| БЕЛОРУССКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
Выпускная работа по
«Основам информационных технологий»
Магистрант
кафедры системного анализа
Чухутина Ольга
Руководители:
ст. преподаватель
Яцков Николай Николаеевич,
ст. преподаватель
Кожич Павел Павлович
Минск – 2009 г.
Оглавление
Оглавление. 2
Список обозначений ко всей выпускной работе. 4
Реферат. 6
Введение. 6
Глава 1.Обзор литературы.. 6
Глава 2.Методика исследований. 8
2.1 Актуальность изучения процессов полимеризации актина. 8
2.2 Реагенты и реакции при полимеризации актина. 8
2.3 Стохастический подход к моделированию биохимических реакций 11
2.4 Алгоритмы имитационного моделирования биохимических реакций 12
2.4.1 Прямой метод. 13
2.4.2 Метод первой реакции. 15
Глава 3.Основные результаты.. 15
Глава 4.Анализ результатов. 17
4.1 Сравнение аналитической и имитационной моделей. 17
4.2 Анализ экспериментальных данных. 19
Заключение. 20
Список литературы к реферату. 21
Список использованных источников. 21
Список публикаций соискателя. 22
Предметный указатель к реферату. 23
Интернет ресурсы в предметной области исследования. 24
Действующий личный сайт в WWW... 25
Граф научных интересов. 26
Презентация магистерской диссертации. 27
Тестовые вопросы.. 28
Список литературы к выпускной работе. 29
Приложения. 31
Список обозначений ко всей выпускной работе
| Реагенты
|
| AT
М
|
g-актин в АТФ форме
|
| AD
М
|
g-актин в АДФ форме
|
| ATF
|
f-актин в АТФ форме
|
| ADF
|
f-актин в АДФ форме
|
| FIB
|
Плюс-концы филамент
|
| FTB
|
Плюс-концы с последним актином в АТФ форме
|
| FDB
|
Плюс-концы с последним актином в АДФ форме
|
| FIP
|
Минус-концы филамент
|
| FTP
|
Минус-концы с последним актином в АТФ форме
|
| FDP
|
Минус-концы с последним актином в АДФ форме
|
| CBM
|
Блокирующий протеин плюс-конца в свободной форме
|
| CBF
|
Блокирующий протеин плюс-конца в связанной форме
|
| FOM
|
Формин в свободной форме
|
| FOF
^
FxB
FxP
|
Формин в связанной форме
Позиция между двумя актинами в филаменте
Все типы плюс-концов филамент
Все типы минус-концов филамент
|
| Реакции
|
| SNUC
|
Нуклеация
|
| FNUC
|
Формин-инициируемая нуклеация
|
| ASTB
|
Элонгация плюс-конца частицей АТФ актина
|
| ASDB
|
Элонгация плюс-конца частицей АДФ актина
|
| ASTP
|
Элонгация минус-конца частицей АТФ актина
|
| ASDP
|
Элонгация минус-конца частицей АДФ актина
|
| ASTF
|
Элонгация плюс-конца с фомином частицей АТФ актина
|
| DITB
|
Диссоциация АТФ актина с плюс-конца
|
| DIDB
|
Диссоциация АДФ актина с плюс-конца
|
| DITP
|
Диссоциация АТФ актина с минус-конца
|
| DIDP
|
Диссоциация АДФ актина с минус-конца
|
| ASCB
|
Блокировка плюс-конца
|
| DICB
|
Разблокировка плюс-конца
|
| CBNU
|
Нуклеация c участием блокирующего протеина
|
| DIFB
|
Отсоединение формина от плюс-конца
|
| ASFB
|
Присоединение формина к плюс-концу
|
| TTOD
|
Переход f-актина из АТФ в АДФ форму
|
| DTOT
|
Переход g-актина из АДФ в АТФ форму
|
Реферат на тему «Применение информационных технологии при моделировании биохимических реакции в процессах полимеризации актина»
Введение
В настоящее время вычислительный эксперимент является одним из основных инструментов развития таких естественнонаучных областей как физика, химия, биология. Как правило, исследуемые в этих научных дисциплинах системы характеризуются значительным числом взаимодействующих компонентов (в свою очередь являющихся подсистемами), многообразием связей между своими компонентами, нелинейностью поведения и, как следствие, сложностью прогнозирования. Поскольку, зачастую, нет простых формул, описывающих поведение модели, а стало быть, и объекта, который описывается моделью, то единственный путь – свести дело к вычислениям, применению численных методов для решения задач.
Одной из наиболее актуальных в настоящее время областей для применения является применение моделирования к молекулярным биологическим процессам и системам. Отличием биологических (клетки) и биомолекулярных (протеины, ДНК, и т.д.) объектов является их чрезвычайная сложность. В то же время, без детальной информации об этих объектах и клеточных процессах невозможна разработка лекарственных препаратов и эффективного лечения таких заболеваний, как различные типы рака, вирусные инфекции, возрастные заболевания (болезнь Альцгеймера, остеопороз).
Глава 1.Обзор литературы
Настоящая работа посвящена построению и анализу моделей полимеризации актина – важного клеточного процесса, обуславливающего динамику клеток. Изучение данного явления кроме научно-исследовательского имеет и прикладное значение. Некоторые патогенные бактерии используют полимеризацию актина для движения внутри инфицированной клетки [1]. Образование и динамика раковых клеток и метастаз в значительной степени зависят от полимеризации актина. Поэтому управление и контроль над данным процессом может позволить снизить активность патогенов и повысить стабильность здоровых клеток.
Несмотря на большое число теоретических работ посвященных этой тематике, формализм моделирования процессов актин-полимеризации не до конца разработан. Таким образом, актуальной является разработка формализма в рамках парарадигмы имитационного моделирования. Хотя подобные работы проводились ранее [2], [3], по-прежнему ощущается недостаток систематического рассмотрения вопроса моделирования актин-полимеризации. Модель Альбертса и Одела разрабатывалась в основном для анализа механических особенностей роста актин-филамент и уделяет очень мало внимания моделированию биохимических реакций. Модель представленная Карлссоном [4] использует крайне неэффективный алгоритм моделирования, и может быть значительно ускорена при использовании алгоритмов моделирования Гиллеспи.
Одним из важнейших вопросов, связанных с данной темой, является построение методики анализа экспериментальных данных с использованием имитационных моделей. Такой подход для обработки результатов биофизических экспериментов уже не нов и использовался при изучении сложных процессов и систем. Он ранее описан в работах групп Схафсмы и Хэмминги [5-7] для задач анализа данных флуоресценции.
Целью данной работы является разработка, реализация и тестирование алгоритмов имитационного моделирования полимеризации актина, позволяющих исследовать влияние различных реагентов и реакций на динамику роста филамент.
В рамках работы были изучены различные процессы, участвующие в полимеризации актина, включая: образование филамент, их элонгацию и диссоциацию мономеров актина на концах филамент, блокировку, фрагментацию, гидролиз , реакции с участием формина. Все разработанные алгоритмы и процессы были реализованы в виде програмного продукта, написанного на языке Java. При выборе языка программирования были учтены несколько очевидных преимуществ, обеспечиваемых Java: независимость от архитектуры компьютера, переносимость, а главное, кроссплатформенность, что позволяет использовать созданную программу на различных компьютерах.
Во второй главе описываются биофизические аспекты процессов актин-полимеризации. Детально рассмотрены основные реагенты и взаимосвязи между ними. Вторая глава работы также описывает методику моделирования химических реакций. Анализируются достоинства и недостатки различных методов моделирования с точки зрения их точности, быстроты вычислений, времени необходимого на разработку конечного алгоритма, применимости для особых случаев.
В третьей главе рассказывается о разработанной модели и ее реализации, обсуждаются сделанные допущения.
Заключительная глава посвящена тестированию алгоритмов и анализу результатов моделирования. Влияние всех учтенных в модели контролирующих механизмов изучено, полученные результаты обоснованы. Также, было проанализировано быстродействие разработанных имитационных алгоритмов. Результаты, полученные с помощью разработанной имитационной модели, были сравнены с аналитическим решением, а также, с экспериментальными данными.
Глава 2.Методика исследований
2.1 Актуальность изучения процессов полимеризации актина
Актин является белком, входящим в состав клеточного скелета, который может находиться в одной из двух взаимопереходящих форм — глобулярной и фибриллярной. Полимеризацией и называется способность глобулярной формы актина (далее g-актин) организовываться в длинные цепи актинов (далее f-актин), которые называются филаментами. Данный процесс играет важную роль в процессах миграции, деления и объединения клеток. Поэтому не удивительно, что нарушение клеточного скелета напрямую связано с такими болезнями как рак, миопарез и нейродегенеративные заболевания [8]. Хотя многочисленные исследования проводятся в настоящее время, пока не существует полного физико-химического описания происходящих процессов. До сих пор еще остается неизвестно, почему рост филаментов происходит достаточно быстро по сравнению с другими процессами, почему удлинение филаментов способно воздействовать на мембрану, вызывая движение клеток в определенном направлении, каков механизм передачи энергии АТФ для роста филаментов, и как можно управлять этими процессами [9].
В данной работе исследуются вопросы, связанные со стохастическим моделированием и анализом процессов полимеризации актина. Поскольку проведение экспериментов с реальными организмами часто не дает возможности получить полной картины процесса, что обуславливает использование имитационных моделей, например, для нахождения скоростей реакции.
2.2 Реагенты и реакции при полимеризации актина
Мономеры актина имеют грушевидную форму, и при полимеризации возникает спирально закрученная полярная нить с различающимися концами: заостренным (минус) и оперенным (плюс) концом (рис.1).
Такие названия появились в связи с тем, что при взаимодействии актиновых нитей с фрагментами молекулы моторного белка миозина образуется комплекс, имеющий под электронным микроскопом стекловидную форму. При этом острые стрелы указывают на заостренный конец, а ее оперение обращено в сторону противоположного конца филамента. Полимеризация актина происходит в две стадии. Первая стадия

