика
|
Относительно малая
|
Поскольку в Конвенции о запрещении биологического и химического оружия нет механизма проверки выполнения этой Конвенции, поскольку в ней не содержатся запреты на исследования с целью создания новых видов такого оружия, в современных условиях чрезвычайно важна позиция самих ученых, использующих в своей работе методы генетической инженерии. Участие ученых-биологов в прямых или косвенных работах по созданию биологического оружия является не только неэтичным, но и преступным деянием.
Литература
БаевА.А. Современная биология как социальное явление. Вопросы философия. 1981, 3,17.
Бароян О. В. Судьба конвенционных болезней. М.: Медицина. 1971. 372 стр.
Дубинин Н. П. Что такое человек. М.: Мысль. 1983. 334 стр.
Дубинин Н. П., Карпец И. И., Кудрявцев В. Н. Генетика, поведение, ответственность. Политиздат. 1982. 304 стр.
Пехов А. П. Социальные проблемы генетики. М.: Знание. 1976. 62 стр.
Пехов А. П. Биология и научно-технический прогресс. М.: Знание. 1984. 64 стр.
Beauchamp Т., Childress J. Principles of Biomedical Ethics. OUP, N. Y. a. Oxford, 1989. 470 pp.
Czifco G. Without Micacles: Universal Selection Theory and the Second Darwinian Revolution. MIT Press. 1996. 386 pp.
Engelhardt H. The Foundation of Bioethics. Oxford University Press. 1986. 398 pp.
Gibson K. R., Ingold T. (Ed.) Tools, Language and Cognition in Human Evolution. Cambridge University Press. 1966. 482 pp.
Современное состояние биологических наук позволяет сформулировать обобщения, основанные на достижениях этих наук, а именно:
1. Жизнь является качественно особой высшей формой движения материи, в основе которой лежит самообновление в виде самовоспроизведения. Ее начальные формы возникли в результате длительного химического процесса из неорганических веществ (необиогенеза).
В основе современной теории происхождения жизни лежат представления о том, что жизнь является результатом исторического одностороннего развития в виде усложнения органических субъединиц и развития их в сложные системы, обладающие свойствами живого.
Живая материя (живые организмы) состоит из множества органических соединений, которые представляют собой соединения углерода в виде крупных молекул (биологических макромолекул). Будучи просты по строению, биологические макромолекулы являются строительными блоками, из которых построены все современные организмы. Специфика организмов любого вида определяется набором присущих только им белков, которые в сочетании с нуклеиновыми кислотами в виде нуклеопротеидов выполняют роль субстрата жизни. Более простые молекулы, из которых состоят макромолекулы, выполняют в клетках несколько функций. Например, аминокислоты служат не только строительными блоками белков, но и являются предшественниками многих гормонов животных, алкалоидов и пигментов растений, а также других макромолекул, тогда как нуклеотиды, являющиеся строительными блоками нуклеиновых кислот, могут служить коферментами и переносчиками энергии.
2. Жизнь дискретна. Она проявляется на многих уровнях, однако элементарными структурно-функциональными единицами жизни являются только клетки, т. к. они представляют собой крайние живые структуры, из которых состоят организмы и с которых начинается жизнь. Поскольку за пределами клеток жизни нет, то воспроизводство клеток заключается в том, что они происходят только от клеток, т. е. все живое происходит только от живого. Формируя многоклеточные организмы, клетки характеризуются сходством строения, различаясь между собой лишь по размерам, форме и функциональной специализации. Все они обладают клеточной мембраной у животных или клеточной стенкой у растений, мембранной системой, ядром, митохондриями (хлоропластами в клетках растений), рибосомами и другими органеллами. Живые клетки представляют собой структуры (изотермические системы органических молекул), способные к самосборке, саморегуляции и самовоспроизводству. Протекающие в клетках реакции синтеза и распада ускоряются ферментами, продуцируемыми самими клетками. Самовоспроизводство клеток контролируется генетически. Исключение по сравнению со свойствами клеток одноклеточных и многоклеточных организмов составляют вирусы, которые представляют собой неживые внеклеточные надмолекулярные структуры, способные к размножению лишь в клетках, где они подчиняют биохимический аппарат клетки с целью производства новых вирусных частиц.
3. Размножение и дифференцировка клеток составляют основу роста и размножения организмов.
Современные экспериментальные разработки показали, что оплодотворение возможно в пробирке, причем яйцеклетки могут быть оплодотворены не только мужскими половыми клетками, но и ядерным материалом, происходящим из соматических клеток. В последнем случае это означает, что генетическая программа яйцеклеток может быть перепрограммирована и что половое размножение высших животных в эксперименте может быть заменено бесполым размножением. Это есть одно из современных доказательств происхождения полового размножения из бесполого.
4. Для всех организмов характерно единство метаболических процессов. Рост и развитие организмов обеспечиваются свободной энергией, которую они поглощают из окружающей среды и которая способна совершать определенную работу. При этом организмы возвращают в среду менее полезную энергию, например, в форме тепла, которая легко рассеивается во Вселенной, превращаясь в энергию хаотического (беспорядочного) движения. Степень рассеивания (обесценивания) энергии определяется энтропией.
Метаболизм есть совокупность катализируемых ферментами процессов, заключающихся, в основном, в обеспечении клеток энергией, получаемой преобразованием энергии солнечного света или расщеплением пищи, поступающей в организм, переводе молекул пищи в блоки, используемые затем для образования макромолекул, сборке биологических макромолекул, а также в синтезе (анаболизме) и распаде (катаболизме) биологических макромолекул, выполняющих специфические функции тех или иных клеток.
Живые организмы являются открытыми системами. Главным источником энергии для живых организмов является лучистая энергия солнечного света, которую улавливают клетки зеленых растений и в процессе фотосинтеза преобразуют эту энергию в химическую энергию углеводов (пищи). Химическая энергия углеводов пищи преобразуется в клетках организмов-гетеротрофов в электрическую, механическую и другие виды энергии. Зеленые растения являются первичными преобразователями энергии Солнца, а фотосинтез зеленых растений и хемосинтез, осуществляемый микроорганизмами отдельных видов, являются одним из этапов в энергетическом обеспечении живого мира. Все живые организмы (кроме растений) обеспечиваются энергией в результате потребления продуктов фотосинтеза или хемосинтеза.
Пищевые вещества в клетках используются с помощью реакций с участием ферментов, причем освобождающаяся при этом энергия запасается в форме макроэргетических связей аденозинтрифосфата (АТФ), который служит в качестве источника энергии для обеспечения многих функций организмов, включая их рост и размножение. АТФ связывает в единое целое системы ферментативных реакций, которые обеспечивают сохранение поступающей из окружающей среды энергии фосфорилированием, с одной стороны, и биосинтез клеточных структур, механизм двигательных и сократительных процессов клеток, а также осмотическую клеточную работу, с другой стороны.
Метаболизм подвержен непрерывной генетической регуляции, которая осуществляется в клетках путем регуляции синтеза ферментов. В клетках синтезируется столько малых молекул, сколько их необходимо для синтеза нуклеиновых кислот, белков, липидов и углеводов. Это позволяет оценивать клетки в качестве чрезвычайно экономичных саморегулируемых химических систем.
5. Для жизни характерны непрерывность, устойчивость и материальная преемственность, которые обеспечиваются наследственностью и изменчивостью организмов. Элементарными единицами наследственности являются гены. Закономерности передачи генов от родителей к их потомству определяются двумя законами наследственности. В соответствии с первым законом (законом расщепления) гены в соматических клетках на их хромосомах содержатся в парах, которые расщепляются при гаметогенезе и расходятся в разные гаметы. В соответствии со вторым законом (законом независимого распределения генов или рекомбинации) расщепление одной пары генов и расхождение их в разные гаметы не зависят друг от друга, но при этом происходит рекомбинация генов, дающая начало организмам с рекомбинантными признаками. Законам наследственности подчиняется весь живой мир, включая человека.
Гены подвержены мутациям, которые сопровождаются изменением количественных и качественных признаков растений, животных и человека. В случае человека мутации проявляются в виде наследственных болезней. Для популяций организмов любого вида характерно генетическое равновесие, определяемое неизменностью содержания нормальных и мутантных генных аллелей. Мутантные организмы являются материалом для действия естественного отбора.