Рисунок.1-Полярная нить актина и круговорот мономеров при полимеризации актина. Рост нити происходит за счет активного присоединения мономеров к оперенному (плюс) концу нити и их более медленной диссоциации с заостренного (минус) конца. По[10]
носит название “нуклеация”, то есть создание ядра (nucleus) или затравки из первых трех мономеров актина. Димер (комплекс двух мономеров) является нестабильной структурой и легко разрушается. Именно нуклеация определяет общую скорость полимеризации. Вторая стадия, удлинение нити, протекает легче, с большей скоростью.
В пробирке мономерный актин может присоединяться и диссоциировать с обоих концов нити, но присоединение происходит быстрее к плюс-концу. Процесс непрерывного присоединения мономеров к оперенному (плюс) концу и их диссоцации с заостренного (минус) представляет собой непрерывный круговорот мономеров. В результате теплого движения филаментов происходит процесс спонтанной фрагментации - самопроизвольный распад цепочки f-актина на две части.
В живой клетке процесс полимеризации актина может быть не похож на такой круговорот мономеров в пробирке, поскольку полярные концы актинового филамента могут быть несвободными.
В клетке присутствуют десятки так называемых актин связывающихся белков, которые сильно влияют на процесс полимеризации актина. Имеется четыре места действия белков: с мономером актина, с плюс-концом , с минус-концом, с боковой поверхностью. Некоторые из них могут блокировать активно растущий оперенный плюс- конец, так называемые кэп-белки, или блокирующие протеины (от англ. cap – шапочка), прекращая таким образом полимеризацию уже существующих нитей и освобождая мономерный актин для построения новых нитей. Другие белки могут разрезать нити актина, формируя тем самым новые фрагменты с заостренными и оперенными концами. Существуют белки, связывающие мономеры актина и таким образом делающие их недоступными для полимеризации (например, профилин), белки, деполимеризующие актин и т.п.[10].
Как f-актин, так и g-актин могут присутствовать в системе в двух состояниях: АТФ-состояние соответствует повышенной вероятности процессов полимеризации, актин в АДФ состоянии быстрее диссоциирует. Переход из АТФ в АДФ состояние происходит, когда молекула актина находится в составе филамента. Обратный процесс возможен для актина в g-форме с участием профилина. Изучение влияния данного протеина достаточно важно, поскольку профилин предположительно является источником энергии для процессов полимеризации. Далее остановимся на некоторых дополнительные протеинах, влияющих на протекающие процессы более подробно:
• АДФ/кофилин – протеин, который может присоединяться к f-актину и стимулировать его отделение от филамента. В таком случае происходит диссоциация мономера, находящегося в любом месте исходной филаменты с ее разделением и образованием двух новых. С одной стороны это производит разрушающий эффект, с другой стороны образование новых концов может ускорять элонгацию при достаточно большой концентрации g-актина
• Кэп-белки (блокирующие протеины) могут присоединяться к свободным концам филаментов, «закрывая» их, то есть, останавливая процессы элонгации и диссоциации для данного конца. Существуют различные виды блокирующих протеинов, которые присоединяются к строго отпределенным концам филаментов (плюс- или минус- концам, оказывая тем самым качественно различное влияние на рост филаментов.
• ARP2/3–комплекс актино-подобных протеинов, имеющих повышенное сродство к минус концам филаментов и может служить матрицей ,на которой происходит образование новых актиновых нитей(нуклеация)(рис.2.а).Кроме того, комплекс может присоединяться к боковым сторонам нитей и таким образом обеспечивать их ветвление(рис.2.б).Эффект ветвления вызывает как правило ускорение элонгации и сильно влияет на механические свойства филамент, которые рассматриваются при исследовании динамических явлений, возникающих в процессе полимеризации актина. Интересно ,что боковые нити актина отходят от основной нити строго под углом 70°,образуя жесткую сеть[10].
• Формин является протеином, ускоряющим нуклеацию филаментов. В процессе роста формин остается прикрепленным к плюс-концу, ускоряя также реакцию элонгации [11].
2.3 Стохастический подход к моделированию биохимических реакций
Имитационное моделирование с использованием парадигмы Монте-Карло основано на получении большого числа реализаций стохастического (случайного) процесса. Данный процесс формируется таким образом, чтобы его вероятностные характеристики совпадали с аналогичными величинами решаемой задачи.
Введем  как функцию, описывающую количество молекул iого вещества в момент времени t. Тогда в момент времени как функцию, описывающую количество молекул iого вещества в момент времени t. Тогда в момент времени 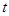 что система находится в состоянии что система находится в состоянии . Чтоб описать изменение ее состояния с течением времени нужно ответить на вопросы: когда наступит следующая реакция и что это будет за реакция. Однако из-за стохастичности реакций эти ответы могут быть даны только с определенной вероятностью. Учитывая предыдущие утверждения, введем функцию . Чтоб описать изменение ее состояния с течением времени нужно ответить на вопросы: когда наступит следующая реакция и что это будет за реакция. Однако из-за стохастичности реакций эти ответы могут быть даны только с определенной вероятностью. Учитывая предыдущие утверждения, введем функцию  , такую, что выражение: , такую, что выражение:
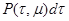 (1) (1)
- определяет вероятность того, что на интервале времени  в системе произойдет реакция в системе произойдет реакция  в объеме V
, если известно, что в момент времени в объеме V
, если известно, что в момент времени 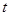 она находилась в состоянии . она находилась в состоянии .
 называется функцией плотности вероятности реакции, поскольку, с математической точки зрения, это объединенная функция плотности вероятности по непрерывному параметру называется функцией плотности вероятности реакции, поскольку, с математической точки зрения, это объединенная функция плотности вероятности по непрерывному параметру   и дискретному и дискретному   . Ценность значений переменных и . Ценность значений переменных и 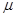 в том, что они дают ответ на те два вопроса, которые были заданы выше: характеризует промежуток времени, за который произошла реакция, а величина непосредственно указывает на номер реакции, которая имела место. Первым шагом на пути поиска приемлемого метода для присваивания численных значений для и является получение аналитического выражения для . в том, что они дают ответ на те два вопроса, которые были заданы выше: характеризует промежуток времени, за который произошла реакция, а величина непосредственно указывает на номер реакции, которая имела место. Первым шагом на пути поиска приемлемого метода для присваивания численных значений для и является получение аналитического выражения для .