6. Генетическим материалом всех организмов является ДНК, сосредоточенная в основном в ядре клеток (в хромосомах). У эукарио-тов ДНК содержится также в митохондриях и хлоропластах клеток. Экстрахромосомная ДНК представлена плазмидами у бактерий. В ДНК в виде генетического кода зашифрована генетическая информация о структуре белков. Генетический код является триплетным, неперекрывающимся, вырожденным, универсальным. Передача и реализация генетической информации о синтезе белков осуществляется в два этапа, называемые транскрипцией и трансляцией. В период транскрипции информация переписывается с молекул ДНК в молекулы мРНК. Трансляция генетической информации происходит на рибосомах с участием тРНК. Признание такого пути передачи генетической информации (ДНК ® РНК ® белок) получило название центральной догмы биологии. Однако у отдельных организмов (вирусов) известна передача генетической информации по схеме РНК ® ДНК ® белок. В составе ДНК обнаруживают как уникальные, так и повторяющиеся последовательности азотистых оснований. Репликация ДНК является полуконсервативной, причем синтез дочерней цепи на одной цепи идет непрерывно, на другой — прерывисто, давая фрагменты, которые затем воссоединяются с помощью ферментов. Современная концепция гена заключается в положении «один ген — одна тРНК — один полипептид».
Цепи ДНК лишь относительно стойки, в результате чего в ДНК могут возникнуть повреждения в основном в виде тиминовых ди-меров. Однако эти повреждения доступны для репарации (восстановления), которая поддерживает стабильность ДНК.
7. Одним из важнейших обобщений биологических наук является теория эволюции, под которой понимают развитие организмов во времени или процесс исторического преобразования на Земле. Основы теории эволюции были заложены Ч. Дарвином. В рамках этой теории признается, что растения и животные в природных условиях подвергаются изменчивости, что их рождается больше, чем позволяют пищевые и другие ресурсы и что это ведет к борьбе за существование между особями, в которой выживают более приспособленные к изменяющимся условиям среды, т. е. происходит естественный отбор наиболее приспособленных. Помимо естественного отбора элементарными факторами эволюции являются также мутационный процесс, популяционные волны, изоляция, дрейф генов, миграция, с действия которых начинаются процессы эволюции в популяции. Однако фактором, направляющим эволюцию, является естественный отбор, который отбирает организмы, свойства которых благоприятны для данной среды и которые дают достаточно многочисленное потомство.
Чрезвычайное разнообразие живых форм (биоразнообразие) является результатом длительного процесса эволюции, причем способность организмов к жизни в различных условиях является результатом эволюции не только их самих, но и эволюции систем их органов. Эволюции нет «хорошей» и нет «плохой». Эволюция может быть лишь только успешной и безуспешной. Организмы, которые хорошо функционируют при определенных условиях и потому дают больше потомства, являются успешными. При других условиях эти же организмы могут быть эволюционно неудачными (безуспешными).
Фундаментальным следствием эволюции является признание того, что все организмы связаны между собой по происхождению, поскольку они происходят от общего предка. Современные доказательства эволюции настолько внушительны, что ее уже можно объяснить не теорией, а понимать как факт. Человек также является результатом эволюции. Эволюционный возраст человека составляет более 150 тыс лет. На ранних этапах эволюции человека действовали те же факторы эволюции, которые действуют в случае эволюции животных и растений, причем направляющим фактором также являлся естественный отбор. Однако как только человек увидел свое отличие от других живых существ, как только начали складываться общественные отношения, биологическая эволюция уступила место социальному совершенствованию человека. В настоящее время естественный отбор почти полностью прекратил свое действие.
8. Живые организмы связаны между собой не только происхождением, но и различными отношениями в процессе их жизни между собой и со средой, в которой они обитают, т.е. они связаны экологически. Эта связь заключается в том, что одни живые организмы (растения и животные) зависят от других организмов. Формируя сообщества, организмы вместе с абиотическими факторами среды образуют экологические системы. В этих системах одни организмы продуцируют органическое вещество (организмы-производители), другие потребляют его (организмы-потребители), а третьи разрушают его (организмы-разрушители).
Экологические системы являются результатом исторического развития природы. Их особенность заключается в том, что для них характерно постоянство, устойчивость и равновесие между компонентами системы. В биосфере непрерывно происходит обмен энергией и материей по схеме организм — среда — организм. Организмы в биосфере нуждаются в углероде, кислороде и азоте, которые необходимы для биосинтеза аминокислот, пуриновых и пиримидиновых оснований, идущих на построение белков и нуклеиновых кислот. По этой причине в биосфере протекают гигантские круговороты углерода, кислорода и азота. Эти круговороты зависят от соотношений между организмами-производителями органического вещества и организмами-потребителями этого вещества. В свою очередь круговороты веществ сопровождаются гигантским круговоротом энергии. Используя энергию Солнца, автотрофные организмы синтезируют углеводы и другие органические соединения, богатые энергией, тогда как гетеротрофные организмы получают энергию из пищи, содержащей углеводы; для потока энергии в биосфере присуща направленность в одном направлении, в котором часть полезной энергии теряется, а количество бесполезной энергии растет.
9. Любое нарушение исторически сложившегося равновесия в природе сопровождается разрушением экологических систем. Следовательно, в основе нарушений в природе лежат нарушения экологического равновесия. В современную эпоху наиболее неблагоприятные воздействия на природу связаны с антропогенным фактором. В погоне за обеспечением собственного благополучия человек разрушает экологические системы, загрязняет природу, насыщает ее различными загрязнителями, в основном радиацией и химическими веществами, которые обладают свойствами мутагенных факторов, вызывая серьезные поражения в генетическом материале человека, сопровождающиеся наследственными болезнями. Между тем осознавая это, человек и дальше будет пользоваться природой, черпая в ней природные ресурсы. В связи с этим возникла гигантской значимости проблема сохранения окружающей среды (природы), необходимость разработки научных основ рационального природопользования, регулирования отношений между человеком и природой. Это регулирование имеет такую значимость в контексте будущего человечества, что позволяет считать его основой дальнейшего развития.
10. Важнейшее достижение в биологических науках относится к генетической инженерии, которое позволяет сделать ряд дополнительных обобщений. Во-первых, генетическая инженерия позволила подтвердить ценность существующих представлений о научной картине мира. Будучи эпохальным методическим достижением, генетическая инженерия не привела к ломке существующих представлений о жизни, но она показала, что эти представления являются справедливыми. Во-вторых, генетическая инженерия подняла на совершенно новый уровень биотехнологию, став ее совершеннейшей методической базой и превратив ее в полноправную сестру механической и химической технологий, созданных ранее. Благодаря генетической открылись невиданные ранее перспективы в сельском хозяйстве, промышленности и медицине.
Однако генетическая инженерия породила и ряд собственных экологических проблем. Признавая необходимость регулирования во взаимоотношениях человека с природой и в генетической инженерии, заключительное обобщение сводится к тому, что регулирование должно стать основной парадигмой развития в XXI в.
В биологических исследованиях используют метрическую систему измерений, с помощью которой измеряют три основные «количества» —длину (объем), массу, время.
В соответствии с Международной системой единиц используют следующие единицы измерений:
Длина: основная единица — метр
1 сантиметр (см) = 0,01 м
1 миллиметр (мм) = 0,001 м
1 микрометр или микрон (мкм) = 0,000001 м
1 нанометр (нм) = 0,000000001 м 1 нанометр — 10 А̊ (ангстремам)
Масса:
основная единица — грамм
1 миллиграмм =° 0,001 г
1 микрограмм = 0,000001 г
1 нанограмм -- 0,000000001 г
Молекулярная масса:
1 дальтон = 1,661´10-24
г
Время: основная единица — секунда
1 миллисекунда = 0,001 сек
1 микросекунда = 0,000001 сек
Объем: основная единица — кубический метр (м3
)
1 кубический сантиметр (см3
) — 0,000001 м3
1 кубический миллиметр (мм3
) = 0,000000001 м3
основная единица — литр
1 миллилитр (мл) = 0,001 л
1 микролитр (л) = 0,000001 л
1 мл всегда равен 1 см3
, а один л = 1 дм3
В соответствии с Международной системой единиц:
Единицы энергии — джоуль (Дж)
1 кал = 4,184 Дж
1500 г. — Установлена 1814 г. — Установлена способность
невозможность выживания экстрактов ячменя превращать
животных крахмал в сахар (Г. Кирхгоф).
в атмосфере, в которой 1823 г. — Отмечены доминантность
не происходит горение и рецессивность признаков
(Леонардо да Винчи), садового гороха (Т. Э. Найт).
1600 г. — Изготовлен первый 1831 г. — Открыто клеточное ядро
микроскоп (Г. Галилей). (Р. Броун).
1628 г. — Открыто кровообращение 1839 г. — Сформулирована
(В. Гарвей). клеточная теория (Т. Шванн,
1651 г. — Сформулировано М. Шлейден).
положение «Все живое 1839 г. — Сформулировано
из яйца» (В. Гарвей). положение о «неживой»
1661 г. — Открыты капилляры природе ферментов
(М. Мальпиги). (Ю. Либих).