Для каждой реакции определим функцию  как количество комбинаций отдельных реагентов , которые возможны в состоянии как количество комбинаций отдельных реагентов , которые возможны в состоянии  . .
Тогда если имеет вид:
 продукт реакции, получим: продукт реакции, получим:
 ; (2) ; (2)
если имеет вид:  продукт реакции, тогда продукт реакции, тогда
 (3) (3)
Таким образом функция представляет собой число всевозможных комбинаций молекул, принимающих участие в реакции μ из набора  .Отсюда: .Отсюда:
 - вероятность того, что на интервале времени - вероятность того, что на интервале времени 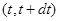 с системой произойдет реакция в объеме V
, если известно, что в момент времени с системой произойдет реакция в объеме V
, если известно, что в момент времени 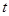 она находилась в состоянии , где она находилась в состоянии , где  - стохастическая скорость реакции. - стохастическая скорость реакции.
Тогда можно вычислить вероятность (1) как произведение(4).
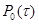 (вероятность того, что интервале времени (вероятность того, что интервале времени  с системой не произойдет никакой реакций в объеме V
, если известно, что в момент времени она находилась в состоянии); с системой не произойдет никакой реакций в объеме V
, если известно, что в момент времени она находилась в состоянии);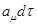 -последующая вероятность, что реакция -последующая вероятность, что реакция  произойдет в интервале произойдет в интервале  : :
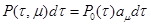 (4) (4)
Для того, чтоб найти выражение для 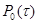 отметим, что отметим, что 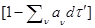 -вероятность того, что за промежуток -вероятность того, что за промежуток  с системой в состоянии с системой в состоянии 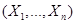 не произойдет никакой реакции. Таким образом: не произойдет никакой реакции. Таким образом:
 (5), (5),
откуда легко получается, что
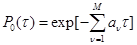 (6) (6)
Подставляя (6) в (4) получим, что функция плотности вероятности реакции, описанная в (1) равна
 , (7) , (7)
где 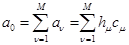 . .
Отметим, что  в (7) - это строгое математическое следствие фундаментальной гипотезы, которое зависит от всех постоянных реакции (не только от в (7) - это строгое математическое следствие фундаментальной гипотезы, которое зависит от всех постоянных реакции (не только от 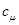 )и от текущего значения молекул всех реагирующих веществ(а не только от реагентов )и от текущего значения молекул всех реагирующих веществ(а не только от реагентов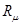 )[21]. )[21].
2.4 Алгоритмы имитационного моделирования биохимических реакций
Для отдельной реализации случайного процесса в каждый момент времени система находится в определенном состоянии. Из этого состояния системы выбирается переход в другое состояние системы на основе вероятностей всех возможных переходов. Ключевым является выбор случайных чисел, используя машинный генератор случайных чисел и их использование для выбора превращений.
Первый из методов, предложенных Гиллеспи, получил название «прямой метод»[12]. Он однозначно вычисляет какая реакция произойдет следующей и когда она произойдет. Второй предложенный метод Гиллеспи назвал «методом первой реакции»[13].Суть метода заключается в генерации для каждой из реакций предполагаемого времени наступления реакции (τ),затем выбирается реакция с наименьшим τ (“первая реакция”)и выполняется данная реакция (μ) в момент времени τ. Позднее был предложен метод τ-leap [14], представляющий собой метод с постоянным шагом по времени. Используя пуассоновскую аппроксимацию для потока реакций, он позволяет оптимизировать время моделирования. Существует широкий спектр задач, где метод τ-leap очень хорошо аппроксимирует стохастичное поведение модели ,однако он является неприменимым для жестких стохастичных задач.
2.4.1 Прямой метод
Этот метод основан на том, что каждая из двумерных функций плотности вероятности может быть записана как произведение двух одномерных функций плотностей, данная операция известна как операция «условия». Введем условную вероятность  как произведение как произведение
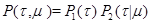 (8) (8)
где –вероятность того, что следующая реакция произойдет в интервале времени от –вероятность того, что следующая реакция произойдет в интервале времени от 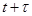 до до  ,а ,а  -вероятность того, что следующей реакцией будет реакция -вероятность того, что следующей реакцией будет реакция 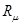 , при условии, что она происходит в момент времени . , при условии, что она происходит в момент времени .
Поскольку
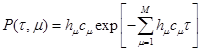 (9), (9),
где  -количество комбинаций молекул отдельных реагентов для реакции -количество комбинаций молекул отдельных реагентов для реакции  , обнаруженных в объеме V
в момент времени t, заменяя , обнаруженных в объеме V
в момент времени t, заменяя 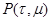 на (9) приходим к результату: на (9) приходим к результату:
 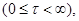 (10) (10)
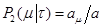  (11), (11),
где для удобства заменим
 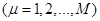 , , . .
Вероятность зафиксировать какую-нибудь реакцию в течение короткого времени  определяется выражением: определяется выражением:
| 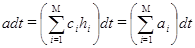
|
(12)
|
В данном выражении  - вероятность зафиксировать хотя бы одну реакцию за единицу времени; - вероятность зафиксировать хотя бы одну реакцию за единицу времени; 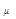 - число всевозможных реакций в системе. Величина - число всевозможных реакций в системе. Величина 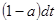 определяет вероятность того, что ни одна реакция не произойдет за время определяет вероятность того, что ни одна реакция не произойдет за время 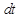 . .
Видно, что в данном случае  не зависит от не зависит от  .Также мы должны учесть, что эти одновариантные функций плотностей нормированы в своей области определения: .Также мы должны учесть, что эти одновариантные функций плотностей нормированы в своей области определения:
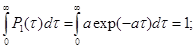 (13) (13)
 (14) (14)
Идеей прямого метода является генерация произвольного значения τ, соответствующему  (13), а затем генерация произвольного целого μ,соответствующего в (14). Результатом является произвольная пара (13), а затем генерация произвольного целого μ,соответствующего в (14). Результатом является произвольная пара  ,помещенная в . ,помещенная в .
Таким образом, время наступления первой реакции рассчитывается методом обратной функции по формуле (15):
|  (15) (16) (15) (16)
|
Номер реакции  определяется методом суперпозиции по формуле (16): определяется методом суперпозиции по формуле (16):
| 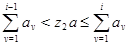 (16) (16)
|
где  - независимые реализации базовой случайной величины - независимые реализации базовой случайной величины

Рисунок
2- Алгоритм прямого метода Гиллеспи
Схема алгоритма моделирования представлен на рисунке 2. Охарактеризуем каждый из блоков:
1. Задание начальных концентраций,скоростей и механизмов реакций, моделируемого объема, времени моделирования, и установление  . .
2. Вычисление 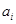 для всех и . для всех и .
3. Вычисление времени наступления реакции, используя  по формуле (15). по формуле (15).
4. Генерирование случайного 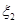 , определяющего номер произошедшей реакции. Определение номера произошедшей реакции по формуле (16). , определяющего номер произошедшей реакции. Определение номера произошедшей реакции по формуле (16).
5. Замена количества молекул согласно произошедшей реакции,
устанавливаем  . .
6. Переход к шагу 2.
Как уже писалось, этот алгоритм использует два случайных числа за итерацию, что занимает времени, пропорционально количеству реакций, для пересчета и μ [12].
2.4.2 Метод первой реакции
Вместе с первым методом Гиллеспи был разработан алгоритм «первой реакции»,который генерирует предполагаемое время наступления каждой из реакции,- время, когда наступит данная реакция, если никакая другая не наступит раньше, потом полагается, что μ -реакция с наименьшим предполагаемым временем, τ переобозначается как 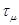 . .
Алгоритм :
1.Задание начальных концентраций,реакций и их скоростей, установление 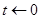 ; ;
2.Вычисление 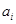 для всех i. для всех i.
3. Генерирование для каждого i с помощью метода обратных функций возможного времени 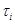 , ,
4.Пусть μ-реакция с наименьшим временем ¸
5.Полагаем τ= ¸.
6.Меняем количество молекул согласно произошедшей реакции μ,устанавливаем ;
7.Переход к шагу 2.
На первый взгляд, эти два метода кажутся очень разными, но они эквивалентны, так как распределения вероятности, используемые для вычисления τ и μ одинаковы. Как уже писалось, данный алгоритм также использует r
случайных чисел за итерацию (r-
количество реакций ), что затратит время, пропорционально r
, чтоб пересчитать все и столько же, чтоб вычислить наименьшее [15].
Глава 3.Основные результаты
Для построения имитационной модели полимеризации актина были сделаны следующие допущения:
· Процесс полимеризации моделируется в кубе,объемом V с периодическими граничными условиями.
· Количество молекул мономеров, например 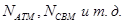 ,в начальный момент времени в объеме V высчитывается из начальных концентрации молекул. ,в начальный момент времени в объеме V высчитывается из начальных концентрации молекул.
· Процесс диффузии происходит намного быстрее ,чем процессы полимеризации, поэтому при моделировании все вещества считаются равнораспределёнными по объему.
Моделировалась динамика роста отдельных филаментов и концентрация всех реагентов в системе. Предполагается, что в начальный момент времени в системе нет филаментов, поэтому начальное состояние задается концентрациями мономеров актина и протеинов в свободной форме.
На рисунке 3 приведена диаграмма, отражающая влияние на процесс полимеризации всех компонентов, реализованных в модели. На схеме представлены как реагенты, так и переходы между ними в результате реакций. Направление ребра в сторону или от эллипса, отображающего конкретную реакцию, соответствует связи с реагентом или продуктом реакции соответственно. Прерывистые линии используются для катализаторов, которые возвращаются в исходное состояние после окончания реакции. Пунктирные линии соответствуют реагентам, которые являются необязательными для протекания соответствующей реакции, однако их участие изменяет скорость реакции.