1665 г. — Обнаружена клеточная 1845 г. — Впервые синтезировано
структура пробки органическое соединение
(Р. Гук). (уксусная кислота) из
1668 г. — Экспериментально неорганических
доказано развитие личинок предшественников.
мух из отложенных яиц 1858 г. — Сформулировано
(Ф. Реди). положение «Каждая клетка
1674 г. — Открыты бактерии и из клетки» (Р. Вирхов).
простейшие (А. Левенгук). 1859 г. — Опубликована книга
1677 г. — Впервые увиден Ч. Дарвина «Происхождение
сперматозоид человека видов путем естественного
(А. Левенгук). отбора, или сохранение
1688 г. — Введено понятие благоприятствуемых пород
о виде как систематической в борьбе за жизнь».
единице (Д. Рей). 1862 г. — Опровергнута теория
1694 г. — Экспериментально самопроизвольного
доказано наличие пола у зарождения (Л. Пастер).
растений (Р. Камерариус). 1862 г. — Показано
1727 г. — Установлено воздушное фотосинтетическое
питание растений происхождение крахмала
(С. Гейлс). (Ю. Сакс).
1753 г. — Разработаны принципы 1862 г. — Открыты явления
систематики организмов и торможения в ЦНС
бинарная номенклатура (И. М. Сеченов).
(К. Линней). 1866 г. — Опубликованы законы
1754 г. — Открыт углекислый газ наследственности (Г. Мендель).
(Дж. Влэк). 1871 г. — Доказано, что способность
1766 г. — Открыт водород ферментировать сахар
(Г. Кавендиш). (превращать его в спирт)
1772 г. — Открыто выделение принадлежит не дрожжевым
кислорода растениями клеткам, а содержащимся в
(Дж. Пристли), них ферментам
1779 г. — Показана связь между (М. М. Манассеина).
светом и зеленой окраской 1871 г. — Открыты нуклеиновые
растений (Ян Ингенхауз). кислоты (Ф. Мишер).
1809 г. — Привлечено внимание к 1875 г. — Доказано, что процессы
влиянию среды на окисления происходят в
изменчивость организмов тканях, а не в крови
(Ж. В. Ламарк). (Э. Пфлюгер).
1875 г. — Дано первое описание 1921 г. — Открыто влияние одной
хромосом (Э. Страсбурге?), части зародыша на другую
1878 г. — Предложен термин и выяснена роль этого явления
«энзим» для обозначения в детерминации частей
ферментов (Ф. В. Кюне). развивающегося зародыша
1883 г. — Сформулирована биоло- (Г. Шпеман).
гическая (фагоцитарная) тео- 1922 г. — Открыт лизоцим
рия иммунитета (А. Флеминг).
(И. И. Мечников). 1923 г. — Охарактеризован
1892 г. — Открыты вирусы фотосинтез в качестве
(Д. И. Ивановский), окислительно-
1893 г. — Открыты восстановительной реакции
нитрифицирующие бактерии (Т. Тунберг).
и объяснена их роль 1924 г. — Опубликована
в круговороте азота естественно-научная теория
(С. Н. Виноградский). происхождения жизни на
1897 г. — Показано, что брожение Земле (А. И. Опарин).
может происходить вне живых 1926 г. — Объяснена роль мутаций
клеток, т. е. начато в естественном отборе
исследование гликолиза (С. С. Четвериков).
(Г. и Э. Вухнеры). 1926 г. — Получена
1898 г. — Открыто двойное кристаллическая уреаза
оплодотворение у цветковых (Д. Сампер).
растений (О. Г. Навашин). 1926 г. — Опубликован труд
1900 г. — Вторичное открытие В. И. Вернадского «Биосфера».
законов наследственности 1931 г. — Открыто дыхательное
(К. Корренс, К. Чермак фосфорилирование на уровне
и Г. де Фриз), клеток (В. А. Энгельгардт).
1900 г. — Открыты группы крови 1932 г. — Появление первого
у человека (К. Ландштейнер). электронного микроскопа
1901 г. — Сформулировано просвечивающего типа
представление об условно- (М. Кноль, Э. Руска).
рефлекторной деятельности 1933 г. — Выделены и
(И. П. Павлов), охарактеризованы ауксины
1903 г. — Привлечено внимание растений (Ф. Кегль).
к роли зеленых растений в 1934 г. — Обоснована центровая
космическом круговороте теория гена (Н. П. Дубинин,
энергии и веществ А. С. Серебровский и др.).
(К. А. Тимирязев). 1937 г. — Открыт цикл
1906 г. — Начато использование трикарбоновых кислот
дрозофилы в качестве (Г. А. Кребс).
экспериментальной 1939 г. — Сформулирована теория
генетической модели природной очаговости
(Т. Морган), трансмиссивных болезней
1910 г. — Доказано сцепление генов (Е. Н. Павловский).
в хромосомах (Т. Морган). 1940 г. — Получен пенициллин
1910 г. —Доказано единство (Г. Флори и Э. Чейн).
брожения и дыхания 1940 г. — Сформулирована теория
(С. П. Костычев). биогеоценозов (В. Н. Сукачев).
1910 г. — Сформулирована теория 1941 г. — Экспериментально
филэмбриогенеза доказано, что синтез
(А. Н. Северцов). бактериальными клетками
1920 г. — Открыта нейросекреция факторов роста
(О. Леви). контролируется генами
1920 г. — Сформулирован закон (Д. Билд и Э. Татум).
гомологических рядов 1943 г. — Доказано существование
наследственности спонтанных мутаций
(Н. И. Вавилов). (С. Лурия и М. Дельбрюк).
1944 г. — Доказана генетическая 1968 г. — Осуществлен химический
роль ДНК (О. Эвери, синтез гена (X. Корана).
С. Маклеод и М. Маккарти). 1968 г. — Открыты рестрикционные
1944 г. — Сформулировано учение эндонуклеазы (М. Месельсон,
о девастации гельминтов Р. Юан, С. Ланн, В. Арьер).
(К. И. Скрябин). 1970 г. — Открыта обратная
1946 г. — Открыта система транскрипция (X. Темин,
рекомбинации у бактерий Д. Балтимор).
(Д. Ледерберг и Э. Татум). 1973 г. — Опубликованы результаты
1948 г. — Обосновано единство первых экспериментов по
принципов управления в молекулярному клонированию
технических системах и (С. Коэн, А. Чанг).
живых организмах (Н. Винер). 1975 г. — Открыты гибридомы и
1951 г. — Сформулировано способ получения
представление о вторичной моноклеточных антител
структуре белков и открыта (Ц. Милыптейн).
а-спираль (Л. Полинг). 1982 г. — Показана возможность
1952 г. — Открыты мигрирующие изменения фенотипа
(транспозируемые) млекопитающих (получения
генетические элементы трансгенных мышей) с
растений (В. Макклинток). помощью рекомбинантных
1953 г. — Сформулирован ы молекул ДНК (Р. Полмитер и
представления о структуре Р. Вринстер).
ДНК (Д. Уотсон и Ф. Крик). 1982 г. — Открыта каталитическая
1957 г. — Запущен второй активность РНК (Т. Чек).
искусственный спутник Земли 1988 г. — Установлен фактор,
с Лайкой на борту (СССР), «лицензирующий» и
I960 г. — Синтезирован хлорофилл позволяющий один раунд
(Р. Вудворд). репликации ДНК на клетку
1960 г. — Установлена (Д. Влау, Р. Лаун).
гибридизация культивируемых 1993 г. — Осуществлены первые
соматических клеток эксперименты по индукции
(Г. Барский), монозиготных близнецов
1961 г. — Определены тип и общая человека (П. Стилман и
природа генетического кода Д. Холл).
(Ф. Крик, Л. Варнет, 1994 г. — Идентификация семейства
С. Вреннер, Р. Уотс-Тобин). гомеотических (Нох) генов,
1961 г. — Начато клонирование которые существенны в
животных (Дж. Гёрдон). определении плана строения
1962 г. — Сформулированы хордовых (К. Кеньон).
представления о регуляции 1995 г. — Установлена возможность
активности генов оплодотворения женских
(Ф. Жакоб и Ж. Моно). половых клеток мужскими
1964 г. — Открыты сперматидами (Ж. Тестарт,
транспозируемые Я. Тесарик и К. Мендоза).
(перемещаемые генетические 1997 г. — Установлена возможность
элементы микроорганизмов) получения (клонирования)
(Э. Кондо и С. Митоухаши). потомства млекопитающих
1966 г. — Расшифрован путем оплодотворения
генетический код яйцеклеток, лишенных ядер,
(М. Ниренберг, М. Очоа, ядрами соматических клеток
X. Корана). (И. Вилмут, К. Кэмпбелл и др.).
659
Абиогенез — спонтанное самозарож- Амнион — мембрана, содержащая жидкость, в которой дение организмов находится эмбрион
Автотрофы — организмы, синтези- Анаболизм (ассимиляция) — эндо
рующие органические соединения из термический процесс уподобления по-
неорганических. ступающих в клетку соединений веще-
Адаптация — процесс формирова- ствам самой клетки (биосинтеза
ния признаков у организмов, обеспечи- компонентов клетки из молекул-пред-
вающих их существование в условиях шественников).
той или иной среды. Анаэробное дыхание — реакции рас-
Аденозивдифосфат (АДФ) — рибо- пада глюкозы без участия кислорода.
нуклеотид-5'-дифосфат, служащий ак- Андрогены — мужские половые гор-
цептором фосфатной группы в энерге- моны.
тическом цикле клеток. Анеуплоидия — мутация в виде на-
Аденояинтрифосфат (АТФ) — соеди- рушении нормального количества хро-
нение с богатыми энергией фосфатны- мосом из-за добавления или удаления
ми связями, вовлеченными в перенос одной или более хромосом.
энергии в клеточном метаболизме в ка- Антибиотик — субстанция, продуци-
честве донора фосфатной группы, руемая микроорганизмом (микроскопи-
Азотфиксация — перевод азотфик- ческим грибом), которая подавляет рост
сирующими микроорганизмами азота других микроорганизмов.
атмосферы (N^) в биологически доступ- Антиген — вещество, преимуще-
ную растворимую форму, ственно белковой природы, вызываю-
Активация аминокислоты — фер- щее при введении в организм позвоноч-
ментативное образование эфирной свя- ных образование антител и способное
эй между карбоксильной группой ами- реагировать с антителами в различных
нокислоты и гидроксильной группой ее реакциях.
тРНК. Антикодон — триплет нуклеотидов
Активный транспорт — перенос ра- тРНК, комплементарный триплету (ко-
створенного вещества через клеточную дону) в мРНК.
мембрану, обеспечиваемый энергией. Антимутаген — фактор, предупреж-
Аллель — альтернативная форма од- дающий мутации.
ного и того же генного локуса гомоло- Антитело — субстанция (защитный
гичной хромосомы, или один из пары белок), продуцируемая в организме в
генов, занимающих определенную по- ответ на введение антигена и способ-
зицию (локус) на гомологичной хромо- ная реагировать с антигеном.
соме и детерминирующий альтернатив- Антропогенез — исторический про-
ный признак, цесс эволюционного становления чело-
Аллостерические ферменты — регу- века.
ляторные ферменты, для которых ха- Апостоз — суицид клеток.
рактерно изменение каталитической Ароморфозы — изменения, которые
активности в процессе нековалентно- поднимают на новый, более высокий
го связывания метаболита в участке, уровень морфофизиологическую органи-
не являющемся каталитическим цен- зацию и жизнедеятельность организмов.
тром. Архебиоз — постепенное возникно-
Аминокислоты — органические со- вение и развитие живой материи в те-
единения, содержащие аминогруппу в чение длительного времени.
(а-положении и карбоксильную груп- Ассимиляция (см. анаболизм).
пу. Являются строительными блоками АТФаза — энерготребующий фер-
в белках, мент, гидролизирующий АТФ до АДФ
Аллопатрическое видообразова- и фосфата.
ние — развитие новых видов в реэуль- АТФ-синтетаза — комплексный фер-
тате изоляции организмов исходной мент, синтезирующий АТФ из АДФ и
популяции географическим барьером, фосфата в процессе фосфорилирования.
Аминоацил — тРНК-синтетаза-фер- Аэроб — организм, живущий в при-
мент, который катализирует обраэова- сутствии кислорода.
ние аминоацил-тРНК благодаря энер- Аэробное дыхание — реакции распа-
гии АТФ. да глюкозы в присутствии кислорода.
Бесполое размножение — размноже- Генетический вектор — ДНК (плаз-
ние без объединения гамет, мид, вирусов), способная к автономной
Бинарная номенклатура — система репликации в клетках и способная при-
иаименования организмов, использую- соединять к себе клонируемый сегмент
щая два названия (родовое и видовое). ДНК.
Биогеоценоз (см. экологическая си- Генетический груз — совокупность
стема). неблагоприятных генов, унаследован-
Биосфера — живая оболочка Земли, ных людьми современных поколений от
Властоцель — полость бластулы, людей предыдущих поколений, а так-
Бластула — стадия эмбрионального же возникающих в результате мутаций
развития, в каждом новом поколении.
Генетический код — способ кодиро-
Вакуоль — пространство в цитоплаз- вания структуры белков.
ме, в котором содержится резервный Генетический мониторинг — слеже-
питательный материал или отбросы, ние за генетическими процессами в по-
Видообразование — образование но- пуляциях человека.
вых видов организмов. Генная инженерия — методология
Водородная связь — слабое электро- конструкции и реконструкции молекул
статическое притяжение электроотри- ДНК.
цательного атома к атому водорода. Генный пул — генетический матери-
который ковалентно связан с другим ал вида на данный период времени.
электроотрицательным атомом. Геном — совокупность генов орга-
Восстановление (репарация) ДНК — низма данного вида.
реконструкция поврежденной ДНК. Генотип — генетическая конститу-
Время подуясизни — время, за ко- ция индивидуального организма или
торое происходит распад половины дан- совокупность генов данного организма.
ного соединения. Гермафродитизм — продукция жен-
Второй закон термодинамики — за- ских и мужских половых клеток осо бью,
кон, в соответствии с которым любой имеющей женские и мужские по-
физический или химический процесс ловыежелезы.
сопровождается ростом энергии. Гетерозигота — организм, у которо-
го какая-либо генная пара представле-
Гамета — зародышевая клетка (яй- на доминантным и рецессивным алле-
цеклетка или сперматозоид), лями.
Гаметогенез — рост и дифференци- Гетероплоидия (см. анеуплоидия).
ровна мужских и женских половых Гетеротроф — организм, нуждаю-
клеток. щийся в энергии и углероде (не способ-
Гаметофит — стадия, связанная с ный к синтезу органического вещества).
образованием гамет в жизненном цик- Гибрид — потомство генетически
ле растений, различных родителей, гетерозиготное
Гаплоид — особь (клетка) с одинар- по одной или многим парам генов.
ным набором хромосом (п — число хро- Гидролиз — разложение соединения
мосом). (молекулы на две или более меньших)
Гаплоидия — мутация в виде умень- реакцией с водой.
шения всего набора хромосом. Гидросфера — водная часть био-
Гемоглобин — гемсодержащий бе- сферы.
лок (дыхательный пигмент) красных Гипотеза — предположение, пробное
клеток крови, объяснение или утверждение, которое
Гемоцианин — дыхательный пиг- нуждается в экспериментальном дока-
мент, содержащий медь и встречаю- аательстве.
щийся в крови у членистоногих и мол- Гистоны — хромосомные основные
люсков. белки.
Ген — сегмент ДНК, контролирую- Гликоген — форма углевода, запаса-
щий синтез одного полипептида. емого в клетках.
Генетическая информация — на- Гликолиз — превращение глюкозы
следственная информация, закодиро- в пировиноградную кислоту (тип бро-
ванная в молекулах ДНК или РНК. жения).
Генетическая карта — чертеж, на ко- Гомозигота — организм, у которого
тором показана последовательность хро- какая-либо генная пара представлена до-
мосомных генов в группах сцепления, минантными или рецессивными генами.
Гомология — сходство структур тела Изогамия — половое размножение
разных организмов, обусловленное их при использовании одинаковых гамет.
общим эмбриональным происхождени- Изотопы — радиоактивные или ста-
ем. Структуры могут иметь различные бильные формы элементов, различаю-
функции. щиеся между собой молекулярной мас-
Гонады — репродуктивные органы, сой, но близкие химически к природным
в которых образуются гаметы (яични- формам элементов.
ки, семенники). Изоферменты — формы одного и
Гормон — химическая субстанция, того же фермента, различающиеся меж-
синтезируемая эндокринной железой и ду собой по сходству к субстратам и по
регулирующая активность клеток, активности.
Градуализм — взгляд на эволюцию Инбридинг — родственное скрещи-
как на процесс, протекающий градуаль- вание организмов.
но во времени. Иидуцибельный фермент — фер-
мент, синтез которого индуцируется
Дальтон — масса одного атома во- своим субстратом. Нормально не выра-
дорода (1,66 х 10~24
г), батывается клеткой.