Рисунок 3-Основные реагенты и реакции в процессе актин-полимеризации
Для генерации событий были реализован вероятностный подход с использованием методики Монте-Карло, представленный в виде трех методов моделирования химических реакций: оригинальный метод Гиллеспи, метод первой реакции и оптимизированный прямой метод Гиллеспи. Метод τ-leap не использовался в данной работе, поскольку по результатам проведенных предварительных тестов доказал свою плохую сходимость при реализации в системах с малым числом молекул.
Реализация же метода Гибсон-Брук в данном случае не возможна, поскольку в соответствующем графе невозможно отразить распад коротких филаментов. Кроме того, из-за сложности системы построение графа становиться неоправданно ресурсоемким. Для филаментов и актинов использована объектная модель, остальные реагенты представлены только числом частиц. Следует отметить, что плюс и минус концы филаментов в различных формах (АДФ и АТФ), представлены как отдельные реагенты, также как и плюс концы филаментов с блокирующим протеином или формином.
Глава 4.Анализ результатов
4.1 Сравнение аналитической и имитационной моделей
Для проверки правильности построения имитационного алгоритма проведено сравнение результатов, полученных с использованием имитационной модели с аналитическим решением для упрощенной системы с меньшим числом реагентов и реакций, приведенным в статье [8]. Анализировалось две системы дифференциальных уравнений.
Аналитическая модель, которая сравнивалась с полученной имитационной дополнительно включает реакции блокировки ,разблокировки и нуклеации с участием блокирующего протеина[8].
 Для тестирования использованы начальные концентрации и скорости реакций, представленные в таблице 1[8,16]. Для тестирования использованы начальные концентрации и скорости реакций, представленные в таблице 1[8,16].
Таблица 12. Начальные параметры, использованные для сравнения моделей (для неуказанных величин начальные значения равны нулю)
| Обозначение
|
Описание
|
Значение
|
| 
|
скорость спонтанной нуклеации
|

|
| 
|
скорость элонгации плюс-конца
|
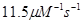
|
| 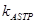
|
скорость элонгации минус-конца
|
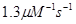
|
| 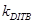
|
скорость диссоциации плюс -конца
|

|
| 
|
скорость диссоциации минус -конца
|

|
| 
|
скорость блокировки плюс-конца
|

|
| 
|
скорость разблокировки плюс-конца
|

|
| 
|
скорость нуклеации с участием блокирующего протеина
|
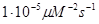
|
| 
|
концентрация g-актина
|

|
| 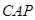
|
концентрация блокирующего протеина
|

|
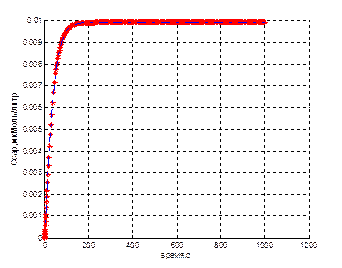
Результаты представлены на рисунке 4.
| 
а
|

б
|
Рисунок 4-Сравнение результатов моделирования. Синие пунктиры –имитационная модель, красные звездочки– аналитическая модель. а – концентрация блокирующего протеина в филаментах (Сcap); б – концентрация f-актина (Сf);
В данном случае наблюдается практические эдентичные результаты, полученные с помощью двух моделей. Это связано с тем, что явления распада коротких филаментов практически не имеет место, в связи с активной нуклеацией и блокировкой филаментов.
Таким образом, полученные результаты позволяют сделать вывод о адекватности и правильности разработанной модели.
4.2 Анализ экспериментальных данных
Для изучения возможности использования разработанной модели для анализа экспериментальных данных,были изучены результаты экспериментов,представленные в статье[17]. В данном эксперименте был использован актин,извлеченный из скелетной мышцы килевидной грудной клетки. Были предварительно очищены g-актины в АТФ форме.В систему также был введен блокирующий протеин  ,извлеченный из кишечной палочки BL21 особым образом. Для изучения динамики процесса полимеризации использовалась силективная флуоресценция пирена.Таким образом в эксперименте были задействованы реакции элонгации,нуклеации ,блокировки и разблокировки.По ходу эксперимента были получены скорости всех реакции. Предполагалось ,что элогация происходит преимущественно на плюс конце филаменты ,поэтому элонгация на минус-конце считается равной нулю,а в связи с активными процессами нуклеации и элонгации реакции диссциации также не были учтены. Все полученные в результате эксперимента данные представлены в таблице 2 . ,извлеченный из кишечной палочки BL21 особым образом. Для изучения динамики процесса полимеризации использовалась силективная флуоресценция пирена.Таким образом в эксперименте были задействованы реакции элонгации,нуклеации ,блокировки и разблокировки.По ходу эксперимента были получены скорости всех реакции. Предполагалось ,что элогация происходит преимущественно на плюс конце филаменты ,поэтому элонгация на минус-конце считается равной нулю,а в связи с активными процессами нуклеации и элонгации реакции диссциации также не были учтены. Все полученные в результате эксперимента данные представлены в таблице 2 .
Таблица 2- Скорости реакции, полученные в результате эксперимента [17]
| Обозначение
|
Описание
|
Значение
|
|
|
скорость спонтанной нуклеации
|

|
|
|
скорость элонгации плюс-конца
|
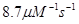
|
|
|
скорость элонгации минус-конца
|
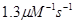
|
|
|
скорость блокировки плюс-конца
|

|
|
|
скорость разблокировки плюс-конца
|

|
|
|
скорость нуклеации с участием блокирующего протеина
|

|
|
|
концентрация g-актина
|

|
Данные из таблицы были использованы при имитационном моделировании и полученная концентрация f-актина была сравнена с результатами эксперимента (рисунок 5).