Дезоксирибонуклеотиды — рибонук- Инсерционная последовательность
леотиды, в которых пентозным компо- ДНК — флангирующие последовательно-
нентом является 2-дезокси-П-рибоза. сти азотистых оснований транспоаонов.
Дегидрогеназы — ферменты, ката- Интерферон — белок, который син-
лизирующие процесс удаления двух тезируется вирусзараженными клетка-
атомов водорода из субстрата, ми животных и человека и который
Дезаминироваяие — процесс фер- препятствует заражению этих клеток
ментативного удаления аминогрупп из другим вирусом.
аминокислот. Интрон — транскрибируемая, но не
Дивергенция признаков — расхож- транслируемая последовательность ДНК.
дение признаков в ходе эволюции организмов.-
Калория — количество тепла, необ-
ДНК — полимер (полинуклеотид), ходимого для повышения температуры
обладающий функцией хранения гене- 1 г воды на 1"С (с 14,5 до 15,5'С).
тической информации. Камбий — ростовой слой стебля и
ДНК-лигаза — фермент, который корней древесных растений.
катализирует образование фосфодиэ- Канцероген — фактор, вызывающий
фирных скелетных связей между 3'- рак.
концом одной цепи ДНК и 5'-концом Капилляры — мелкие кровеносные
другой цепи. сосуды, соединяющие артерии с венами.
ДНК-полимераза — фермент, ката- Кариотип — диплоидный набор хро-
лизирующий синтез ДНК из дезокси- мосом, определяемый их числом, вели-
рибонуклеотид-5'-трифосфатов. чиной и формой.
Дрейф генов — изменения в часто- Катаболизм (диссимиляция) — экзо-
тах генов у организмов малых популя- термический процесс распада сложных
ций в результате случайных скрещи- веществ в организме с освобождением
ваний. энергии.
Доминантность — подавление одним Каталитический центр — участок
аллелем проявления другого (рецессив- фермента, который вовлечен в катали-
ного) в паре аллелей, тический процесс.
Дыхание — расщепление молекул Квант — наименьшая единица энер-
питательного вещества путем их окис- гии света.
ления, сопровождаемого освобождени- кДНК (комплементарная) — ДНК,
ем энергии, синтезируемая обратной транскрипта-
зой и комплементарная цепи мРНК.
Естественный отбор — сохранение Киназа — фермент, катализирую-
наиболее благоприятных индивидуаль- щий с помощью АТФ фосфорилирова-
ных различий, обеспечивающих выжи- ние молекулы-акцептора.
вание организмов и их приспособление Клон — вегетативное потомство од-
к среде, ной особи, возникшее бесполым путем.
Коацерваты — агрегаты коллоид-
Зигота — оплодотворенная яйцек- ных капель, удерживаемых вместе
летка, электростатическими зарядами.
Ковалентная связь — химическая мов во времени, ведущие к новым урон-
связь, образуемая парой электронов, ням сложности и появлению организ-
Код генетический — система запи- мов, принадлежащих к более крупным
си генетической информации в ДНК. таксономическим единицам по сравне-
Кодон — триплет азотистых основа- нию с видом.
ний, кодирующий место одной амино- Мезодерма — слой зародышевых
кислоты в полипептиде. клеток между эктодермой и энтодер-
Коллинеарность — линейное соот- мой.
ветствие между нуклеотидной после- Мейоз — редукционное деление ядер
довательностью ДНК и кодируемой ею клеток репродуктивных органов, сопро-
последовательности аминокислот в вождающееся уменьшением числа хро-
белке. мосом в ядре.
Комменсализм — форма существова- Меристема — недифференцирован-
ния организмов двух видов, при кого- ная эмбриональная ткань растений.
рой организмы одного вида извлекают Метаболизм — совокупность хими-
выгоду, не повреждая организмы дру- ческих реакций в клетках (организме),
гого вида. Заключающихся в синтезе и распаде
Конвергентная эволюция — эволю- сложных молекул с образованием про-
ция сходных признаков неродственных межуточных продуктов (метаболитов).
видов. Метаболит — продукт реакций ка-
Конститутивные ферменты — фер- таболизма.
менты метаболизма, нормально присут- Метагенез — вторичная смена поко-
ствующие в клетках, лений.
Конформация — трехмерная струк- Металлофермент — фермент, содер-
тура макромолекул, жащий ион металла в качестве просте-
Конъюгация — форма полового раз- тической группы.
множения, при которой два организма Метамерия — сегментация тела.
формируют между собой контакты и Микроэволюция — мелкие генети-
обмениваются генетическим материа- ческие изменения, ведущие к видооб-
лом. разованию.
Кофактор — низкомолекулярное Митоз — механизм деления сомати-
органическое или неорганическое со- ческих клеток.
единение, которое необходимо для про- Митотический цикл — совокупность
явления активности фермента, процессов, происходящих в клетке от
Кофермент — небелковый органи- одного деления до другого.
ческий компонент фермента, ответ- Мониторинг генетический — слеже-
ственный за его активность (часто ви- ние за генетическими процессами в по-
тамин). пуляциях.
Креационизм — учение о сверхъес- Моносомия — мутация в виде поте-
тественном сотворении Земли и живо- ри одного гомолога в хромосомной паре.
го мира. Молярный раствор — раствор, в ко-
Кребса цикл — серия ферментатив- тором 1 моль вещества, под которым
ных реакций, конвертирующих пиро- понимают молекулярную массу хими-
виноградную кислоту в двуокись угле- ческого элемента, выраженную в грам-
рода, воду и энергию. ' мах, растворен в 1 литре раствора.
Кретинизм — патологическое состо- Морфогенез — совокупность процес-
яние, являющееся результатом тирои- сов, определяющих структурную орга-
дизма до полового созревания, ниаацию клеток и тканей, а также об-
Кроссинговер — обмен частями щую морфологию организмов.
гомологичных хромосом в процессе Морфология — форма или структу-
мейоза. ра организма.
Мутаген — фактор, вызывающий
Литосфера — подразделение биосфе- мутации.
ры (поверхность Земли). Мутагенез — процесс образования
мутаций.
Макромолекула — молекула, моле- Мутация — изменение генетическо-
кулярная масса которой составляет от го материала, сопровождающееся изме-
нескольких тысяч до сотен миллионов, нениями фенотипа.
Макроэволюция — крупномасштаб- Мутуализм — взаимовыгодная связь
ные генетические изменения органиа- между организмами разных видов.
Необиогенез — процесс образования Открытая система — система, кото-
живых форм из неорганических ве- рая обменивается материей и энергией
ществ (в виде химической эволюции), окружающей среды.
Нонсенс-код-кодон,детермини-
рующий окончание синтеза полипеп- Палиндром ДНК — сегмент ДНК, в
тидной цепи. котором обе цепи обладают осевой сим-
Нонсенс-мутация — мутация, сопро- метрией относительно центра.
вождающаяся преждевременным окон- Панспермия — занос жизни на Зем-
чанием синтеза полипептидной цепи. лю с других планет.
Норма реакции — разнообразие фе- Паразит — организм, который жи-
нотипов, возникакпцих в результате вет за счет другого организма.
взаимодействия генотипа с разными Параллельная эволюция — разви-
факторами среды, тие сходств у организмов эволюционно
Нуклеаза — фермент, который спо- сходных линий в результате действия
собен гидролизовать межнуклеотидные сходных факторов селекции.
связи в нуклеиновой кислоте. Партеногенез — развитие организма
Нуклеозид — соединение пуриново- из неоплодотворенной яйцеклетки.
го или пиримидинового азотного оено- Пенетравтноеть — частота проявле-
вания, ковалентно связанного с пен- ния гена, измеряемая частотой появ-
тозой. ления признака популяции.
Нуклеотид — фундаментальная Пептид — две или более аминокис-
структура нуклеиновой кислоты (нук- лоты, ковалентно связанные между со-
леозид, у которого одна из гидроксиль- бой пептидными связями (амидными
ных групп пентоаы фосфорилирована). связями между a-аминогруппой одной
аминокислоты и a-карбокоильной груп-
Обратная мутация — мутация, со- пой другой аминокислоты).
провождающаяся восстановлением ди- Первый закон термодинамики —
кого фенотипа, закон, устанавливающий, что в ходе
Обратная трапскриптаза — РНК-за- всех процессов во Вселенной ее общая
висимая ДНК-полимераза. энергия не остается постоянной.
Овогенез — развитие яйцеклеток. Пиноцитоз — поглощение жидко-
Окислитеяьно-восстановительная стей и высокомолекулярных веществ
реакция — реакция, в которой проис- клетками за счет выпячиваний плаама-
ходит перенос электронов от донорной тической мембраны.
молекулы к акцепторной молекуле. Плазма — жидкая часть крови.