Рисунок 5-Сравнение результатов имитационного моделирования с результатами, полученными в результате эксперимента[29]
Таким образом, хорошая согласованность результатов позволяет сделать вывод о том, разработанная имитационная модель адекватна и соответствует реальным процессам полимеризации актина, происходящих в живых клетках и растворах.
Заключение
В работе построена имитационная модель процесса полимеризации актина, проведена верификация разработанной модели на аналитической модели и на экспериментальных данных. По результатам работы можно сделать следующие выводы:
· Блокирующий протеин не изменяет стационарного состояния f-актина, а ускоряет его достижение. Распределение филаментов по длинам меняется в сторону уменьшения при введении данного протеина в систему.
· Введение реакции фрагментации также не изменяет концентрации f-актина в стационарном состоянии, а лишь уменьшает среднюю длину филаментов в системе. Благодаря данной реакции филаментов становится больше в системе.
· Введение реакции старения актина в филаментах уменьшает значение стационарной концетрации f-актина и среднюю длину филаментов в системе, поскольку концы филаментов АДФ состоянии диссоциируют более интенсивно.
· Формин даже в небольших концентрациях значительно ускоряет наступление стационарного состояния. Значение концентрации f-актина в стационарном состоянии остается неизменным, за исключением случаев, когда концентрация формина становится на один порядок меньше начальной концентрации g-актина в АТФ форме. В таком случае наблюдается резкое снижение концентрации f-актина и момент достижения максимальной концентрации f-актина сдвигается по временной оси влево. При очень малых начальных концентрациях формина (порядка ~10^-4) наблюдается рост длин филаментов с увеличением концентрации формина. Однако, при дальнейшем увеличении концентрации наблюдается уменьшение их средней длинны.
· Наибольшим быстродействием обладает оптимизированный прямой метод Гиллеспи, поскольку при моделировании процесса полимеризации актина вероятности наступления различных реакций отличаются на несколько порядков, что позволяет использовать данный факт при оптимизации.
Построенную модель планируется в дальнейшем усовершенствовать и использовать для анализа экспериментальных данных, полученных с помощью FRAP- и флуоресцентной спектроскопии .
Список литературы к реферату
Список использованных источников
1. Soo, F.S. and J.A. Theriot, Large-scale quantitative analysis of sources of variation in the ac
tin polymerization-based movement of Listeria monocytogenes. Biophys. J., 2005. 89(1): p. 703-23.
2. Alberts, J.B. and G.M. Odell, In silico reconstitution of Listeria propulsion exhibits nano-saltation. PLoS Biol., 2004. 2(12): p. e412.
3. Carlsson, A.E., M.A. Wear, and J.A. Cooper, End versus Side Branching by Arp2/3 Complex. Biophys. J., 2004. 86: p. 1074-81.
4. Yatskou, M.M., et al., Nonisotropic excitation energy transport in organized molecular systems:
Monte Carlo simulation-based analysis of time-resolved fluorescence. J. Phys. Chem. A, 2001. 105(41): p. 9498-9508.
5. Nazarov, P.V., et al., FRET study of membrane proteins: simulation-based fitting for analysis of membrane protein embedment and association. Biophys. J., 2006. 91(2): p. 454-466.
6. Nazarov, P.V., et al., Artificial neural network modification of simulation-based fitting: application to a protein-lipid system. J. Chem. Inf. Comput. Sci., 2004. 44(2): p. 568-74.
7. Romero, S., et al., Formin is a processive motor that requires profilin to accelerate actin assembly and associated ATP hydrolysis. Cell, 2004. 119(3): p. 419-29.
8. Halavatyi ,A, et al.,,. An integrative simulation model linking major biochemical reactions of actin-polymerization to structural properties of actin filaments.2008.p 1-6.
9. Gillespie, D.T., Approximate accelerated stochastic simulation of chemically reacting systems. J. Phys. Chem., 2001. 115(4): p. 1716-33.
10. Клячко, Н.Л. Биологическая подвижность и полимеризация актина,2000.с.2-4
11. Kovar, D.R., Molecular details of formin-mediated actin assembly. Curr. Opin. Cell Biol., 2006. 18(1): p. 11-7.
12. Gillespie, D.T., Exact stochastic simulation of coupled chemical reactions. J. Phys. Chem., 1977. 81(25): p. 2340-2361.
13. Gillespie, D.T., A general method for numerically simulating the stochastic time evolution of coupled chemical reactions. J. Comput. Phys., 1976. 22: p. 403-434.
14. Cao, Y., H. Li, and L. Petzold, Efficient formulation of the stochastic simulation algorithm for chemically reacting systems. J Chem Phys, 2004. 121(9): p. 4059-67.
15. Cao, Y., et al., The numerical stability of leaping methods for stochastic simulation of chemically reacting systems. J Chem Phys, 2004. 121(24): p. 12169-78.
16. Gibson, M.A. and J. Bruck, Efficient Exact Stochastic Simulation of Chemical Systems with Many Species and Many Channels. J. Phys. Chem., 2000. 104: p. 1876-89.
17. Pollard, T.D., Rate constants for the reactions of ATP- and ADP-actin with the ends of actin filaments. J Cell Biol, 1986. 103: p. 2747-54.
18. Carlsson, A.E., Actin Polymerization Overshoots and Hydrolysis as Assayed by Pyrene Fluorescence, 2008 p. 140-70.
19. Carlsson, A.E., Structure of Autocatalytically Branched Actin Solutions. PhysRevLett, 2004. 92(23): p. 238102-1-4.
Список публикаций соискателя
1 -А.Чухутина О.В. Имитационное моделирование биохимических реакций//Сборник работ 64-й научной конференции студентов и аспирантов Белгосуниверситета,2007.
2 -А.Чухутина О.В. Имитационное моделирование биохимических реакций//Материалы V республиканской научной конференции молодых ученых и студентов “Современные проблемы математики и вычислительной техники”,Брест,Беларусь,2007.
3 -А.Чухутина О.В. Построение и анализ имитационных моделей полимеризации актина//Сборник работ 65-й научной конференции студентов и аспирантов Белгосуниверситета,2008.
Предметный указатель к реферату
ARP2/3–комплекс, 9
АДФ/кофилин, 9
Актин, 7
Aлгоритм «первой реакции», 14
Глобулярная форма актина, 7
Димер, 8
Имитационное моделирование, 10
Кэп-белки, 9
Мономеры, 7
Прямой метод, 12
Фибриллярная форма актина, 7
Филамента, 7
Формин, 10
Интернет ресурсы в предметной области исследования
http://www.cell.com/biophysj/ - данный источник представляет собою наиболее полное собрание научных статей в области биологии, биохимического моделирования. Удобный интерфейс облегчает поиск нужного материала.
http://elsevier.com/ -Журнал о теоретической биологии. Ведущий форум, освещающий теоретические работы в области биологического прогресса. Освещает очень широкий круг интересов исследователей в разных областях биологии.