Окислительное фосфорилирова- Плазмида — бактериальная экстрах -
ние — реакция ферментативного пре- ромосомная молекула ДНК.
вращения АДФ в АТФ, сопровождав- Полиплоидия — мутация в виде уве-
мая одномоментным переносом личения кратного набора хромосом.
электронов от субстрата к молекуляр- Полипептид — цепь аминокислот,
ному кислороду, соединенных между собой пептидными
Онтогенез — индивидуальная исто- связями.
рия организма. Полинуклеотид — последователь-
Оператор — последовательность ность нуклеотидов, ковалентно связан-
ДНК, с которой связывается белок-реп- ных между собой путем соединения фос-
рессор. фодиэфирным мостиком 3'-положения
Оперон — единица координирован- пентозы одного нуклеотида с 5'-положе-
ной экспрессии генов, нием следующего за ним нуклеотида.
Оплодотворение — объединение га- Популяция — совокупность особей
мет, сопровождающееся образованием одного вида, населяющих определен-
зиготы. ную территорию, скрещивающихся
Осмос — диффузия воды через по- между собой и изолированных от дру-
лупроницаемую мембрану, разделяю- гих популяций этого вида.
щую два раствора, из меньшей концен- Приматы — отряд плацентарных
трации в большую, млекопитающих (обезьяны, человек).
Осмотическое давление — давле- Прокариоты — организмы, не име-
ние, которое является результатом ющие ядерной оболочки.
диффузии воды через мембрану (из Протеолитический фермент — фер-
менылей концентрации раствора в мент, катализирующий гидролиз бел-
болылую). ков или пептидов.
Радиоактивный изотоп — изотоп, в Спора — репродуктивная клетка в
котором происходит переход нестабиль- бесполом размножении организмов.
ного ядра в стабильное. Структурный ген — ген, который
Размножение — свойство организ- кодирует структуру белка.
мов производить потомство, или способ- Сцепление— одновременное насле-
ность организмов к самовоспроизведе- дование двух или более генов, локали-
нию, продолжению своего вида. зованных на одной хромосоме.
Раса — исторически сложившаяся
группировка людей, характеризующих- Таксой — единица классификации
ся сходством морфологических и фи- организмов.
зиологических свойств, а также общ- Теория — научное объяснение хоро-
ностью занимаемых ими территорий, шо установленных фактов.
Расогенез—процесс образования рас. Тиминовый димер— ковалентно
Регенерация — восстановление уте- соединенные два тиминовых остатка в
рянных или поврежденных частей тела. молекуле ДНК.
Регуляторный ген — ген, контроли- Токсин — субстанция, продуцируе-
рующий экспрессию другого гена. мая одним организмом и токсичная для
Рекомбинация — новое сочетание других организмов.
генов, устанавливаемое в ходе мейоза Топоизомеразм — ферменты, вызы-
или генно-инженерным путем, вающие сверхскручивание ДНК.
Рекомбинантная ДНК — ДНК, обра- Трансаминазы — ферменты, катали-
зованная воссоединением последова- зирующие перенос аминогрупп от од-
тельностей разного происхождения, ного метаболита к другому.
Репрессор — белок, блокирующий Транзиция — перекрестная замена
транскрипцию ДНК путем связывания оснований в ДНК.
с оператором. Трансверсия — замена одного осно-
Рестриктазы — ферменты «разреза- вания на другое в ДНК.
ющие» ДНК в определенных участках. Транскрипция — переписывание
Ретровирус — вирус, содержащий информации с ДНК в мРНК.
РНК и РНК-зависимую ДНК-полимера- Трансляция — реализация генети-
зу (обратную транскриптазу, ревертазу). ческой информации.
Рецессивный ген — ген, который не Транспозон — мобильный, переме-
экспрессируется в присутствии доми- щающийся генетический элемент (сег-
нантного аллеля. мент ДНК).
Рибонуклеаза — фермент, катализи- Транспорт электронов — переход
рующий гидролиз межнуклеотидных электронов от субстрата к кислороду в
связей в РНК. дыхательной цепи.
РНК — рибонуклеиновая кислота. Трисомия — мутация в виде превраще-
ния дубликата хромосомы в трипликат.
Сбраживание — расщепление глю- Тропосфера — подразделение био-
козы в отсутствие кислорода с образо- сферы (нижняя часть атмосферы).
ванием энергии.
Сведберг (S) — единица скорости Фагоцитоз — переваривание твер-
седиментации (осаждения) частиц при дых частиц клетками.
центрифугировании. Фенотип — совокупность внешних и
Свободная энергия — часть общей внутренних признаков организма.
энергии системы, благодаря которой си- Фермент — органический катализатор.
стема может выполнять работу (при Ферментация — аэробное дыхание
постоянных температуре и давлении), посредством ферментов.
Система — совокупность материи. Филогенез — историческое развитие
изолированная от окружающей среды видов.
(остальной материи во Вселенной). Фитопланктон — плавающие фито-
Симбиоз — сожительство организ- синтезирующие организмы.
мов разных видов, из которого оба из- фосфорилирование — образование
влекают выгоду, фосфатного производного биологической
Сперматогенез — развитие половых молекулы в результате ферментативно-
клеток. го переноса фосфатной группы от АТФ.
Сплайсинг — ферментативное при- Фотосинтез — синтез углеводов зе-
соединение одного сегмента ДНК к дру- леными растениями под влиянием сол-
гому после вырезания интронов. нечного света.
Хемосинтез — процесс, в котором Чистая линия — потомство орга-
энергия для синтеза органических пи- низма, полученное в результате его
тательных веществ образуется в резуль- самоопыления или самооплодотво-
тате окисления простых неорганичес- рения.
ких соединений.
Хемотаксис — движение клеток к Эволюция — развитие организмов
химическому фактору среды, и преобразование их во времени.
Химерная ДНК — рекомбинантная Экологическая система (биогеоце-
ДНК, созданная генно-инясенерным ме- ноз) — элементарная единица биосфе-
тодом. ры в виде совокупности живых орга-
Химическая эволюция — постепен- низмов и неживых элементов на опре-
ная трансформация неживого вещества деленной территории.
и энергии в живые клетки. Экологическая сукцессия — замена
Хищничество — взаимодействие сообществ одних организмов сообще-
организмов, при котором один (хищ- ствами других организмов.
ник) убивает другого (жертву). Эксоц — транскрибируемая и транс-
Хлоропласт — клеточная органелла, лируемая последовательность ДНК.
содержащая хлорофилл. Экспрессивность — уровни (сила)
Хорион — мембрана эмбриона у реп- фенолитической выраженности гена.
тилий, птиц и млекопитающих. Эктодерма — наружный слой гаст-
Хроматин—вещество хромосом, рулы (наружный зародышевый лис-
Хромосома — структурное образова- ток).
ние ядра клеток, содержащее гены. Электрофорез — перемещение заря-
женных частиц в электрическом поле.
Целлюлоза — сложный углевод, со- Эмбрион — организм на ранней ста-
держащийся в клеточных стенках рас- дии развития.
тений, Эндонуклеаза — фермент, гидроли-
Центральная догма биологии — зирующий внутренние фосфодиэфир-
принцип, в соответствии о которым ге- ные связи в ДНК.
нетическая информация передается по Эндосперм — питательная ткань се-
схеме ДНК -> РНК -> белок, мени.
Цитозоль — водная фаза цитоплаз- Энтодерма — внутренний слой гас-
мы с растворенными в ней вещества- трулы (внутренний зародышевый лис-
ми. ток).
Цитокинез — разделение цитоплаз- Энтропия — уменьшение свободной
мы с образованием двух дочерних кле- энергии и повышение той части внут-
ток. ренней энергии системы, которая яв-
Цитоплазма — клеточная субстан- ляется мерой степени неупорядоченно-
ция, окружающая ядро. сти системы.
Цитохром С — одна из групп желе- Эпителий — слой клеток, покры-
зосодержащих белковых субстанций, вающих поверхность полостей орга-
функционирующих в качестве кофер- низма.
мента в дыхании клеток. Эритроцит — красная клетка крови.
Эстрогены — женские половые гор-
Чередование поколений — тип раз- моны.
множения, в котором есть чередование Эукариот — организм, клетки кото-
полового и бесполого поколений, рого содержат оформленное ядро.
Биофизика
Биохимия
Ботанический журнал
Генетика
Журнал общей биологии
Журнал эволюционной биохимии и физиологии
Зоологический журнал
Известия РАН (серия биологическая)
Микробиологический журнал
Сельскохозяйственная биология
Успехи современной биологии
Цитология
Экология
Экология и жизнь
Annual Review of Cell Biology
Annual Review of Biochemistry
Annual Review of Ecology and Systematics
Annual Review of Genetics
Annual Review of Immunology
Annual Review of Microbiology
Annual Review of Physiology
Nature
 Александр ПЕХОВ
Александр ПЕХОВ
профессор,
доктор биологических наук,
заслуженный деятель науки
Российской Федерации,
Академик
Международной Академии Наук
Высшей Школы
Российский университет
дружбы народов
СОДЕРЖАНИЕ
ОТ АВТОРА......................................................................................................................................................................................... 5
Методические рекомендации............................................................................................................................................... 7
ВВЕДЕНИЕ.......................................................................................................................................................................................... 8
Раздел I
БИОРАЗНООБРАЗИЕ (разнообразие
живого мира)
Глава I
ПРИНЦИПЫ И МЕТОДЫ
КЛАССИФИКАЦИИ ОРГАНИЗМОВ............................................................. 19
§1 Искусственные системы................................................................................................................................................. 20
§2 Естественные системы.................................................................................................................................................... 21
§3 Методы классификации.................................................................................................................................................. 22
Вопросы для обсуждения...................................................................................................................................................... 23
Литература................................................................................................................................................................................... 23
Глава II
РАЗНООБРАЗИЕ РАСТЕНИЙ.............................................................................................................................. 24
§4 Надцарство доящерные организмы
(PROCARYOTA)......................................................................................... 25
§5 Надцарство ядерные организмы (EUCARYOTA).................................................................................................. 28
Вопросы для обсуждения...................................................................................................................................................... 41
Литература................................................................................................................................................................................... 42
Глава III
РАЗНООБРАЗИЕ ЖИВОТНЫХ......................................................................................................................... 43
§ 6 Подцарство простейшие (PROTOZOA)....................................................................................................................... 43
§ 7 Подцарство многоклеточные (METAZOA).............................................................................................................. 49
Вопросы для обсуждения...................................................................................................................................................... 71
Литература................................................................................................................................................................................... 72
Глава IV
Разнообразие вирусов............................................................................................................................... 73
§8 Общие свойства вирусов.................................................................................................................................................. 73
§9 Вирусы животных, растений и бактерий.................................................................................................................. 76
§10 Происхождение вирусов................................................................................................................................................ 80
Вопросы для обсуждения...................................................................................................................................................... 82
Литература................................................................................................................................................................................... 82
Раздел II
Живые системы: клетка, организм.............................................................................................. 83
Глава V
СУЩНОСТЬ ЖИЗНИ, СВОЙСТВА И УРОВНИ ОРГАНИЗАЦИИ ЖИВОГО.................................... 84
§11 Сущность и субстрат жизни......................................................................................................................................... 84
§12 Свойства живого............................................................................................................................................................... 85
§13 Уровни организации живого....................................................................................................................................... 89
Вопросы для обсуждения...................................................................................................................................................... 94
Литература................................................................................................................................................................................... 95
Глава VI
КЛЕТКА - ОСНОВНАЯ ФОРМА ОРГАНИЗАЦИИ ЖИВОЙ МАТЕРИИ............................................ 96
§14 Методы изучения клеток.............................................................................................................................................. 98
§15 Структурно-функциональная организация
прокариотических клеток.................................................. 98
§16 Структурно-функциональная организация
эукариотических клеток................................................ 101
§17 Химический состав....................................................................................................................................................... 114
§18 Размножение клеток..................................................................................................................................................... 128
§19 Ткани животных и растений.................................................................................................................................... 133
§20 Эволюция клеток и тканей......................................................................................................................................... 139
Вопросы для обсуждения.................................................................................................................................................... 142
Литература................................................................................................................................................................................. 143
Глава VII
ОБМЕН ВЕЩЕСТВ И ЭНЕРГИИ.................................................................................................................. 145
§21 Анаболизм и катаболизм............................................................................................................................................ 145
§22 Поступление вещества в клетки............................................................................................................................. 149
§ 23 Фотосинтез. Хемосинтез.......................................................................................................................................... 153
§ 24 Подготовка энергии к использованию
(дыхание)....................................................................................... 156
§25 Использование энергии в клетках.......................................................................................................................... 163
§26 Метаболизм на уровне организмов....................................................................................................................... 164
§27 Происхождение типов обмена................................................................................................................................. 166
Вопросы для обсуждения.................................................................................................................................................... 167
Литература................................................................................................................................................................................. 168
Глава VIII
РАЗМНОЖЕНИЕ, РОСТ И ИНДИВИДУАЛЬНОЕ РАЗВИТИЕ ОРГАНИЗМОВ....................... 169
§ 28 Бесполое размножение............................................................................................................................................... 170
§29 Половое размножение. Сперматогенез
и овогенез...................................................................................... 171
§30 Оплодотворение.............................................................................................................................................................. 181
§31 Чередование поколений.............................................................................................................................................. 184
§32 Половой диморфизм. гермафродитизм................................................................................................................ 186
§33 Онтогенез, его типы и периодизация.................................................................................................................... 187
§ 34 Промбриональный и эмбриональный
периоды.............................................................................................. 190
§ 35 Гистогенез и органогенез.......................................................................................................................................... 193
§ 36 Постэмбриональный период..................................................................................................................................... 199
§ 37 Онтогенез растений...................................................................................................................................................... 212
§38 Происхождение способов размножения............................................................................................................. 214
Вопросы для обсуждения.................................................................................................................................................... 215
Литература................................................................................................................................................................................. 216
Раздел III
НАСЛЕДСТВЕННОСТЬ
и ИЗМЕНЧИВОСТЬ ОРГАНИЗМОВ...................................................... 217
Глава IX
НАСЛЕДСТВЕННОСТЬ, НЕПРЕРЫВНОСТЬ ЖИЗНИ И СРЕДА.................................................... 218
§ 39 Наследственность и непрерывность жизни...................................................................................................... 218
§ 40 Наследственность, изменчивость и среда....................................................................................................... 219
§ 41 Методы, генетические модели и
уровни изучения неслественности................................................ 226
Вопросы для обсуждения.................................................................................................................................................... 228
Литература................................................................................................................................................................................. 229
Глава X
ГЕНЕТИЧЕСКИЙ МАТЕРИАЛ......................................................................................................................... 230
§42 Химия и структура ДНК............................................................................................................................................... 232
§ 43 Ядерные (хромосомные) детерминанты
наследственности.................................................................. 236
§ 44 Экстраядерные (экстрахромосомные)
детерминанты наследственности..................................... 239
§ 45 Транспортируемые генетические элементы.................................................................................................... 242
§ 46 Репликация ДНК и хромосом.................................................................................................................................. 244
§ 47 Мутации............................................................................................................................................................................. 249
§ 48 Репарация повреждений ДНК.................................................................................................................................. 262
§49 Эволюция генов и геномов клеток......................................................................................................................... 266
Вопросы для обсуждения.................................................................................................................................................... 268
Литература................................................................................................................................................................................. 269
Глава XI
ЗАКОНОМЕРНОСТИ ПЕРЕДАЧИ
ГЕНЕТИЧЕСКОЙ ИНФОРМАЦИИ.