www.intelligen.com –сайт компании INTELLIGEN,которая занимается разработкой программ имитационного моделирования. На сайте есть возможность скачать данные программные продукты. Есть документация по их использованию.
http://www.eurosim.info/ - FEDERATION OF EUROPEAN SIMULATION SOCIETIES–Федерация Европейских ассоциации моделирования, которая обеспечивает европейски форум для региональных и национальных сообществ, занимающихся различными видами моделирования.
http://como.cheng.cam.ac.uk/ -вебсайт научной группы Кембриджского университета, занимающейся численным моделированием.
Действующий личный сайт в
WWW
http://chukhutsina.narod.ru/
Граф научных интересов
аспиранта Чухутиной О.В., факультет радиофизики и электроники
Специальность физическая электроника
| Смежные специальности
| 01.01.05 –теория вероятностей и математическая статистика
|
| 1. Стохастический анализ и стохастические дифференциальные уравнения.
2. Случайные процессы специального вида, включая процессы массового обслуживания.
3. Статистика случайных процессов, полей и временных рядов.
4. Вероятностно-статистическое моделирование.
|
|
Основная специальность
| 05.13.18 – математическое моделирование, численные методы и комплексы программ
|
| 1. Исследование и разработка методов и принципов построения математических моделей.
2. Развитие, обоснование и применение математических моделей для решения актуальных научных задач естествознания (в биохимии.)
|
|
Сопутствующие специальности
| 03.00.04 – биохимия
|
| 1. Взаимосвязь химического строения, структуры и функций белков. Пути денатурации белковой глобулы. Современные биохимические методы исследования белков.
2. Биохимия регуляторных процессов.
3. Разработка биохимических методов исследования живых систем и получения веществ с заданными свойствами.
|
| 02.00.03 – органическая химия
|
| 1. Изучение строения и свойств органических соединений с использованием химических, физико-химических и физических методов исследования и теоретических расчетов.
2. Изучение реакционной способности и механизмов реакций органических соединений.
3. Препаративная органическая химия, разработка методов органического синтеза, его теории и практики, молекулярный дизайн, комбинаторная химия.
4. Прикладная органическая химия.
5. Промышленная органическая химия и научные основы технологии органического синтеза.составом и структурой.
|
|
Презентация магистерской диссертации
Презентация к данной работе выполнена в Microsoft Office PowerPoint 2003 и может быть скачена с личного сайта соискателя (http://chukhutsina.narod.ru/presentIT.ppt ).
Тестовые
вопросы
<question type="close" id="59">
<text>(Чухутина Ольга) Что из перечисленного является отличием XHTML от HTML:</text>
<answers type="request">
<answer id="1" right="1">в XHTML применяется синтаксис XML.</answer>
<answer id="2" right="1">в XHTML все элементы обязаны иметь закрывающий тег.</answer>
<answer id="3" right="1">Документы XHTML должны использовать нижний регистр для всех имен элементов и атрибутов HTML.</answer>
<answer id="4" right="0">HTML представляет собой словарь XML.</answer>
</answers>
</question>
<question type="close" id="559">
<text>(Чухутина Ольга) Что из перечисленного не является характерной чертой аналитических моделей:</text>
<answers type="request">
<answer id="1" right="0">возможность получать информацию только о средних значениях рассматриваемых величин.</answer>
<answer id="2" right="1">возможность рассматривать присутствие различных шумов, связанных с дискретностью системы.</answer></answer>
<answer id="3" right="0">замена реализаций случайных величин их средними значениями.</answer>
</answers>
</question>
Список литературы к выпускной работе
1. Soo, F.S. and J.A. Theriot, Large-scale quantitative analysis of sources of variation in the actin polymerization-based movement of Listeria monocytogenes. Biophys. J., 2005. 89(1): p. 703-23.
2. Alberts, J.B. and G.M. Odell, In silico reconstitution of Listeria propulsion exhibits nano-saltation. PLoS Biol., 2004. 2(12): p. e412.
3. Carlsson, A.E., M.A. Wear, and J.A. Cooper, End versus Side Branching by Arp2/3 Complex. Biophys. J., 2004. 86: p. 1074-81.
4. Yatskou, M.M., et al., Nonisotropic excitation energy transport in organized molecular systems: Monte Carlo simulation-based analysis of time-resolved fluorescence. J. Phys. Chem. A, 2001. 105(41): p. 9498-9508.
5. Nazarov, P.V., et al., FRET study of membrane proteins: simulation-based fitting for analysis of membrane protein embedment and association. Biophys. J., 2006. 91(2): p. 454-466.
6. Nazarov, P.V., et al., Artificial neural network modification of simulation-based fitting: application to a protein-lipid system. J. Chem. Inf. Comput. Sci., 2004. 44(2): p. 568-74.
7. Romero, S., et al., Formin is a processive motor that requires profilin to accelerate actin assembly and associated ATP hydrolysis. Cell, 2004. 119(3): p. 419-29.
8. Halavatyi ,A, et al.,,. An integrative simulation model linking major biochemical reactions of actin-polymerization to structural properties of actin filaments.2008.p 1-6.
9. Gillespie, D.T., Approximate accelerated stochastic simulation of chemically reacting systems. J. Phys. Chem., 2001. 115(4): p. 1716-33.
10. Клячко, Н.Л. Биологическая подвижность и полимеризация актина,2000.с.2-4
11. Kovar, D.R., Molecular details of formin-mediated actin assembly. Curr. Opin. Cell Biol., 2006. 18(1): p. 11-7.
12. Gillespie, D.T., Exact stochastic simulation of coupled chemical reactions. J. Phys. Chem., 1977. 81(25): p. 2340-2361.
13. Gillespie, D.T., A general method for numerically simulating the stochastic time evolution of coupled chemical reactions. J. Comput. Phys., 1976. 22: p. 403-434.
14. Cao, Y., H. Li, and L. Petzold, Efficient formulation of the stochastic simulation algorithm for chemically reacting systems. J Chem Phys, 2004. 121(9): p. 4059-67.
15. Cao, Y., et al., The numerical stability of leaping methods for stochastic simulation of chemically reacting systems. J Chem Phys, 2004. 121(24): p. 12169-78.
16. Gibson, M.A. and J. Bruck, Efficient Exact Stochastic Simulation of Chemical Systems with Many Species and Many Channels. J. Phys. Chem., 2000. 104: p. 1876-89.
17. Pollard, T.D., Rate constants for the reactions of ATP- and ADP-actin with the ends of actin filaments. J Cell Biol, 1986. 103: p. 2747-54.
18. Carlsson, A.E., Actin Polymerization Overshoots and Hydrolysis as Assayed by Pyrene Fluorescence, 2008 p. 140-70.
19. Carlsson, A.E., Structure of Autocatalytically Branched Actin Solutions. PhysRevLett, 2004. 92(23): p. 238102-1-4.
Приложения
|