ГЕНЕТИЧЕСКАЯ ОРГАНИЗАЦИЯ ХРОМОСОМ........................................................................................................... 269
§50 Доминантность и рецессивность............................................................................................................................ 270
§51 Расщепление (сегрегация) генов............................................................................................................................. 272
§52 Независимое распределение генов....................................................................................................................... 277
§ 53 Хромосомные основы расщепления
и независимого перераспределения генов......................... 286
§54 Наследственность, сцепленная с полом............................................................................................................. 287
§55 Сцепление и кроссинговер......................................................................................................................................... 291
§ 56 Молекулярные механизмы и генетический
контроль рекомбинаций................................................. 295
§57 Линейный порядок и тонкое строение генов.................................................................................................... 296
§ 58 Современная кнцепция гена.................................................................................................................................... 297
Вопросы для обсуждения.................................................................................................................................................... 299
Литература................................................................................................................................................................................. 301
Глава XII
ДЕЙСТВИЕ ГЕНОВ............................................................................................................................................. 302
§ 59 Структура и свойства генетического кода....................................................................................................... 303
§60 Транскрипция и трансляция....................................................................................................................................... 305
§ 61 Митохондриальный и хлоропластный
генетические коды..................................................................... 316
§ 62 Универсальность и происхождение
генетического кода.......................................................................... 317
§ 63 Генетический контроль экспрессии генов
Вопросы для обсуждения.................................................. 326
Литература................................................................................................................................................................................. 326
Глава XIII
НОРМАЛЬНАЯ И ПАТОЛОГИЧЕСКАЯ
НАСЛЕДСТВЕННОСТЬ ЧЕЛОВЕКА...................... 328
§ 64 Методы изучения наследственности
человека............................................................................................ 328
§ 65 Нормальная наследственность............................................................................................................................. 332
§ 66 Наследственность и поведение............................................................................................................................. 337
§ 67 Генетическая индивидуальность.......................................................................................................................... 340
§ 68 Патологическая наследственность...................................................................................................................... 342
§ 69 Генетические принципы диагностики,
лечения и профилактики наследственных
болезней 356
Вопросы для обсуждения.................................................................................................................................................... 359
Литература................................................................................................................................................................................. 360
Раздел IV
ЭВОЛЮЦИЯ ОРГАНИЧЕСКОГО МИРА.................................................................................................. 361
Глава XIV
ТЕОРИЯ ЭВОЛЮЦИИ ..................................................................................................................................... 362
§ 70 Представления об эволюции до
ЧарлзаДарвина............................................................................................ 362
§ 71 Ч. Дарвин и его теория эволюции........................................................................................................................... 368
§ 72 Современные представления о
происхождении жизни.............................................................................. 371
§ 73 Ход, главные направления и доказательства
эволюции........................................................................... 377
§ 74 Учение о микроэволюции и видообразование................................................................................................ 394
§75 Гипотеза нейтральности молекулярной
эволюции...................................................................................... 405
§ 76 Антидарвиновские концепции эволюции......................................................................................................... 405
Вопросы для обсуждения.................................................................................................................................................... 407
Литература................................................................................................................................................................................. 407
Глава XV
ПРОИСХОЖДЕНИЕ ЧЕЛОВЕКА................................................................................................................. 408
§ 77 Взгляды на антропогенез в прошлом................................................................................................................... 409
§ 78 Концепция животного происхождения
человека........................................................................................ 411
§ 79 Этапы антропогенеза.................................................................................................................................................. 414
§ 80 Прародина человека..................................................................................................................................................... 426
§ 81 Факторы антропогенеза............................................................................................................................................. 428
§ 82 Расы и их происхождение. Расизм........................................................................................................................ 430
§ 83 Экологическое разнообразие современного
человека............................................................................... 442
§ 84 Культурное развитие человека............................................................................................................................... 445
Вопросы для обсуждения.................................................................................................................................................... 447
Литература................................................................................................................................................................................. 448
Глава XVI
ЭВОЛЮЦИЯ СИСТЕМ ОРГАНОВ............................................................................................................... 448
§ 85 Покровы тела................................................................................................................................................................... 449
§ 86 Скелет.................................................................................................................................................................................. 450
§ 87 Пищеварительная система....................................................................................................................................... 453
§ 88 Дыхательная система................................................................................................................................................. 457
§ 89 Кровеносная и лимфатическая системы........................................................................................................... 458
§ 90 Выделительная система............................................................................................................................................ 462
§ 91 Нервная система............................................................................................................................................................ 465
§ 92 Эндокринная система................................................................................................................................................. 468
§ 93 Репродуктивная система........................................................................................................................................... 469
Вопросы для обсуждения.................................................................................................................................................... 471
Литература................................................................................................................................................................................. 471
Раздел V
Экология............................................................................................................................................................. 472
Глава XVII
Организм и среда..................................................................................................................................... 475
§ 94 Абиотические факторы............................................................................................................................................... 475
§ 95 Биотические факторы.................................................................................................................................................. 480
§ 96 Факторы защиты организма (иммунитет)....................................................................................................... 484
§ 97 Пространство, местообитания,
биомы, сообщства...................................................................................... 490
§ 98 Популяции........................................................................................................................................................................ 493
§ 99 Среда обитания, ареалы и экологические
ниши............................................................................................ 496
Вопросы для обсуждения.................................................................................................................................................... 498
Литература................................................................................................................................................................................. 498
Глава XVIII
БИОСФЕРА И ЧЕЛОВЕК............................................................................................................................ 500
§ 100 Подразделения биосферы........................................................................................................................................ 500
§ 101 Экологические системы.......................................................................................................................................... 502
§ 102 Круговорот веществ................................................................................................................................................... 505
§ 103 Устойчивость экосистем. Сукцессии................................................................................................................ 506
§ 104 Антропогенные воздействия и направления................................................................................................. 508
этих воздействий................................................................................................................................................................... 508
§ 105 Охрана природы и среды обитания..................................................................................................................... 515
Вопросы для обсуждения.................................................................................................................................................... 517
Литература................................................................................................................................................................................. 518
Раздел VI
БИОЛОГИЯ, ГЕНЕТИЧЕСКАЯ ИНЖЕНЕРИЯ
и БИОТЕХНОЛОГИЯ........................................ 519
Глава XIX
ГЕННАЯ ИНЖЕНЕРИЯ................................................................................................................................... 521
§ 106 Выделение ДНК............................................................................................................................................................ 522
§ 107 Ферменты-рестриктазы и рестрикция ДНК...................................................................................................... 523
§ 108 Генеические векторы................................................................................................................................................. 526
§ 109 Конструирование рекомбинантных
молекул ДНК..................................................................................... 527
§ 110 Введение рекомбинатных молекуд
ДНК в клетки..................................................................................... 528
Вопросы для обсуждения.................................................................................................................................................... 530
Литература................................................................................................................................................................................. 531
Глава XX
КЛЕТОЧНАЯ ИНЖЕНЕРИЯ.......................................................................................................................... 532
§ 111 Клеточная инженерия у человека
и животных............................................................................................ 532
§ 112 Клеточная инженерия у растений....................................................................................................................... 535
Вопросы для обсуждения.................................................................................................................................................... 537
Литература................................................................................................................................................................................. 537
Глава XXI
НАПРАВЛЕНИЯ ГЕНЕТИЧЕСКОЙ ИНЖЕНЕРИИ............................................................................ 538
§ 113 Производство пищи.................................................................................................................................................... 538
§ 114 Производство источников энергии
и новых материалов....................................................................... 542
§ 115 Генетическая инженерия и медицина.............................................................................................................. 543
§ 116 Экологические проблемы генетической
инженерии................................................................................. 546
Вопросы для обсуждения.................................................................................................................................................... 547
Литература................................................................................................................................................................................. 547
Глава XXII
ФИЛОСОФСКИЕ, СОЦИАЛЬНЫЕ
И ЭТИЧЕСКИЕ ПРОБЛЕМЫ БИОЛОГИИ................... 548
Литература............................................................................................................................................................................... 559
ЗАКЛЮЧЕНИЕ............................................................................................................................................................................. 560
ПРИЛОЖЕНИЯ............................................................................................................................................................................. 566
Единицы измерения............................................................................................................................................................... 566
Некоторые знаментательные даты в
развитии биологии.................................................................................. 567
Словарь........................................................................................................................................................................................ 570
Названия наиболее известных периодических
изданий по биологии........................................................ 577
АЛЕКСАНДР ПЕТРОВИЧ ПЕХОВ
БИОЛОГИЯ С ОСНОВАМИ ЭКОЛОГИИ
Генеральный директор А Л. Кноп
Директор издательства О. В. Смирнова
Главный редактор Ю.А. Сандулов
Художественный редактор С. Л. Шапиро
Оригинал-макет подготовила К. Н, Белькова
Выпускающие А В. Яковлев, Н. К. Белякова
ЛР № 065466 от 21.10.97 г.
Гигиенический сертификат 78.01.07.952.Т.11668.01.99 от 19.01.99,
выдан ЦГСЭН в СПб
Издательство «ЛАНЬ»
lan@lpbl.spb.ru
www.lanpbl.spb.ru
193012, Санкт-Петербург, пр. Обуховской обороны, 277,
издательство: тел.: (812)262-2495, 262-1178;
pbl@lpbl.spb.ru (издательский отдел),
склад № 1:факс: (812)267-2792, 267-1368.
trade@lpbl.spb.ru (торговый отдел),
193029,
пр. Елизарова, 1,
склад № 2: (812)265-0088, 567-5493, 567-1445.
root@lanpbl. spb. ru
Филиал в Москве:
Москва, 7-я ул. Текстильщиков, 5, тел.: (095) 919-96-00.
Филиал в Краснодаре:
350072, Краснодар, ул. Зиповская, 7, тел.: (8612)57-97-81.
Сдано в набор 20.0S.99. Подписано в печать 16.02.2000.
Бумага газетная. Формат eOxSS1
/^. Гарнитура Школьная.
Печать офсетная. Печ. л. 42. Уч.-изд. л. 42.
Тираж 5000 экз. Заказ № 156.
Отпечатано с готовых диапозитивов в АООТ «Типография „Правда"
. 191119, С.-Петербург, Социалистическая ул., 14.
|